Search for Partner Proteins of A. thaliana Immunophilins Involved in the Control of Plant Immunity

 ,
, 
Abstract
:1. Introduction
2. Results
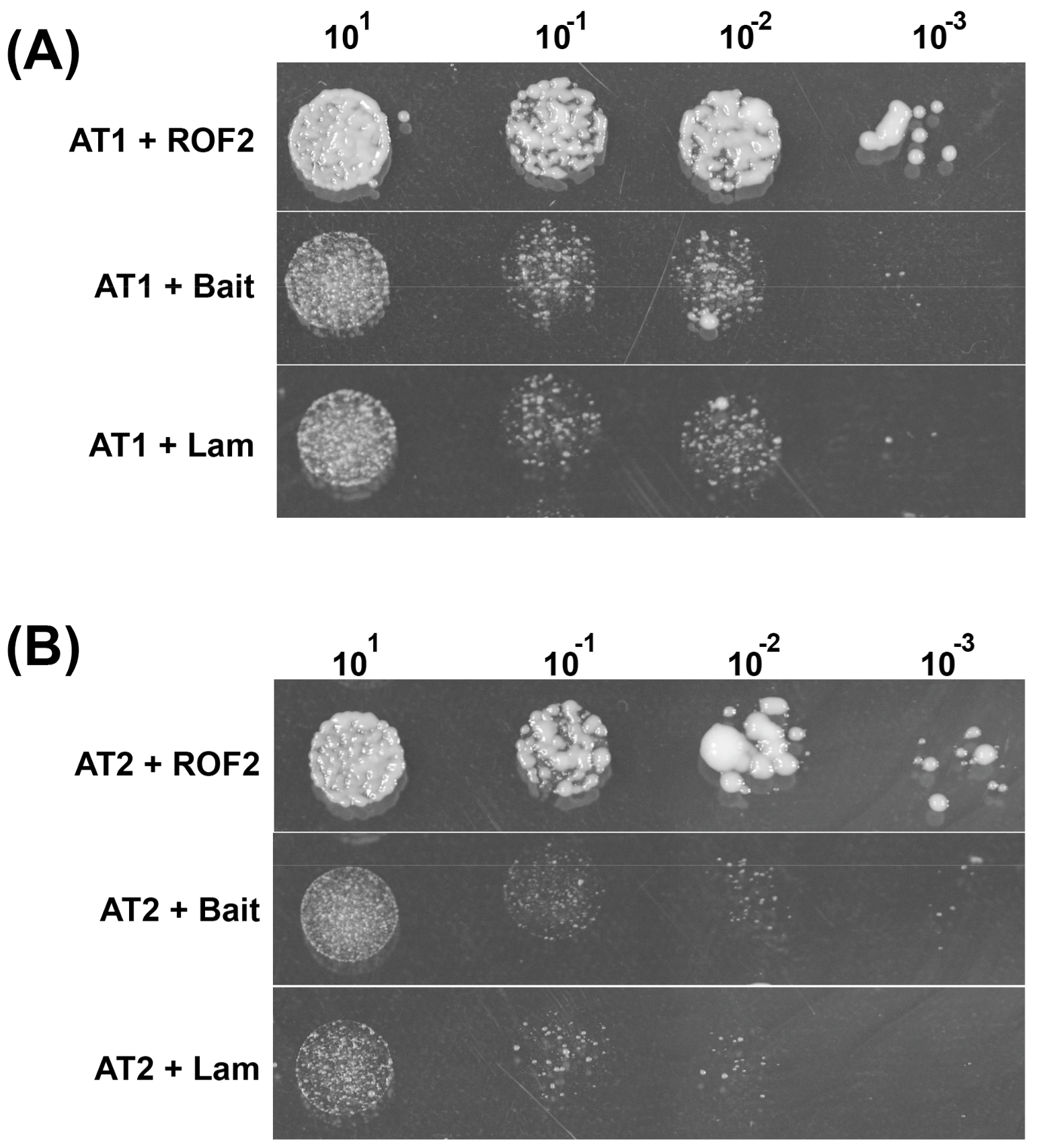
2.1. ROF2 Interaction Partners
2.2. Site-Directed Mutagenesis of Immunophilins
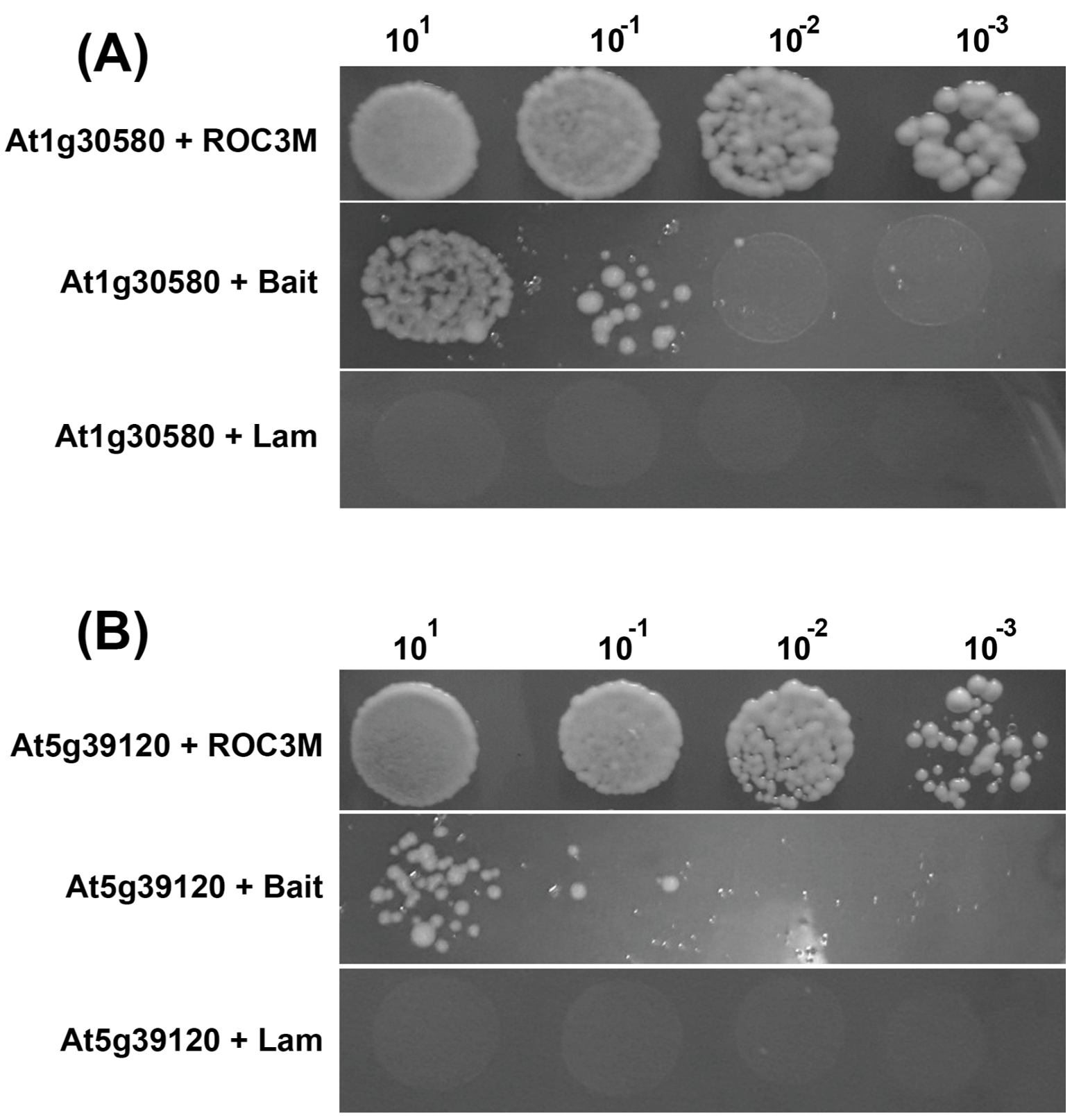
2.3. Interaction Partners for ROC3M
2.4. CYP57M Protein Partners
3. Discussion
4. Materials and Methods
4.1. Amplification and Cloning of Immunophilin cDNA Sequences and Identification of Interaction Partners
- ROF2 sequence (5′–3′): FKB-For (EcoRI) CATACgaattcGAAGACGATTTCGACACGCA and FKB-Rev (SalI) CATACgtcgacTCATGCCTTGGTGTCAATAC;
- ROC3 cDNA (5′–3′): ROC3-For (NdeI) CATACcatatgGCAACAAACCCTAAAGTCTA and ROC3-Rev (BamHI) CATACggatccgAACCTCCACCTGTACATGTG;
- AtCYP57 (5′–3′): CYP-For (EcoRI) CATACgaattcTCGACGGTGTACGTGCTAGA and CYP-Rev (PstI) CATACctgcagTCAGGCAAGAGATTTTCCAG.
