Pineal Calcification, Melatonin Production, Aging, Associated Health Consequences and Rejuvenation of the Pineal Gland


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
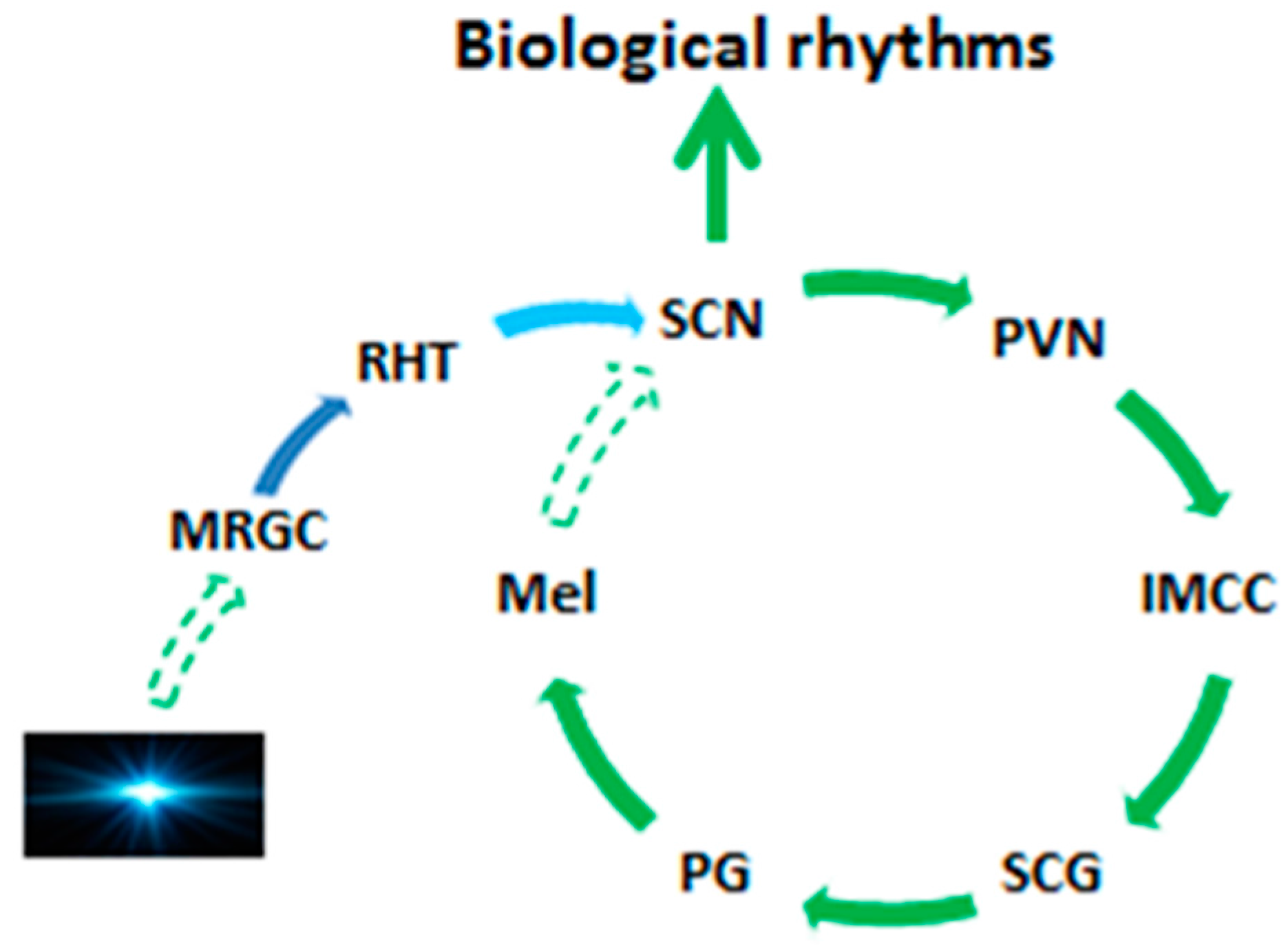
2. Pineal Gland and the Melatonin Circadian Rhythm
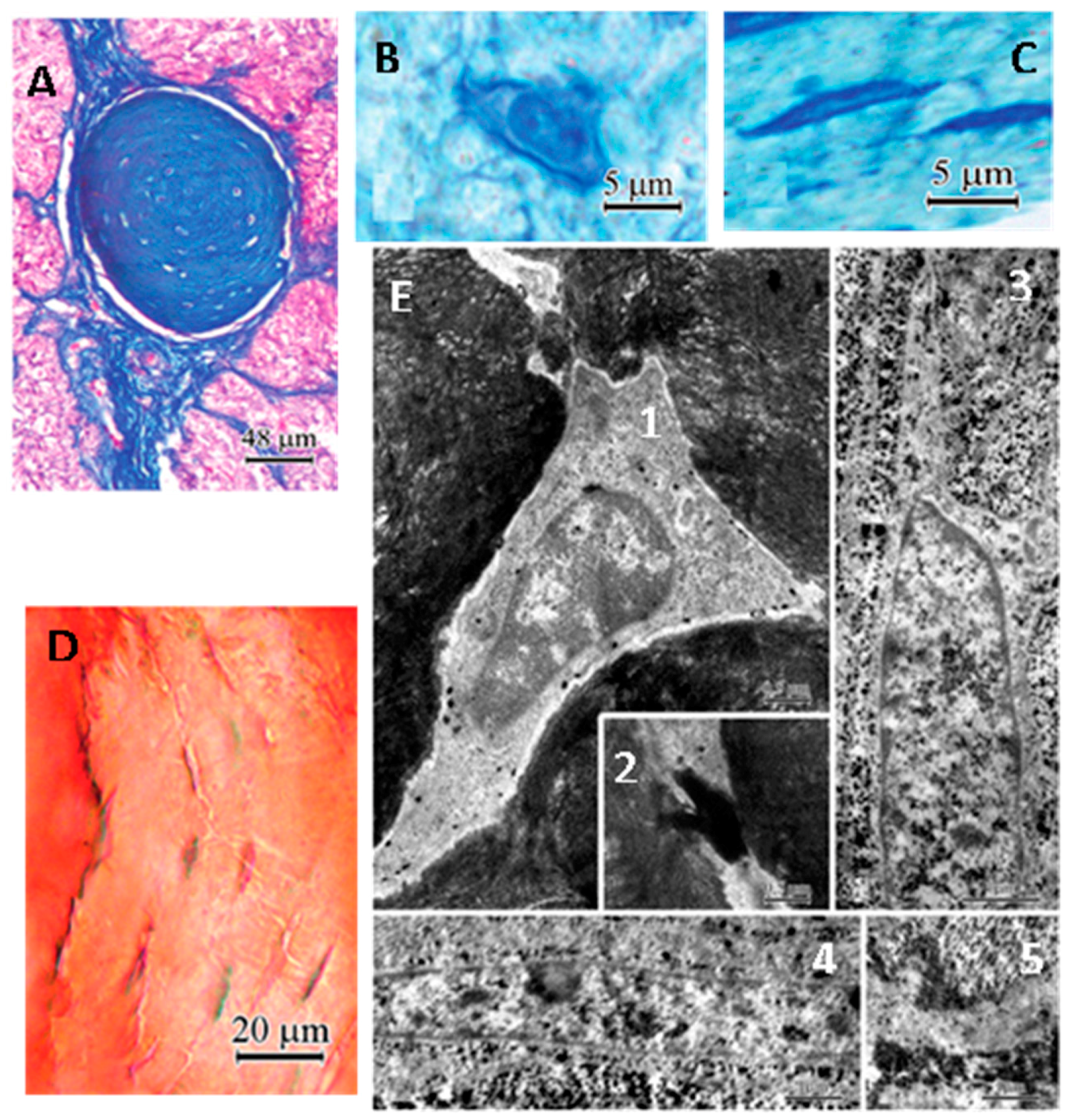
3. Pineal Gland Calcification (PGC), Melatonin Production, Neurodegenerative Diseases and Aging
4. Potential Mechanisms for PGC Formation
- (1)
- Chronic vascular inflammation: The pineal gland has a complicated vascular system with abundance of arteries, fenestrated capillaries and veins. Especially the filtration rate of blood in pineal gland is in excess of most organs and it is only second to the kidney in terms of blood flow. These make the gland venerable to the chronic vascular inflammation during aging or certain disorders. The vascular inflammation mobilizes the MSCs migration and adhesion in the gland or promotes the de novo MSCs proliferation due to the increased levels of pro-inflammatory cytokines, TGF-β or TNF-α. The crosstalk between vascular MSCs and inflammatory mediators, especially, interleukin-22, lead to MSCs proliferation, migration and osteogenic differentiation [266,267] under the influence of high levels of pineal melatonin and finally PGC formation.
