A Comprehensive Genome Survey Provides Novel Insights into Bile Salt Hydrolase (BSH) in Lactobacillaceae
by
 , ,
, ,
Lifeng Liang
1,2,† ,
,
Yunhai Yi
1,3,†,
Yunyun Lv
1,3,
Junwei Qian
2,4,
Xuejing Lei
2,4 and
Gengyun Zhang
1,2,4,* 1
BGI Education Center, University of Chinese Academy of Sciences, Shenzhen 518083, China
2
State Key Laboratory of Agricultural Genomics, BGI-Shenzhen, Shenzhen 518083, China
3
Shenzhen Key Lab of Marine Genomics, Guangdong Provincial Key Lab of Molecular Breeding in Marine Economic Animals, BGI Academy of Marine Sciences, BGI Marine, BGI, Shenzhen 518083, China
4
Key Lab of Genomics, Chinese Ministry of Agriculture, BGI-Shenzhen, Shenzhen 518083, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Molecules 2018, 23(5), 1157; https://doi.org/10.3390/molecules23051157
Submission received: 25 April 2018
/
Revised: 9 May 2018
/
Accepted: 10 May 2018
/
Published: 11 May 2018
Abstract
:Bile salt hydrolase (BSH) is a well-known enzyme that has been commonly characterized in probiotic bacteria, as it has cholesterol-lowering effects. However, its molecular investigations are scarce. Here, we build a local database of BSH sequences from Lactobacillaceae (BSH–SDL), and phylogenetic analysis and homology searches were employed to elucidate their comparability and distinctiveness among species. Evolutionary study demonstrates that BSH sequences in BSH–SDL are divided into five groups, named BSH A, B, C, D and E here, which can be the genetic basis for BSH classification and nomenclature. Sequence analysis suggests the differences between BSH-active and BSH-inactive proteins clearly, especially on site 82. In addition, a total of 551 BSHs from 107 species are identified from 451 genomes of 158 Lactobacillaceae species. Interestingly, those bacteria carrying various copies of BSH A or B can be predicted to be potential cholesterol-lowering probiotics, based on the results of phylogenetic analysis and the subtypes that those previously reported BSH-active probiotics possess. In summary, this study elaborates the molecular basis of BSH in Lactobacillaceae systematically, and provides a novel methodology as well as a consistent standard for the identification of the BSH subtype. We believe that high-throughput screening can be efficiently applied to the selection of promising candidate BSH-active probiotics, which will advance the development of healthcare products in cholesterol metabolism.
1. Introduction
Currently, cardiovascular disease (CVD) is one of the main causes of health problems and mortality worldwide [1]. Although multiple risk factors could give rise to CVD, elevated serum cholesterol is the major cause correlating to the progression of CVD in humans. Numerous epidemiological and clinical studies observed that the high serum level of low-density lipoprotein cholesterol (LDL-C) and total cholesterol (TC) were strongly associated with the incidence of CVD [2]. Cholesterol in the human body is absorbed from food in the small intestine or de-novo synthesized in the liver. Cholesterol is broken down into the primary bile salts cholic acid (CA) or chenodeoxycholic acid (CDCA) under enzymolysis by more than 17 hepatic enzymes. CA is subsequently combined with taurine or glycine, to form taurocholic acid (TCA) and glycocholic acid (GCA), while CDCA utilizes it to synthesize taurochenodeoxycholic acid (TCDCA) and glycochenodeoxycholic acid (GCDCA) [3,4]. The two groups within each macromolecular compound are conjugated. In addition, these four compounds are then accumulated in the body and have been reported to cause adverse effects, including cytotoxicity [5], severe cancer-promoting consequences [6], colonic and oesophageal carcinogenesis in the context of obesity [7,8], and even liver tumour development [9].
In the intestine, TCA, GCA, TCDCA and GCDCA can be decomposed into unconjugated bile acids (CA, CDCA) and amino acid residues via hydrolysis by bile salt hydrolase (BSH, EC 3.5.1.24), which is mainly derived from Bacteroides, Clostridium, Lactobacillus, Bifidobacterium and Listeria. A small proportion of CA and CDCA will be 7α-dehydroxylated into secondary bile salts (DCA and LCA) by the actions of Clostridium and Eubacterium [10]. Farnesoid X receptor (FXR) and G-protein-coupled bile salt receptor 1 (GPBAR1, also known as TGR5) are then stimulated by CA, CDCA, DCA or LCA, in order to regulate host metabolism, such as increasing the cholesterol efflux and limiting the bile salt synthesis in hepatocytes. The majority of CA and CDCA are reabsorbed in the intestinal lumen, and the rest are excreted into feces. Thus, the presence of gastrointestinal bacteria carrying active BSH could accelerate cholesterol metabolism and reduce serum cholesterol levels, thus lowering the risk of CVD [11,12,13,14,15]. Previous studies have indicated that hypercholesterolemic adults treated with certain probiotics, such as Lactobacillus reuteri NCIMB 30242 [16], Lactobacillus acidophilus La5 [17], and Lactobacillus plantarum CECT 7527, 7528 and 7529 [18], could have lower serum TC and LDL-C levels than before. Interestingly, most of these probiotics were proven to be BSH-active. Therefore, BSH is an important enzyme that can affect the regulation of host serum cholesterol metabolism.
BSH is a cytoplasmic enzyme belonging to the N-terminal nucleophile (NTN-) hydrolase superfamily. Variable numbers of bsh genes exist in most Lactobacillaceae species (Table S1), and their function is divergent. For instance, most Lactobacillus plantarum carrying four bsh genes, such as WCFS1 [19] and ST-III [20], were not able to hydrolyze bile salts (BAs) with high efficiency. It has been proposed that penicillin acylase could contribute to the consumption of BAs, as it is also a member of the NTN-hydrolase superfamily [21]. Another study demonstrated that two BSHs had different substrate specificities in Lactobacillus acidophilus NCFM [22]. In addition, only one of the two bsh genes in Lactobacillus fermentum MTCC 8711 performed BSH activity [23]. Moreover, many strains carrying multiple bsh gene copies have been reported to be bile resistant, but the responsible genes have not been identified. Thus, it is challenging to find out the exact BSH enzymes that could hydrolyze BAs efficiently.
Although several bsh gene-sequence analyses have been reported in common probiotics, the bsh gene distribution, evolutionary relationship, and the correlations between sequence characteristics and BA hydrolyzation activity in Lactobacillaceae are still unclear. With the rapid increase in available genome data of Lactobacillaceae, it is possible and imperative to further explore bsh gene biodiversity. In this study, a BSH sequence seed database was built to probe into the characteristics of BSH-active sequences both in their evolutionary relationship and active sites, revealing a reliable model for BSH classification. In addition, a large-scale genome survey in Lactobacillaceae enhances our recognition of BSH-active probiotics. Taken together, this information provides novel insights into bsh in Lactobacillaceae, and a high-throughput approach to select potential cholesterol-lowering probiotics.
2. Results
2.1. Establishment of BSH Database in Lactobacillaceae
A total of 60 bile salt hydrolase protein sequences in 34 Lactobacillaceae species were collected from the Kyoto Encyclopedia of Genes and Genomes (KEGG) database to build a bile salt hydrolase seed database (BSH–SDL) (Table S1). Every BSH protein sequence was selected from the representative strain in each species, except that two strains were selected from Lactobacillus johnsonii.
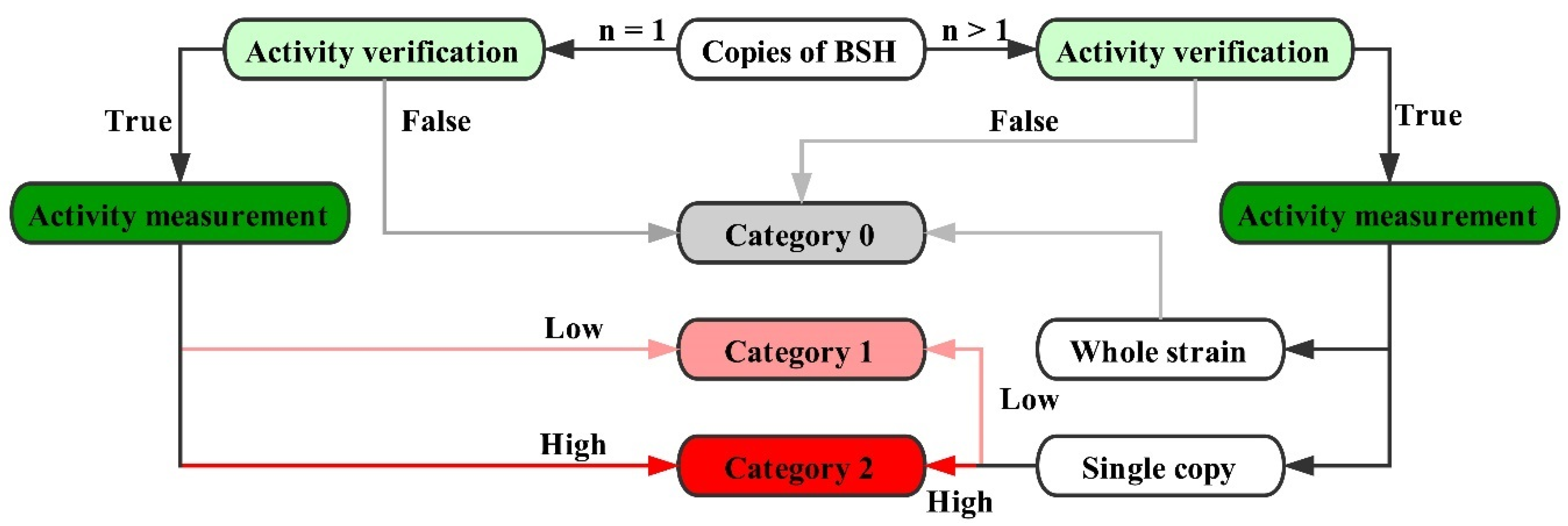
All these sequences were divided into three categories based on their level of biological activation in hydrolyzing BAs, according to previous studies. As shown in Figure 1, if a strain carries multiple bsh gene copies, the sequences with high BSH activity that have been confirmed by previous studies were graded as “2”, while those with low BSH activity were “1”. If a BSH-active strain was found to have only one bsh gene in the genome, this BSH was noted as “2”. If the bsh genes and BSH activity in a strain have not been verified, all BSHs of this strain were noted as “0”. If the relevant experiments for a strain have not been reported, but several bsh-homologous genes exist in the genome, these bsh genes were still marked as “0”. All the scoring numbers are shown in the “Category” column of Table S1. Accurate data collection and complete information for each strain are the fundamental basis of our analysis.