4.2. Site-Directed Mutagenesis
4.3. Cotransformation of Yeast Cells
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pogorelko, G.V.; Mokryakova, M.V.; Fursova, O.V.; Abdeeva, I.A. Characterization of three Arabidopsis thaliana immunophilin genes involved in the plant defense response against Pseudomonas syringae. Gene 2014, 538, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Laloi, C.; Appel, K.; Danon, A. Reactive oxygen signalling: The latest news. Curr. Opin. Plant Biol. 2004, 7, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Fraire-Velázquez, S.; Rodríguez-Guerra, R.; Sánchez-Calderón, L. Abiotic and Biotic Stress Response Crosstalk-Plants. In Abiotic and Biotic Stress Response Crosstalk in Plants-Physiological, Biochemical and Genetic Perspectives; Shanker, A., Venkateswarlu, B., Eds.; InTech Open Access Company: Rijeka, Croatia, 2011; pp. 1–26. ISBN 978-953-307-672-0. [Google Scholar]
- Spoel, S.H.; Dong, X. Making sense of hormone crosstalk during plant immune response. Cell Host Microbe 2008, 3, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Ishikawa, A.; Jikumaru, Y.; Seki, M.; Umezawa, T.; Asami, T.; Maruyama-Nakashita, A.; Kudo, T.; Shinozaki, K.; Yoshida, S.; et al. Antagonistic interaction between systemic acquired resistance and the abscisic acid-mediated abiotic stress response in Arabidopsis. Plant Cell 2008, 20, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- Bartoli, C.G.; Casalongué, C.A.; Simontacchi, M.; Marquez-Garcia, B.; Foyer, C.H. Interactions between hormone and redox signaling pathways in the control of growth and cross-tolerance to stress. Environ. Exp. Bot. 2013, 94, 73–88. [Google Scholar] [CrossRef]
- Nurnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate immunity in plants and animals: Striking similarities and obvious differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling mechanisms in pattern-triggered Immunity (PTI). Mol. Plant. 2015, 8, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.Q.; Jiang, Z.H.; Yi, Y.Y.; Yang, Y.; Ke, L.P.; Pei, Z.M.; Zhu, S. Biotic and Abiotic Stresses Activate Different Ca2+ Permeable Channels in Arabidopsis. Front. Plant Sci. 2017, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhu, S.; Ye, R.; Xue, Y.; Chen, A.; An, L.; Pei, Z.M. Relationship between NaCl- and H2O2-induced cytosolic Ca2+ increases in response to stress in rabidopsis. PLoS ONE 2013, 8, e76130. [Google Scholar] [CrossRef]
- Espinosa-Diez, C.; Miguel, V.; Mennerich, D.; Kietzmann, T.; Sánchez-Pérez, P.; Cadenas, S.; Lamas, S. Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol. 2015, 6, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.A. ROS in biotic interactions. Physiol. Plant 2010, 138, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G. Cell signaling. H2O2, a necessary evil for cell signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef] [PubMed]
- Asai, S.; Ohta, K.; Yoshioka, H. MAPK signaling regulates nitric oxide and NADPH oxidase-dependent oxidative bursts in Nicotiana benthamiana. Plant Cell 2008, 20, 1390–1406. [Google Scholar] [CrossRef] [PubMed]
- Aumüller, T.; Jahreis, G.; Fischer, G.; Schiene-Fischer, C. Role of prolyl cis/trans isomers in cyclophilin-assisted Pseudomonas syringae AvrRpt2 protease activation. Biochemistry 2010, 49, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Tzelepis, G.; Zouhar, M.; Ryšánek, P.; Dixelius, C. The immunophilin repertoire of lasmodiophora brassicae and functional analysis of PbCYP3 cyclophilin. Mol. Genet. Genom. 2018, 293, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Iterative Threading ASSEmbly Refinement. Available online: https://zhanglab.ccmb.med.umich.edu/I-TASSER/ (accessed on 18 April 2018).