- (2)
- Brain tissue hypoxia: Many pathological conditions cause brain tissue hypoxia including hypertension, sleep apnea, stroke, and even respiratory disorders. Hypoxia-inducible factor (HIF)-1α is an important regulator of MSCs and it promotes the proliferation, migration and adhesion of MSCs in the hypoxic areas [268,269,270] including to the pineal gland. Generally, hypoxia increases bone resorption and suppresses osteoblastic differentiation and bone-formation [271,272]. However, this may not be applied to the pineal gland. During the dark phase, the pineal produces high levels of melatonin. Under the hypoxic condition, melatonin would promote the osteoblast differentiation and mineralization of MSCs via the p38 MAPK and PRKD1 signaling pathways [273]. In addition, melatonin also inhibits the activity of the osteoclast and osteoclatogenesis [274,275], especially under inflammatory conditions [276]. These processes favor PGC formation under hypoxic conditions.
- (3)
- Intracranial pressure: Some cells of the pineal gland are “swimming” in the third ventricle and, as a result, they are influenced by the intracranial pressure. Intracranial pressure usually increases with cerebral disorders such as idiopathic intracranial hypertension, brain trauma and stroke [277], and even Alzheimer’s disease [278]. The high pressure may impede the pineal filtration rate and induce endoepithelial cell damage by chrono-inflammation. The pressure also promotes the bone remodeling and mineralization, thus, PGC formation.
5. Rejuvenation of Pineal Gland?
6. Conclusions
Conflicts of Interest
References
- Golan, J.; Torres, K.; Staśkiewicz, G.J.; Opielak, G.; Maciejewski, R. Morphometric parameters of the human pineal gland in relation to age, body weight and height. Folia Morphol. 2002, 61, 111–113. [Google Scholar]
- Tan, D.X.; Manchester, L.C.; Fuentes-Broto, L.; Paredes, S.D.; Reiter, R.J. Significance and application of melatonin in the regulation of brown adipose tissue metabolism: Relation to human obesity. Obes. Rev. 2011, 12, 167–188. [Google Scholar] [CrossRef] [PubMed]
- Hintermann, E.; Jenö, P.; Meyer, U.A. Isolation and characterization of an arylalkylamine N-acetyltransferase from Drosophila melanogaster. FEBS Lett. 1995, 375, 148–150. [Google Scholar] [CrossRef]
- Itoh, M.T.; Hattori, A.; Sumi, Y.; Suzuki, T. Day-night changes in melatonin levels in different organs of the cricket (Gryllus bimaculatus). J. Pineal Res. 1995, 18, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Vivien-Roels, B.; Pevet, P.; Beck, O.; Fevre-Montange, M. Identification of melatonin in the compound eyes of an insect, the locust (Locusta migratoria), by radioimmunoassay and gas chromatography-mass spectrometry. Neurosci. Lett. 1984, 49, 153–157. [Google Scholar] [CrossRef]
- Ralph, C.L. The pineal gland and geographical distribution of animals. Int. J. Biometeorol. 1975, 19, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Grosshans, M.; Vollmert, C.; Vollstaedt-Klein, S.; Nolte, I.; Schwarz, E.; Wagner, X.; Leweke, M.; Mutschler, J.; Kiefer, F.; Bumb, J.M. The association of pineal gland volume and body mass in obese and normal weight individuals: A pilot study. Psychiatr. Danub. 2016, 28, 220–224. [Google Scholar] [PubMed]
- Bumb, J.M.; Schilling, C.; Enning, F.; Haddad, L.; Paul, F.; Lederbogen, F.; Deuschle, M.; Schredl, M.; Nolte, I. Pineal gland volume in primary insomnia and healthy controls: A magnetic resonance imaging study. J. Sleep Res. 2014, 23, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Bryden, M.M.; Griffiths, D.J.; Kennaway, D.J.; Ledingham, J. The pineal gland is very large and active in newborn antarctic seals. Experientia 1986, 42, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Cuello, A.C.; Tramezzani, J.H. The epiphysis cerebri of the Weddell seal: Its remarkable size and glandular pattern. Gen. Comp. Endocrinol. 1969, 12, 154–164. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Sainz, R.M.; Mayo, J.C.; León, J.; Reiter, R.J. Physiological ischemia/reperfusion phenomena and their relation to endogenous melatonin production: A hypothesis. Endocrine 2005, 27, 149–158. [Google Scholar] [CrossRef]
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of melatonin, the pineal gland factor that lightens melanocytes. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Pavel, S.; Petrescu, S. Inhibition of gonadotrophin by a highly purified pineal peptide and by synthetic arginine vasotocin. Nature 1966, 212, 1054. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.K.; Kopylov, A.T.; Vaskovsky, B.V.; Ryzhak, G.A.; Lin’kova, N.S. Identification of Peptide AEDG in the Polypeptide Complex of the Pineal Gland. Bull. Exp. Biol. Med. 2017, 164, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.R.; Persinger, M.A. Application of transcerebral, weak (1 microT) complex magnetic fields and mystical experiences: Are they generated by field-induced dimethyltryptamine release from the pineal organ? Percept. Mot. Skills 2003, 97, 1049–1050. [Google Scholar] [CrossRef] [PubMed]
- Guchhait, R.B. Biogenesis of 5-methoxy-N,N-dimethyltryptamine in human pineal gland. J. Neurochem. 1976, 26, 187–190. [Google Scholar] [PubMed]
