Authentication of the Herbal Medicine Angelicae Dahuricae Radix Using an ITS Sequence-Based Multiplex SCAR Assay



Abstract
:1. Introduction
2. Results
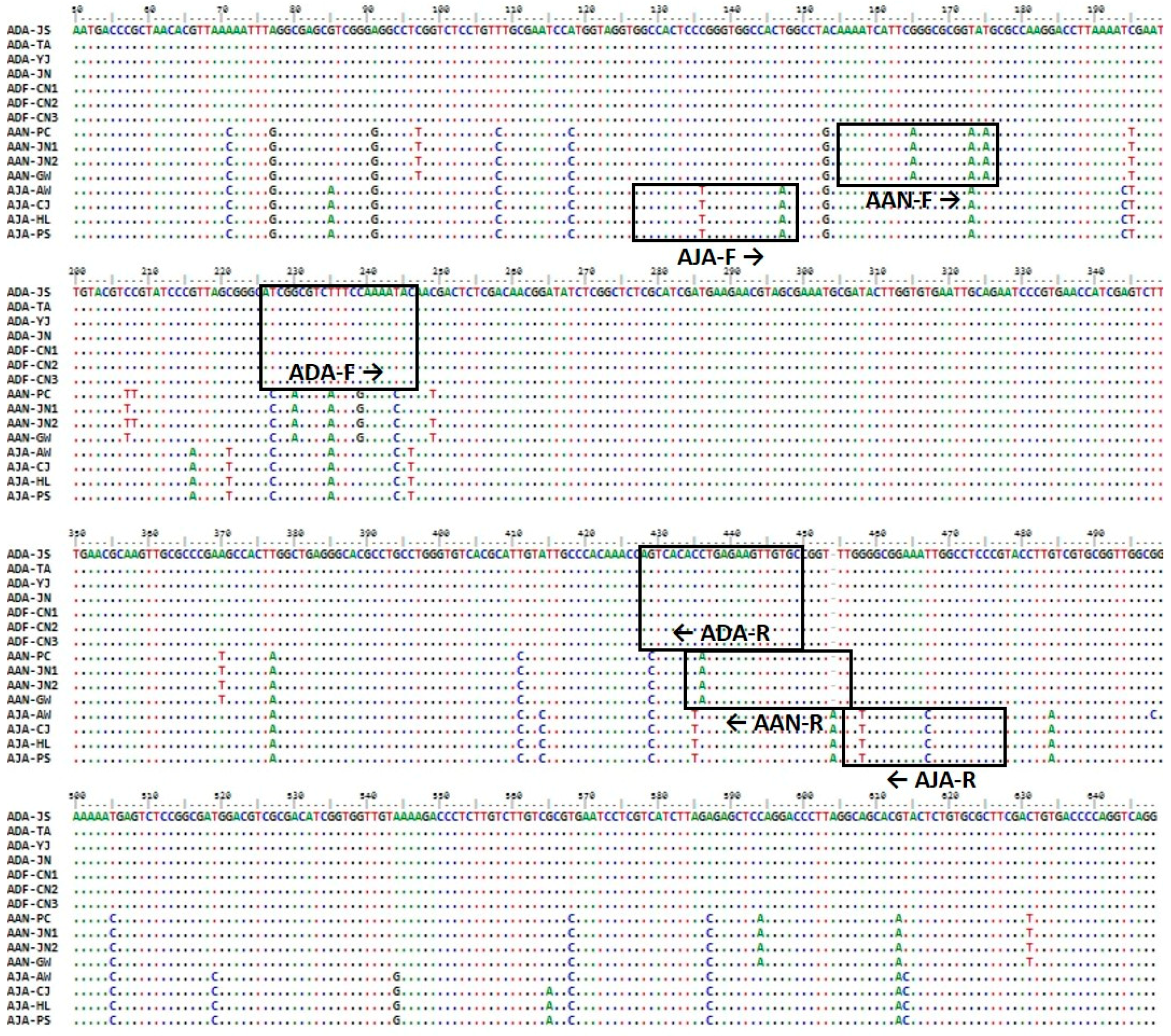
2.1. Nucleotide Sequence and Phylogenetic Analyses
2.2. Development of Species-Specific SCAR Markers
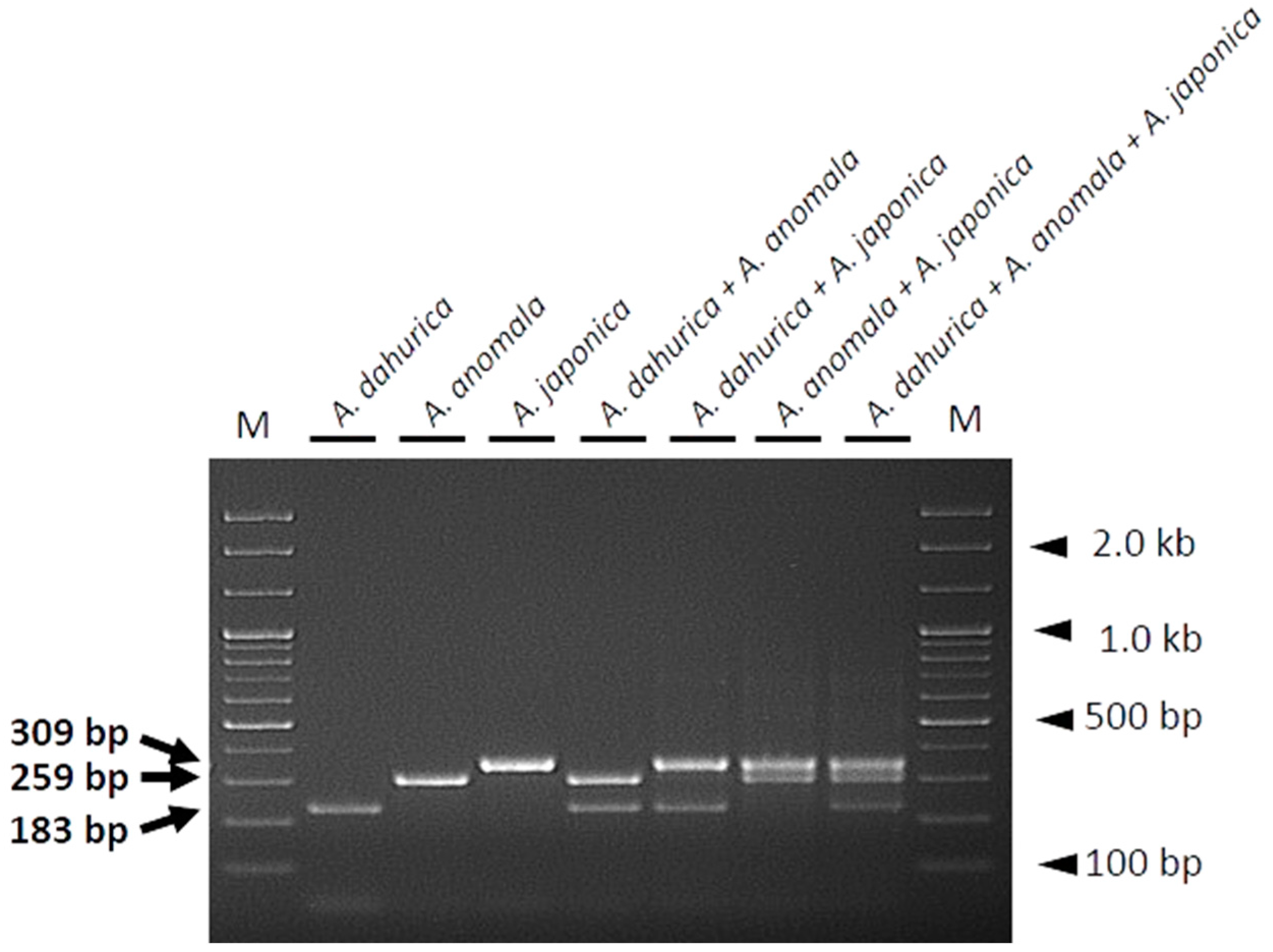
2.3. Development of A Multiplex SCAR Assay for the Authentication of Herbal Medicines
3. Discussion
4. Materials and Methods
4.1. Plant Material and Herbal Medicines
4.2. Genomic DNA Extraction
4.3. PCR Amplification and Sequencing of The ITS Region
4.4. Nucleotide Sequence, Phylogenetic Analyses and SCAR Marker Development
4.5. Development of Multiplex SCAR Assay and Monitoring of Commercial Herbal Medicines
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. WHO Traditional Medicine Strategy: 2014–2023; WHO Press: Geneva, Switzerland, 2013. [Google Scholar]
- Mishra, P.; Kumar, A.; Nagireddy, A.; Mani, D.N.; Shukla, A.K.; Tiwari, R.; Sundaresan, V. DNA barcoding: An efficient tool to overcome authentication challenges in the herbal market. Plant Biotechnol. J. 2016, 14, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Kiran, U.; Khan, S.; Mirza, K.J.; Ram, M.; Abdin, M. SCAR markers: A potential tool for authentication of herbal drugs. Fitoterapia 2010, 81, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Seethapathy, G.S.; Balasubramani, S.P.; Venkatasubramanian, P. nrDNA ITS sequence based SCAR marker to authenticate Aconitum heterophyllum and Cyperus rotundus in Ayurvedic raw drug source and prepared herbal products. Food Chem. 2014, 145, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Pang, X.; Song, J.; Shi, L.; Yao, H.; Han, J.; Leon, C. A renaissance in herbal medicine identification: From morphology to DNA. Biotechnol. Adv. 2014, 32, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sucher, N.J.; Carles, M.C. Genome-based approaches to the authentication of medicinal plants. Planta Med. 2008, 74, 603–623. [Google Scholar] [CrossRef] [PubMed]
- Moon, B.C.; Kim, W.J.; Han, K.S.; Yang, S.; Kang, Y.; Park, I.; Piao, R. Differentiating authentic Adenophorae Rdix from its adulterants in commercially-processed samples using multiplexed ITS sequence-based SCAR markers. Appl. Sci. 2017, 7, 660. [Google Scholar] [CrossRef]
- CBOL Plant Working Group. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and using a plant DNA barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef] [PubMed]
