1. Introduction
Irreversible antagonists play a pivotal role in the investigation of receptors, being useful for differentiating receptor types and subtypes and being essential in the assessment of receptor reserve. Receptor reserve is an integrative measure of the response-inducing ability of the interaction between an agonist and receptor system consisting of a receptor and postreceptorial signalling [
1,
2]. This term is coined to elucidate the phenomenon, that is, in some cases, the stimulation of a submaximal fraction of receptors can apparently elicit the maximal effect. Accordingly, detection and quantification of receptor reserve is based on the comparison of
E/
c curves generated in the absence and presence of an irreversible receptor antagonist. As receptor reserve depends on the tissue, agonist and effect, receptor reserve values should be determined for each tissue, agonist and effect being of practical importance [
1,
2].
The A
1 adenosine receptor (A
1 receptor) is a member of the ancient and ubiquitous adenosine receptor family that exerts complex regulatory functions in almost all tissues [
3,
4,
5]. In the heart, the A
1 receptor mediates strong negative tropic effects including negative inotropy on the ventricle and, even more, on the atrium [
6], as a component of the A
1 adenosinergic cardioprotection against ischemic/hypoxic stress [
7,
8,
9]. Accordingly, in earlier studies, we aimed to assess the guinea pig atrial A
1 receptor reserve for the direct negative inotropic effect of several A
1 adenosine receptor agonists (the term direct means that experiments were conducted on atria lacking prior stimulation of contractility). For this purpose, concentration-response (
E/
c) curves were constructed in the absence and presence of 8-cyclopentyl-
N3-[3-(4-(fluorosulfonyl)benzoyloxy)propyl]-
N1-propylxanthine (FSCPX). After its first appearance in a report from 1994 [
10], FSCPX was tested and then utilized as a selective and irreversible A
1 receptor antagonist in numerous investigations [
11,
12,
13,
14,
15,
16,
17,
18,
19].
In the case of stable, synthetic full agonists (such as
N6-cyclopentyladenosine: CPA), the A
1 receptor reserve could be successfully determined with the operational model of agonism and Furchgott’s method, two procedures providing a quantitative result. The A
1 receptor reserve proved to be extremely large for the direct negative inotropic effect [
17]. This finding could be well illustrated by the fact that FSCPX was unable to significantly decrease the maximal effects of these agonists, although, based on results of Furchgott’s method, about 80–90% of the A
1 receptors were inactivated by FSCPX. Instead, FSCPX only shifted the
E/
c curves of the stable agonists to the right, as if it were a competitive antagonist [
17]. However, our attempt failed to assess the A
1 receptor reserve for the direct negative inotropic effect of adenosine, the physiological agonist, when naïve and FSCPX-pre-treated adenosine
E/
c curves were evaluated with the operational model and Furchgott’s method. This failure was probably due to the too short half-life of adenosine in our ex vivo experimental system that impeded adenosine
E/
c curves to get saturated [
17].
To address this challenge, an alternative receptor reserve-estimating method had to be developed for adenosine [
18]. The essence of this method is the use of
S-(2-hydroxy-5-nitrobenzyl)-6-thioinosine (NBTI), a selective inhibitor of the equilibrative and NBTI-sensitive nucleoside transporter (ENT1; SLC29A1) [
20]. As the physiological adenosine transport is directed into the cells (e.g., cardiomyocytes) [
21,
22], NBTI prevents adenosine, administered for the
E/
c curve, from the intracellular degradation and reutilization, allowing enough time for the exogenous adenosine to exert its effect. However, NBTI also prevents endogenous adenosine from the intracellular elimination that leads to a rise of the interstitial adenosine concentration. Importantly, this latter phenomenon occurs already before the generation of the adenosine
E/
c curve (and it exists throughout the experiment, till NBTI is present). As the surplus endogenous adenosine accumulated by NBTI consumes in part the response capacity of A
1 receptors (and their postreceptorial signalling) before constructing the adenosine
E/
c curve, the response to adenosine (detected by the given adenosine
E/
c curve) will show an apparent diminution. Thus, effect values of adenosine
E/
c curves generated in the presence of NBTI should be corrected for the distortion caused by the increased endogenous adenosine level in the interstitium. For this correction, we apply a procedure based on the receptorial responsiveness method (RRM) [
23,
24]. Finally, the corrected FSCPX-naïve and FSCPX-pre-treated adenosine
E/
c curves (generated in the presence of NBTI) are compared. Information on the receptor reserve in question can be obtained from the distance of the final (saturated) parts of the corrected curves: small distance means great receptor reserve, while large distance indicates a small one. Although our receptor reserve-estimating method provides only qualitative results, the A
1 receptor reserve for the direct negative inotropic effect of adenosine could be assessed and has been found similarly large as that for the synthetic agonists, in both eu- [
18] and hyperthyroid states [
19].