- Nichols, D.E. N,N-dimethyltryptamine and the pineal gland: Separating fact from myth. J. Psychopharmacol. 2017, 32, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, K.; Haraguchi, S.; Vaudry, H. 7α-Hydroxypregnenolone regulating locomotor behavior identified in the brain and pineal gland across vertebrates. Gen. Comp. Endocrinol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J. Pineal Melatonin: Cell Biology of Its Synthesis and of Its Physiological Interactions. Endocr. Rev. 1991, 12, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P.; Wurtman, R.J. Hydroxyindole-O-methyl transferases in rat pineal, retina and harderian gland. Endocrinology 1972, 91, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Nagle, C.A.; Cardinali, D.P.; Rosner, J.M. Retinal and pineal hydroxyindole-O-methyl transferases in the rat: Changes following cervical sympathectomy, pinealectomy or blinding. Endocrinology 1973, 92, 1560–1564. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, R.A.; Johnson, L.B.; Reiter, R.J. Regulation of melatonin in the harderian glands of golden hamsters. J. Pineal Res. 1989, 6, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Pelaez, A.; Reiter, R.J.; Howes, K.A.; Puig-Domingo, M.; Vaughan, M.K.; Troiani, M.E.; Little, J.C. Harderian gland N-acetyltransferase activity in the male Syrian hamster: Effects of gonadectomy, short photoperiod exposure, or subcutaneous melatonin implants. Endocr. Res. 1988, 14, 121–130. [Google Scholar] [PubMed]
- Menendez-Pelaez, A.; Reiter, R.J.; Gonzalez-Brito, A.; Howes, K.A. N-Acetyltransferase activity and indole contents of the male Syrian hamster Harderian gland: Changes during the light:Dark cycle. Arch. Biochem. Biophys. 1989, 268, 20–25. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Back, K.; Manchester, L.C.; Alatorre-Jimenez, M.A.; Reiter, R.J. On the significance of an alternate pathway of melatonin synthesis via 5-methoxytryptamine: Comparisons across species. J. Pineal Res. 2016, 61, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin metabolism in the central nervous system. Curr. Neuropharmacol. 2010, 8, 168–181. [Google Scholar] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Kim, T.-K.; Janjetovic, Z.; Slominski, R.M.; Zmijewski, J.W. Melatonin, mitochondria, and the skin. Cell. Mol. Life Sci. 2017, 74, 3913–3925. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Itoh, M.T.; Miyata, M.; Ishikawa, S.; Sumi, Y. Detection of melatonin, its precursors and related enzyme activities in rabbit lens. Exp. Eye Res. 1999, 68, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Alkozi, H.A.; Pintor, J. TRPV4 activation triggers the release of melatonin from human non-pigmented ciliary epithelial cells. Exp. Eye Res. 2015, 136, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Alkozi, H.A.; Perez de Lara, M.J.; Pintor, J. Melatonin synthesis in the human ciliary body triggered by TRPV4 activation: Involvement of AANAT phosphorylation. Exp. Eye Res. 2017, 162, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Mayo, J.C.; Sainz, R.M.; Leon, J.; Bandyopadhyay, D. Neurally-mediated and neurally-independent beneficial actions of melatonin in the gastrointestinal tract. J. Physiol. Pharmacol. 2003, 54 (Suppl. 4), 113–125. [Google Scholar] [PubMed]
- Reiter, R.J.; Rosales-Corral, S.; Coto-Montes, A.; Boga, J.A.; Tan, D.-X.; Davis, J.M.; Konturek, P.C.; Konturek, S.J.; Brzozowski, T. The photoperiod, circadian regulation and chronodisruption: The requisite interplay between the suprachiasmatic nuclei and the pineal and gut melatonin. J. Physiol. Pharmacol. 2011, 62, 269–274. [Google Scholar] [PubMed]
- Tijmes, M.; Pedraza, R.; Valladares, L. Melatonin in the rat testis: Evidence for local synthesis. Steroids 1996, 61, 65–68. [Google Scholar] [CrossRef]
- Itoh, M.T.; Ishizuka, B.; Kudo, Y.; Fusama, S.; Amemiya, A.; Sumi, Y. Detection of melatonin and serotonin N-acetyltransferase and hydroxyindole-O-methyltransferase activities in rat ovary. Mol. Cell. Endocrinol. 1997, 136, 7–13. [Google Scholar] [CrossRef]
- Khan, Z.A.; Yumnamcha, T.; Rajiv, C.; Sanjita Devi, H.; Mondal, G.; Devi, S.D.; Bharali, R.; Chattoraj, A. Melatonin biosynthesizing enzyme genes and clock genes in ovary and whole brain of zebrafish (Danio rerio): Differential expression and a possible interplay. Gen. Comp. Endocrinol. 2016, 233, 16–31. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, J.; Li, Y.; Zhu, K.; Xu, Z.; Song, Y.; Song, Y.; Liu, G. Melatonin-related genes expressed in the mouse uterus during early gestation promote embryo implantation. J. Pineal Res. 2015, 58, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Qi, W.B.; Zhang, M.; Weintraub, S.T.; Cabrera, J.; Sainz, R.M.; Mayo, J.C. Identification of highly elevated levels of melatonin in bone marrow: Its origin and significance. Biochim. Biophys. Acta 1999, 1472, 206–214. [Google Scholar] [CrossRef]
- Conti, A.; Conconi, S.; Hertens, E.; Skwarlo-Sonta, K.; Markowska, M.; Maestroni, J.M. Evidence for melatonin synthesis in mouse and human bone marrow cells. J. Pineal Res. 2000, 28, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.; Lacasse, A.-A.; Lanoix, D.; Sagrillo-Fagundes, L.; Boulard, V.; Vaillancourt, C. Placental melatonin system is present throughout pregnancy and regulates villous trophoblast differentiation. J. Pineal Res. 2015, 59, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Lanoix, D.; Guérin, P.; Vaillancourt, C. Placental melatonin production and melatonin receptor expression are altered in preeclampsia: New insights into the role of this hormone in pregnancy. J. Pineal Res. 2012, 53, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, K.; Itoh, M.T.; Takahashi, N.; Tarumi, W.; Ishizuka, B. The rat oocyte synthesises melatonin. Reprod. Fertil. Dev. 2013, 25, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Rosengarten, H.; Meller, E.; Friedhoff, A.J. In vitro enzymatic formation of melatonin by human erythrocytes. Res. Commun. Chem. Pathol. Pharmacol. 1972, 4, 457–465. [Google Scholar] [PubMed]
- Launay, J.M.; Lamaître, B.J.; Husson, H.P.; Dreux, C.; Hartmann, L.; Da Prada, M. Melatonin synthesis by rabbit platelets. Life Sci. 1982, 31, 1487–1494. [Google Scholar] [CrossRef]