Four bsh genes (lp_0067, lp_2572, lp_3362 and lp_3536) were found in Lactobacillus plantarum WCFS1, and their BSH activities have been verified by molecular cloning experiments. Those results have shown that lp_0067 expressed a BSH which had the highest BSH activity, while other bsh genes had lower activity. Thus, lp_0067 was graded as “2”, while other bsh genes were noted as “1” [19]. The Pediococcus acidilactici genome contains only one bsh gene, and its BSH activity has been reported [24], so this BSH was noted as “2”. Lactobacillus fermentum was proven to have cholesterol-lowering effects and several bsh gene copies were identified; however, the detailed information of functional properties about each bsh gene was not reported. In this case, all bsh genes of Lactobacillus fermentum were marked with “0” [23]. Although Pediococcus damnosus possesses one bsh gene, related BSH activity properties have not been reported yet, so it was noted as “0”. Therefore, there are 18 sequences with high BSH activity, four with lower BSH activity, and 38 without BSH-active records in total in BSH–SDL.
2.2. Phylogenetic Analysis
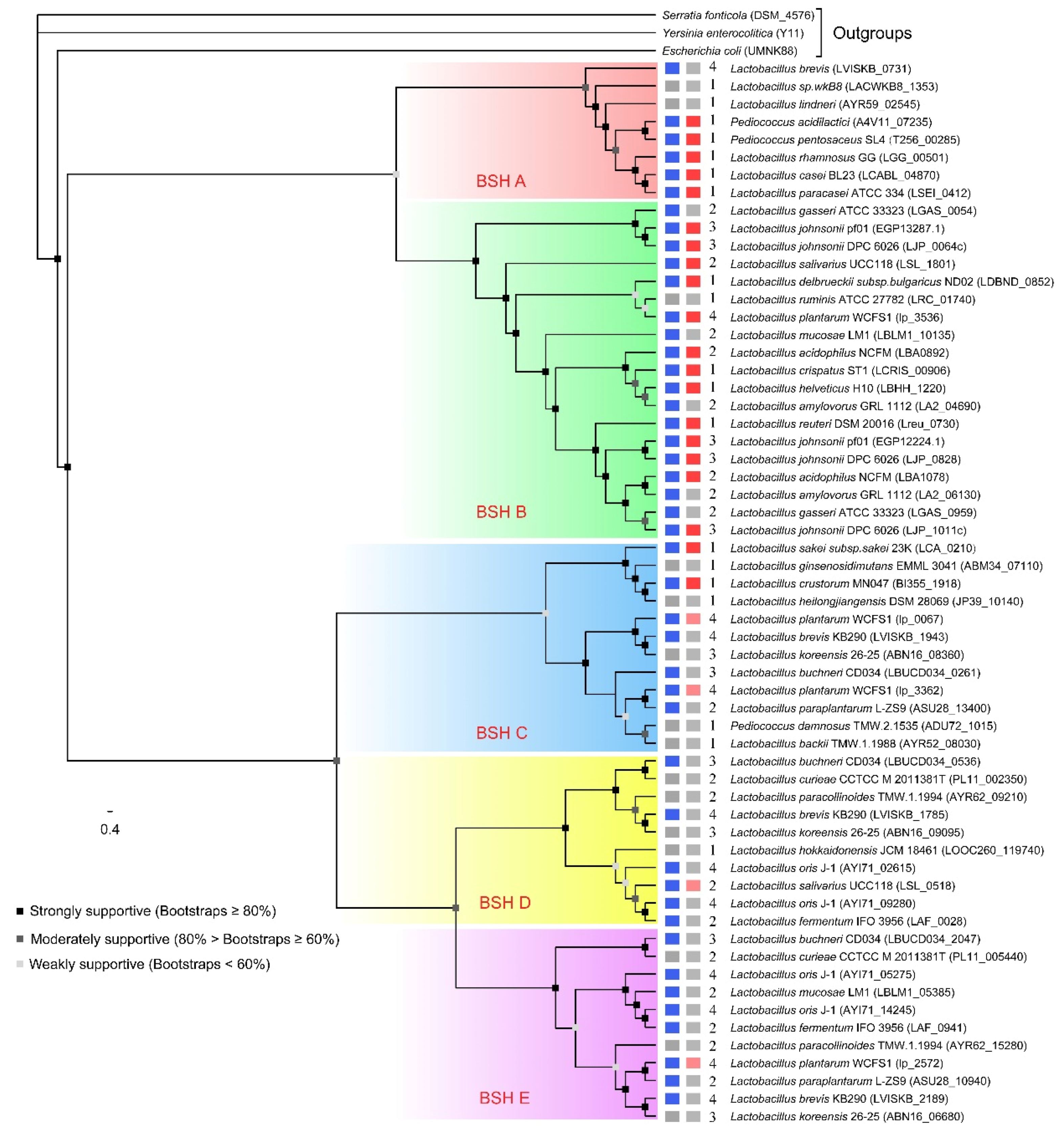
Phylogenetic analysis was employed to reveal the evolutionary relationship among BSH proteins in BSH–SDL. Five distinct clades obviously emerged from the phylogenetic tree (Figure 2). It is interesting to note that almost all BSHs with high activity are gathered in two clades. Therefore, five subtypes of BSH are presumed to exist in Lactobacillaceae species based on the results, and here we named them “BSH A”, “BSH B”, “BSH C”, “BSH D” and “BSH E”. Most BSH A and BSH B proteins have been reported to exhibit greater BSH activity according to previous studies, while the majority of BSH C, BSH D and BSH E proteins have not been verified to be BSH-active yet.
There are eight proteins classified in the BSH A subtype, including five BSH sequences verified in Pediococcus (Pediococcus acidilactici and Pediococcus pentosaceus), Lactobacillus casei group (Lactobacillus casei and Lactobacillus paracasei), Lactobacillus rhamnosus, and three BSHs with unknown activity, which were predicted from Lactobacillus brevis, Lactobacillus lindneri and Lactobacillus sp. wkB8. There are 18 BSH proteins from 12 species in the BSH B subtype, and they have been identified in commonly applied probiotics, including Lactobacillus plantarum, Lactobacillus gasseri, Lactobacillus johnsonii, Lactobacillus salivarius, Lactobacillus reuteri, Lactobacillus acidophilus and Lactobacillus delbrueckii subsp. bulgaricus, Lactobacillus mucosae, Lactobacillus amylovorus, and Lactobacillus helveticus, except an unproven BSH in Lactobacillus ruminis (Figure 2).
Two proteins with high BSH activity from Lactobacillus sakei and Lactobacillus crustorum were assigned to the BSH C subtype group, while the other proteins in this group have not demonstrated high BSH activity. The majority of proteins in BSH C, BSH D and BSH E were predicted to be bile salt hydrolases but without experimental substantiation. They mainly come from species with multiple BSH copies, except for four BSHs with relatively lower activity in Lactobacillus plantarum and Lactobacillus salivarius. Moreover, a strain with multiple BSH copies has diverse BSH subtypes. For instance, four BSHs of Lactobacillus plantarum are classified into three BSH subtypes, BSH B, BSH C and BSH E, and four BSH copies of Lactobacillus brevis were distributed in BSH A, BSH C, BSH D and BSH E subtypes.