However, during the process of determining receptor reserve with our qualitative method, we encountered an astonishing phenomenon that, in the presence of NBTI, FSCPX pre-treatment apparently increased the response to adenosine (when the FSCPX-NBTI co-treated adenosine
E/
c curve was compared to the solely NBTI-treated one) [
18,
19]. Starting from this pharmacological paradox, a recent
in silico study from our laboratory has raised the possibility that FSCPX pre-treatment may modify the effect of NBTI through a mechanism other than blocking the A
1 receptor [
25]. To the best of our knowledge, no other paper has reported any effect of FSCPX other than permanent A
1 receptor inactivation (even as a hypothesis).
Our assumption about an interference between effects of FSCPX and NBTI implies that our receptor reserve-estimating method should be revisited. In the present study, therefore, our primary goal was to justify or refute our assumption about this interference. Furthermore, our goal was to improve our receptor reserve-estimating method to eliminate the distorting effect resulted from this unexpected interference. Finally, we aimed to compare results obtained from the original and improved forms of our method. For these purposes, we have included a new protocol to the old ones used in previous studies (i.e., [
18,
19]) (see:
Table 1). This amendment has offered new information on the mechanism of action of FSCPX, furthermore it has made our receptor reserve-estimating method free of the disturbing effect stemming from the interference between actions of FSCPX and NBTI.
3. Discussion
The main experimental finding of the present study is that a pre-treatment with FSCPX, a chemical widely considered to be a selective and irreversible A
1 receptor antagonist, selectively influences the different effect components of NBTI, a selective nucleoside transport inhibitor, that are apparent on
E/
c curves of adenosine and CPA, two A
1 receptor agonists, in the isolated guinea pig atrium. While FSCPX pre-treatment considerably counteracts the depressive (
Emax-decreasing) effect of NBTI on
E/
c curves of both adenosine and CPA, it does not significantly affect the extensive action of NBTI on
EC50 of the adenosine
E/
c curve. Thus, our recent proposal, that is, FSCPX blunts the effect of NBTI [
25], should be refined in light of the new results. It seems that FSCPX pre-treatment inhibits the effects of NBTI that are mediated by elevating the interstitial level of endogenous but not exogenous adenosine. Consequently, the target of FSCPX (other than the A
1 receptor) cannot be the ENT1 transporter or any other molecules that participate in the mediation of the effect of NBTI on the level of exogenous adenosine. Conversely, the target in question may be a (or some) molecule(s) that is (are) associated exclusively with the effect of NBTI exerted on the level of endogenous adenosine, for example, enzymes contributing to the interstitial adenosine formation.
In a recent investigation dealing with the
in silico modelling of our receptor reserve-estimating method [
25], we have found that
E/
c curves resulted from studies using this method [
18,
19] could not be simulated without supposing an interference between effects of FSCPX and NBTI. Based on this finding, we hypothesized that FSCPX might modify ENT1 in a way that it allows ENT1 to transport adenosine but impedes NBTI to inhibit this transport [
25]. In the present study, in order to explore the interference between effects of FSCPX and NBTI, we have amended the protocols of our receptor reserve-estimating method to enable detecting the influence of FSCPX and NBTI co-treatment on the CPA
E/
c curve.
