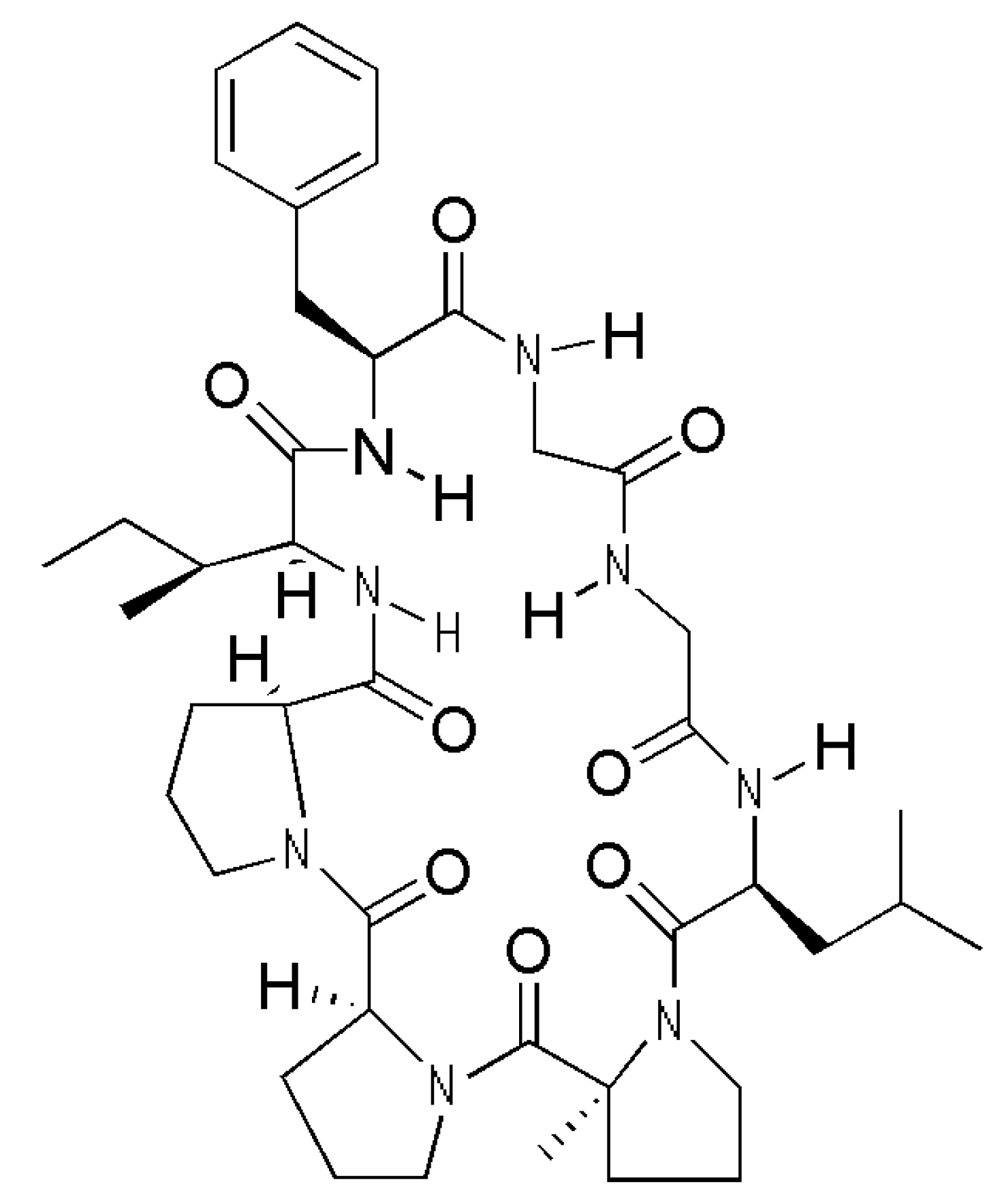
Heterophyllin B Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Macrophages by Suppressing the PI3K/Akt Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
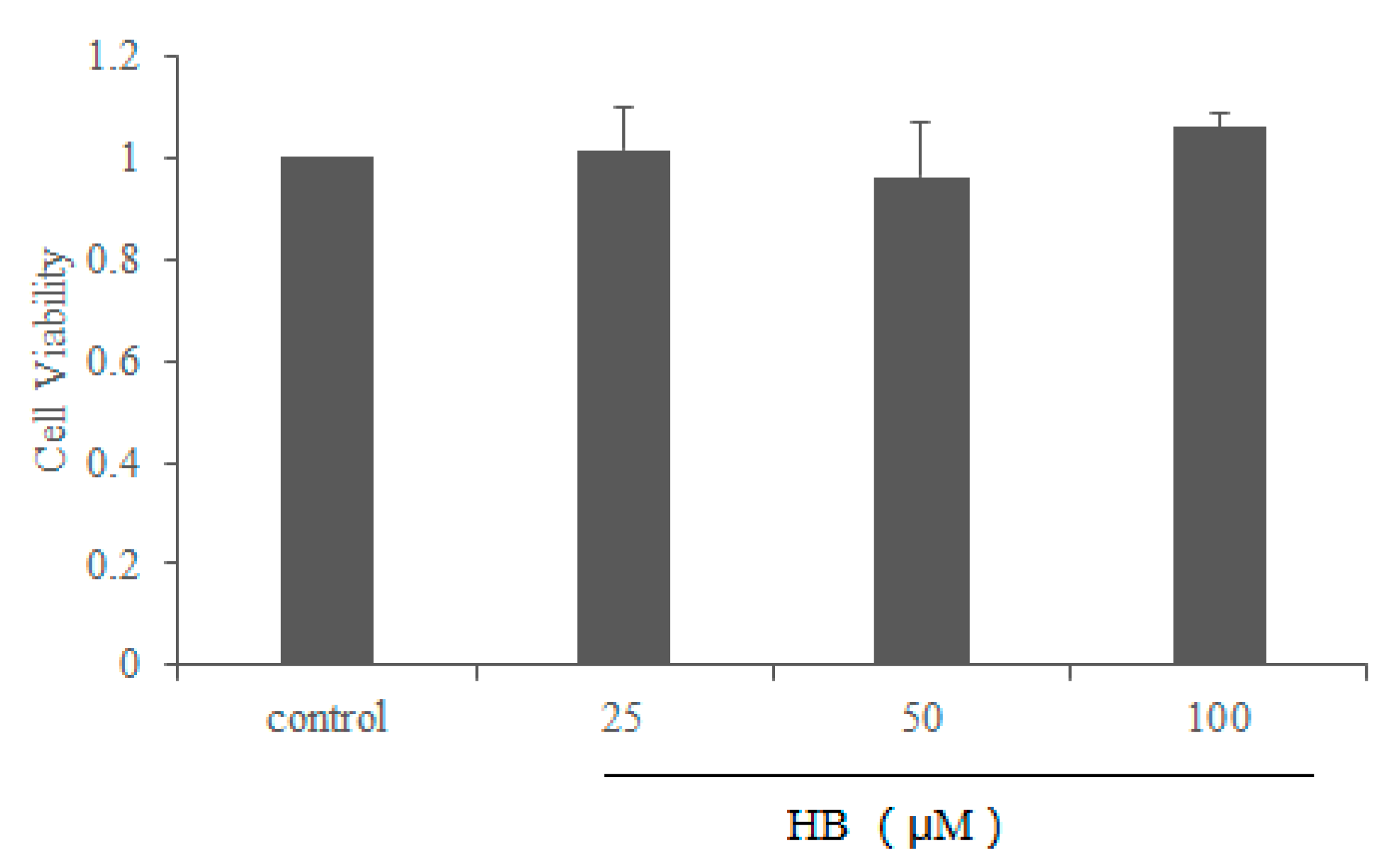
2.1. Effect of HB on Cell Viability
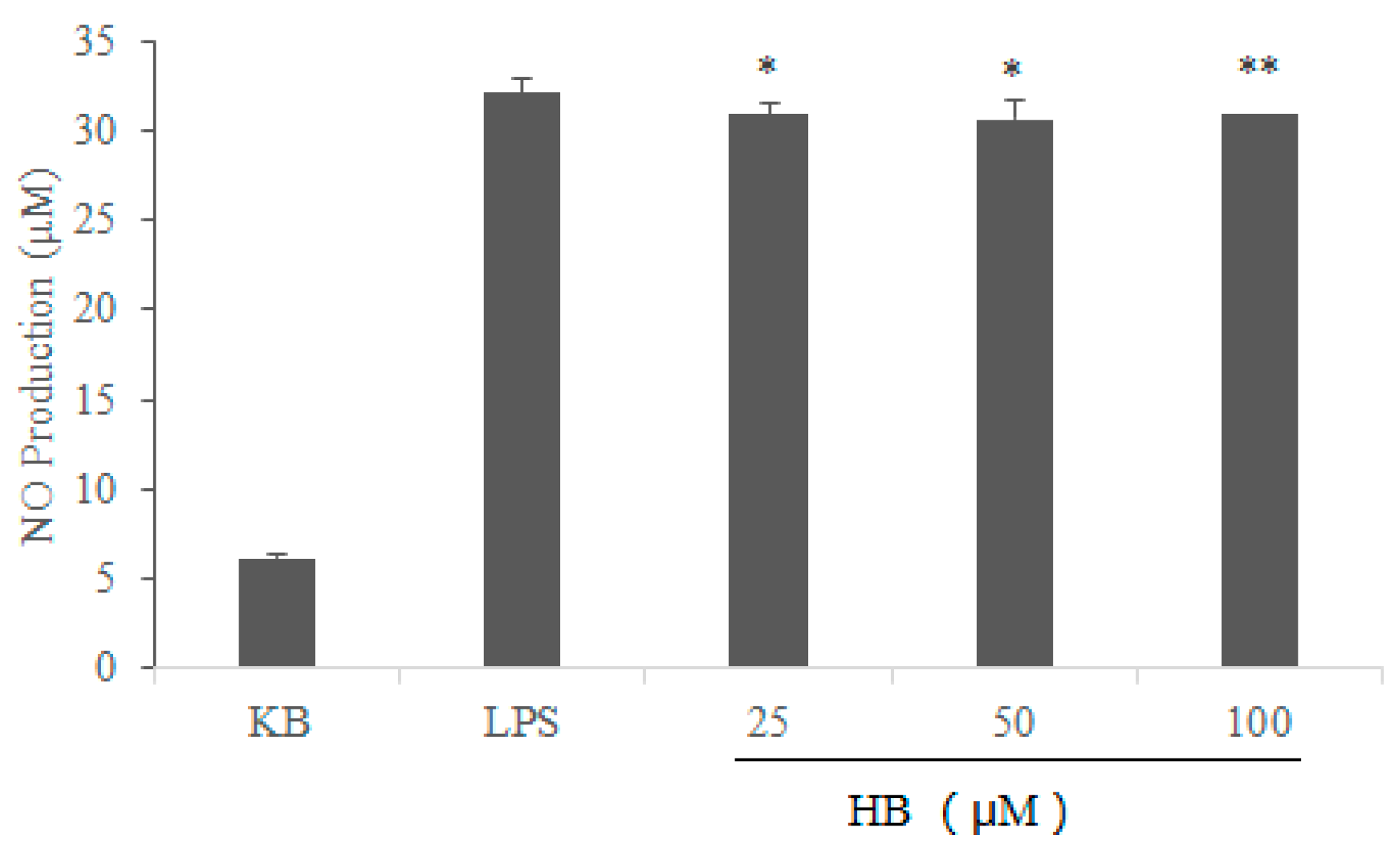
2.2. Effects of HB on LPS-Induced Generation of NO in RAW264.7 Cells
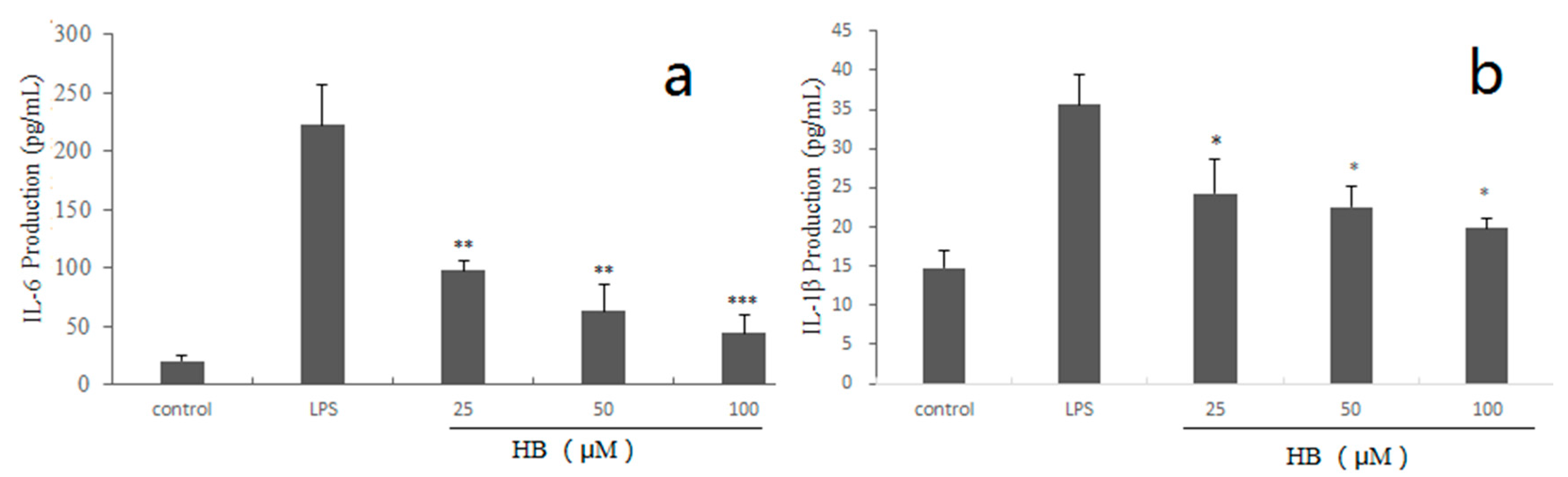
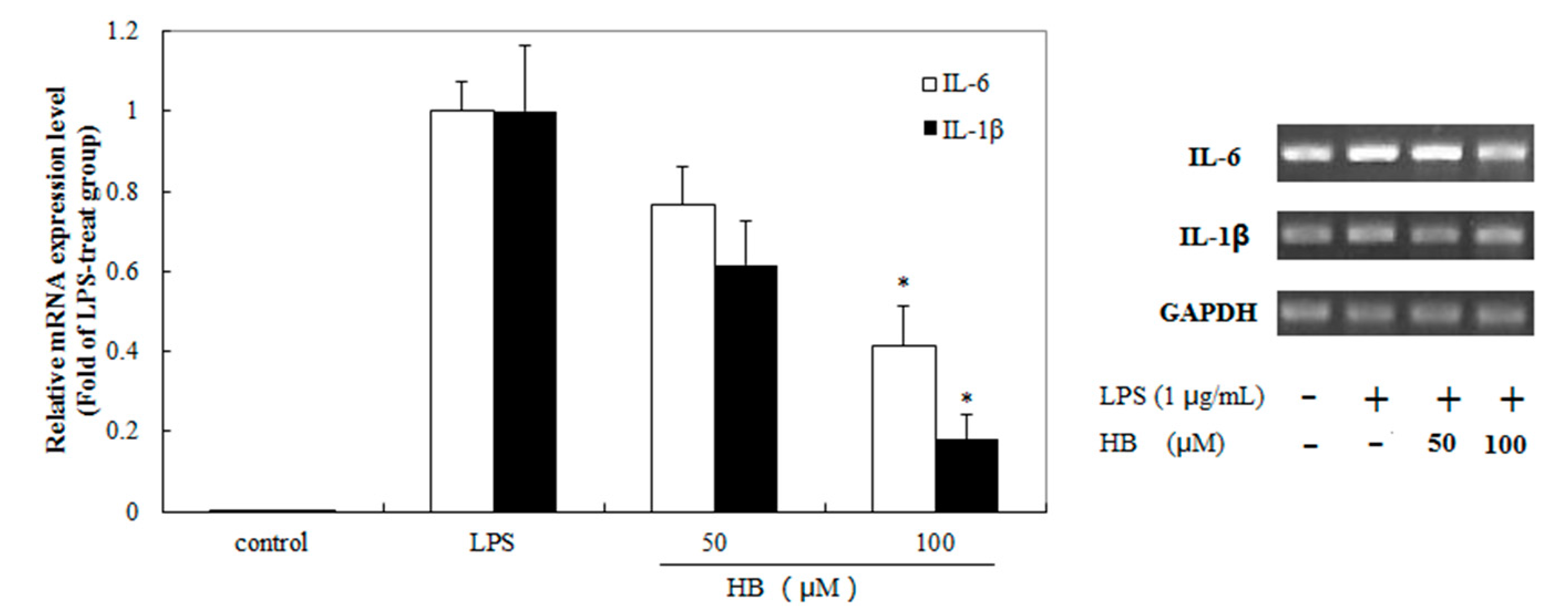
2.3. Effects of HB on LPS-Induced Generation of IL-6, IL-1β in RAW264.7 Cells
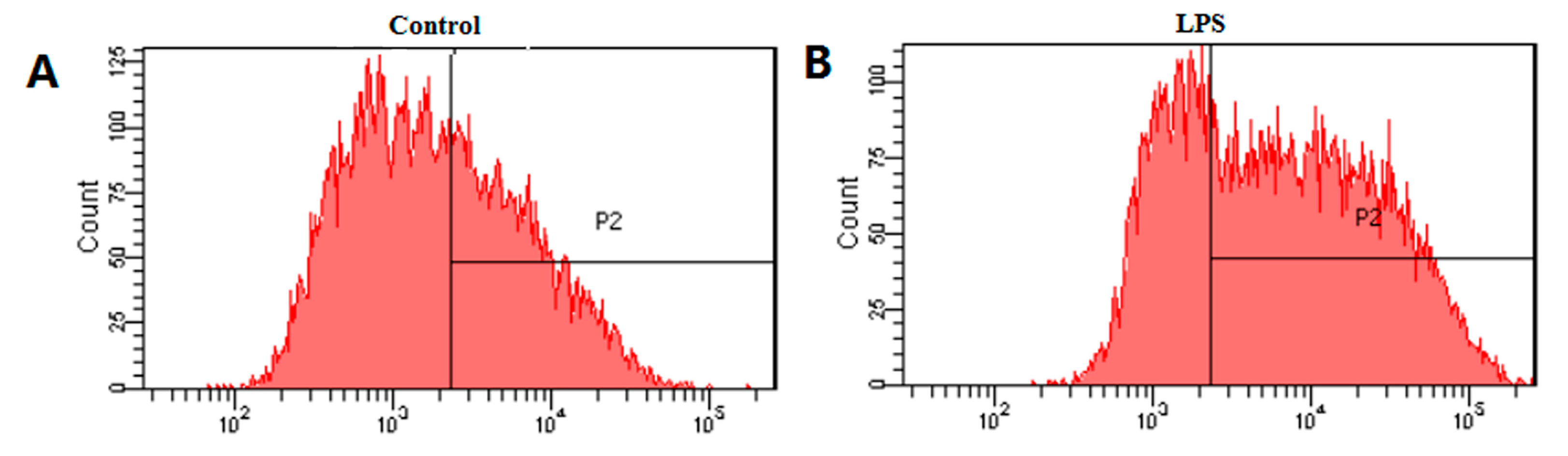
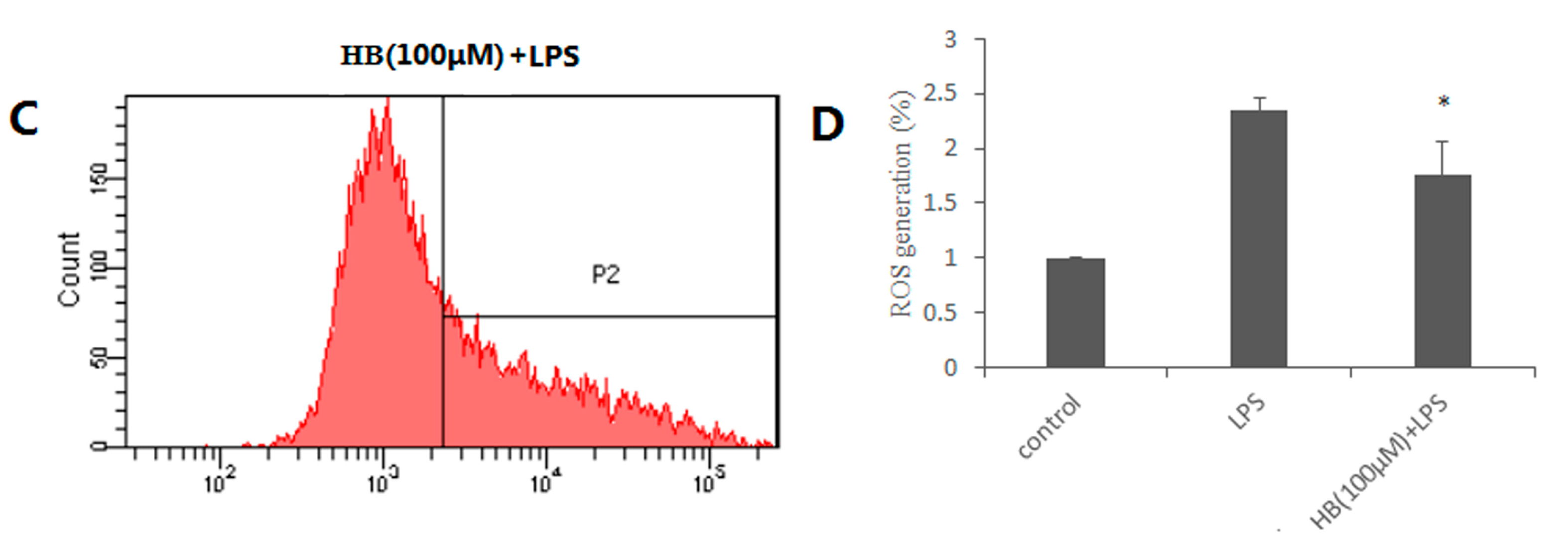
2.4. Effects of HB on LPS-Induced Generation of ROS
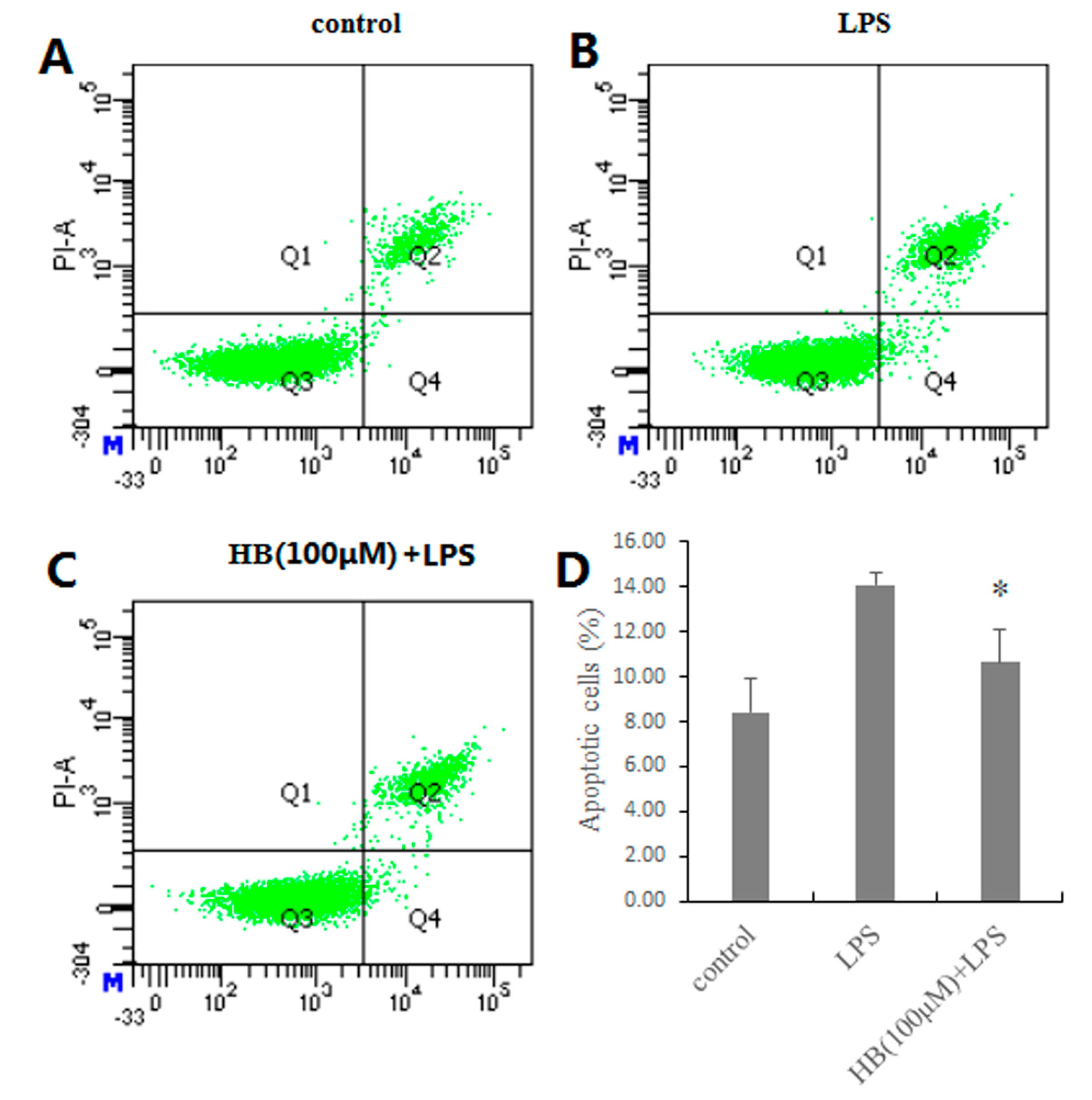
2.5. Effects of HB on LPS-Induced Apoptosis in RAW 264.7 Cells
2.6. Effects of HB Treatment on LPS-Induced IL-6 and IL-1β Expression in RAW 264.7 Cells
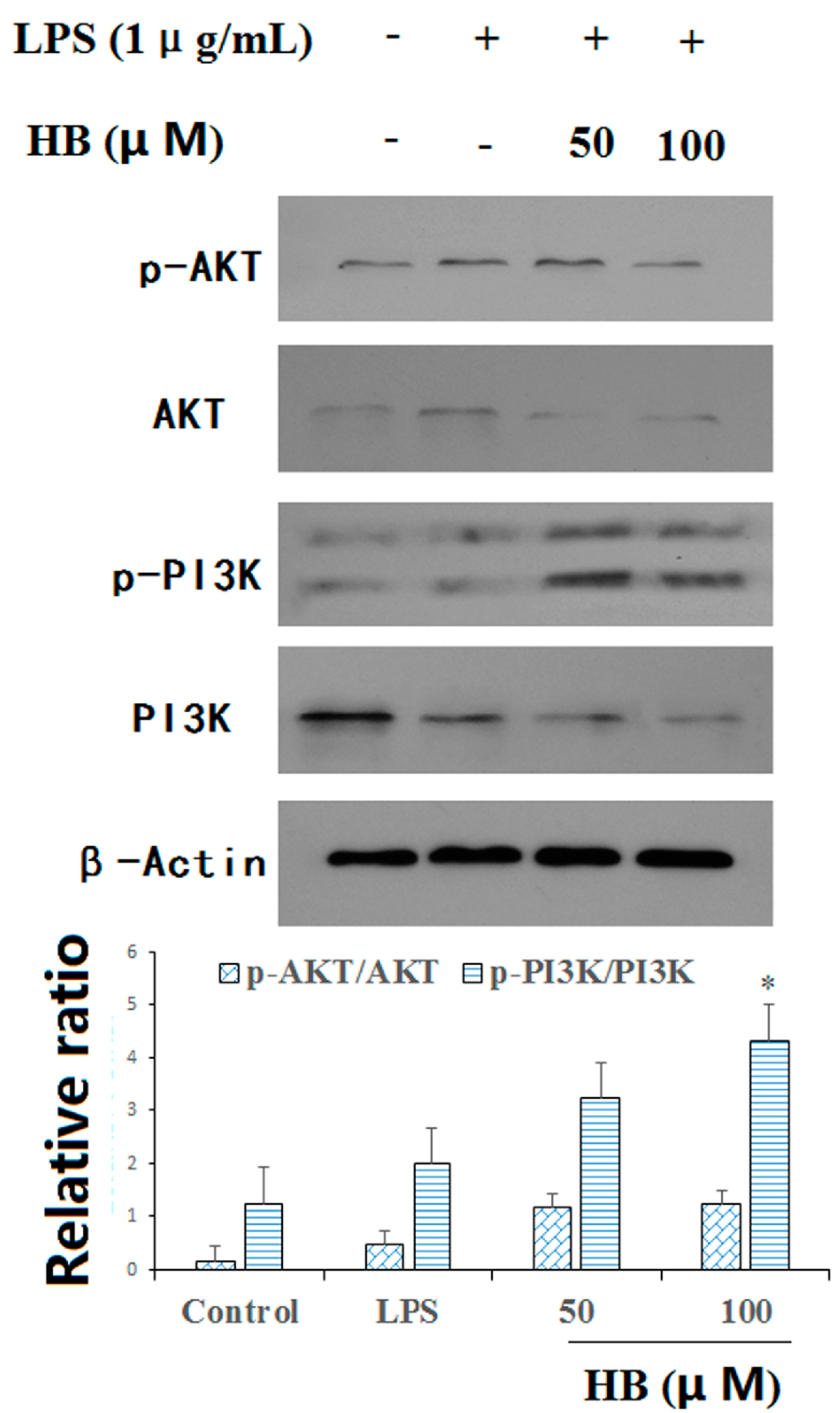
2.7. HB Suppressed LPS-Induced PI3K and AKT Phosphorylation
3. Discussion
4. Materials and Methods
4.1. Drugs and Chemicals