- Carrillo-Vico, A.; Calvo, J.R.; Abreu, P.; Lardone, P.J.; García-Mauriño, S.; Reiter, R.J.; Guerrero, J.M. Evidence of melatonin synthesis by human lymphocytes and its physiological significance: Possible role as intracrine, autocrine, and/or paracrine substance. FASEB J. 2004, 18, 537–539. [Google Scholar] [CrossRef] [PubMed]
- Park, O.K.; Yoo, K.-Y.; Lee, C.H.; Choi, J.H.; Hwang, I.K.; Park, J.H.; Kwon, Y.-G.; Kim, Y.-M.; Won, M.-H. Arylalkylamine N-acetyltransferase (AANAT) is expressed in astrocytes and melatonin treatment maintains AANAT in the gerbil hippocampus induced by transient cerebral ischemia. J. Neurol. Sci. 2010, 294, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.D.; Mora-Santos, M.; Naji, L.; Carrascosa-Salmoral, M.P.; Naranjo, M.C.; Calvo, J.R. Evidence of melatonin synthesis and release by mast cells. Possible modulatory role on inflammation. Pharmacol. Res. 2010, 62, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Uz, T.; Qu, T.; Sugaya, K.; Manev, H. Neuronal expression of arylalkylamine N-acetyltransferase (AANAT) mRNA in the rat brain. Neurosci. Res. 2002, 42, 309–316. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.-X.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell. Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Paredes, S.D.; Korkmaz, A.; Sainz, R.M.; Mayo, J.C.; Fuentes-Broto, L.; Reiter, R.J. The changing biological roles of melatonin during evolution: From an antioxidant to signals of darkness, sexual selection and fitness. Biol. Rev. 2010, 85, 607–623. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Blask, D.E.; Talbot, J.A.; Barnett, M.P. Nature and the time course of seizures associated with surgical removal of the pineal gland from parathyroidectomized rats. Exp. Neurol. 1973, 38, 386–397. [Google Scholar] [CrossRef]
- Tosches, M.A.; Bucher, D.; Vopalensky, P.; Arendt, D. Melatonin Signaling Controls Circadian Swimming Behavior in Marine Zooplankton. Cell 2014, 159, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.F.; Ralph, C.L. Pineal and serum melatonin at midday and midnight following pinealectomy or castration in male rats. J. Exp. Zool. 1975, 193, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Pelham, R.W. A Serum Melatonin Rhythm in Chickens and Its Abolition by Pinealectomy. Endocrinology 1975, 96, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J.; Tetsuo, M.; Markey, S.P.; Goodwin, F.K.; Kopin, I.J. Pinealectomy abolishes plasma melatonin in the rat. J. Clin. Endocrinol. Metab. 1980, 50, 204–205. [Google Scholar] [CrossRef] [PubMed]
- Suofu, Y.; Li, W.; Jean-Alphonse, F.G.; Jia, J.; Khattar, N.K.; Li, J.; Baranov, S.V.; Leronni, D.; Mihalik, A.C.; He, Y.; et al. Dual role of mitochondria in producing melatonin and driving GPCR signaling to block cytochrome c release. Proc. Natl. Acad. Sci. USA 2017, 114, E7997–E8006. [Google Scholar] [CrossRef] [PubMed]
- McMahon, D.G.; Iuvone, P.M.; Tosini, G. Circadian organization of the mammalian retina: From gene regulation to physiology and diseases. Prog. Retin. Eye Res. 2014, 39, 58–76. [Google Scholar] [CrossRef] [PubMed]
- Buonfiglio, D.C.; Malan, A.; Sandu, C.; Jaeger, C.; Cipolla-Neto, J.; Hicks, D.; Felder-Schmittbuhl, M.-P. Rat retina shows robust circadian expression of clock and clock output genes in explant culture. Mol. Vis. 2014, 20, 742–752. [Google Scholar] [PubMed]
- Underwood, H.; Binkley, S.; Siopes, T.; Mosher, K. Melatonin rhythms in the eyes, pineal bodies, and blood of Japanese quail (Coturnix coturnix japonica). Gen. Comp. Endocrinol. 1984, 56, 70–81. [Google Scholar] [CrossRef]
- Yu, H.S.; Pang, S.F.; Tang, P.L. Increase in the level of retinal melatonin and persistence of its diurnal rhythm in rats after pinealectomy. J. Endocrinol. 1981, 91, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.S.; Pang, S.F.; Tang, P.L.; Brown, G.M. Persistence of circadian rhythms of melatonin and N-acetylserotonin in the serum of rats after pinealectomy. Neuroendocrinology 1981, 32, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Hardeland, R.; Lopez-Burillo, S.; Mayo, J.C.; Sainz, R.M.; Reiter, R.J. Melatonin: A hormone, a tissue factor, an autocoid, a paracoid, and an antioxidant vitamin. J. Pineal Res. 2003, 34, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Goldman, H.; Wurtman, R.J. Flow of Blood to the Pineal Body of the Rat. Nature 1964, 203, 87–88. [Google Scholar] [CrossRef] [PubMed]
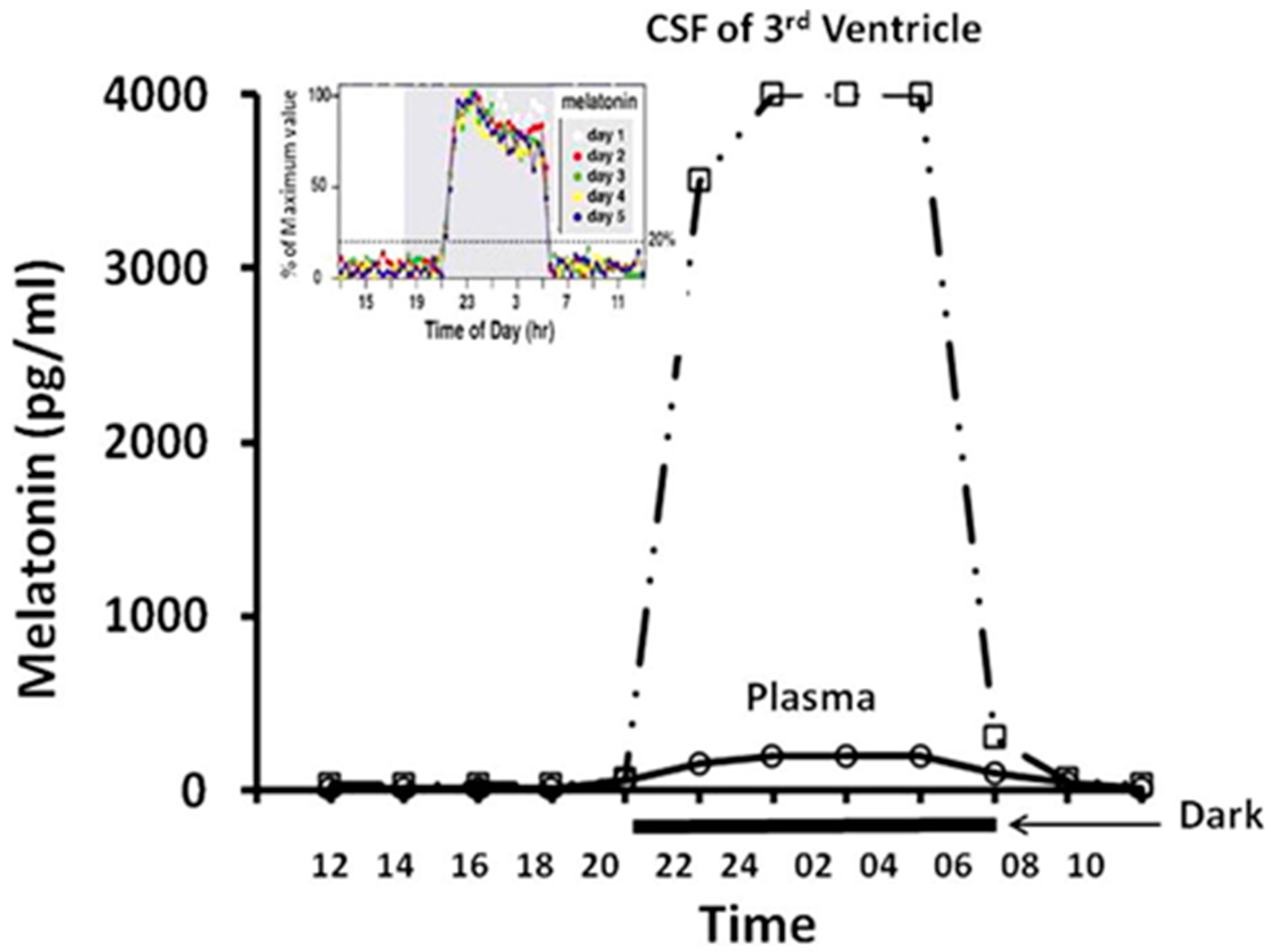
- Tan, D.-X.; Manchester, L.C.; Reiter, R.J. CSF generation by pineal gland results in a robust melatonin circadian rhythm in the third ventricle as an unique light/dark signal. Med. Hypotheses 2016, 86, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.S.; Dixit, A.S. Morphology and ultrastructural studies of pineal organ of the tree sparrow (Passer montanus). Micron 2014, 58, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Yalcin, A.; Ceylan, M.; Bayraktutan, O.F.; Sonkaya, A.R.; Yuce, I. Age and gender related prevalence of intracranial calcifications in CT imaging; data from 12,000 healthy subjects. J. Chem. Neuroanat. 2016, 78, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, M.T.; Oh, C.; Raju, A.; Choudhri, A.F. Physiologic pineal region, choroid plexus, and dural calcifications in the first decade of life. AJNR Am. J. Neuroradiol. 2015, 36, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Lilja, B. The tentorial pressure cone, its significance and its diagnosis through dislocation of the calcified pineal body. Acta Radiol. 1948, 30, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Goree, J.A.; Wallace, K.K.; Bean, R.L. The pineal tomogram. Visualization of the faintly calcified pineal gland. Am. J. Roentgenol. Radium Ther. Nucl. Med. 1963, 89, 1209–1211. [Google Scholar] [PubMed]