- Techen, N.; Parveen, I.; Pan, Z.; Khan, I.A. DNA barcoding of medicinal plant material for identification. Curr. Opin. Biotechnol. 2014, 25, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.W.; Liao, Y.J.; Hung, Y.S.; Chang, J.C.; Sung, J.M. Development of ITS sequence based SCAR markers for discrimination of Paphiopedilum armeniacum, Paphiopedilum micranthum, Paphiopedilum delenatii and their hybrids. Sci. Hortic. 2011, 127, 405–410. [Google Scholar] [CrossRef]
- Babaei, S.; Talebi, M.; Bahar, M. Developing an SCAR and ITS reliable multiplex PCR-based assay for safflower adulterant detection in saffron samples. Food Control 2014, 35, 323–328. [Google Scholar] [CrossRef]
- Zhu, Y.P. Chinese Materia Medica: Chemistry, Pharmacology and Applications; CRC Press: Boca Raton, FL, USA, 1998; ISBN 978-9-05702-285-2. [Google Scholar]
- Defining Dictionary for Medicinal Herbs in Korea Institute of Oriental Medicine. Available online: http://boncho.kiom.re.kr/codex/ (accessed on 11 June 2018). (In Korean).
- The Korea Food and Drug Administration. The Korean Pharmacophoeia, 11th ed.; Korea Food and Drug Administration: Seoul, Korea, 2014; Volume 2, pp. 43–44. (In Korean)
- National Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2015; Volume 1, pp. 105–106. [Google Scholar]
- Zheng, Y.M.; Shen, J.Z.; Wang, Y.; Lu, A.X.; Ho, W.S. Anti-oxidant and anti-cancer activities of Angelica dahurica extract via induction of apoptosis in colon cancer cells. Phytomedicine 2016, 23, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Q.; Song, Y.L.; Zhu, Z.X.; Su, C.; Zhang, X.; Wang, J.; Shi, S.P.; Tu, P.F. Anti-inflammatory dimeric furanocoumarins from the roots of Angelica dahurica. Fitoterapia 2015, 105, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.W.; Sun, J.G.; Chan, J.Y.W.; Yang, L.; Wu, S.H.; Fung, K.P.; Liu, F.Y. Anticancer effects of imperatorin isolated from Angelica dahurica: Induction of apoptosis in HepG2 cells through both death-receptor-and mitochondria-mediated pathways. Chemotherapy 2011, 57, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.M.; Moon, T.C.; Hong, T.G.; Park, K.M.; Son, J.K.; Chang, H.W. 5-Methoxy-8-(2-hydroxy-3-buthoxy-3-methylbutyloxy)-psoralen isolated from Angelica dahurica inhibits cyclooxygenase-2 and 5-Lipoxygenase in mouse bone marrow-derived mast cells. Arch. Pharm. Res. 2008, 31, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.H.; Chae, H.S.; Oh, Y.C.; Choi, J.G.; Lee, Y.S.; Jang, H.J.; Kim, J.H.; Kim, Y.C.; Sohn, D.H.; Park, H. Anti-nociceptive and anti-inflammatory effects of Angelicae Dahuricae Radix through inhibition of the expression of inducible nitric oxide synthase and NO production. Am. J. Chin. Med. 2008, 36, 913–928. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.J.; Lee, J.Y.; Oh, M.S.; Pak, Y.K.; Park, K.S.; Oh, T.H.; Yune, T.Y. Inhibition of inflammation and oxidative stress by Angelica dahuricae radix extract decreases apoptotic cell death and improves functional recovery after spinal cord injury. J. Neurosci. Res. 2012, 90, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.H.; Kim, J.H.; Park, S.M.; Lee, B.C.; Pyo, H.B.; Park, H.D. New cosmetic agents for skin whitening from Angelica dahurica. J. Cosmet. Sci. 2006, 57, 11–21. [Google Scholar] [PubMed]
- Yuan, Q.J.; Zhang, B.; Jiang, D.; Zhang, W.J.; Lin, T.Y.; Wang, N.H.; Chiou, S.J.; Huang, L.Q. Identification of species and materia medica within Angelica L. (Umbelliferae) based on phylogeny inferred from DNA barcodes. Mol. Ecol. Resour. 2015, 15, 358–371. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Moon, B.C.; Yang, S.; Han, K.S.; Choi, G.; Lee, A.Y. Rapid authentication of the herbal medicine plant species Aralia continentalis Kitag. and Angelica biserrata C.Q. Yuan and R.H. Shan using ITS2 sequences and multiplex-SCAR markers. Molecules 2016, 21, 270. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.D.; Nahar, L. Natural medicine: The genus Angelica. Curr. Med. Chem. 2004, 11, 1479–1500. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Downie, S.R.; Yu, Y.; Zhang, X.; Chen, W.; He, X.; Liu, S. Molecular systematics of Angelica and allied genera (Apiaceae) from the Hengduan Mountains of China based on nrDNA ITS sequences: Phylogenetic affinities and biogeographic implications. J. Plant Res. 2009, 122, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Ganie, S.H.; Upadhyay, P.; Das, S.; Sharma, M.P. Authentication of medicinal plants by DNA markers. Plant Gene 2015, 4, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Heubi, G. New aspects of DNA-based authentication of Chinese medicinal plants by molecular biological techniques. Planta Med. 2010, 76, 1963–1974. [Google Scholar] [CrossRef]