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. NO Production Assay
4.5. ELISA Assays
4.6. Measurement of Intracellular ROS
4.7. Apoptosis Analysis
4.8. qRT-PCR Analysis
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cheng, B.C.Y.; Ma, X.Q.; Kwan, H.Y.; Tse, K.W.; Cao, H.H.; Su, T.; Shu, X.; Wu, Z.Z.; Yu, Z.L. A herbal formula consisting of Rosae Multiflorae Fructus and Lonicerae Japonicae Flos inhibits inflammatory mediators in LPS-stimulated RAW 264.7 macrophages. J. Ethnopharmacol. 2014, 153, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mazza, G. Inhibitory effects of anthocyanins and other phenolic compounds on nitric oxide production in LPS/IFN-γ-activated RAW 264.7 macrophages. J. Agric. Food Chem. 2002, 50, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.K.; Surh, Y.J. Inflammation: Gearing the journey to cancer. Mutat. Res. 2008, 659, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Ning-Hua, T.; Jun, Z.; Chang-Xiang, C.; Shou-Xun, Z. Cyclopeptides from the roots of Pseudostellaria heterophylla. Phytochemistry 1993, 32, 1327–1330. [Google Scholar] [CrossRef]
- Tan, N.H.; Zhou, J. Plant cyclopeptides. Chem. Rev. 2006, 106, 840–895. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Müller, A.; Miess, H.; Grossb, H. Cyclic lipopeptides as antibacterial agents–potent antibiotic activity mediated by intriguing mode of actions. Int. J. Med. Microbiol. 2014, 304, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.B.; Tan, N.H.; Zhang, F.; Lu, Y.Q.; He, M.; Zhou, J. Cyclopeptides and amides from Pseudostellaria heterophylla (Caryophyllaceae). Helv. Chim. Acta 2003, 86, 3376–3379. [Google Scholar] [CrossRef]
- Fleury, C.; Mignotte, B.; Vayssière, J.L. Mitochondrial reactive oxygen species in cell death signaling. Biochimie 2002, 84, 131–141. [Google Scholar] [CrossRef]
- Sauer, H.; Wartenberg, M.; Hescheler, J. Reactive Oxygen Species as Intracellular Messengers during Cell Growth and Differentiation. Cell. Physiol. Biochem. 2001, 11, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Cuschieri, J.; Maier, R.V. Oxidative stress, lipid rafts, and macrophage reprogramming. Antioxid. Redox Signal. 2007, 9, 1485–1498. [Google Scholar] [CrossRef] [PubMed]