The effects of FSCPX and NBTI on the
E/
c curves of adenosine receptor agonists can be divided into three components: Effects mediated by influencing the interstitial level of endogenous adenosine (that were earlier attributed exclusively to NBTI [
26,
27]), effects mediated
via affecting the interstitial level of exogenous adenosine (also ascribed to NBTI [
28]) and effects resulted from the permanent inactivation of a fraction of the A
1 receptor population (attributed to FSCPX [
11]).
In terms of the shape of our
E/
c curves, differentiation of effects of “endogenous” and “exogenous” adenosine originates in the features of our experimental model. The “effects mediated by influencing the interstitial level of endogenous adenosine” refers to consequences of a rise in the interstitial adenosine concentration developed
before the construction of the
E/
c curve of an adenosine receptor agonist. This occurs when NBTI, administered to the atria before the generation of the
E/
c curve, reduces the elimination of adenosine produced interstitially in the course of the tissue adenosine turnover. In contrast, the “effects mediated via affecting the interstitial level of exogenous adenosine” means outcomes of an attenuated decrease (so a relative increase) in the interstitial adenosine concentration
during the construction of an adenosine
E/
c curve. This is the case when NBTI decreases the elimination of adenosine administered as
E/
c curve doses. The point is that, if receptors (with their signalling) respond to a stimulus before the generation of an
E/
c curve, this response will be reflected in the subsequent
E/
c curve as a deficit in the responsiveness of the receptors (and their signalling). This is the motif of RRM [
23,
24] and that is why RRM can be used to correct
E/
c curves distorted by a response evoked before the construction of the
E/
c curve [
18]. As a result, increase in interstitial concentrations of endogenous and exogenous adenosine (upon our experimental conditions) exert the opposite effect on the main parameters (
Emax,
EC50) of an adenosine
E/
c curve (
Table 3).
In the case of an NBTI-treated CPA
