Role of Phenolic Acids from the Rhizosphere Soils of Panax notoginseng as a Double-Edge Sword in the Occurrence of Root-Rot Disease

Abstract
:1. Introduction
2. Results
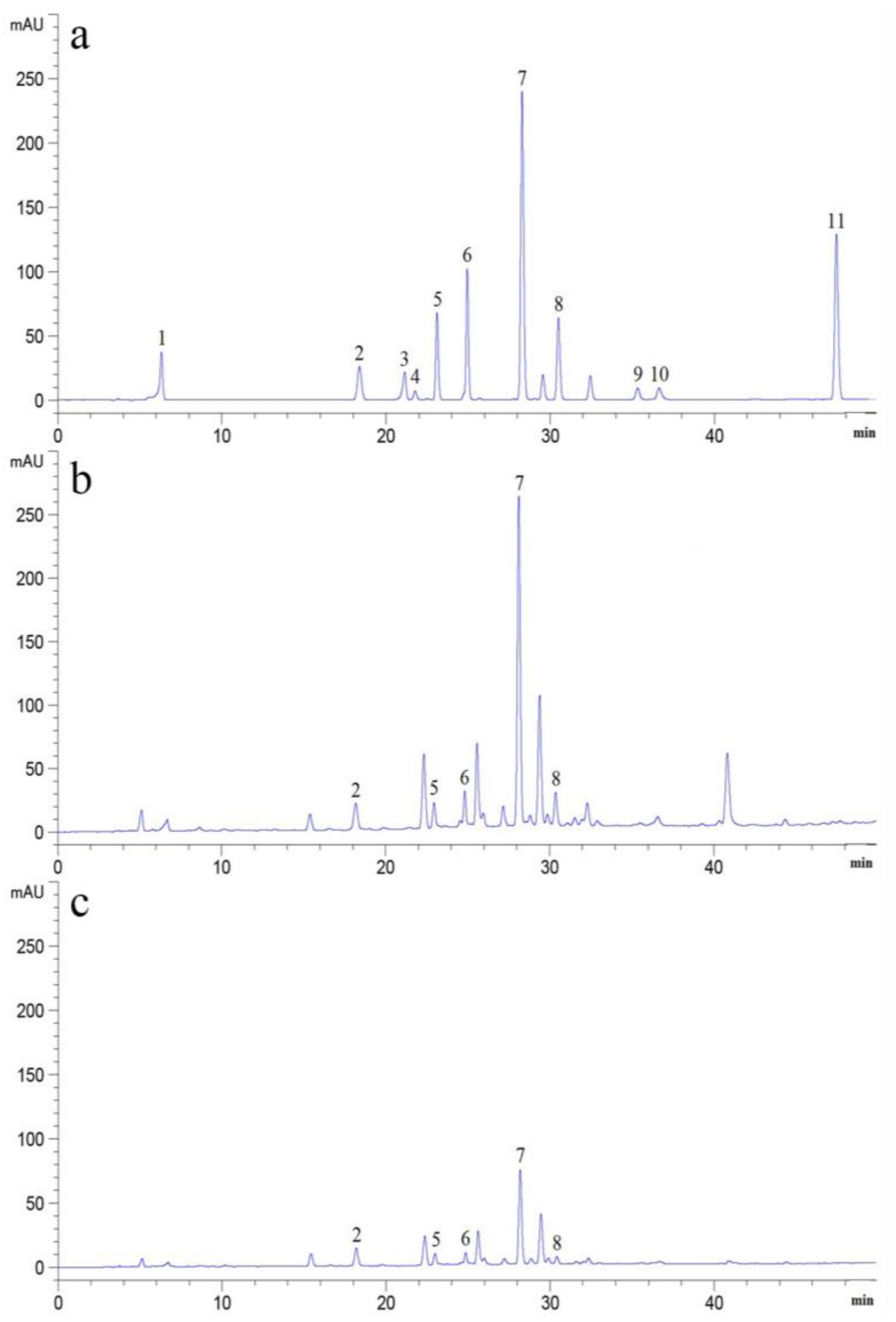
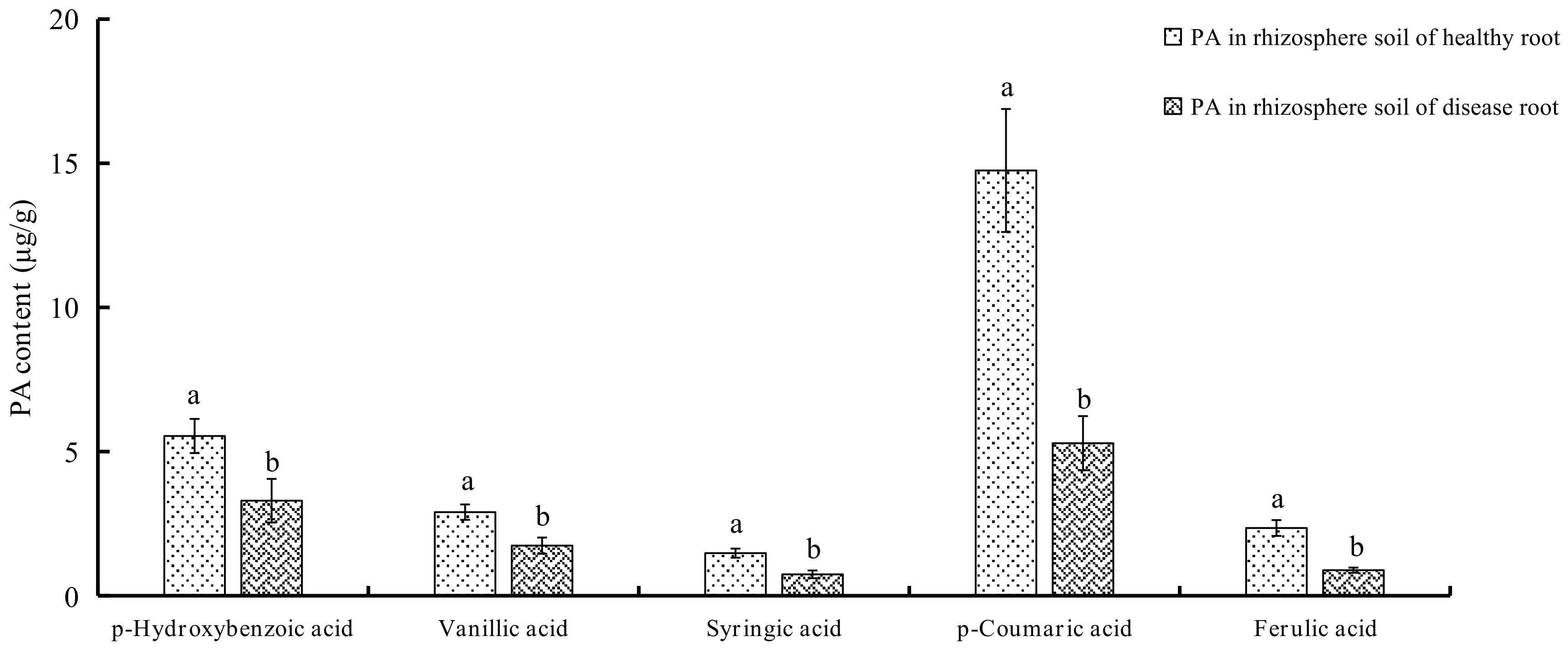
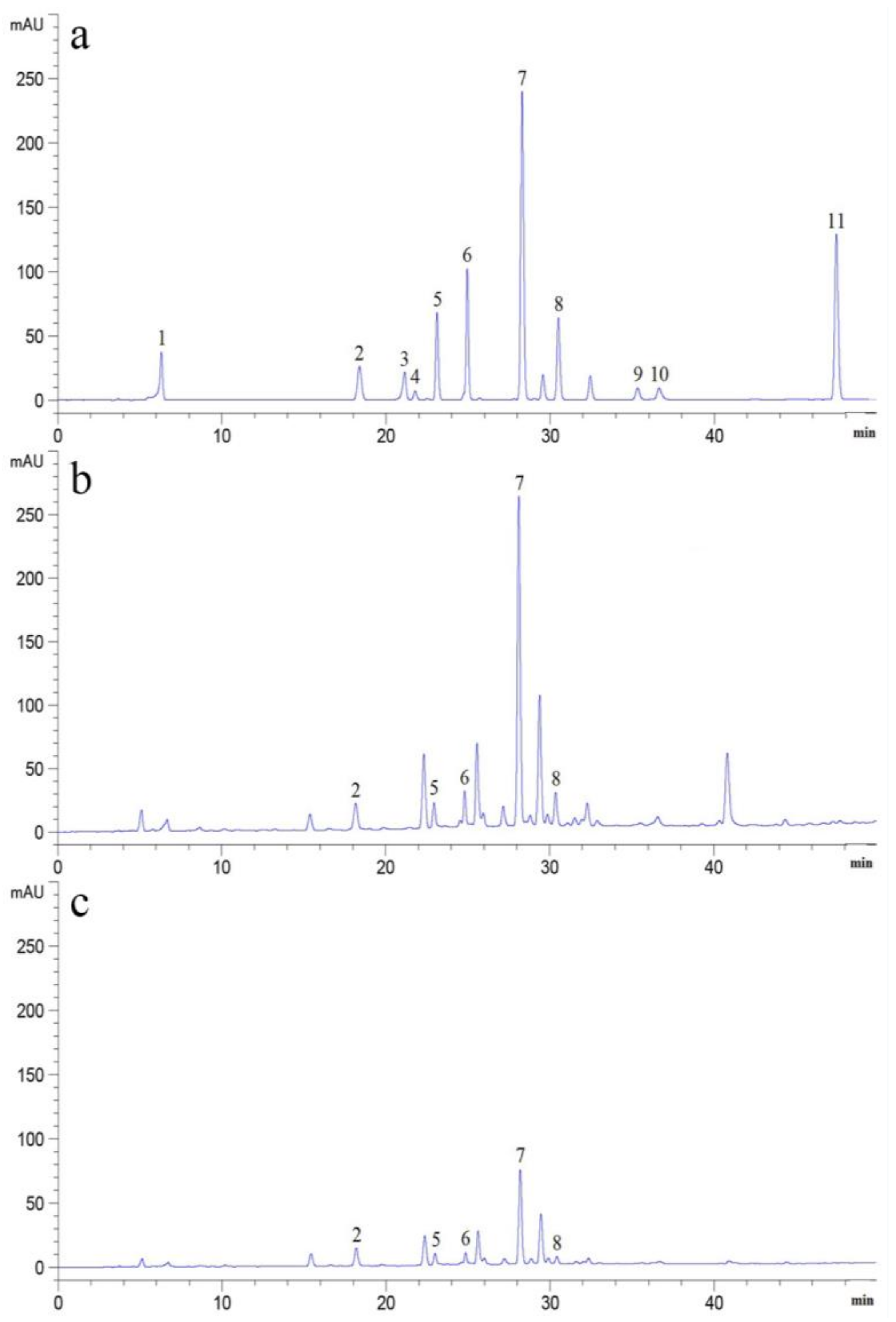
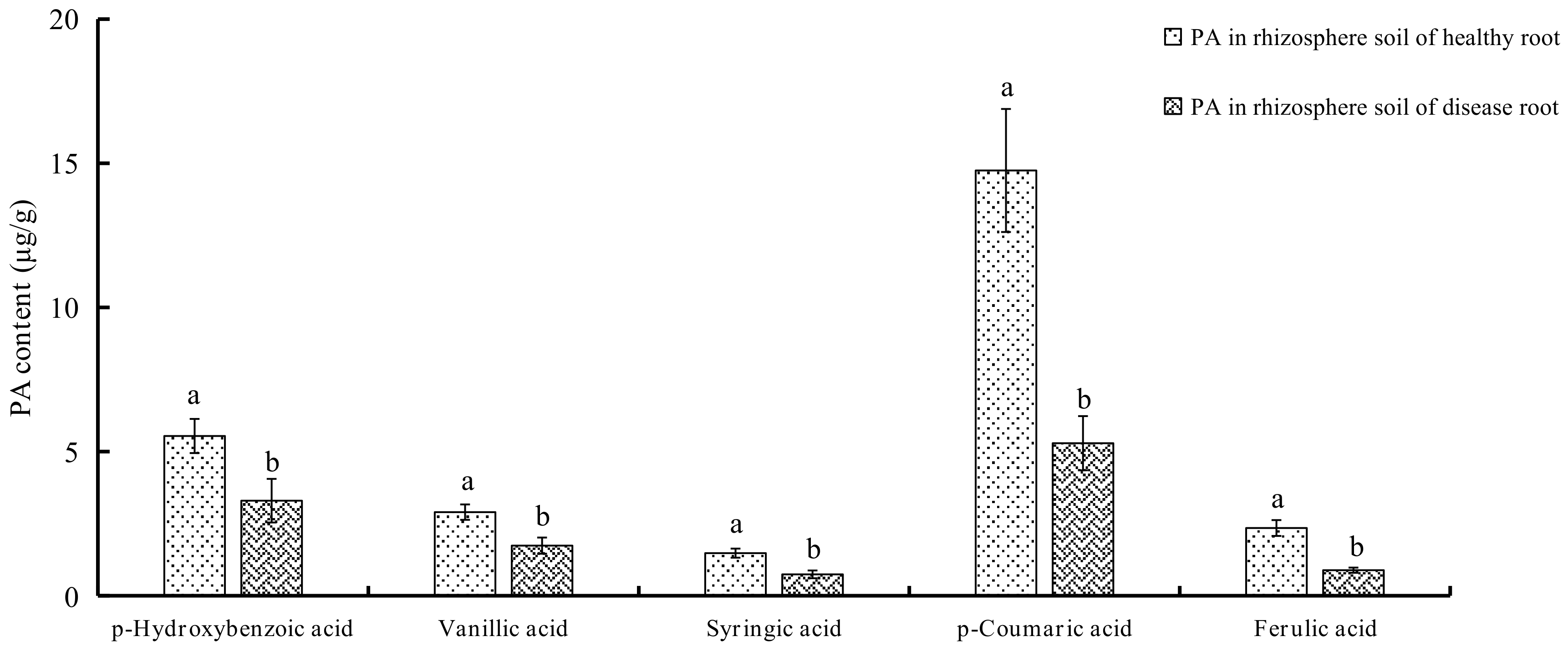
2.1. Determination of PAs in Soils
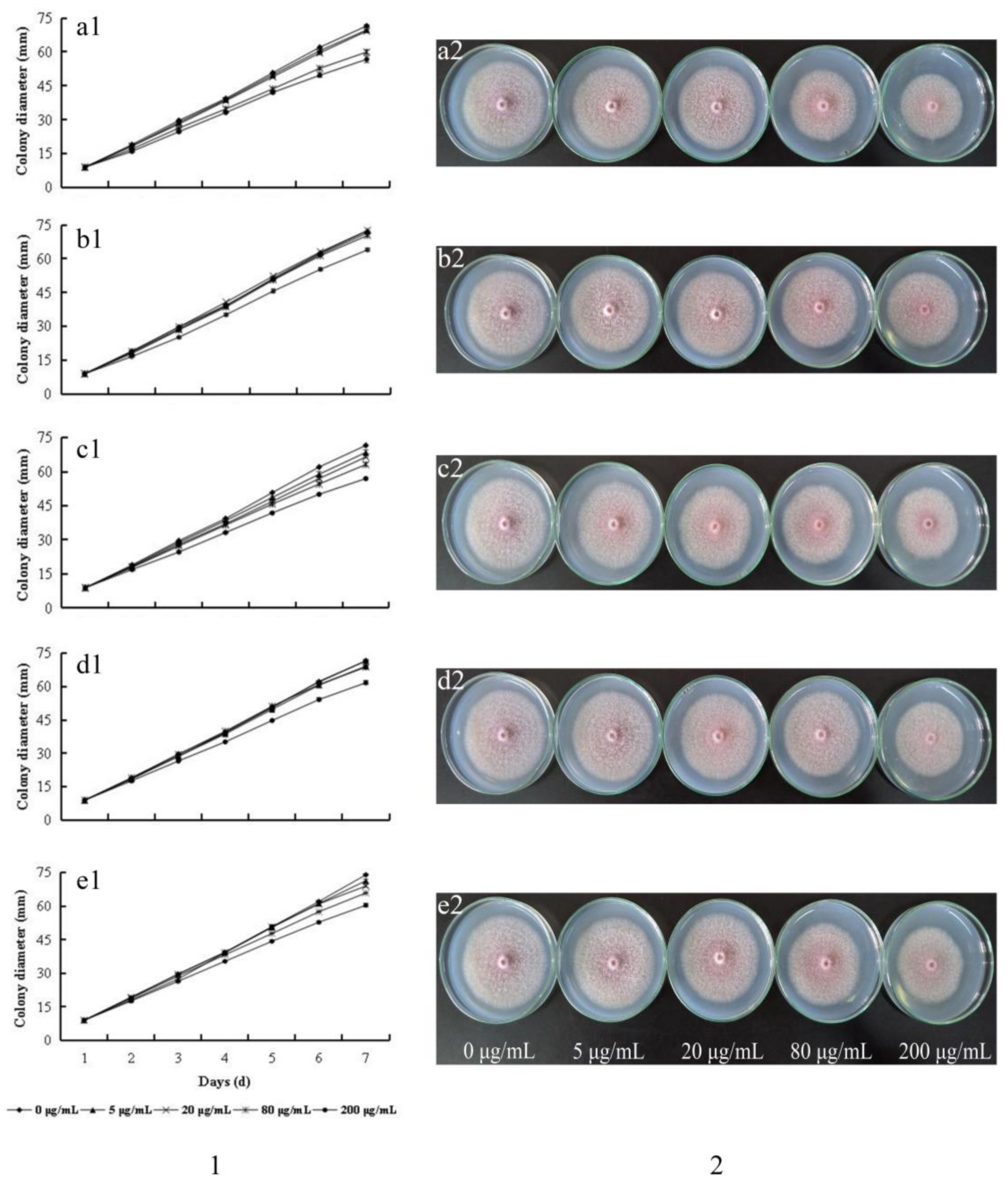
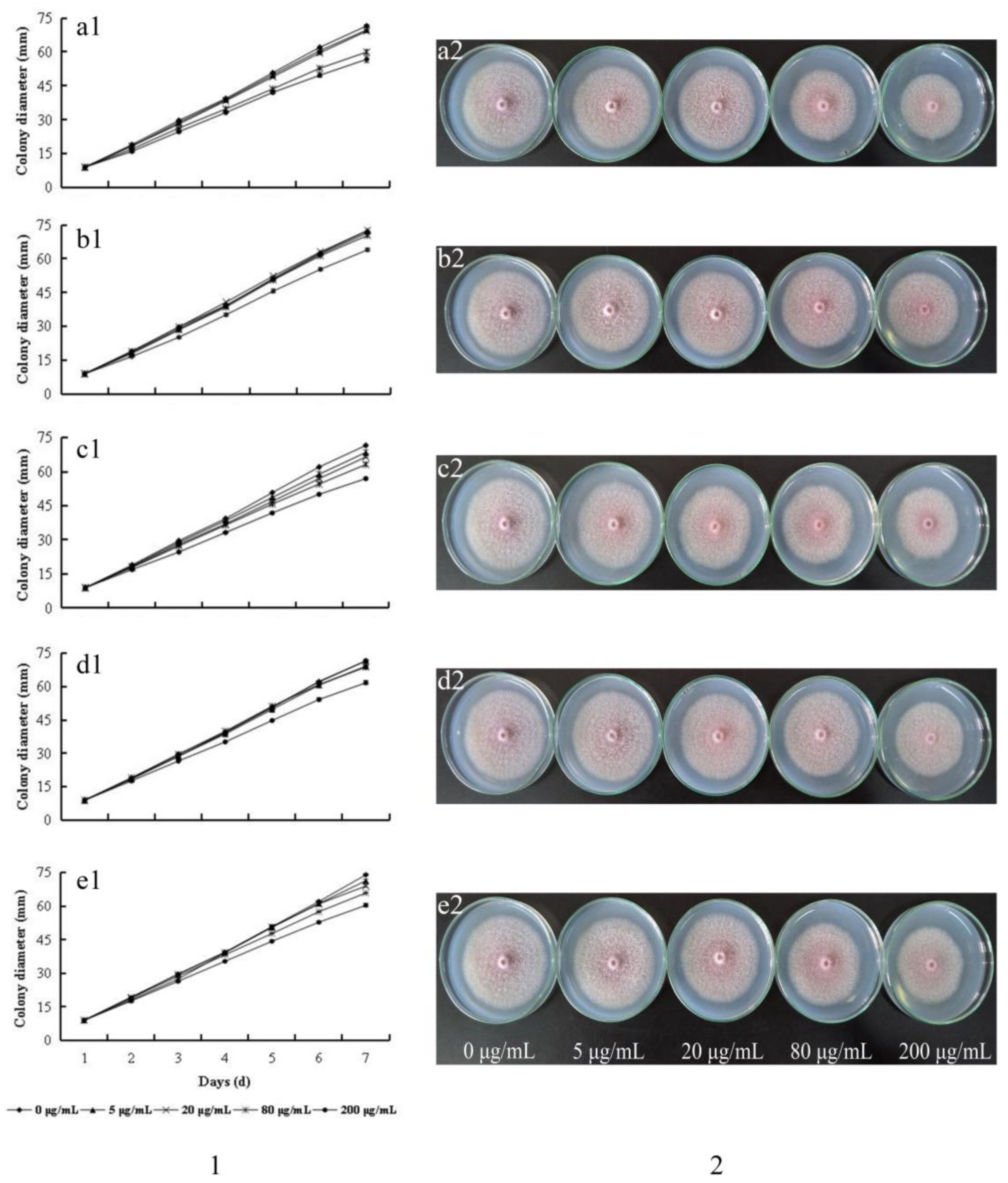
2.2. Growth Inhibition of F. oxysporum
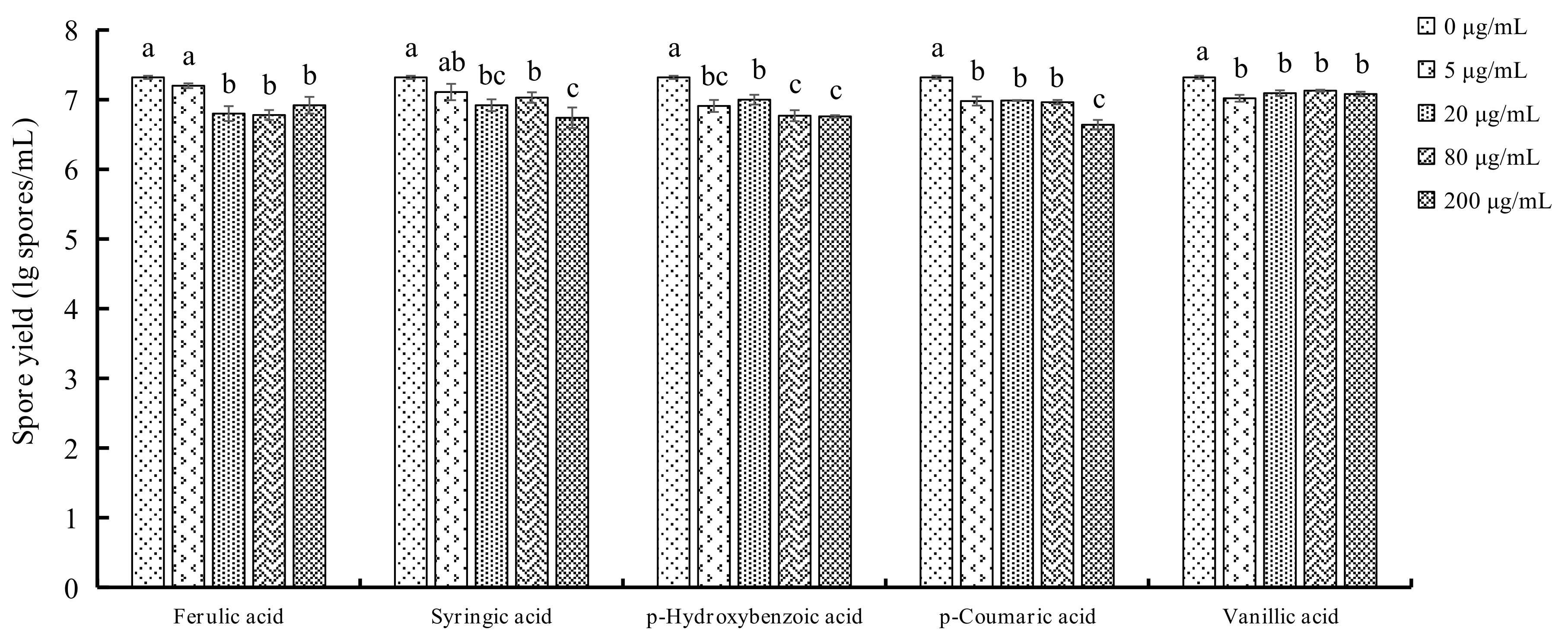
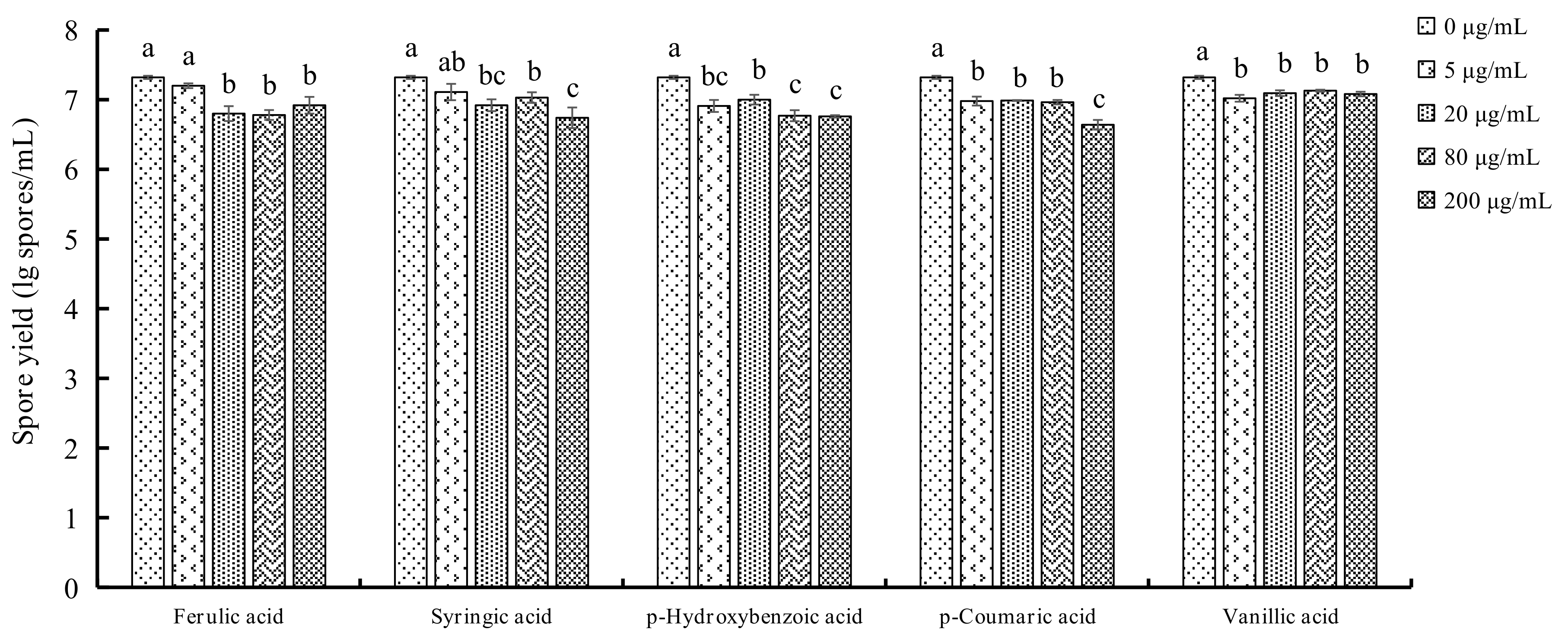
2.3. Determination of Spore Yields of F. oxysporum
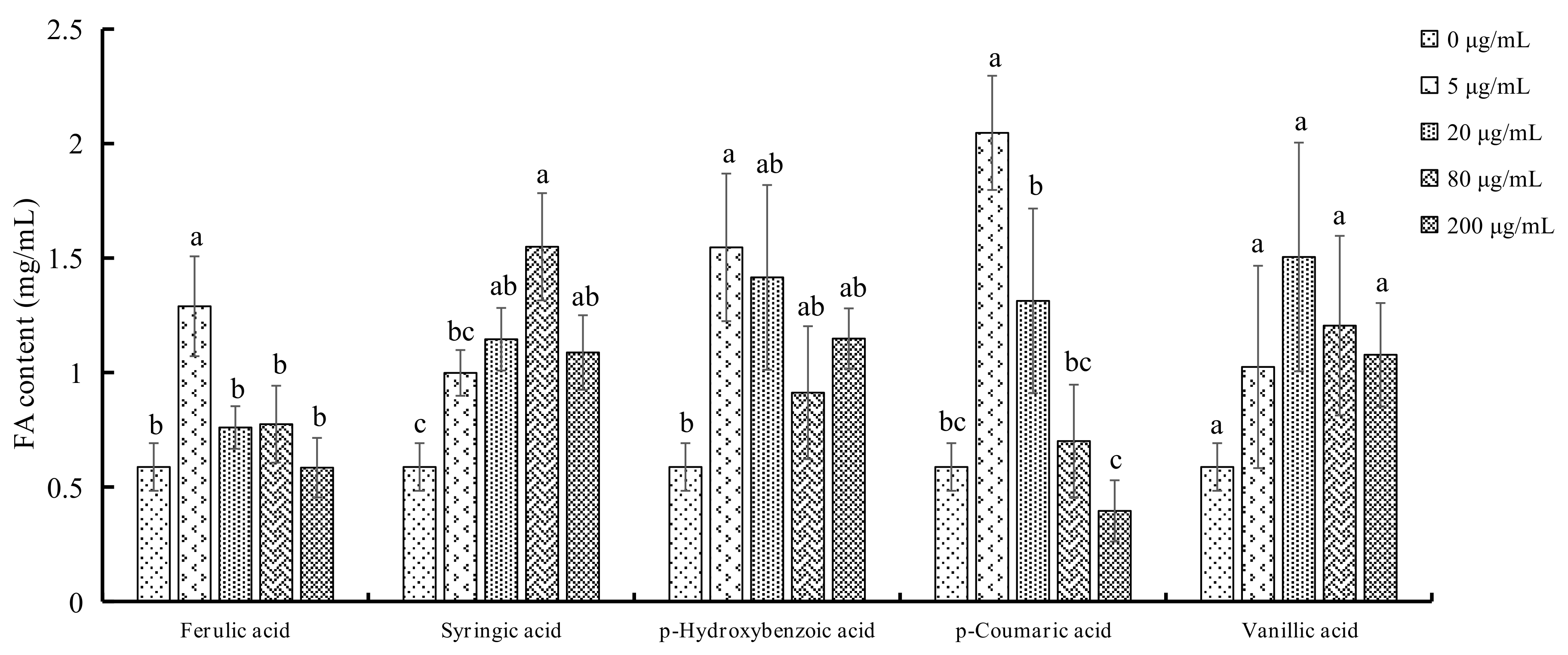
2.4. FA Production in Both F. oxysporum and P. notoginseng
2.5. Effects of FA on the Growth of P. notoginseng
3. Materials and Methods
3.1. Soil Materials
3.2. Indicator Fungi
3.3. Determination of PAs in Soil Samples
3.4. Determination of F. oxysporum Growth
3.5. Determination of the Spore Yields of F. oxysporum
3.6. Determination of FA in Both F. oxysporum and P. notoginseng
3.7. The Effect of FA on the Growth of P. notoginseng
3.8. Statistical Analysis