- Ali, M.A.; Gyulai, G.; Hidvegi, N.; Kerti, B.; Al Hemaid, F.M.A.; Pandey, A.K.; Lee, J. The changing epitome of species identification-DNA barcoding. Saudi J. Biol. Sci. 2014, 21, 204–231. [Google Scholar] [CrossRef]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly variable chloroplast markers for evaluating plant phylogeny at low taxonomic levels and for DNA barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Erickson, D.L. DNA barcodes: Genes, genomics, and bioinformatics. Proc. Natl. Acad. Sci. USA 2008, 105, 2761–2762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, B.C.; Kim, W.J.; Park, I.; Sung, G.-H.; Noh, P. Establishment of a PCR assay for the detection and discrimination of authentic Cordyceps and adulterant species in food and herbal medicines. Molecules 2018, 23, 1932. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. PCR protocols: A Guide to Methods and Applicationsl; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Feliner, G.N.; Larena, B.G.; Aguilar, J.F. Fine-scale Geographical Structure, Intra-individual Polymorphism and Recombination in Nuclear Ribosomal Internal Transcribed Spacers in Armeria (Plumbaginaceae). Ann. Bot. 2004, 93, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: An User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. Available online: http://brownlab.mbio.ncsu.edu/JWB/papers/1999Hall1.pdf (accessed on 4 July 2017).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of A. dahurica, A. dahurica var. formosana, A. anomala, A. japonica, and commercial herbal medicines are available from the authors and the Korean Herbarium of Standard Herbal Resources (Index Herbariorum code KIOM). |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Herbal Name | Collection Site | Collection Date | Voucher Number | Abbreviation |
|---|---|---|---|---|---|
| A. dahurica (Hoffm.) Benth. & Hook.f. ex Franch. & Sav. | Angelicae Dahuricae Radix | Beonam, Jangsu, Jeonbuk, Korea | 2015-09-11 | KIOM201501015740 | ADA-JS |
| Iwon, Taean, Chungnam, Korea | 2015-08-25 | KIOM201501015772 | ADA-TA | ||
| Punggi, Yeongju, Gyeongbuk, Korea | 2017-08-09 | KIOM201801020615 | ADA-YJ | ||
| Gohan, Jeongseon, Gangwon, Korea | 2017-08-10 | KIOM201801020618 | ADA-JN | ||
| A. dahurica var. formosana (Boissieu) Yen | Angelicae Dahuricae Radix | Nangang, Harbin, Heilongjiang, China | 2014-08-06 | 2014CHINA1-1 | ADF-CN1 |
| 2014-08-06 | 2014CHINA1-2 | ADF-CN2 | |||
| 2014-08-06 | 2014CHINA1-3 | ADF-CN3 | |||
| A. anomala Avé-Lall. | - | Bongpyeong, Pyeongchang, Gangwon, Korea | 2015-07-29 | KIOM201501015646 | AAN-PC |