- Rodriguez, L.; Gonzalez-Guzman, M.; Diaz, M.; Rodrigues, A.; Izquierdo-Garcia, A.C.; Peirats-Llobet, M.; Fernandez, M.A.; Antoni, R.; Fernandez, D.; Marquez, J.A.; et al. C2-domain abscisic acid-related proteins mediate the interaction of PYR/PYL/RCAR abscisic acid receptors with the plasma membrane and regulate abscisic acid sensitivity in Arabidopsis. Plant Cell 2014, 26, 4802–4820. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jiang, L.; Wang, G.; Yu, L.; Zhang, Q.; Xin, Q.; Wu, W.; Gong, Z.; Chen, Z. Structural insights into the abscisic acid stereospecificity by the ABA receptors PYR/PYL/RCAR. PLoS ONE 2013, 8, e67477. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Yin, P.; Li, W.; Wang, L.; Yan, C.; Lin, Z.; Wu, J.Z.; Wang, J.; Yan, S.F.; Yan, N. The molecular basis of ABA-independent inhibition of PP2Cs by a subclass of PYL proteins. Mol. Cell 2011, 42, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Asselbergh, B.; Achuo, A.E.; Höfte, M.; Van Gijsegem, F. Abscisic acid deficiency leads to rapid activation of tomato defence responses upon infection with Erwinia chrysanthemi. Mol. Plant Pathol. 2008, 9, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, N.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3544. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Ton, J.; Ent, V.D.S.; Hulten, V.M.; Pozo, M.; Oosten, V.V.; Loon, L.C.; Mauch-Mani, B.; Turlings, T.C.J.; Pieterse, C.M.J. Priming as a mechanism behind induced resistance against pathogens; insects and abiotic stress. IOBC/Wprs Bull. 2009, 44, 3–13. [Google Scholar]
- Melotto, M.; Underwood, W.; Koczan, J.; Nomura, K.; He, S.Y. Plant stomata function in innate immunity against bacterial invasion. Cell 2006, 126, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Luna, E.; Pastor, V.; Robert, J.; Flors, V.; Mauch-Mani, B.; Ton, J. Callose deposition: A multifaceted plant defense response. Mol. Plant Microbe Interact. 2011, 24, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.F.; Talle, B.; Wilson, Z.A. Anther and pollen development: A conserved developmental pathway. J. Integr. Plant Biol. 2015, 57, 876–891. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.P.; Badruzsaufari, E.; Schenk, P.M.; Manners, J.M.; Desmond, O.J.; Ehlert, C.; Maclean, D.J.; Ebert, P.R.; Kazan, K. Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell 2004, 16, 3460–3479. [Google Scholar] [CrossRef] [PubMed]
- Schweighofer, A.; Kazanaviciute, V.; Scheikl, E.; Teige, M.; Doczi, R.; Hirt, H.; Schwanninger, M.; Kant, M.; Schuurink, R.; Mauch, F.; et al. The PP2C-Type Phosphatase AP2C1, Which Negatively Regulates MPK4 and MPK6, Modulates Innate Immunity, Jasmonic Acid, and Ethylene Levels in Arabidopsis. Plant Cell 2007, 19, 2213–2224. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, Y.J.; Kim, M.H.; Kwak, J.M. MAPK Cascades in Guard Cell Signal Transduction. Front. Plant Sci. 2016, 7, 80. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Rasool, S.; Gul, A.; Sheikh, S.A.; Akram, N.A.; Ashraf, M.; Kazi, A.M.; Gucel, S. Jasmonates: Multifunctional Roles in Stress Tolerance. Front. Plant Sci. 2016, 7, 813. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.Y.; Li, M.W.; Yung, Y.L.; Wen, C.Q.; Lam, H.M. The unconventional P-loop NTPase OsYchF1 and its regulator OsGAP1 play opposite roles in salinity stress tolerance. Plant Cell Environ. 2013, 36, 2008–2020. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rubio, V.; Lieberman, M.W.; Shi, Z.Z. OLA1, an Obg-like ATPase, suppresses antioxidant response via nontranscriptional mechanisms. Proc. Natl. Acad. Sci. USA-Biol. Sci. 2009, 106, 15356–15361. [Google Scholar] [CrossRef] [PubMed]
- Giraud, M.F.; Leonard, G.A.; Field, R.A.; Berlind, C.; Naismith, J.H. RmlC, the third enzyme of dTDP-L-rhamnose pathway, is a new class of epimerase. Nat. Struct. Biol. 2000, 7, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liao, B.; Qi, H.; Xie, L.J.; Huang, L.; Tan, W.J.; Zhai, N.; Yuan, L.B.; Zhou, Y.; Yu, L.J.; et al. Autophagy contributes to regulation of the hypoxia response during submergence in Arabidopsis thaliana. Autophagy 2015, 11–12, 2233–2246. [Google Scholar] [CrossRef] [PubMed]
- Tadege, M.; Bucher, M.; Stahli, W.; Suter, M.; Dupuis, I.; Kuhlemeier, C. Activation of plant defense responses and sugar efflux by expression of pyruvate decarboxylase in potato leaves. Plant J. 1998, 16, 661–671. [Google Scholar] [CrossRef]
- Xiang, L.; Etxeberria, E.; Van den Ende, W. Vacuolar protein sorting mechanisms in plants. FEBS J. 2013, 280, 979–993. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.; Pan, S.; Zouhar, J.; Avila, E.L.; Girke, T.; Raikhel, N.V. The Vegetative Vacuole Proteome of Arabidopsis thaliana Reveals Predicted and Unexpected Proteins. Plant Cell 2004, 16, 3285–3303. [Google Scholar] [CrossRef] [PubMed]
- Kursteiner, O.; Dupuis, I.; Kuhlemeier, C. The Pyruvate decarboxylase1 Gene of Arabidopsis Is Required during Anoxia but Not OtherEnvironmental Stresses. Plant Physiol. 2003, 132, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Song, Y.; Zhuang, K.; Li, L.; Xia, Y.; Shen, Z. Proteomic Analysis of Copper-Binding Proteins in Excess Copper-Stressed Roots of Two Rice (Oryza sativa L.) Varieties with Different Cu Tolerances. PLoS ONE 2015, 10, e0125367. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Number of Clones | Partner Protein | Cotransformation ID | Confirmation |
|---|---|---|---|
| 2 | Calcium-dependent lipid-binding domain-containing protein | At1g70790 (АТ1) | + |
| 2 | Putative protein phosphatase | At2g30020 (АТ2) | + |
| 3 | ATPase, F1 complex, OSCP/delta subunit | At4g00895 (АТ4) | − |
| 1 | 60S ribosomal protein L27a-3 mRNA | False positive | N/A |
| 1 | 40S ribosomal protein S20 | False positive | N/A |
| 1 | H2A_12 | False positive | N/A |
| Number of Clones | Partner Protein | Cotransformation ID | Confirmation |
|---|---|---|---|
| 2 | ENGD-1, GTP-binding protein | At1g30580 | + |
| 1 | RmlC-like cupin superfamily | At5g39120 | + |