E/
c curve, the situation is relatively simple: The surplus endogenous adenosine produced by NBTI will consumes the response capacity of the cell-surface A
1 receptors (and their postreceptorial pathways) before the construction of the
E/
c curve, therefore NBTI decreases
Emax and increases
EC50 of the CPA
E/
c curve [
26]. In the case of an NBTI-treated adenosine
E/
c curve, the situation is more complex: Consequences of the elevated endogenous adenosine level will combine with effects stemming from the preservation of adenosine administered for the
E/
c curve, which latter effects tend to increase
Emax and to decrease
EC50 of the adenosine
E/
c curve. As a resultant, NBTI reduces both
Emax and
EC50 of the adenosine
E/
c curve [
28] (
Table 3).
Before the emergence of our recent results [
25], action of FSCPX appeared the simplest: similar increase in
EC50 of the CPA and adenosine
E/
c curves, consistent with the irreversible A
1 receptor antagonist property of FSCPX and the large A
1 receptor reserve for the direct negative inotropy in the guinea pig atrium [
17,
18,
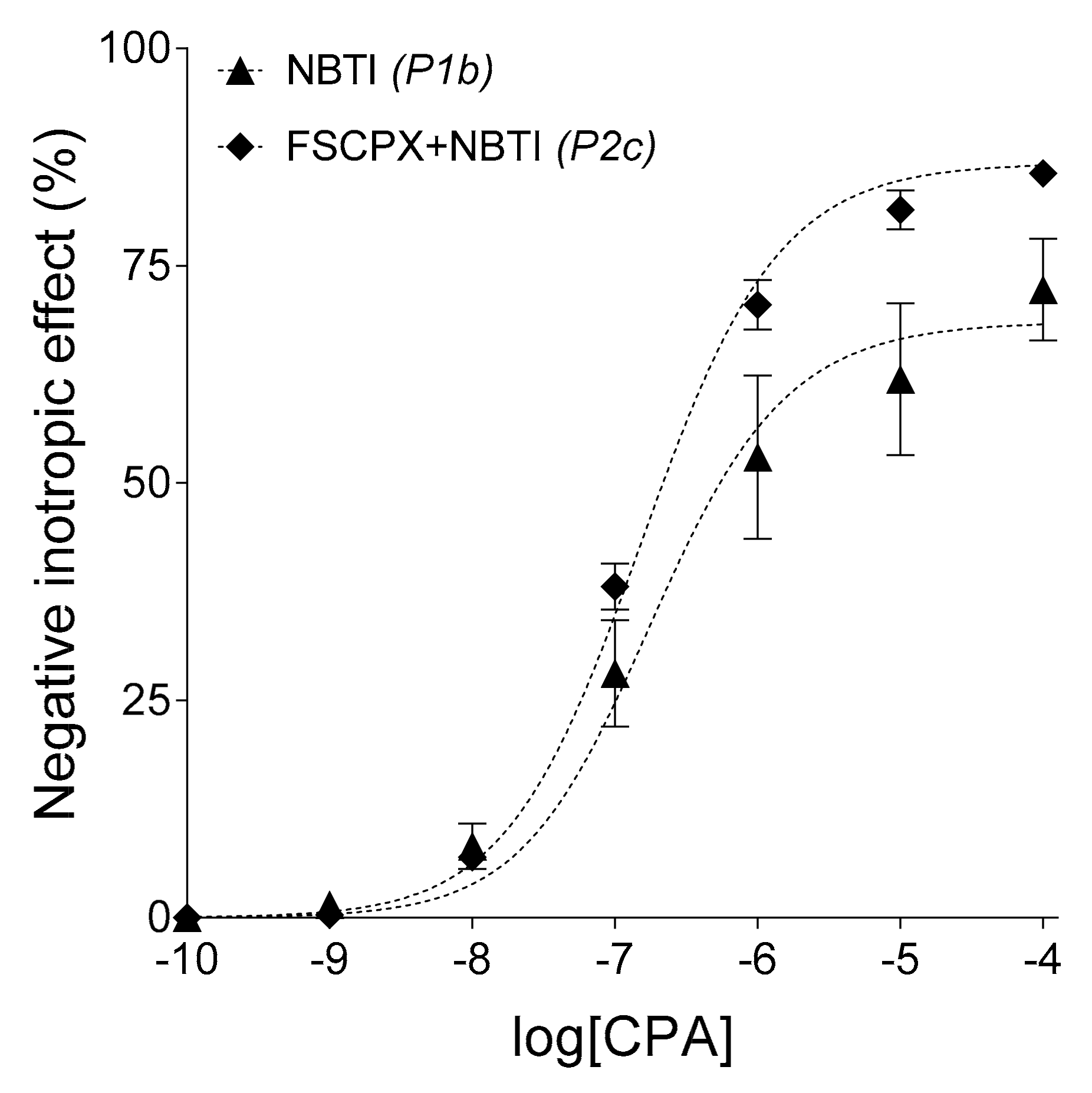
19]. In the present study, however, when we applied an FSCPX pre-treatment before the construction of the CPA
E/
c curve in the presence of NBTI, the resulting effect was an almost complete cancelling of NBTI’s effects with the preservation of the action of FSCPX (
Figure 2A,
Table 2). This curve constellation cannot be explained with the irreversible A
1 receptor antagonist property of FSCPX. If FSCPX were a simple irreversible antagonist, it would have only shifted the NBTI-treated CPA
E/
c curve to the right (increasing its
EC50). In turn, when an FSCPX-NBTI co-treatment was applied in the case of an adenosine
E/
c curve, the
Emax-decreasing effect of NBTI was only cancelled, while the
EC50-decreasing effect of NBTI remained practically intact (
Figure 2B,
Table 2). This curve constellation further contradicts the prevailing concept being FSCPX only an irreversible A
1 receptor antagonist, moreover it also shows that the additional effect of FSCPX cannot be an equal inhibition of all effects of NBTI.