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guo, H.B.; Cui, X.M.; An, N.; Cai, G.P. Sanchi ginseng (Panax notoginseng (Burkill) F.H. Chen) in China: Distribution, cultivation and variations. Genet. Resour. Crop Evolut. 2010, 57, 453–460. [Google Scholar] [CrossRef]
- Miao, Z.Q.; Li, S.D.; Liu, X.Z.; Chen, Y.J.; Li, Y.H.; Wang, Y.; Guo, R.J.; Xia, Z.Y.; Zheng, K.Q. The causal microorganisms of Panax notoginseng root rot disease. Sci. Agric. Sin. 2006, 39, 1371–1378. [Google Scholar] [CrossRef]
- He, C.N.; Bi, W.; Jiao, X.L.; Gao, W.W. Variation of ginsenosides in the infected roots of American ginseng (Panax quinquefolium). China J. Chin. Mater. Med. 2008, 33, 2905. [Google Scholar] [CrossRef]
- Paulitz, T.C. Low Input No-till Cereal Production in the Pacific Northwest of the U.S.: The challenges of root diseases. Eur. J. Plant Pathol. 2006, 115, 271–281. [Google Scholar] [CrossRef]
- Gapillout, I.; Milat, M.L.; Blein, J.P. Effects of fusaric acid on cells from tomato cultivars resistant or susceptible to Fusarium oxysporum f.sp. Lycopersici. Eur. J. Plant Pathol. 1996, 102, 127–132. [Google Scholar] [CrossRef]