2.3. Sequence Analysis of BSH
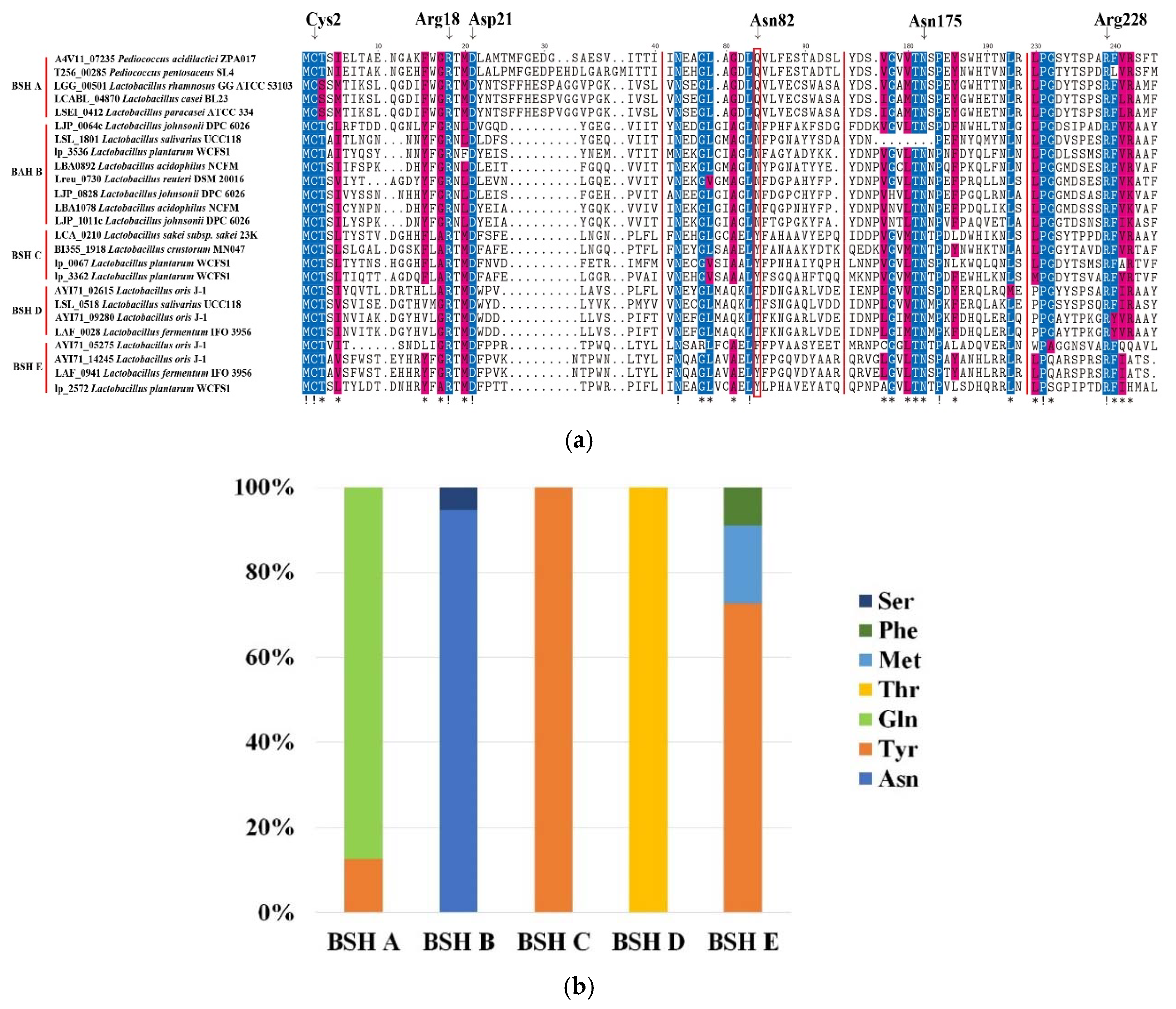
Previous study has identified six catalytic residues as active sites of BSH, including Cys2, Arg18, Asp21, Asn82, Asn175 and Arg228 [25]. Multiple-sequence alignment of BSH protein sequences in BSH–SDL showed that Cys2, Arg18, Asp21, Asn175 and Arg228 were highly conserved among all BSH subtypes, whereas Cys2, Arg18 and Asp21 were absent in few BSH A sequences (Figure 3a, Table S2). The site 82 was highly diverse in comparison to five other sites, including the Ser, Phe, Met, Thr, Gln, Tyr and Asn types (Figure 3b). Gln82 and Asn82 were commonly seen in this catalytic site of BSH A and BSH B, accounting for 87.5% and 94.74% in all protein sequences, respectively. In addition, the site 82 of BSH C and BSH D were all Tyr82 and Thr82, respectively, while BSH E possessed three types, including Tyr82 (72.73%), Met82 (18.18%) and Phe82 (9.09%) (Table S2). It was indicated that the site 82 was closely related to the activity of BSH, and Gln82 and Asn82 were assumed to be essential requirements for BSH with high hydrolase activity. Furthermore, the residues near catalytic sites were rather conserved, while the other sites were not. It is demonstrated that although BSH protein sequences have low homology, some of them still have a similar ability to perform hydrolytic reactions.
2.4. Genome Survey of bsh Genes in Lactobacillaceae Species
Of 194 Lactobacillaceae species in the database of the National Center for Biotechnology Information (NCBI), 158 species had records of whole genome data by the end of year 2017. A total of 451 genomes from 158 Lactobacillaceae species were obtained to search for bsh genes. As a result, 551 BSH-homologous proteins were identified from 107 species. bsh-homologous genes were absent in 51 species. Among those species abundant in BSH-homologous proteins, which accounted for almost one-third of the species surveyed, Lactobacillus plantarum, Lactobacillus oris and Lactobacillus brevis are known as BSH-rich species, and eight BSH-rich species with more than three newly detected bsh genes were discovered, including Lactobacillus acidifarinae, Lactobacillus antri, Lactobacillus farraginis, Lactobacillus odoratitofui, Lactobacillus rapi, Lactobacillus spicheri, Lactobacillus zymae and Lactobacillus parabuchneri.
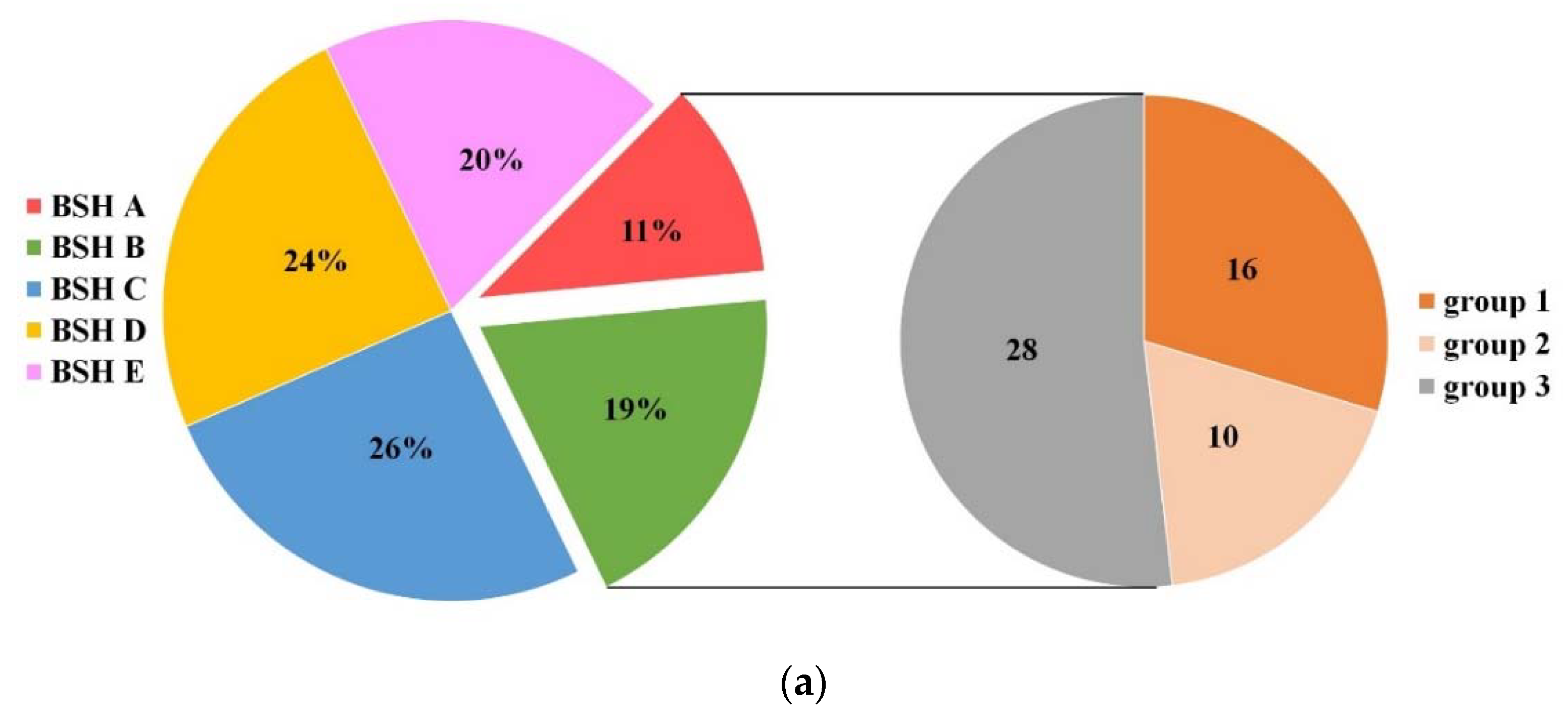
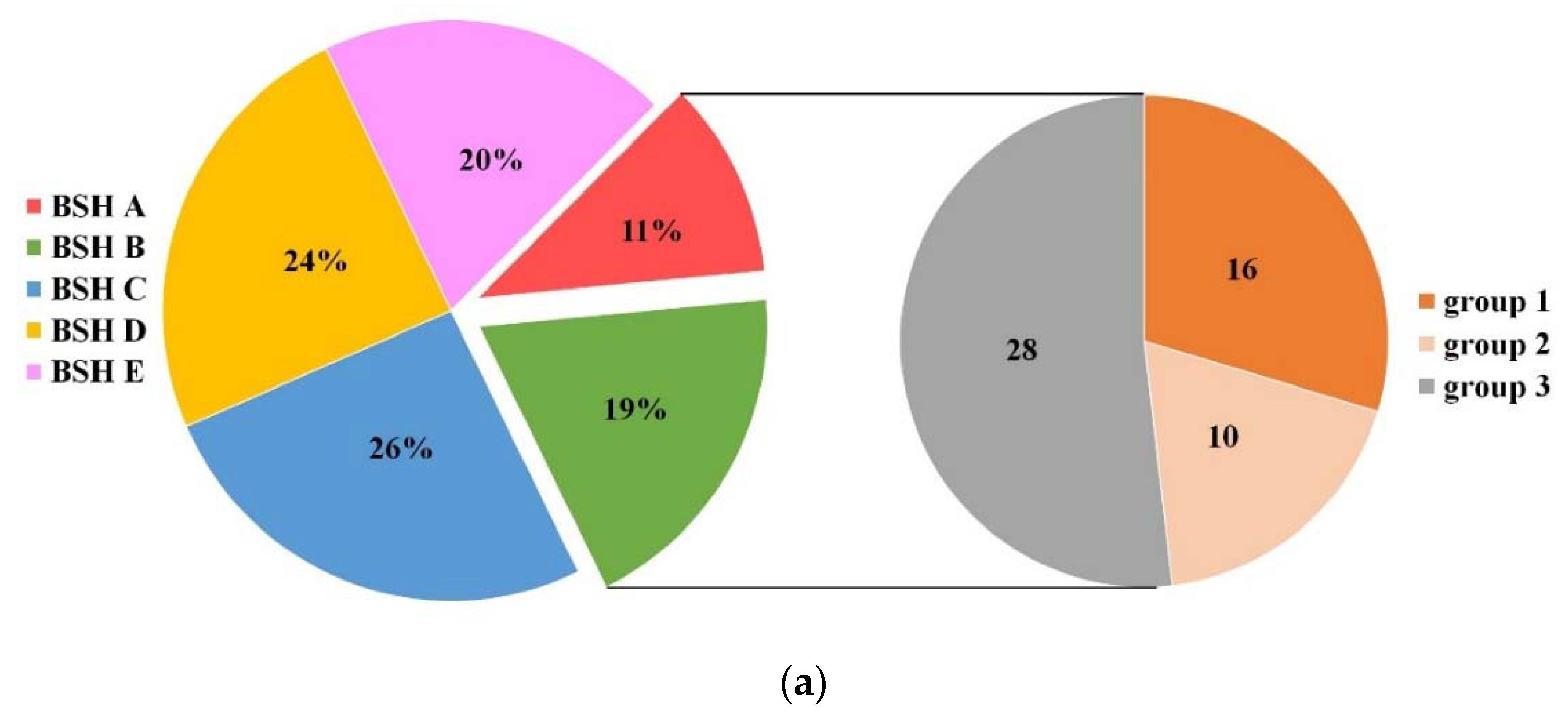
Homology searches identified the BSH subtypes of 551 BSH homologous proteins via their matched protein sequences in BSH–SDL. BSH proteins of the BSH C subtype were the most abundant BSHs in Lactobacillaceae species, while BSH A had the fewest (Figure 4a, Table S3). In addition, BSH A and BSH B proteins were identified in 54 species, and apart from the 16 species in BSH–SDL that exhibited BSH activity, 38 species were predicted to have potentially high BSH activity based on the results of phylogenetic and catalytic-residue analysis in this study. Interestingly, reported BSH activity studies suggest that 10 species of these 38 species are capable of hydrolyzing BAs or have resistance to bile, including Lactobacillus agilis, Lactobacillus animalis, Lactobacillus gallinarum, Lactobacillus helveticus, Lactobacillus ingluviei, Lactobacillus intestinalis, Lactobacillus kefiranofaciens, Lactobacillus panis, Lactobacillus taiwanensis and Lactobacillus vaginalis, and the other 28 species do not yet have relevant research conducted on them, including Lactobacillus lindneri, Lactobacillus hominis, Lactobacillus harbinensis, Lactobacillus ruminis, Lactobacillus intestinalis and so on (Figure 4a, Table S4). In Table 1, several representative BSH-active strains with diverse BSH subtypes are listed. It is demonstrated that our predictive results of BSH subtypes identified in their whole genome are consistent with their biological activities verified by previous studies.
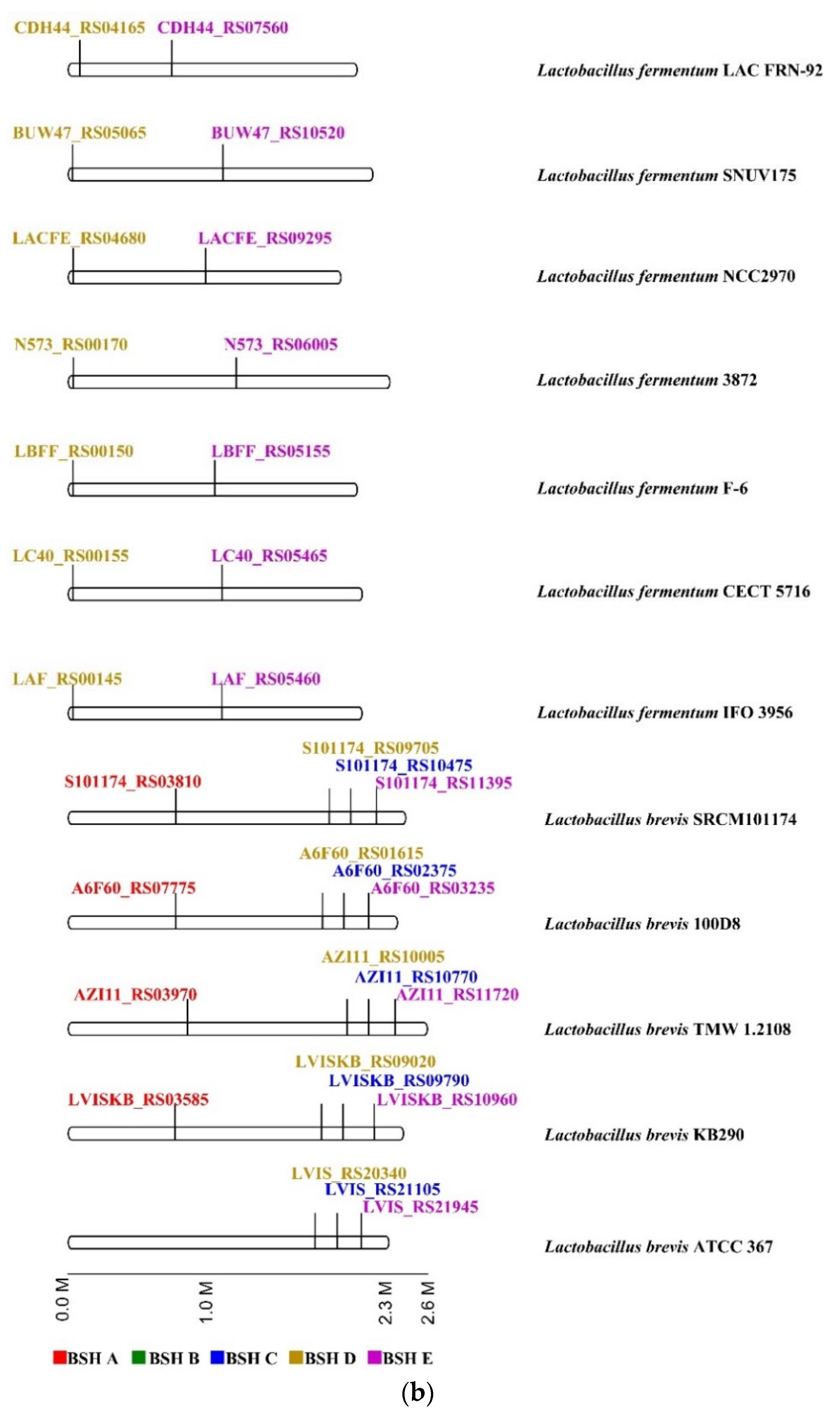