Based on
in silico results, recently we hypothesized that FSCPX might impede the inhibitory action of NBTI on ENT1 [
25]. It should be reminded that all effects of NBTI stem from the inhibition of ENT1 [
20], by which NBTI reduces the physiologically inward transmembranous adenosine transport and thus increases the interstitial concentration of adenosine of any origin [
21,
22]. The above-mentioned hypothesis would agree with the present results obtained from CPA
E/
c curves (
Figure 2A,
Table 2) but it contrasts with the current results yielded from adenosine
E/
c curves (
Figure 2B,
Table 2). Such a great increase in
Emax together with the preservation of
EC50, seen in the case of the FSCPX-NBTI co-treated adenosine
E/
c curve as compared to the only NBTI-treated one, can hardly be imagined if we suppose the inhibition of all effects of NBTI (and inhibition of ENT1 would decrease all NBTI-evoked effects). Interestingly, earlier studies [
18,
19] yielded results, for which this previous hypothesis appeared to work. However, depression of the solely NBTI-treated adenosine
E/
c curve in those studies was considerably smaller than that in the present investigation, although relative positions of the control, FSCPX-pre-treated, NBTI-treated and FSCPX-NBTI co-treated adenosine
E/
c curves were the same in all above-mentioned studies.
Taking all together, the target of FSCPX, to exert its effect other than A
1 receptor inactivation, should be a molecule that is related to the effect of NBTI that is mediated by increasing the interstitial level of endogenous adenosine. Enzymes contributing (exclusively or at least predominantly) to the interstitial adenosine formation are possible candidates. It can be speculated that FSCPX, which ruins the binding site for adenosine in the A
1 receptor, can do the same with the binding site of one (or some) enzyme(s) participating in the interstitial adenosine production. If it were so, FSCPX pre-treatment would decrease the interstitial adenosine level. As the resting interstitial adenosine concentration elicits undetectably small inotropic effect in the guinea pig atrium [
29], the supposed diminution of it by FSCPX would be also undetectable. However, under conditions with reduced interstitial adenosine producing capacity resulted from an FSCPX pre-treatment, NBTI could elevate the interstitial level of endogenous adenosine to a smaller extent than otherwise. This reduced increase in the interstitial adenosine level (before generating an
E/
c curve) would cause easily detectable alterations in
E/
c curves of both adenosine and CPA: Tendency to a smaller decrease in
Emax and to a smaller increase in
EC50 (
Table 3). It is also important that, supposing the above-mentioned additional mechanism of action for FSCPX, the protective effect of NBTI on the exogenous adenosine would freely prevail. Regarding the shape of our CPA and adenosine
E/
c curves (
Figure 2A,B,
Table 2), this is exactly the case in the present investigation. The only concern might be the fact that, after FSCPX pre-treatment, NBTI did not cause a significantly greater decrease in
EC50 of the adenosine
E/
c curve as compared to the control state than NBTI alone. However, this phenomenon can be ascribed to the irreversible antagonist property of FSCPX. When comparing the FSCPX-NBTI co-treated adenosine
E/
c curve to the solely FSCPX-pre-treated one, the sinistral displacement (decrease in
EC50) is much larger than it is if comparing the NBTI-treated adenosine
E/
c curve to the control one (
Figure 2A,B,
Table 2). Thus, we have improved our hypothesis that is (from the appearance of this writing) as follows: FSCPX pre-treatment modifies the action of NBTI on
E/
c curves of adenosine receptor agonists via limiting the interstitial adenosine producing capacity, perhaps by inhibiting one (or some) interstitial adenosine forming enzyme(s) (
Table 3). (Theoretically, an alternative mechanism for FSCPX to decrease the interstitial adenosine level could be the enhancement of elimination of interstitial adenosine. However, this mechanism is unlikely because it would lower the concentration of both exogenous and endogenous adenosine, a possibility that is not supported by the present results)
A phenomenon that is also worthwhile to address is the slight increase of the response to CPA at low concentrations in the case of the FSCPX-NBTI co-treated CPA
E/
c curve, as compared to its adequate counterpart, the FSCPX-pre-treated CPA
E/
c curve (
Figure 2A). This may be explained by supposing that CPA, in agreement with our previous observation [
23], undergoes a minor elimination in the atrial tissue. As in the case of the NBTI-treated and FSCPX-NBTI co-treated adenosine
E/
c curves in comparison with the control and FSCPX-pre-treated ones, respectively (
Figure 2B), the increased response to the given adenosine receptor agonist shows the protective effect of NBTI that manifests predominantly at low concentrations (
Figure 2A,B). Thus, for further studies in this topic, use of
N6-cyclohexyladenosine (CHA), a selective A
1 receptor full agonist showing higher resistance against adenosine-handling enzymes [
30], should be considered.