- Drexler, J.; Meaney, T.F.; McCormack, L.J. The calcified pineal body and carcinoma. Clevel. Clin. Q. 1957, 24, 242–247. [Google Scholar] [CrossRef]
- Drexler, J. The calcified pineal body and mammary carcinoma. Cancer 1963, 16, 1616–1617. [Google Scholar] [CrossRef]
- Sandyk, R. The relationship of thought disorder to third ventricle width and calcification of the pineal gland in chronic schizophrenia. Int. J. Neurosci. 1993, 68, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Sandyk, R. Pineal calcification in relation to menopause in schizophrenia. Int. J. Neurosci. 1992, 67, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sandyk, R.; Kay, S.R. Abnormal EEG and calcification of the pineal gland in schizophrenia. Int. J. Neurosci. 1992, 62, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Bersani, G.; Garavini, A.; Taddei, I.; Tanfani, G.; Nordio, M.; Pancheri, P. Computed tomography study of pineal calcification in schizophrenia. Eur. Psychiatry 1999, 14, 163–166. [Google Scholar] [CrossRef]
- Sandyk, R.; Kay, S.R. Pineal calcification in schizophrenia. Relationship to age of onset and tardive dyskinesia. Schizophr. Res. 1991, 5, 85–86. [Google Scholar] [CrossRef]
- Doyle, A.J.; Anderson, G.D. Physiologic Calcification of the Pineal Gland in Children on Computed Tomography: Prevalence, Observer Reliability and Association with Choroid Plexus Calcification. Acad. Radiol. 2006, 13, 822–826. [Google Scholar] [CrossRef] [PubMed]
- Bojkowski, C.J.; Arendt, J. Factors influencing urinary 6-sulphatoxymelatonin, a major melatonin metabolite, in normal human subjects. Clin. Endocrinol. 1990, 33, 435–444. [Google Scholar] [CrossRef]
- Commentz, J.C.; Fischer, P.; Stegner, H.; Winkler, P.; Helmake, K.; Willig, R.P. Pineal calcification does not affect melatonin production. J. Neural Transm. Suppl. 1986, 21, 481–502. [Google Scholar]
- Kay, S.R.; Sandyk, R. Experimental models of schizophrenia. Int. J. Neurosci. 1991, 58, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Friedland, R.P.; Luxenberg, J.S.; Koss, E. A quantitative study of intracranial calcification in dementia of the Alzheimer type. Int. Psychogeriatr. 1990, 2, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Turgut, A.T.; Karakaş, H.M.; Ozsunar, Y.; Altın, L.; Ceken, K.; Alıcıoğlu, B.; Sönmez, I.; Alparslan, A.; Yürümez, B.; Celik, T.; et al. Age-related changes in the incidence of pineal gland calcification in Turkey: A prospective multicenter CT study. Pathophysiology 2008, 15, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Tessonneaud, A.; Locatelli, A.; Caldani, M.; Viguier-Martinez, M.C. Bilateral lesions of the suprachiasmatic nuclei alter the nocturnal melatonin secretion in sheep. J. Neuroendocrinol. 1995, 7, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.J.; Jansen, H.T.; Kao, C.C.; Kuehl, D.E.; Jackson, G.L. Disruption of reproductive rhythms and patterns of melatonin and prolactin secretion following bilateral lesions of the suprachiasmatic nuclei in the ewe. J. Neuroendocrinol. 1995, 7, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Bittman, E.L.; Crandell, R.G.; Lehman, M.N. Influences of the paraventricular and suprachiasmatic nuclei and olfactory bulbs on melatonin responses in the golden hamster. Biol. Reprod. 1989, 40, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Rudeen, P.K.; Banks, A.F.; Rollag, M.D. Acute effects of unilateral or bilateral superior cervical ganglionectomy on rat pineal N-acetyltransferase activity and melatonin content. Experientia 1979, 35, 691–692. [Google Scholar] [CrossRef] [PubMed]
- Saboureau, M.; Vivien-Roels, B.; Pevet, P. Pineal melatonin concentrations during day and night in the adult hedgehog: Effect of a light pulse at night and superior cervical ganglionectomy. J. Pineal Res. 1991, 11, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Gillette, M.U.; Tischkau, S.A. Suprachiasmatic nucleus: The brain’s circadian clock. Recent Prog. Horm. Res. 1999, 54, 33–58. [Google Scholar] [PubMed]
- McArthur, A.J.; Gillette, M.U.; Prosser, R.A. Melatonin directly resets the rat suprachiasmatic circadian clock in vitro. Brain Res. 1991, 565, 158–161. [Google Scholar] [CrossRef]
- Waly, N.E.; Hallworth, R. Circadian Pattern of Melatonin MT1 and MT2 Receptor Localization in the Rat Suprachiasmatic Nucleus. J. Circadian Rhythms 2015, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kandalepas, P.C.; Mitchell, J.W.; Gillette, M.U. Melatonin signal transduction pathways require e-box-mediated transcription of Per1 and Per2 to reset the SCN clock at dusk. PLoS ONE 2016, 11, e0157824. [Google Scholar] [CrossRef] [PubMed]
- Gauer, F.; Masson-Pévet, M.; Skene, D.J.; Vivien-Roels, B.; Pévet, P. Daily rhythms of melatonin binding sites in the rat pars tuberalis and suprachiasmatic nuclei; evidence for a regulation of melatonin receptors by melatonin itself. Neuroendocrinology 1993, 57, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Hastings, M.H.; Herbert, J.; Martensz, N.D.; Roberts, A.C. Annual reproductive rhythms in mammals: Mechanisms of light synchronization. Ann. N. Y. Acad. Sci. 1985, 453, 182–204. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Isaki, K.; Sack, R.L.; Lewy, A.J. Free-running melatonin, sleep propensity, cortisol and temperature rhythms in a totally blind person. Jpn. J. Psychiatry Neurol. 1992, 46, 210–212. [Google Scholar] [CrossRef] [PubMed]