- Toyoda, H.; Hashimoto, H.; Utsumi, R.; Kobayashi, H.; Ouchi, S. Detoxification of fusaric acid by a fusaric acid-resistant mutant of Pseudomonas solanacearum and its application to biological control of Fusarium wilt of tomato. Phytopathology 1988, 78, 1307–1311. [Google Scholar] [CrossRef]
- Mao, Z.S.; Long, Y.J.; Zhu, S.S.; Chen, Z.J.; Wei, F.G.; Zhu, Y.Y.; He, X.H. Progress in research on Panax notoginseng root rot disease. J. Chin. Med. Mater. 2013, 36, 2051–2054. [Google Scholar] [CrossRef]
- Zanardo, D.I.L.; Lima, R.B.; Ferrarese, M.D.L.L.; Bubna, G.A.; Ferrarese-filho, O. Soybean root growth inhibition and lignification induced by p-coumaric acid. Environ. Exp. Bot. 2009, 66, 25–30. [Google Scholar] [CrossRef]
- Yan, J.; Bi, H.H.; Liu, Y.Z.; Zhang, M.; Zhou, Z.Y.; Tan, J.W. Phenolic Compounds from Merremia umbellata subsp. Orientalis and their allelopathic effects on arabidopsis seed germination. Molecules 2010, 15, 8241–8250. [Google Scholar] [CrossRef] [PubMed]
- Diley, D.; Barber, S.A. Salt Accumulation at the Soybean (Glycine Max. (L.) Merr.) Root-Soil Interface. Soil Sci. Soc. Am. J. 1970, 34, 154–155. [Google Scholar] [CrossRef]
- He, C.N.; Gao, W.W.; Yang, J.X.; Bi, W.; Zhang, X.S.; Zhao, Y.J. Identification of autotoxic compounds from fibrous roots of Panax quinquefolium L. Plant Soil 2009, 318, 63–72. [Google Scholar] [CrossRef]