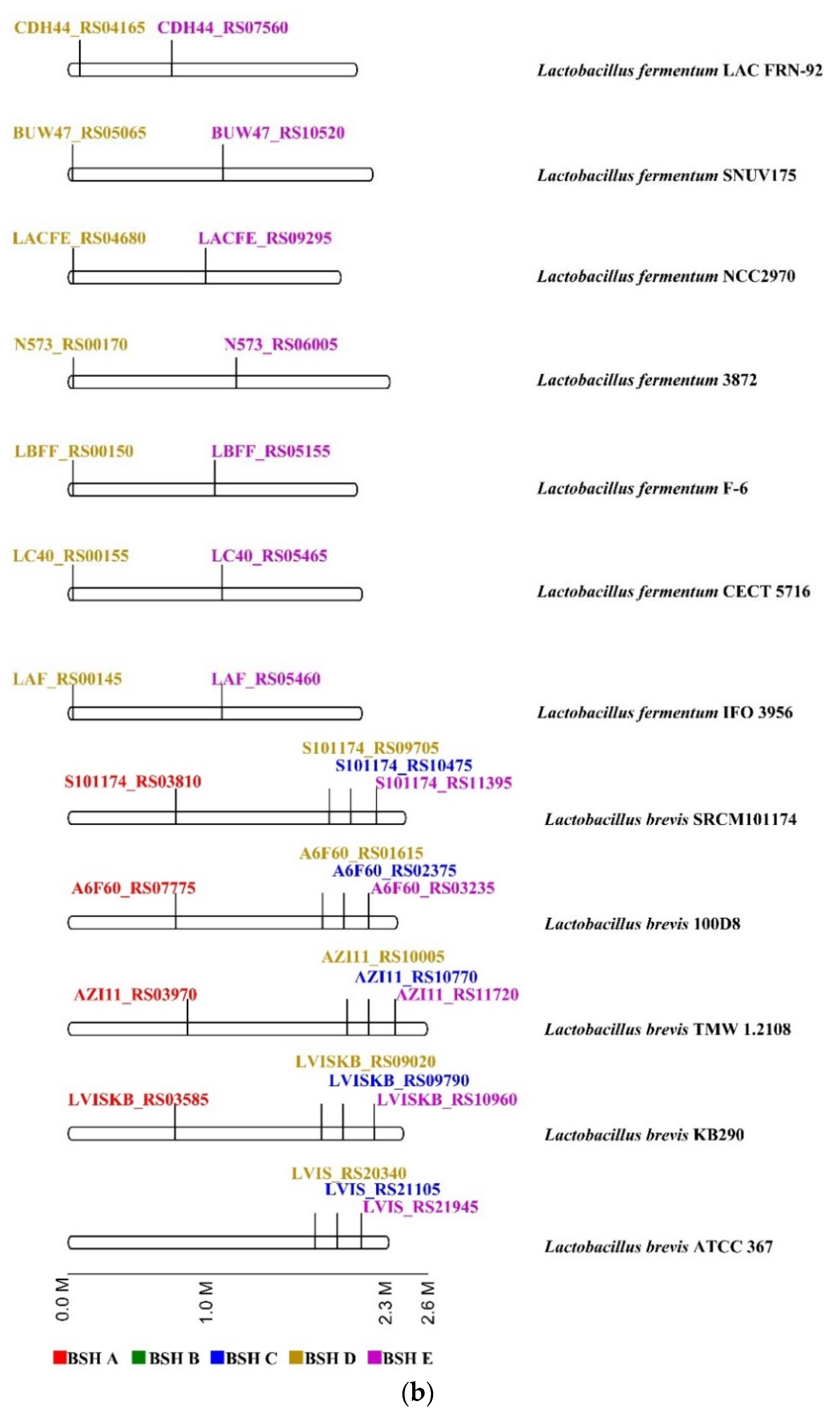
Moreover, the complete genomes of species carrying multiple bsh copies were employed to investigate the genomic location of bsh genes. It was revealed that various BSH subtype genes of one strain are not clustered. Concurrently, the locations of one specific bsh gene subtype in different strains of the same species are relatively fixed (Figure 4b, Figure S1). For instance, it is fully illustrated that two different BSH subtype genes of Lactobacillous fermentum are distributed in the different positions of seven complete genomes of Lactobacillous fermentum strains, and four BSH subtypes of Lactobacillous brevis are located, in a relatively fixed positioning pattern, at five genomes of strains in this species (Figure 4b). The same results were also exhibited in other Lactobacillaceae species and strains (Table S5), while different species showed different bsh distribution patterns. Therefore, it is demonstrated that bsh genes have significant variation among species, while they are comparatively conserved within strains of one species.
3. Discussion
BSHs have attracted much attention because of their vital role in cholesterol metabolism, which makes them a common indicator for screening probiotics. Although several BSH studies have been investigated in a number of strains, in-depth exploration in Lactobacillaceae is limited. Here, a comprehensive molecular investigation into BSHs in Lactobacillaceae was performed. In this study, BSHs of Lactobacillaceae were divided into five subtypes based on their phylogenetic relationship, and BSH A and BSH B are two subtypes possessing high hydrolysis activity. Six catalytic residues were compared among 60 proteins of five BSH subtypes. It was observed that the diversity of amino acid residues in site 82 was a key factor to distinguish BSH subtypes. Identification of BSH-homologous proteins in 451 genomes from 158 Lactobacillaceae species revealed that 54 species were predicted to be BSH-active bacteria, as bsh genes of BSH A or BSH B subtype exist in their genomes. While 28 species have not been reported with any BSH-related research, they are regarded as potential bacterial strains with BSH activity. For example, Lactobacillus intestinalis was isolated from the intestinal tract and it was reported that a high-fat diet changed its abundance in the gastrointestinal tract of rats [32]; however, detailed BSH verification and other characteristics were unclear. Therefore, these 28 species could be valuable resources to discover novel probiotics with higher BSH activity, and this study provides new clues to related intestinal microbiology studies.
Some Lactobacillaceae species carrying multiple BSH copies have been reported in previous studies, but their functional annotations and nomenclature of BSH subtypes were not consistent among species. Therefore, a standardized BSH classification, which is based on phylogenetic analysis of BSH proteins on BSH–SDL, is provided in this study. Newly discovered BSH protein sequences could distinguish their subtypes by referring to five BSH subtypes proposed in our analysis results. It would be helpful for later researchers to recognize BSH protein sequences comprehensively and to share their research results more easily. This high-throughput approach bridges the gap between traditional isolation analysis and bioinformatics.
In the KEGG database, 34 Lactobacillaceae species carrying bsh genes were recorded, while in this study, BSH-homologous protein sequences have been identified in 107 Lactobacillaceae species. Many studies have demonstrated that BSH A and BSH B subtype proteins had greater BSH activity. Therefore, the combination of previous observations and BSH phylogenetic relationship analysis results could be employed to infer whether a strain expressing BSH-homologous protein exhibits BSH-active ability or not. In our results, Lactobacillus sp. wk88, Lactobacillus lindneri, Lactobacillus heilongjiangensis, Lactobacillus ginsenosidimutans and Lactobacillus ruminis are considered as potential BSH-active probiotics (Figure 2). In addition, it is practicable to identify the exact contributory molecules for cholesterol-lowering effects in multiple BSH proteins of Lactobacillus mucosae LM1 (LBLM1_10135). For Lactobacillus amylovorus GRL 1112 and Lactobacillus gasseri ATCC 33323, two BSH proteins in each are predicted to be BSH-active in hydrolyzing BAs.