Further observation of the present study is that the scatter of
E/
c curve data is the biggest for the solely NBTI-treated curves. Interestingly, this big scatter caused by NBTI has been largely prevented by the FSCPX-pre-treatment (
Figure 2A,B,
Figure 3). It may be speculated that the cause of the exceptionally big scatter in the presence of NBTI is due to the considerable individual variability of enzyme activities contributing to the interstitial adenosine production that, if inhibited by FSCPX, makes the response to adenosine receptor agonists more homogenous.
A godsend for this investigation was that NBTI exerted very strong
Emax-decreasing (for both CPA and adenosine
E/
c curves:
Figure 2A,B) and
EC50-increasing (for the CPA
E/
c curve:
Figure 2A) effects during the experiments presented herein, in comparison with earlier studies [
18,
19,
26,
27]. In our interpretation, these effects of NBTI on the
E/
c curves of adenosine receptor agonists can be ascribed to the action of NBTI on the interstitial level of endogenous adenosine (
Table 3). Thus, it is reasonable to assume that, in our present experiments, atria possessed exceptionally high interstitial adenosine forming capacity, allowing both NBTI and FSCPX to exert especially strong effects on the interstitial level of endogenous adenosine. Consistently, we have found the highest
cx value ever in our praxis that characterizes well the efficiency of NBTI in the present study (
Table 4). For the reason mentioned in the previous paragraph, the efficiency of FSCPX (to elicit its additional effect) could only manifest in its interaction with NBTI: The FSCPX pre-treatment produced a considerably large diminution in
cx (from 100.2 nM to 6.73 nM). These strong effects observed concerning FSCPX and NBTI enabled us to improve our previous assumption on the mechanism of action of FSCPX other than A
1 receptor inactivation.
The
cx values provided the opportunity to create hypothetical adenosine
E/
c curves free of the disturbing effect of the interstitial accumulation of endogenous adenosine in response to NBTI (
Figure 4). These hypothetical adenosine
E/
c curves allow us to compare our improved receptor reserve-estimating method with the original one. The FSCPX-NBTI co-treated adenosine
E/
c curve, corrected with the original method, ran unequivocally above its counterpart corrected with the improved method (
Figure 4). This phenomenon corroborates the assumption made previously [
25] that the original method, used for two earlier studies [
18,
19], overestimates and thereby overcorrects the distortion caused by NBTI, if previous FSCPX pre-treatment has occurred. Since this inaccuracy affects responses belonging to the high adenosine concentrations only to a small extent and since these responses form the basis to estimate receptor reserve (via comparing them with the corresponding response values of the corrected NBTI-treated adenosine
E/
c curve [
18,
19]), the original method over measures the receptor reserve in question only to a slight extent (
Figure 4). Of course, application of the improved method is recommended in the future.
4. Materials and Methods
4.1. Materials
The following chemicals were used: Adenosine, N6-cyclopentyladenosine (CPA), 8-cyclopentyl-N3-[3-(4-(fluorosulfonyl)benzoyloxy)propyl]-N1-propylxanthine (FSCPX) and S-(2-hydroxy-5-nitrobenzyl)-6-thioinosine (NBTI), purchased from Sigma (St. Louis, MO, USA).