- Wu, H.M.; Wu, L.K.; Wang, J.Y.; Zhu, Q.; Lin, S.; Xu, J.H.; Zheng, C.L.; Chen, J.; Qin, X.J.; Fang, C.X.; et al. Mixed phenolic acids mediated proliferation of pathogens Talaromyces helicus and Kosakonia sacchari in continuously monocultured Radix pseudostellariae rhizosphere soil. Front. Microbiol. 2016, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Ling, N.; Raza, W.; Ma, J.H.; Huang, Q.W.; Shen, Q.R. Identification and role of organic acids in watermelon root exudates for recruiting Paenibacillus polymyxa SQR-21 in the rhizosphere. Eur. J. Soil Biol. 2011, 47, 374–379. [Google Scholar] [CrossRef]
- Wu, H.S.; Raza, W.; Fan, J.Q.; Sun, Y.G.; Bao, W.; Shen, Q.R. Cinnamic acid inhibits growth but stimulates production of pathogenesis factors by in vitro cultures of Fusarium oxysporum f.sp. niveum. J. Agric. Food Chem. 2008, 56, 1316–1321. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.C.; Flematti, G.R.; Ghisalberti, E.L.; Sivasithamparam, K.; Chakraborty, S.; Obanor, F.; Barbetti, M.J. Mycotoxins produced by Fusarium species associated with annual legume