Although amino acid residues of site 82 in BSH proteins are predicted to be closely related to BSH activity in this study, site-directed mutagenesis of bsh genes on this site was rarely found to provide direct proof of this conclusion. Experimental data from Lactobacillus salivarius [33] seemed to support this point indirectly, as it showed that the BSH mutation of Asn82 into Ala82 significantly decreased its capability of catalyzing TCA, while other mutant amino acids in other sites also influenced the BSH activity. However, catalytic residues of Cys2, Arg18, Asp21, Asn175 and Arg228 are highly conserved, and it would be easier to distinguish the biological effects of amino acid residues in site 82 and to explore their relationship with BSH-active subtypes. Our analysis provides new site information and sequence characteristics for further BSH exploration.
4. Materials and Methods
4.1. Establishment of BSH–SDL
All of the bile salt hydrolase (choloylglycine hydrolase, ko: K01442) protein sequences from Lactobacillaceae, which have records in the KEGG database (http://www.kegg.jp/dbget-bin/www_bget?ko:K01442), were collected in our local database. Those redundant protein sequences were removed by mutual BLASTP alignment (e-value: 1e-10), and sequences with more than 90% identity were deleted. A nonredundant bsh-seed database BSH–SDL was built for further analysis.
4.2. Identification of BSH in Lactobacillaceae
A total of 194 Lactobacillaceae species were recorded in NCBI by the end of August 2017, and 158 species have genome data, thus a total of 451 whole-genome-derived protein sequence datasets of Lactobacillaceae strains from 158 Lactobacillaceae species were retrieved from the NCBI RefSeq database (https://www.ncbi.nlm.nih.gov/) (Table S6). Subsequently, protein sequences in BSH–SDL were employed to search against 451 protein datasets using BLASTP (e-value: 1e-5), in order to identify homologous BSH sequences in different strains. Those hits with identity less than 60% were removed, and the best hit was retained for the following analysis using an in-house pipeline.
4.3. Genomic Location of bsh
The whole complete genomes from 451 Lactobacillaceae strains were selected to investigate the genomic location of bsh genes. To normalize the starting position of the 451 genomes, the dnaA gene was regarded as the initial gene. The gene location had been readjusted by an in-house Python script if dnaA was not the initial gene in the records of NCBI. The presentation of genomic location of bsh was accomplished by TBtools software (version 0.49991, CJ-Chen, Guangzhou, China) [34].
4.4. Sequence Alignment and Phylogenetic Analysis
The protein sequences in BSH–SDL were used for the construction of the phylogenetic tree. At first, MUSCLE (version 3.7, CV, Mill Valley, CA, USA) [35] was employed to perform multiple sequence alignment with default parameters. Sequence analysis was further shown by TeXshade (version 1.25, Eric Beitz, Tübingen, Germany) [36]. Then, the phylogenetic tree was constructed by PhyML (version 3.1, LIRMM, Université de Montpellier, Montpellier, France) [37] with best-fit protein evolution model (WAG+I+G+F), which was confirmed by Prottest 3 software (version 3.4.2, University of Vigo, Vigo, Spain) [38], and with a bootstrapping value of 1000. Final tree visualization was processed by iToL (version 4.2, biobyte solutions GmbH, Heidelberg, Germany) [39].