- Palm, L.; Blennow, G.; Wetterberg, L. Correction of non-24-hour sleep/wake cycle by melatonin in a blind retarded boy. Ann. Neurol. 1991, 29, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J.; Newsome, D.A. Different types of melatonin circadian secretory rhythms in some blind subjects. J. Clin. Endocrinol. Metab. 1983, 56, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Sack, R.L.; Lewy, A.J.; Blood, M.L.; Stevenson, J.; Keith, L.D. Melatonin administration to blind people: Phase advances and entrainment. J. Biol. Rhythms 1991, 6, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Lockley, S.W.; Skene, D.J.; James, K.; Thapan, K.; Wright, J.; Arendt, J. Melatonin administration can entrain the free-running circadian system of blind subjects. J. Endocrinol. 2000, 164, R1–R6. [Google Scholar] [CrossRef] [PubMed]
- Ibañez Rodriguez, M.P.; Noctor, S.C.; Muñoz, E.M. Cellular basis of pineal gland development: Emerging role of microglia as phenotype regulator. PLoS ONE 2016, 11, e0167063. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Manchester, L.C.; Qin, L.; Reiter, R.J. Melatonin: A mitochondrial targeting molecule involving mitochondrial protection and dynamics. Int. J. Mol. Sci. 2016, 17, 2124. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Manchester, L.C.; Liu, X.; Rosales-Corral, S.A.; Acuna-Castroviejo, D.; Reiter, R.J. Mitochondria and chloroplasts as the original sites of melatonin synthesis: A hypothesis related to melatonin’s primary function and evolution in eukaryotes. J. Pineal Res. 2013, 54, 127–138. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, J.; Zhang, Z.; Yang, M.; Li, Y.; Tian, X.; Ma, T.; Tao, J.; Zhu, K.; Song, Y.; et al. Mitochondria Synthesize Melatonin to Ameliorate Its Function and Improve Mice Oocyte’s Quality under in Vitro Conditions. Int. J. Mol. Sci. 2016, 17, 939. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, C.; Zheng, X.; Guo, Y.; Zhou, F.; Shan, D.; Liu, X.; Kong, J. Plant mitochondria synthesize melatonin and enhance the tolerance of plants to drought stress. J. Pineal Res. 2017, 63, e12429. [Google Scholar] [CrossRef] [PubMed]
- Krakowski, G.; Cieciura, L. Ultrastructural studies on the pinealocyte mitochondria during the daytime and at night. J. Pineal Res. 1985, 2, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Casado, M.E.; Rusanova, I.; Aranda, P.; Fernández-Ortiz, M.; Sayed, R.K.A.; Fernández-Gil, B.I.; Hidalgo-Gutiérrez, A.; Escames, G.; López, L.C.; Acuña-Castroviejo, D. In Vivo Determination of Mitochondrial respiration in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated zebrafish reveals the efficacy of melatonin in restoring mitochondrial normalcy. Zebrafish 2017. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Wang, Z.; Chen, Z.; Jin, H.; Chen, C.; Chai, S.; Lv, H.; Yang, L.; Hu, Y.; Dong, R.; et al. Melatonin attenuates chronic cough mediated by oxidative stress via transient receptor potential melastatin-2 in guinea pigs exposed to particulate matter 2.5. Physiol. Res. Available online: https://www.bioportfolio.com/resources/pmarticle/1932561/Melatonin-attenuates-chronic-cough-mediated-by-oxidative-stress-via-transient-receptor-potential.html (accessed on 30 January 2018).
- Parameyong, A.; Charngkaew, K.; Govitrapong, P.; Chetsawang, B. Melatonin attenuates methamphetamine-induced disturbances in mitochondrial dynamics and degeneration in neuroblastoma SH-SY5Y cells. J. Pineal Res. 2013, 55, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Suwanjang, W.; Abramov, A.Y.; Charngkaew, K.; Govitrapong, P.; Chetsawang, B. Melatonin prevents cytosolic calcium overload, mitochondrial damage and cell death due to toxically high doses of dexamethasone-induced oxidative stress in human neuroblastoma SH-SY5Y cells. Neurochem. Int. 2016, 97, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Ning, J.; Feng, N.; Li, Z.; Liu, Z.; Wang, Y.; Wang, Y.; Li, X.; Huo, C.; Jia, X.; et al. Dynamin-related protein 1-mediated mitochondrial fission contributes to post-traumatic cardiac dysfunction in rats and the protective effect of melatonin. J. Pineal Res. 2017, e12447. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Du, W.; Li, Y.; Shi, C.; Hu, N.; Ma, S.; Wang, W.; Ren, J. Effects of melatonin on fatty liver disease: The role of NR4A1/DNA-PKcs/p53 pathway, mitochondrial fission, and mitophagy. J. Pineal Res. 2017, 64, e12450. [Google Scholar] [CrossRef] [PubMed]
- Chuang, J.-I.; Pan, I.-L.; Hsieh, C.-Y.; Huang, C.-Y.; Chen, P.-C.; Shin, J.W. Melatonin prevents the dynamin-related protein 1-dependent mitochondrial fission and oxidative insult in the cortical neurons after 1-methyl-4-phenylpyridinium treatment. J. Pineal Res. 2016, 61, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Parameyong, A.; Govitrapong, P.; Chetsawang, B. Melatonin attenuates the mitochondrial translocation of mitochondrial fission proteins and Bax, cytosolic calcium overload and cell death in methamphetamine-induced toxicity in neuroblastoma SH-SY5Y cells. Mitochondrion 2015, 24, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Pi, H.; Zhang, L.; Zhang, N.; Li, Y.; Zhang, H.; Tang, J.; Li, H.; Feng, M.; Deng, P.; et al. Melatonin prevents abnormal mitochondrial dynamics resulting from the neurotoxicity of cadmium by blocking calcium-dependent translocation of Drp1 to the mitochondria. J. Pineal Res. 2016, 60, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, L.; Li, Y.; Gao, J. Melatonin increases human cervical cancer HeLa cells apoptosis induced by cisplatin via inhibition of JNK/Parkin/mitophagy axis. In Vitro Cell. Dev. Biol. Anim. 2018, 54, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, D.; Zhu, P.; Hu, S.; Hu, N.; Ma, S.; Zhang, Y.; Han, T.; Ren, J.; Cao, F.; et al. Melatonin suppresses platelet activation and function against cardiac ischemia/reperfusion injury via PPARγ/FUNDC1/mitophagy pathways. J. Pineal Res. 2017, 63, e12438. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhang, Y.; Hu, S.; Shi, C.; Zhu, P.; Ma, Q.; Jin, Q.; Cao, F.; Tian, F.; Chen, Y. Melatonin protects cardiac microvasculature against ischemia/reperfusion injury via suppression of mitochondrial fission-VDAC1-HK2-mPTP-mitophagy axis. J. Pineal Res. 2017, 63, e12413. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Shrestha, S.; Li, J.; Yu, X.; Chen, J.; Yan, F.; Ying, G.; Gu, C.; Wang, L.; Chen, G. Melatonin-mediated mitophagy protects against early brain injury after subarachnoid hemorrhage through inhibition of NLRP3 inflammasome activation. Sci. Rep. 2017, 7, 2417. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Domínguez, N.; Ordóñez, R.; Fernández, A.; Méndez-Blanco, C.; Baulies, A.; Garcia-Ruiz, C.; Fernández-Checa, J.C.; Mauriz, J.L.; González-Gallego, J. Melatonin-induced increase in sensitivity of human hepatocellular carcinoma cells to sorafenib is associated with reactive oxygen species production and mitophagy. J. Pineal Res. 2016, 61, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-W.; Hong, J.-M.; Lee, S.-M. Melatonin enhances mitophagy and mitochondrial biogenesis in rats with carbon tetrachloride-induced liver fibrosis. J. Pineal Res. 2016, 60, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Chao, H.; Li, Z.; Xu, X.; Liu, Y.; Hou, L.; Liu, N.; Ji, J. Melatonin attenuates traumatic brain injury-induced inflammation: A possible role for mitophagy. J. Pineal Res. 2016, 61, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Coto-Montes, A.; Boga, J.A.; Rosales-Corral, S.; Fuentes-Broto, L.; Tan, D.-X.; Reiter, R.J. Role of melatonin in the regulation of autophagy and mitophagy: A review. Mol. Cell. Endocrinol. 2012, 361, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Domínguez, N.; Méndez-Blanco, C.; Carbajo-Pescador, S.; Fondevila, F.; García-Palomo, A.; González-Gallego, J.; Mauriz, J.L. Melatonin enhances sorafenib actions in human hepatocarcinoma cells by inhibiting mTORC1/p70S6K/HIF-1α and hypoxia-mediated mitophagy. Oncotarget 2017, 8, 91402–91414. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-J.; Zhuang, J.; Zhu, H.-Y.; Shen, Y.-X.; Tan, Z.-L.; Zhou, J.-N. Cultured rat cortical astrocytes synthesize melatonin: Absence of a diurnal rhythm. J. Pineal Res. 2007, 43, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Markus, R.P.; Fernandes, P.A.; Kinker, G.S.; da Silveira Cruz-Machado, S.; Marçola, M. Immune-Pineal Axis—Acute inflammatory responses coordinate melatonin synthesis by pinealocytes and phagocytes. Br. J. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Silva, S.; Ximenes, V.F.; Livramento, J.A.; Catalani, L.H.; Campa, A. High concentrations of the melatonin metabolite, N1-acetyl-N2-formyl-5-methoxykynuramine, in cerebrospinal fluid of patients with meningitis: A possible immunomodulatory mechanism. J. Pineal Res. 2005, 39, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira Cruz-Machado, S.; Pinato, L.; Tamura, E.K.; Carvalho-Sousa, C.E.; Markus, R.P. Glia-pinealocyte network: The paracrine modulation of melatonin synthesis by tumor necrosis factor (TNF). PLoS ONE 2012, 7, e40142. [Google Scholar] [CrossRef] [PubMed]
- Villela, D.; Atherino, V.F.; de Lima, L.S.; Moutinho, A.A.; do Amaral, F.G.; Peres, R.; Martins de Lima, T.; da Torrão, A.S.; Cipolla-Neto, J.; Scavone, C.; et al. Modulation of pineal melatonin synthesis by glutamate involves paracrine interactions between pinealocytes and astrocytes through NF-κB activation. BioMed Res. Int. 2013, 2013, 618432. [Google Scholar] [CrossRef] [PubMed]
- Souza-Teodoro, L.H.; Dargenio-Garcia, L.; Petrilli-Lapa, C.L.; da Souza, E.S.; Fernandes, P.A.C.M.; Markus, R.P.; Ferreira, Z.S. Adenosine triphosphate inhibits melatonin synthesis in the rat pineal gland. J. Pineal Res. 2016, 60, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Welsh, M.G. CSF-contacting pinealocytes in the pineal recess of the Mongolian gerbil: A correlative scanning and transmission electron microscope study. Dev. Dyn. 1983, 166, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Krstić, R. Scanning electron microscope observations of the canaliculi in the rat pineal gland. Experientia 1975, 31, 1072–1074. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Kim, S.J.; Cruz, M.H.C. Delivery of pineal melatonin to the brain and SCN: Role of canaliculi, cerebrospinal fluid, tanycytes and Virchow-Robin perivascular spaces. Brain Struct. Funct. 2014, 219, 1873–1887. [Google Scholar] [CrossRef] [PubMed]
- Krstić, R.V. Scanning electron microscopic study of the freeze-fractured pineal body of the rat. Cell Tissue Res. 1979, 201, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Quay, W.B. Pineal canaliculi: Demonstration, twenty-four-hour rhythmicity and experimental modification. Dev. Dyn. 1974, 139, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Tricoire, H.; Locatelli, A.; Chemineau, P.; Malpaux, B. Melatonin enters the cerebrospinal fluid through the pineal recess. Endocrinology 2002, 143, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Tricoire, H.; Møller, M.; Chemineau, P.; Malpaux, B. Origin of cerebrospinal fluid melatonin and possible function in the integration of photoperiod. Reprod. Suppl. 2003, 61, 311–321. [Google Scholar] [PubMed]
- Legros, C.; Chesneau, D.; Boutin, J.A.; Barc, C.; Malpaux, B. Melatonin from cerebrospinal fluid but not from blood reaches sheep cerebral tissues under physiological conditions. J. Neuroendocrinol. 2014, 26, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Sanchez-Barcelo, E.; Mediavilla, M.D.; Reiter, R.J. Significance of high levels of endogenous melatonin in Mammalian cerebrospinal fluid and in the central nervous system. Curr. Neuropharmacol. 2010, 8, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Leston, J.; Harthé, C.; Mottolese, C.; Mertens, P.; Sindou, M.; Claustrat, B. Is pineal melatonin released in the third ventricle in humans? A study in movement disorders. Neurochirurgie 2015, 61, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X. Role of CSF in the transport of melatonin. J. Pineal Res. 2002, 33, 61. [Google Scholar] [CrossRef] [PubMed]