pastures and ‘sheep feed refusal disorders’ in Western Australia. Mycotoxin Res. 2011, 27, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.K.; Sousadias, M.G. Fusaric acid content of swine feed stuffs. J. Agric. Food Chem. 1993, 41, 2296–2298. [Google Scholar] [CrossRef]
- Dong, X.; Ling, N.; Wang, M.; Shen, Q.R.; Guo, S.W. Fusaric acid is a crucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Plant Phys. Biochem. 2012, 60, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, V.; Kanoujia, N. Biological management of vascular wilt of tomato caused by Fusarium oxysporum f.sp. Lycospersici by plant growth-promoting rhizobacterial mixture. Biol. Control 2001, 57, 85–93. [Google Scholar] [CrossRef]
- Shaharoona, B.; Naveed, M.; Arshad, M.; Zahir, Z.A. Fertilizer-dependent efficiency of Pseudomonads for improving growth, yield, and nutrient use efficiency of wheat (Triticum aestivum L.). Appl. Microbiol. Biotechnol. 2008, 79, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Matern, U.; Grimmig, B.; Kneusel, R.E. Plant cell wall reinforcement in the disease-resistance response: Molecular composition and regulation. Can. J. Bot. 1995, 73, 511–517. [Google Scholar] [CrossRef]