Adenosine was dissolved in 36 °C modified Krebs-Henseleit buffer (Krebs solution) containing (in mM): NaCl: 118, KCl: 4.7, CaCl2: 2.5, NaH2PO4: 1, MgCl2: 1.2, NaHCO3: 24.9, glucose: 11.5, ascorbic acid: 0.1, dissolved in redistilled water. CPA was dissolved in ethanol:water (1:4) solution (v/v). Dimethyl-sulfoxide (DMSO) was used as a solvent for FSCPX and NBTI. All stock solutions were adjusted to a concentration of 10 mM, except for the adenosine stock solution used to achieve 3 mM concentration in the bathing medium (it was 20 mM and was prepared always freshly before each use). Adenosine and CPA stock solutions were diluted with Krebs solution.
4.2. Animals, Preparations and Protocols
All experiments were carried out between 6–29 March 2018. The animal use protocols were approved by the Committee of Animal Research, University of Debrecen, Hungary (25/2013/DE MÁB). Male Hartley guinea pigs, weighing 600–800 g, were guillotined and then left atria were quickly removed and mounted at 10 mN resting tension in 10 mL vertical organ chambers (Experimetria TSZ-04; Experimetria Kft, Budapest, Hungary) containing Krebs solution, oxygenated with 95% O2 and 5% CO2 (36 °C; pH = 7.4). Atria were paced by platinum electrodes (3 Hz, 1 ms, twice the threshold voltage) by means of a programmable stimulator (Experimetria ST-02; Experimetria Kft, Budapest, Hungary) and power amplifier (Experimetria PST-02; Experimetria Kft, Budapest, Hungary). The contractile force was characterized by the amplitude of the isometric twitches, which were measured by a transducer (Experimetria SD-01; Experimetria Kft, Budapest, Hungary) and strain gauge (Experimetria SG-01D; Experimetria Kft, Budapest, Hungary) and recorded by a polygraph (Medicor R-61 6CH Recorder; Medicor Művek, Budapest, Hungary).
The atria were divided into seven groups (P1a, P1b, P2a, P2b, P2c, P3 and P4;
n = 6–10) according to the seven experimental protocols of the present study. The protocols were the same as those used for our previous investigations [
18,
19], except for a rearrangement of protocol numbering and introduction of a new protocol, P2c that was carried out first in the present study (
Table 1).
4.3. Empirical Characterization of the E/c Curves
All
E/
c curves were fitted to the Hill equation that is a simple and reliable empirical model of receptor function [
31]:
where:
E: The effect (that was defined as a percentage decrease in the initial contractile force of atria);
c: The concentration of the agonist (administered for the given
E/
c curve);
Emax: The maximal effect;
EC50: The agonist concentration producing half-maximal effect;
n: The Hill coefficient (slope factor).
The Hill equation was fitted to both individual and averaged E/c curve data.
4.4. Quantification of the Distortion Produced by NBTI in the CPA E/c Curves
NBTI, via selective inhibition of the inward adenosine transport, increases the interstitial concentration of endogenous adenosine in the atrium under well-oxygenated conditions [
26,
27]. Thus, our
E/
c curves constructed in the presence of NBTI were distorted by a surplus interstitial adenosine concentration that was developed already before the construction of the
E/
c curve. To quantify this extra adenosine concentration, RRM was performed. Specifically, the averaged CPA
E/
c curves of P1b (only NBTI-treated) and P2c (FSCPX-NBTI co-treated) groups were fitted to Equation (2), which contained the empirical parameters of the averaged CPA
E/
c curve of the P1a (control) or P2b (solely FSCPX-pre-treated) group, respectively:
where:
E’: The distorted effect (calculated conventionally);
Emax,
EC50,
n: empirical CPA
E/
c curve parameters (provided by Equation (1)) that define the basic (i.e., NBTI-free) conditions;
c: The concentration of CPA (administered for the
E/
c curve);
cx: The variable parameter of Equation (2) that indicates the CPA concentration that is equieffective with the surplus interstitial adenosine concentration accumulated by NBTI.