5. Conclusions
In this study, the genetic basis of bsh genes in Lactobacillaceae species has been explored systematically. Phylogenetic analysis indicates that BSH can be classified into five subtypes, which provides a general principle for future nomenclature of BSH protein. Further multiple sequence alignment demonstrates distinct characteristics of BSH A, B, C, D and E in Lactobacillaceae, particularly in site 82. We point out that Gln82 and Asn82 are two critical amino acids distinguishing BSH-active subtype proteins from other BSHs with lower BA hydrolysis activity. In addition, a large-scale high-throughput identification of bsh genes in genomes of all available Lactobacillaceae species provides a novel approach to select potential BSH-active candidate probiotics for cholesterol-lowering effects. Genome location of bsh in various species also indicates its conservation and diversity in Lactobacillaceae. These findings provide novel insights into BSH, and valuable resources for further research on cholesterol-lowering probiotics.
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/1420-3049/23/5/1157/s1. Figure S1: Genome location of bsh genes in 13 Lactobacillaceae species; Table S1: Detailed information about protein sequences in BSH–SDL; Table S2: Statistics of six catalytic residues of BSH subtypes from all BSH protein sequences in BSH–SDL; Table S3: Statistics of mapped BSH subtypes in 158 species; Table S4: Statistics of mapped information about BSH A and BSH B in 54 species; Table S5: The genomic location of bsh in representative Lactobacillaceae species; Table S6: The NCBI ftp list of 451 Lactobacillaceae genomes.
Author Contributions
G.Z. and L.L. conceived and designed the study; L.L. analyzed the data; Y.L. performed the phylogenetic analysis; J.Q. and X.L. participated in discussions and provided resource support; L.L. and Y.Y. wrote the paper.
Funding
This research was funded by Shenzhen Municipal Government grant number No. KY20160306.
Acknowledgments
We would like to thank Guangdong Province key laboratory of crop germplasm research and application for assistance with computing resources support.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| CVD | cardiovascular disease |
| TC | total cholesterol |
| LDL-C | low-density lipoprotein cholesterol |
| BAs | bile acids |
| CA | cholic acid |
| CDCA | chenodeoxycholic acid |
| TCA | taurocholic acid |
| TCDCA | taurochenodeoxycholic acid |
| GCA | glycocholic acid |
| GCDCA | glycochenodeoxycholic acid |
| BSH | bile salt hydrolase |
| BSH–SDL | bile salt hydrolase seed database of Lactobacillaceae |
| FXR | farnesoid X receptor |
| GPBAR1/TGR5 | G-protein-coupled bile acid receptor 1 |
References
- Lee, C.M.Y.; Barzi, F.; Woodward, M.; Batty, G.D.; Giles, G.G.; Wong, J.W.; Jamrozik, K.; Lam, T.H.; Ueshima, H.; Kim, H.C.; et al. Adult height and the risks of cardiovascular disease and major causes of death in the Asia-Pacific region: 21,000 deaths in 510,000 men and women. Int. J. Epidemiol. 2009, 38, 1060–1071. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Liu, X.M.; Zhang, Q.X.; Shen, Z.; Tian, F.W.; Zhang, H.; Sun, Z.H.; Zhang, H.P.; Chen, W. Influence of consumption of probiotics on the plasma lipid profile: A meta-analysis of randomised controlled trials. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E.; Kok, E.; Javitt, N.B. Bile acid synthesis in man: Metabolism of 7α-hydroxycholesterol-(14)C and 26-hydroxycholesterol-(3)H. J. Clin. Investig. 1972, 51, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Xie, G.; Jia, W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.; Jaeschke, H.; Copple, B.L. Bile acids induce inflammatory genes in hepatocytes: A novel mechanism of inflammation during obstructive cholestasis. Am. J. Pathol. 2011, 178, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, H.; Bernstein, C.; Payne, C.M.; Dvorak, K. Bile acids as endogenous etiologic agents in gastrointestinal cancer. World J. Gastroenterol. WJG 2009, 15, 3329–3340. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, C.; Holubec, H.; Bhattacharyya, A.K.; Nguyen, H.; Payne, C.M.; Zaitlin, B.; Bernstein, H. Carcinogenicity of deoxycholate, a secondary bile acid. Arch. Toxicol. 2011, 85, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Quante, M.; Bhagat, G.; Abrams, J.; Marache, F.; Good, P.; Lee, M.D.; Lee, Y.; Friedman, R.; Asfaha, S.; Dubeykovskaya, Z.; et al. Bile acid and inflammation activate gastric cardia stem cells in a mouse model of Barrett’s-like metaplasia. Cancer Cell 2012, 21, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Wang, X.; Huang, F.; Zhao, A.; Chen, W.; Yan, J.; Zhang, Y.; Lei, S.; Ge, K.; Zheng, X.; et al. Dysregulated hepatic bile acids collaboratively promote liver carcinogenesis. Int. J. Cancer 2016, 139, 1764–1775. [Google Scholar] [CrossRef] [PubMed]
- Molinaro, A.; Wahlström, A.; Marschall, H.-U. Role of bile acids in metabolic control. Trends Endocrinol. Metab. 2018, 29, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, X.; Wang, J.; Wu, F.; Sui, Y.; Yang, L.; Wang, Z. Lactobacillus plantarum strains as potential probiotic cultures with cholesterol-lowering activity. J. Dairy Sci. 2013, 96, 2746–2753. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp. Diabetes Res. 2012, 2012, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ooi, L.-G.; Liong, M.-T. Cholesterol-lowering effects of probiotics and prebiotics: A review of in vivo and in vitro findings. Int. J. Mol. Sci. 2010, 11, 2499–2522. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, G.; Liu, X.-M.; Zhang, Q.-X.; Tian, F.-W.; Zhang, H.; Zhang, H.-P.; Chen, W. Research advances with regards to clinical outcome and potential mechanisms of the cholesterol-lowering effects of probiotics. Clin. Lipidol. 2012, 7, 501–507. [Google Scholar] [CrossRef]
- Chand, D.; Avinash, V.S.; Yadav, Y.; Pundle, A.V.; Suresh, C.G.; Ramasamy, S. Molecular features of bile salt hydrolases and relevance in human health. Biochim. Biophys. Acta 2017, 1861, 2981–2991. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Martoni, C.J.; Parent, M.; Prakash, S. Cholesterol-lowering efficacy of a microencapsulated bile salt hydrolase-active Lactobacillus reuteri NCIMB 30242 yoghurt formulation in hypercholesterolaemic adults. Br. J. Nutr. 2012, 107, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V.; Akbarian-Moghari, A. Effect of probiotic yogurt containing Lactobacillus acidophilus and Bifidobacterium lactis on lipid profile in individuals with type 2 diabetes mellitus. J. Dairy Sci. 2011, 94, 3288–3294. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, M.C.; Lajo, T.; Carrión, J.M.; Cuñé, J. Cholesterol-lowering efficacy of Lactobacillus plantarum CECT 7527, 7528 and 7529 in hypercholesterolaemic adults. Br. J. Nutr. 2013, 109, 1866–1872. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.M.; Bongers, R.S.; de Vos, W.M.; Kleerebezem, M. Functional analysis of four bile salt hydrolase and penicillin acylase family members in Lactobacillus plantarum WCFS1. Appl. Environ. Microbiol. 2008, 74, 4719–4726. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Sun, K.; Wu, Z.; Yao, J.; Guo, B. All 4 bile salt hydrolase proteins are responsible for the hydrolysis activity in Lactobacillus plantarum ST-III. J. Food Sci. 2011, 76, M622–M628. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.S.; Brannigan, J.A.; Prabhune, A.A.; Pundle, A.V.; Dodson, G.G.; Dodson, E.J.; Suresh, C.G. Structural and functional analysis of a conjugated bile salt hydrolase from Bifidobacterium longum reveals an evolutionary relationship with penicillin V acylase. J. Biol. Chem. 2006, 281, 32516–32525. [Google Scholar] [CrossRef] [PubMed]
- McAuliffe, O.; Cano, R.J.; Klaenhammer, T.R. Genetic analysis of two bile salt hydrolase activities in Lactobacillus acidophilus NCFM. Appl. Environ. Microbiol. 2005, 71, 4925–4929. [Google Scholar] [CrossRef] [PubMed]
- Jayashree, S.; Pooja, S.; Pushpanathan, M.; Rajendhran, J.; Gunasekaran, P. Identification and characterization of bile salt hydrolase genes from the genome of Lactobacillus fermentum MTCC 8711. Appl. Biochem. Biotechnol. 2014, 174, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Asghar, S.; Arif, M.; Nawaz, M.; Muhammad, K.; Ali, M.A.; Ahmad, M.D.; Iqbal, S.; Anjum, A.A.; Khan, M.; Nazir, J. Selection, characterisation and evaluation of potential probiotic Lactobacillus spp. isolated from poultry droppings. Benef. Microbes 2016, 7, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Lodola, A.; Branduardi, D.; De Vivo, M.; Capoferri, L.; Mor, M.; Piomelli, D.; Cavalli, A. A Catalytic mechanism for cysteine N-Terminal nucleophile hydrolases, as revealed by free energy simulations. PLoS ONE 2012, 7, e32397. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Gu, Q. Complete genome sequence of Lactobacillus plantarum LZ95, a potential probiotic strain producing bacteriocins and B-group vitamin riboflavin. J. Biotechnol. 2016, 229, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Song, S.; Huang, X.; Feng, C.; Liu, G.; Huang, J.; Xie, M. Screening and identification of functional Lactobacillus specific for vegetable fermentation. J. Food Sci. 2012, 78, M84–M89. [Google Scholar] [CrossRef] [PubMed]
- Deraz, S.F.; Karlsson, E.N.; Khalil, A.A.; Mattiasson, B. Mode of action of acidocin D20079, a bacteriocin produced by the potential probiotic strain, Lactobacillus acidophilus DSM 20079. J. Ind. Microbiol. Biotechnol. 2007, 34, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.P.; Valeriano, V.D.; Kim, G.-B.; Kang, D.-K. Molecular cloning, characterization and comparison of bile salt hydrolases from Lactobacillus johnsonii PF01. J. Appl. Microbiol. 2013, 114, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Allain, T.; Chaouch, S.; Thomas, M.; Vallée, I.; Buret, A.G.; Langella, P.; Grellier, P.; Polack, B.; Bermúdez-Humarán, L.G.; Florent, I. Bile-salt-hydrolases from the probiotic strain Lactobacillus johnsonii La1 mediate anti-giardial activity in vitro and in vivo. Front. Microbiol. 2017, 8, 2707. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, C.-L.; Sun, Y.; Li, A.-L.; Liu, F.; Meng, X.-C. Microencapsulation of Lactobacillus rhamnosus GG by transglutaminase cross-linked soy protein isolate to improve survival in simulated gastrointestinal conditions and yoghurt. J. Food Sci. 2016, 81, M1726–M1734. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, V.; Kaakoush, N.O.; Maloney, C.A.; Raipuria, M.; Huinao, K.D.; Mitchell, H.M.; Morris, M.J. Changes in gut microbiota in rats fed a high fat diet correlate with obesity-associated metabolic parameters. PLoS ONE 2015, 10, e0126931. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Fang, F.; Qiu, Y.; Yang, Q.; Chen, J. Substrate specificities of bile salt hydrolase 1 and its mutants from Lactobacillus salivarius. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2014, 30, 445–454. [Google Scholar]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a toolkit for biologists integrating various HTS-data handling tools with a user-friendly interface. bioRxiv 2018. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Beitz, E. TeXshade: Shading and labeling of multiple sequence alignments using LaTeX2e. Bioinformatics 2000, 16, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available. |
Figure 1.