- De Ascensao, A.R.; Dubery, I.A. Soluble and wall-bound phenolics and phenolic polymers in Musa acuminata roots exposed to elicitors from Fusarium oxysporum f.sp. cubense. Phytochemistry 2003, 63, 679–686. [Google Scholar] [CrossRef]
- Inderjit. Soil microorganisms: An important determinant of allelopathic activity. Plant Soil 2005, 274, 227–236. [Google Scholar] [CrossRef]
- Shaw, L.J.; Morris, P.; Hooker, J.E. Perception and modification of plant flavonoid signals by rhizosphere microorganisms. Environ. Microbiol. 2006, 8, 1867–1880. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, N.; Ahmed, N.; Ghani, M.Y.; Sofi, P.A. Role of phenolic compounds in resistance to chilli wilt. Commun. Biom. Crop Sci. 2009, 4, 52–61. [Google Scholar]
- Mandal, S.; Mitra, A.; Mallick, N. Time course study on accumulation of cell wall-bound phenolics and activities of defense enzymes in tomato roots in relation to Fusarium wilt. World J. Microbiol. Biotechnol. 2009, 25, 795–802. [Google Scholar] [CrossRef]
- Yuan, F.; Zhang, C.L.; Shen, Q.R. Effect and mechanism of phenol compounds in alleviating cucumber Fusarium wilt. Sci. Agric. Sin. 2004, 37, 545–551. [Google Scholar] [CrossRef]