Determination of cx belonging to the P1b group (that characterizes the effect of NBTI treatment on the interstitial concentration of endogenous adenosine) was a part of both the original and improved forms of our receptor reserve-estimating method, while assessment of cx related to the P2c group (characterizing the effect of FSCPX-NBTI co-treatment on the interstitial level of endogenous adenosine) appeared first in the improved version.
4.5. Correction of Effect Values of Adenosine E/c Curves Distorted by NBTI (Improved Method)
The effect values of adenosine
E/
c curves biased by NBTI were corrected by means of
cx values obtained from CPA
E/
c curves distorted by NBTI. First, the effect belonging to
cx was determined using the Hill equation:
where:
Ex: The effect evoked solely by the extra interstitial adenosine accumulated by NBTI;
cx: The appropriate theoretical CPA concentration provided by Equation (2);
Emax,
EC50,
n: Empirical parameters of the appropriate CPA
E/
c curve (provided by Equation (1)).
When Ex was calculated for the correction of the averaged, only NBTI-treated adenosine E/c curve of the P4 group, cx belonging to the averaged CPA E/c curve of the P1b group (that was also solely NBTI-treated) and empirical parameters of the averaged CPA E/c curve of the P1a group (control) were substituted into Equation (3). In turn, when Ex was computed for correcting the averaged, FSCPX-NBTI co-treated adenosine E/c curve of the P4 group, cx related to the averaged CPA E/c curve of the P2c group (that was FSCPX-NBTI co-treated as well) and empirical parameters of the averaged CPA E/c curve of the P2b group (that was only FSCPX-pre-treated) were written into Equation (3).
From the distorted effects and their corresponding
Ex values, corrected effects were computed by means of Equation (4):
where:
E: The corrected effect;
E’: The effect distorted by the surplus interstitial adenosine produced by NBTI;
Ex: The effect of the extra interstitial adenosine accumulated by NBTI (yielded by Equation (3)).
These corrected effect values (obtained from the procedure described above) are unique in that they reflect the action of NBTI on the adenosine E/c curve without the distortion caused by the interstitially accumulated endogenous adenosine.
4.6. Correction of Effect Values of Adenosine E/c Curves Distorted by NBTI (Original Method)
For the sake of comparison, the averaged, FSCPX-NBTI co-treated adenosine
E/
c curve of the P4 group was corrected with
cx belonging to the averaged CPA
E/
c curve of the P1b group (that was only NBTI-treated) as well, consistent with the procedure used in our previous studies [
18,
19]. (The averaged, NBTI-treated adenosine
E/
c curve of the P4 group was corrected once, because both methods specified the same
cx for this correction)
4.7. Data Analysis
Each atrium was required to meet three criteria in order to qualify for inclusion in the further evaluation: (i) the resting contractile force had to reach 1 mN before the first E/c curve; (ii) the mechanical activity of the paced atrium had to be regular; (iii) the response to 10 μM adenosine obtained from the first E/c curve was required to be within the mean ± 2 SD range (i.e., outliers were excluded). The mean and SD were computed using atria meeting the first two criteria. All experimental outcomes conforming to these three criteria were further processed.
According to the recommendation [
32], concentrations (
c as agonist concentration,
EC50 and
cx) in the equations used for curve fitting were expressed as common logarithms.
Normality of data was checked using Shapiro-Wilk normality test. Two data sets, if passed the normality test, were compared with unpaired t test. If not, Mann-Whitney U test was used. More than two data sets were compared using one-way ANOVA (with Geisser-Greenhouse correction) followed by Tukey post-testing (herein, all data sets undergone multiple comparison passed the normality test). Difference of means (or medians) was considered significant at p < 0.05.
Curve fitting and statistical analysis were performed with GraphPad Prism 7.04 for Windows (GraphPad Software Inc., La Jolla, CA, USA), while other calculations were made by means of Microsoft Excel 2016 (Microsoft Co., Redmond, WA, USA).

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}