| 1 | At1g72370 40S ribosomal protein Sa-1 | False positive | N/A |
| 1 | At2g36830 GAMMA-TIP, TIP1 | False positive | N/A |
| 1 | At1g12050 putative fumarylacetoacetase | False positive | N/A |
| 1 | H2A_12 | False positive | N/A |
| Number of Clones | Partner Protein | Cotransformation ID | Confirmation |
|---|---|---|---|
| 1 | Putative pyruvate decarboxylase-1 Pdc1 | At4g33070 | + |
| 1 | Сlathrin adaptor complex medium subunit family protein At5g05010 | At5g05010 | + |
| 1 | At4g24280 chloroplast heat shock protein 70-1 | False positive | N/A |
| 1 | At1g70800 ENHANCED BENDING 1 | False positive | N/A |
| 1 | At3g53430 60S ribosomal Protein L12 | False positive | N/A |
| 1 | At4g03520 thioredoxin M2 | False positive | N/A |
| 1 | At1g70850 MLP-like protein 34 | False positive | N/A |
| 1 | At2g36830 GAMMA-TIP, TIP1 | False positive | N/A |
| Gene ID | Forward Primer, 5′–3′ | Reverse Primer, 5′–3′ |
|---|---|---|
| ТОР1/At5g | TOP-For (NdeI) NNcatatgTTAATGGCGACTCCAACG | TOP-Rev (BamHI) NNggatccTTAAGCAGAAGCAGAGGCAGC |
| АТ1/At1g70790 | AT1-For (NdeI) NNcatatgGAAGATAAACCATTAGGGAT | AT1-Rev (XhoI) NNctcgagTTAGTCCAATCGTTTTGTCGGCA |
| AT2/At2g300020 | AT2-For (NdeI) NNcatatgTCTTGCTCCGTCGCCGTA | AT2-Rev (BamHI) NNggatccCTATATGAACTGGCGTAAAGG |
| At4/At4g00895 | AT4-For (NdeI) NNcatatgGATACTCTCTCAGCATCC | AT4-Rev (BamHI) NNggatccTCATCAAGAAACCCAGACAAG |
| At1g30580 | At1g30580-For(EcoRI) gaatccATGCCTCCGAAAGCCAAAG | At1g30580-Rev(XhoI) ctcgagTCATTTCTTCCCACCACCGGA |
| At4g33070 | At4g33070-For(EcoRI) gaattcATGGACACCAAAATCGGATC | At4g33070-Rev(XhoI) ctcgagCTACTGAGGATTGGGAGGAC |
| At5g05010 | At5g05010- For(EcoRI) gaattcATGGTTGTGCTTGCTGCTG | At5g05010-Rev(BamHI) ggatccTCATATGACTTGATAGTTCTGG |
| At5g39120 | At5g39120-For(EcoRI) gaattcATGAAGGTGTCCATGTCTC | At5g39120-Rev(XhoI) ctcgagTTAGTTTTTAAACTTGGCCTC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdeeva, I.A.; Pogorelko, G.V.; Maloshenok, L.G.; Mokrykova, M.V.; Fursova, O.V.; Bruskin, S.A. Search for Partner Proteins of A. thaliana Immunophilins Involved in the Control of Plant Immunity. Molecules 2018, 23, 953. https://doi.org/10.3390/molecules23040953
Abdeeva IA, Pogorelko GV, Maloshenok LG, Mokrykova MV, Fursova OV, Bruskin SA. Search for Partner Proteins of A. thaliana Immunophilins Involved in the Control of Plant Immunity. Molecules. 2018; 23(4):953. https://doi.org/10.3390/molecules23040953
Chicago/Turabian StyleAbdeeva, Inna A., Gennady V. Pogorelko, Liliya G. Maloshenok, Maria V. Mokrykova, Oksana V. Fursova, and Sergey A. Bruskin. 2018. "Search for Partner Proteins of A. thaliana Immunophilins Involved in the Control of Plant Immunity" Molecules 23, no. 4: 953. https://doi.org/10.3390/molecules23040953
APA StyleAbdeeva, I. A., Pogorelko, G. V., Maloshenok, L. G., Mokrykova, M. V., Fursova, O. V., & Bruskin, S. A. (2018). Search for Partner Proteins of A. thaliana Immunophilins Involved in the Control of Plant Immunity. Molecules, 23(4), 953. https://doi.org/10.3390/molecules23040953