- Di Pietro, A.; Garcia Maceira, F.I.; Meglecz, E.; Roncero, M.I. A MAP kinase of the vascular wilt fungus Fusarium oxysporum is essential for root penetration and pathogenesis. Mol. Microbiol. 2001, 39, 1140–1152. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Q.; Chen, Z.J.; Wei, M.L.; Huang, T.W.; Ke, J.H.; Cui, X.M. Preliminary study on effects of ferulic acid allelopathic on Panax notoginseng. Spec. Wild Econ. Anim. Plant Res. 2008, 30, 39–41. [Google Scholar] [CrossRef]
- Ren, L.X.; Huo, H.W.; Zhang, F.; Hao, W.Y.; Xiao, L.; Dong, C.X.; Xu, G.H. The component difference between rice and watermelon root exudates which lead to their ecological roles on pathogenic fungus and watermelon defense. Plant Signal. Behav. 2016, 11, e1187357. [Google Scholar] [CrossRef] [PubMed]
- Shaik, A.B.; Ahil, S.B.; Govardhanam, R.; Senthi, M.; Khan, R.; Sojitra, R.; Kumar, S.; Srinivas, A. Antifungal effect and protective role of Ursolic acid and three phenolic derivatives in the management of Sorghum Grain Mold under field conditions. Chem. Biodivers. 2016, 13, 1158–1164. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (ppm) | Wilting Rate (%) | Disease Index | FA Concent in Plants (μg/g) |