Scoring rules of BSH.
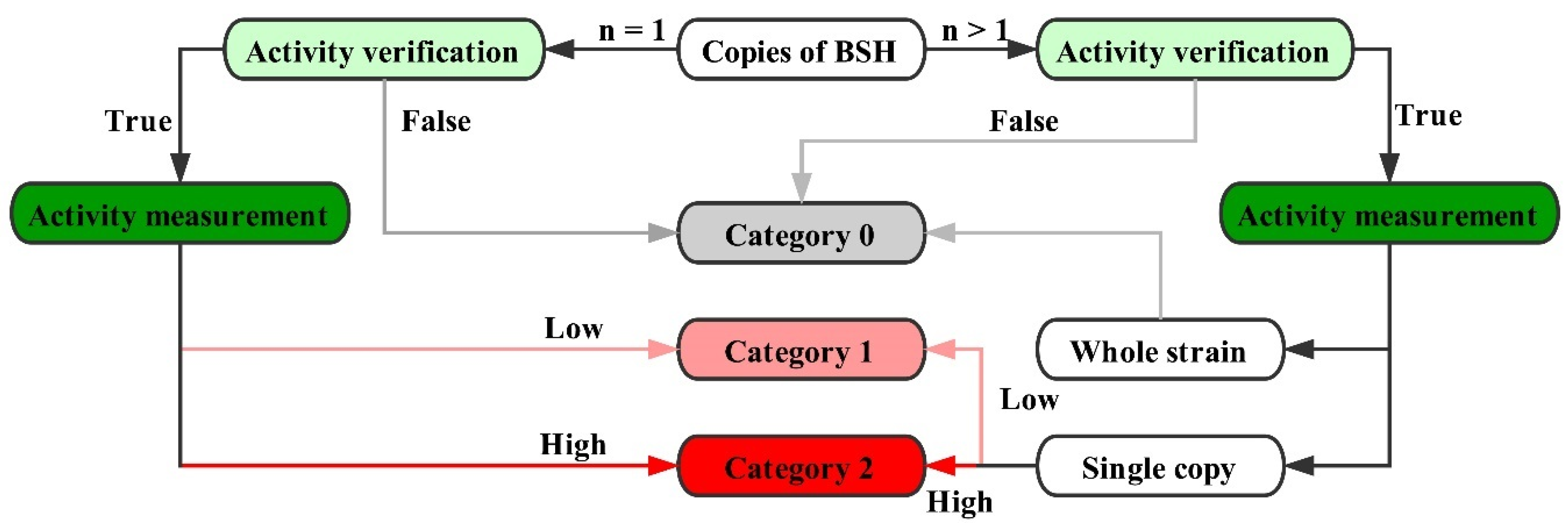
Figure 2.
Phylogenetic tree of BSHs in 34 Lactobacillaceae species. The BSH subtypes are shaded by various colors. BSH A, B, C, D and E are shown in red, green, blue, yellow and purple, respectively. On the right-hand side, the rectangle of the first column denotes experimental reports of this strain or bsh genes; blue means it has been reported before and grey means it has not. The second rectangle denotes the scoring type of this BSH (see more details in Table S1), categories 2, 1 and 0 are marked with red, light red and grey, respectively. The number beside it indicates the copy number of the bsh gene in this strain.
Figure 2.
Phylogenetic tree of BSHs in 34 Lactobacillaceae species. The BSH subtypes are shaded by various colors. BSH A, B, C, D and E are shown in red, green, blue, yellow and purple, respectively. On the right-hand side, the rectangle of the first column denotes experimental reports of this strain or bsh genes; blue means it has been reported before and grey means it has not. The second rectangle denotes the scoring type of this BSH (see more details in Table S1), categories 2, 1 and 0 are marked with red, light red and grey, respectively. The number beside it indicates the copy number of the bsh gene in this strain.
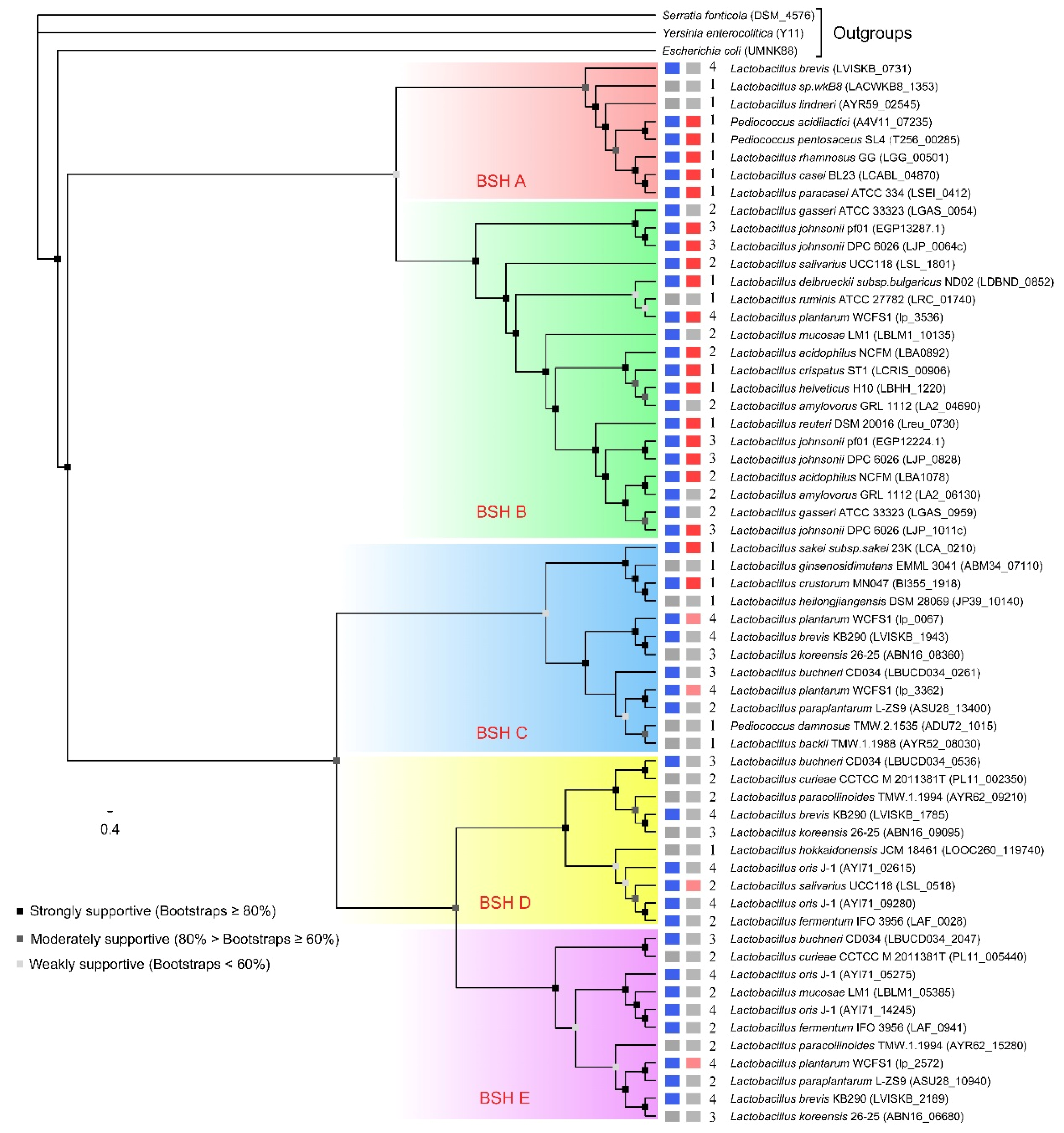
Figure 3.