- Lucas, K.; Maes, M. Role of the Toll Like receptor (TLR) radical cycle in chronic inflammation: Possible treatments targeting the TLR4 pathway. Mol. Neurobiol. 2013, 48, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.A.; Hwang, Y.J.; Song, J. Antioxidant activities and oxidative stress inhibitory effects of ethanol extracts from Cornus officinalis on raw 264.7 cells. BMC Complement. Altern. Med. 2016, 16, 196. [Google Scholar] [CrossRef] [PubMed]
- Shu, M.; Hu, X.R.; Hung, Z.A.; Huang, D.D.; Zhang, S. Effects of tanshinone IIA on fibrosis in a rat model of cirrhosis through heme oxygenase-1, inflammation, oxidative stress and apoptosis. Mol. Med. Rep. 2016, 13, 3036–3042. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.O.; Pang, L.; Dong, N.; Yang, S. LC-ESI-MS/MS analysis and pharmacokinetics of heterophyllin B, a cyclic octapeptide from Pseudostellaria heterophylla in rat plasma. Biomed. Chromatogr. 2015, 29, 1693–1699. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Ng, T.B. Concurrent isolation of a Kunitz-type trypsin inhibitor with antifungal activity and a novel lectin from Pseudostellaria heterophylla roots. Biochem. Biophys. Res. Commun. 2006, 342, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.; Lin, S.; Dai, Q.; Zhang, H.; Hu, J. Antitussive activity of Pseudostellaria heterophylla (Miq.) Pax extracts and improvement in lung function via adjustment of multi-cytokine levels. Molecules 2011, 16, 3360–3370. [Google Scholar] [CrossRef] [PubMed]
- Higashimoto, T.; Panopoulos, A.; Hsieh, C.L.; Zandi, E. TNFα induces chromosomal abnormalities independent of ROS through IKK, JNK, p38 and caspase pathways. Cytokine 2006, 34, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Sizemore, N.; Leung, S.; Stark, G.R. Activation of phosphatidylinositol 3-kinase in response to interleukin-1 leads to phosphorylation and activation of the NF-κB p65/RelA subunit. Mol. Cell. Boil. 1999, 19, 4798–4805. [Google Scholar] [CrossRef]