|---|---|---|---|
| 0 | 0.00 b | 0.00 c | 0 c |
| 50 | 13.33 ± 3.33 a | 6.66 ± 1.93 b | 5.3571 ± 0.2572 bc |
| 100 | 16.67 ± 3.33 a | 11.11 ± 2.22 b | 15.0364 ± 3.9238 b |
| 200 | 20.00 a | 18.89 ± 1.11 a | 36.5687 ± 5.9505 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.-M.; Cheng, Y.-X.; Ma, Y.-N.; Chen, C.-J.; Xu, F.-R.; Dong, X. Role of Phenolic Acids from the Rhizosphere Soils of Panax notoginseng as a Double-Edge Sword in the Occurrence of Root-Rot Disease. Molecules 2018, 23, 819. https://doi.org/10.3390/molecules23040819
Zhao Y-M, Cheng Y-X, Ma Y-N, Chen C-J, Xu F-R, Dong X. Role of Phenolic Acids from the Rhizosphere Soils of Panax notoginseng as a Double-Edge Sword in the Occurrence of Root-Rot Disease. Molecules. 2018; 23(4):819. https://doi.org/10.3390/molecules23040819
Chicago/Turabian StyleZhao, Ya-Meng, Yong-Xian Cheng, Yu-Nan Ma, Chuan-Jiao Chen, Fu-Rong Xu, and Xian Dong. 2018. "Role of Phenolic Acids from the Rhizosphere Soils of Panax notoginseng as a Double-Edge Sword in the Occurrence of Root-Rot Disease" Molecules 23, no. 4: 819. https://doi.org/10.3390/molecules23040819