Analysis of catalytic residues of BSH proteins. (a) Multiple-sequence alignment of five BSH subtypes in representative species. Six catalytic residues are marked by top arrow, and the site 82 is highlighted by the red box. The residues with sequence identity >50% and >80% were shaded by blue and yellow blocks, respectively; “!” and “*” were noted in the bottom side which residues identity =100% and >80%, respectively. (b) amino acid statistics of site 82 in six BSH subtypes from all BSH–SDL sequences (see more details of other sites in Table S2).
Figure 3.
Analysis of catalytic residues of BSH proteins. (a) Multiple-sequence alignment of five BSH subtypes in representative species. Six catalytic residues are marked by top arrow, and the site 82 is highlighted by the red box. The residues with sequence identity >50% and >80% were shaded by blue and yellow blocks, respectively; “!” and “*” were noted in the bottom side which residues identity =100% and >80%, respectively. (b) amino acid statistics of site 82 in six BSH subtypes from all BSH–SDL sequences (see more details of other sites in Table S2).
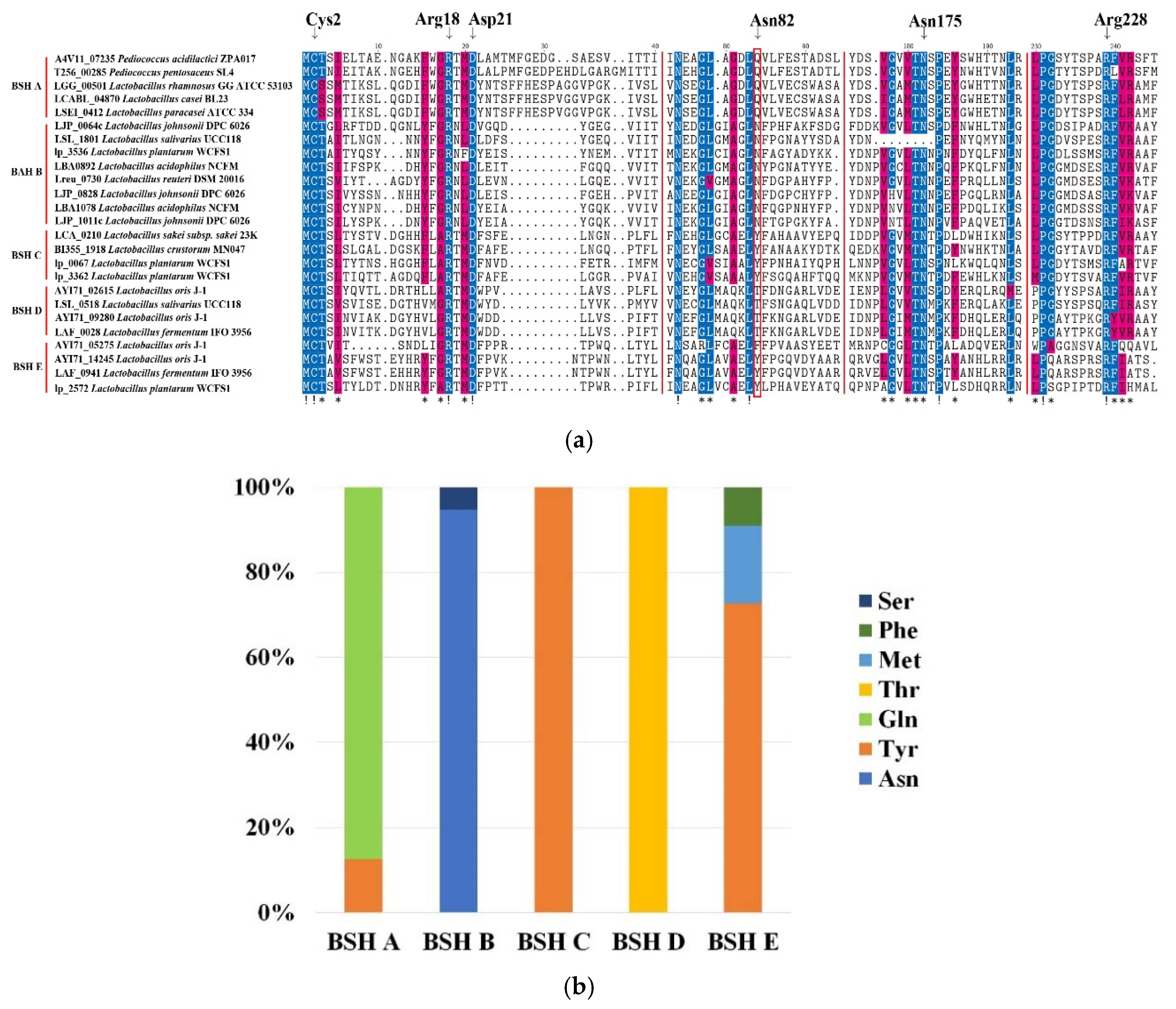
Figure 4.
Distribution of BSH subtypes in 107 Lactobacillaceae species. (a) Proportion of each BSH subtype in 107 species. 54 species carrying BSH A or BSH B subtypes accounted for 30% in total. Group 1 consists of 16 species which have been verified for BSH activity, and is included in BSH–SDL, and the other 38 species in group 2 (10 potential cholesterol-lowering bacteria) and group 3 (28 species without related investigation) are predicted to be BSH-active based on this study; (b) genomic location of bsh in Lactobacillus fermentum and Lactobacillus brevis. Those colors marked on BSH subtypes (a) and gene ID (b) indicate their corresponding colors on BSH subtypes in the phylogenetic tree of Figure 2.
Figure 4.
Distribution of BSH subtypes in 107 Lactobacillaceae species. (a) Proportion of each BSH subtype in 107 species. 54 species carrying BSH A or BSH B subtypes accounted for 30% in total. Group 1 consists of 16 species which have been verified for BSH activity, and is included in BSH–SDL, and the other 38 species in group 2 (10 potential cholesterol-lowering bacteria) and group 3 (28 species without related investigation) are predicted to be BSH-active based on this study; (b) genomic location of bsh in Lactobacillus fermentum and Lactobacillus brevis. Those colors marked on BSH subtypes (a) and gene ID (b) indicate their corresponding colors on BSH subtypes in the phylogenetic tree of Figure 2.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biological activity of representative predicted BSH-active species with diverse BSH subtypes.
Table 1.
Biological activity of representative predicted BSH-active species with diverse BSH subtypes.
| Species | BSH Subtypes | Tolerable Conditions | Substrate Specificities | Strains | |||||
|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | Bile Salt (%) | pH | |||
| Lactobacillus plantarum | 1 | 2 | 1 | 1.0 | 2.0 | GCA | ST-III [20] | ||
| 1 | 2 | 1 | - | - | GCA | WCFS1 [19] | |||
| 1 | 2 | 1 | 0.3 | 2.0 | - | LZ95 [26] | |||
| 1 | 1 | 1 | 1.0 | 2.0 | - | NCU116 [27] | |||
| Lactobacillus acidophilus | 2 | 0.2 | - | TCA, GCA, TCDCA | NCFM [22] | ||||
| 2 | 0.3 | 2.0 | - | DSM 20079 [28] | |||||
| Lactobacillus johnsonii | 2 | 0.5 | - | - | PF01 [29] | ||||
| 2 | 0.5 | 2.0 | GCDCA, TCDCA | La1 (also known as NCC533) [30] | |||||
| Lactobacillus rhamnosus | 1 | 2.0 | 2.5 | - | GG (ATCC 53103) [31] | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liang, L.; Yi, Y.; Lv, Y.; Qian, J.; Lei, X.; Zhang, G. A Comprehensive Genome Survey Provides Novel Insights into Bile Salt Hydrolase (BSH) in Lactobacillaceae. Molecules 2018, 23, 1157. https://doi.org/10.3390/molecules23051157
AMA Style
Liang L, Yi Y, Lv Y, Qian J, Lei X, Zhang G. A Comprehensive Genome Survey Provides Novel Insights into Bile Salt Hydrolase (BSH) in Lactobacillaceae. Molecules. 2018; 23(5):1157. https://doi.org/10.3390/molecules23051157
Chicago/Turabian StyleLiang, Lifeng, Yunhai Yi, Yunyun Lv, Junwei Qian, Xuejing Lei, and Gengyun Zhang. 2018. "A Comprehensive Genome Survey Provides Novel Insights into Bile Salt Hydrolase (BSH) in Lactobacillaceae" Molecules 23, no. 5: 1157. https://doi.org/10.3390/molecules23051157