- Chan, J.Y.H.; Chan, S.H.H. Activation of endogenous antioxidants as a common therapeutic strategy against cancer, neurodegeneration and cardiovascular diseases: A lesson learnt from DJ-1. Pharmacol. Ther. 2015, 156, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gonzalez, M.I.; Meinkoth, J.L.; Field, J.; Kazanietz, M.G.; Tennekoon, G.I. Lysophosphatidic acid promotes survival and differentiation of rat Schwann cells. J. Biol. Chem. 2003, 278, 9585–9591. [Google Scholar] [CrossRef] [PubMed]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Dery, M.C.; Leblanc, V.; Shooner, C.; Asselin, E. Regulation of Akt expression and phosphorylation by 17β-estradiol in the rat uterus during estrous cycle. Reprod. Biol. Endocrinol. 2003, 1, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, Y.Y.; Song, J.H.; Shin, Y.K.; Han, E.S.; Lee, C.S. Depressant effects of ambroxol and erdosteine on cytokine synthesis, granule enzyme release, and free radical production in rat alveolar macrophages activated by lipopolysaccharide. Basic Clin. Pharmacol. Toxicol. 2003, 92, 173–179. [Google Scholar] [CrossRef]
- Du, X.; Shi, Z.; Peng, Z.; Zhao, C.; Zhang, Y.; Wang, Z.; Li, X.; Liu, G.; Li, X. Acetoacetate Induces Hepatocytes Apoptosis by the ROS-Mediated MAPKs Pathway in Ketotic Cows. J. Cell. Physiol. 2017, 232, 3296–3308. [Google Scholar] [CrossRef] [PubMed]
- Afri, M.; Frimer, A.A.; Cohen, Y. Active oxygen chemistry within the liposomal bilayer. Part IV: Locating 2′,7′-dichlorofluorescein(DCF), 2′,7′-dichlorodihydrofluorescein(DCFH) and 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) in the lipid bilayer. Chem. Phys. Lipids 2004, 131, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Judware, R.; McCormick, T.S.; Yun, J.K.; Lapetina, E.G. Apoptosis of RAW264. 7 peritoneal macrophages: Relationship between apoptosis and integrin expression. In Nitric Oxide and the Cell: Proliferation, Differentiation, and Death; Princeton University Press: Princeton, NJ, USA, 2017; p. 271. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ci, X.; Ren, R.; Xu, K.; Li, H.; Yu, Q.; Song, Y.; Wang, D.; Li, R.; Deng, X. Schisantherin a exhibits anti-inflammatory properties by down-regulating NF-κB and MAPK signaling pathways in lipopolysaccharide-treated RAW 264.7 cells. Inflammation 2010, 33, 126–136. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; You, L.; Yin, X.; Liu, Y.; Leng, X.; Wang, W.; Sai, N.; Ni, J. Heterophyllin B Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Macrophages by Suppressing the PI3K/Akt Pathways. Molecules 2018, 23, 717. https://doi.org/10.3390/molecules23040717
Yang C, You L, Yin X, Liu Y, Leng X, Wang W, Sai N, Ni J. Heterophyllin B Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Macrophages by Suppressing the PI3K/Akt Pathways. Molecules. 2018; 23(4):717. https://doi.org/10.3390/molecules23040717
Chicago/Turabian StyleYang, Chunjing, Longtai You, Xingbin Yin, Yi Liu, Xin Leng, Wenping Wang, Na Sai, and Jian Ni. 2018. "Heterophyllin B Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Macrophages by Suppressing the PI3K/Akt Pathways" Molecules 23, no. 4: 717. https://doi.org/10.3390/molecules23040717