| Gohan, Jeongseon, Gangwon, Korea | 2017-08-10 | KIOM201801020679 | AAN-JN1 | ||
| Gohan, Jeongseon, Gangwon, Korea | 2017-08-10 | KIOM201801020685 | AAN-JN2 | ||
| Gunwi, Gyeongbuk, Korea | 2013-07-14 | KIOM201501011489 | AAN-GW | ||
| A. japonica A. Gray | - | Aewol, Jeju, Jeju, Korea | 2007-03-30 | KIOM201501011678 | AJA-AW |
| Chuja, Jeju, Jeju, Korea | 2015-07-15 | KIOM201501015106 | AJA-CJ | ||
| Hallim, Jeju, Jeju, Korea | 2016-12-05 | KIOM201701019409 | AJA-HL | ||
| Pyoseon, Seogwipo, Jeju, Korea | 2016-12-06 | KIOM201801020395 | AJA-PS |
| Species | Length of ITS (bp) | Aligned Length (bp) | Intra-Species Variability (%) 1 | Inter-Species Variability (%) 1 | Species-Specific Mutations | |
|---|---|---|---|---|---|---|
| Indels | Substitutions | |||||
| A. dahurica | 689 | 690 | 0.0000 ± 0.0000 | 0.0476 ± 0.0053 | 0 | 18 |
| A. dahurica var. formosana | ||||||
| A. anomala | 689 | 690 | 0.0017 ± 0.0012 | 0.0423 ± 0.0021 | 0 | 10 |
| A. japonica | 690 | 690 | 0.0015 ± 0.0010 | 0.0461 ± 0.0074 | 1 | 16 |
| Species | Primer Name 1 | Primer Sequence (5′→3′) | PCR Product Size (bp) |
|---|---|---|---|
| A. dahurica | ADA-F | ATCGGCGTCTTTCCAAAATGC | 183 |
| A. dahurica var. formosana | ADA-R | GCACAACTTCTCAGGTGTGCCT | |
| A. anomala | AAN-F | AAAATCATTCAGGCGCGGAGAG | 259 |
| AAN-R | AAACCGGCACAACTTCTCATGT | ||
| A. japonica | AJA-F | GGCCACTCCTGGGTGGCCAGAG | 309 |
| AJA-R | CGGGAGGCCAGTTTCCGCCAGA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noh, P.; Kim, W.J.; Yang, S.; Park, I.; Moon, B.C. Authentication of the Herbal Medicine Angelicae Dahuricae Radix Using an ITS Sequence-Based Multiplex SCAR Assay. Molecules 2018, 23, 2134. https://doi.org/10.3390/molecules23092134
Noh P, Kim WJ, Yang S, Park I, Moon BC. Authentication of the Herbal Medicine Angelicae Dahuricae Radix Using an ITS Sequence-Based Multiplex SCAR Assay. Molecules. 2018; 23(9):2134. https://doi.org/10.3390/molecules23092134
Chicago/Turabian StyleNoh, Pureum, Wook Jin Kim, Sungyu Yang, Inkyu Park, and Byeong Cheol Moon. 2018. "Authentication of the Herbal Medicine Angelicae Dahuricae Radix Using an ITS Sequence-Based Multiplex SCAR Assay" Molecules 23, no. 9: 2134. https://doi.org/10.3390/molecules23092134
APA StyleNoh, P., Kim, W. J., Yang, S., Park, I., & Moon, B. C. (2018). Authentication of the Herbal Medicine Angelicae Dahuricae Radix Using an ITS Sequence-Based Multiplex SCAR Assay. Molecules, 23(9), 2134. https://doi.org/10.3390/molecules23092134