- Paredes, S.D.; Korkmaz, A.; Manchester, L.C.; Tan, D.X.; Reiter, R.J. Phytomelatonin: A review. J. Exp. Bot. 2009, 60, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tan, D.X.; Lei, Q.; Chen, H.; Wang, L.; Li, Q.; Gao, Y.; Kong, J. Melatonin and its potential biological functions in the fruits of sweet cherry. J. Pineal Res. 2013, 55, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Wang, L.; Tan, D.X.; Zhao, Y.; Zheng, X.-D.; Chen, H.; Li, Q.; Zuo, B.; Kong, J. Identification of genes for melatonin synthetic enzymes in “Red Fuji” apple (Malus domestica Borkh cv. Red) and their expression and melatonin production during fruit development. J. Pineal Res. 2013, 55, 443–451. [Google Scholar] [PubMed]
- Byeon, Y.; Back, K. Low melatonin production by suppression of either serotonin N-acetyltransferase or N-acetylserotonin methyltransferase in rice causes seedling growth retardation with yield penalty, abiotic stress susceptibility, and enhanced coleoptile growth under anoxic conditions. J. Pineal Res. 2016, 60, 348–359. [Google Scholar] [PubMed]
- Ramakrishna, A.; Giridhar, P.; Sankar, K.U.; Ravishankar, G.A. Melatonin and serotonin profiles in beans of Coffea species. J. Pineal Res. 2012, 52, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Zanghi, B.M.; Manchester, L.C.; Reiter, R.J. Melatonin identified in meats and other food stuffs: Potentially nutritional impact. J. Pineal Res. 2014, 57, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar] [PubMed]
- Nagata, C.; Nagao, Y.; Shibuya, C.; Kashiki, Y.; Shimizu, H. Association of vegetable intake with urinary 6-sulfatoxymelatonin level. Cancer Epidemiol. Prev. Biomark. 2005, 14, 1333–1335. [Google Scholar] [CrossRef] [PubMed]
- Oba, S.; Nakamura, K.; Sahashi, Y.; Hattori, A.; Nagata, C. Consumption of vegetables alters morning urinary 6-sulfatoxymelatonin concentration. J. Pineal Res. 2008, 45, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Howatson, G.; Bell, P.G.; Tallent, J.; Middleton, B.; McHugh, M.P.; Ellis, J. Effect of tart cherry juice (Prunus cerasus) on melatonin levels and enhanced sleep quality. Eur. J. Nutr. 2012, 51, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Garrido, M.; Paredes, S.D.; Cubero, J.; Lozano, M.; Toribio-Delgado, A.F.; Munoz, J.L.; Reiter, R.J.; Barriga, C.; Rodriguez, A.B. Jerte valley cherry-enriched diets improve nocturnal rest and increase 6-sulfatoxymelatonin and total antioxidant capacity in the urine of middle-aged and elderly humans. J. Gerontol. Ser. A 2010, 65A, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Kilic, E.; Ozdemir, Y.G.; Bolay, H.; Keleştimur, H.; Dalkara, T. Pinealectomy aggravates and melatonin administration attenuates brain damage in focal ischemia. J. Cereb. Blood Flow Metab. 1999, 19, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Sainz, R.M.; Reiter, R.J.; Mayo, J.C.; Cabrera, J.; Tan, D.X.; Qi, W.; Garcia, J.J. Changes in lipid peroxidation during pregnancy and after delivery in rats: Effect of pinealectomy. J. Reprod. Fertil. 2000, 119, 143–149. [Google Scholar] [CrossRef] [PubMed]
- De Butte, M.; Fortin, T.; Pappas, B.A. Pinealectomy: Behavioral and neuropathological consequences in a chronic cerebral hypoperfusion model. Neurobiol. Aging 2002, 23, 309–317. [Google Scholar] [CrossRef]
- Ates, O.; Cayli, S.; Gurses, I.; Yucel, N.; Iraz, M.; Altinoz, E.; Kocak, A.; Yologlu, S. Effect of pinealectomy and melatonin replacement on morphological and biochemical recovery after traumatic brain injury. Int. J. Dev. Neurosci. 2006, 24, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Borlongan, C.V.; Sumaya, I.; Moss, D.; Kumazaki, M.; Sakurai, T.; Hida, H.; Nishino, H. Melatonin-secreting pineal gland: A novel tissue source for neural transplantation therapy in stroke. Cell Transplant. 2003, 12, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Del Rio-Hortega, P. Cytology and cellular pathology of the nervous system. In Pineal Gland; Penfield, W., Ed.; Hoeber: New York, NY, USA, 1932; pp. 637–703. [Google Scholar]
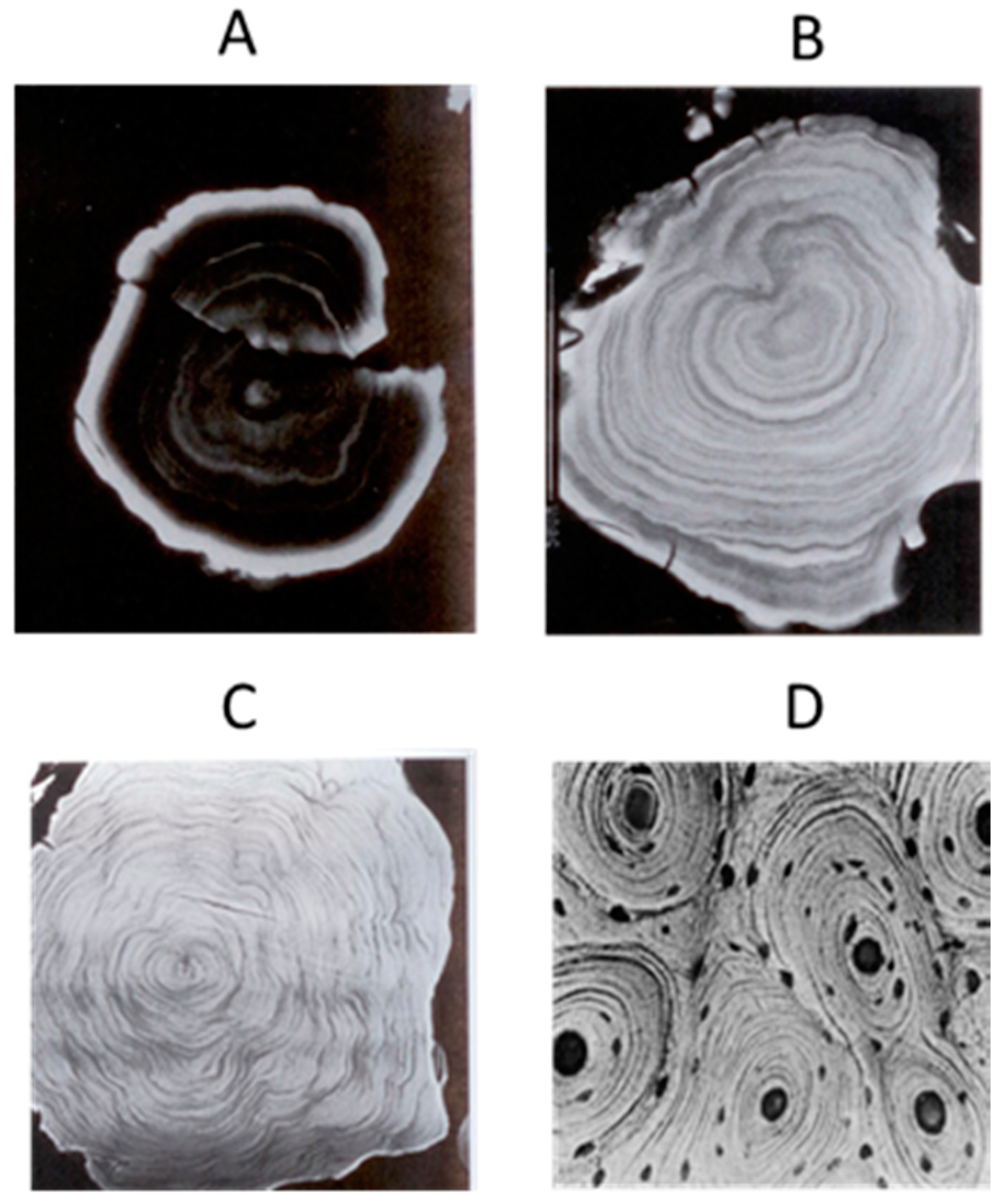
- Welsh, M.G. Pineal calcification: Structural and functional aspects. Pineal Res. Rev. 1985, 3, 41–68. [Google Scholar]
- Vígh, B.; Szél, A.; Debreceni, K.; Fejér, Z.; Manzano e Silva, M.J.; Vígh-Teichmann, I. Comparative histology of pineal calcification. Histol. Histopathol. 1998, 13, 851–870. [Google Scholar] [PubMed]
- Japha, J.L.; Eder, T.J.; Goldsmith, E.D. Calcified inclusions in the superficial pineal gland of the mongolian gerbil, Meriones unguiculatus. Acta Anat. 1976, 94, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Przybylska-Gornowicz, B.; Lewczuk, B.; Prusik, M.; Bulc, M. Pineal concretions in turkey (Meleagris gallopavo) as a result of collagen-mediated calcification. Histol. Histopathol. 2009, 24, 407–415. [Google Scholar] [PubMed]
- Maślińska, D.; Laure-Kamionowska, M.; Deręgowski, K.; Maśliński, S. Association of mast cells with calcification in the human pineal gland. Folia Neuropathol. 2010, 48, 276–282. [Google Scholar] [PubMed]
- Winkler, P.; Helmke, K. Age-related incidence of pineal gland calcification in children: A roentgenological study of 1044 skull films and a review of the literature. J. Pineal Res. 1987, 4, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Champney, T.H.; Joshi, B.N.; Vaughan, M.K.; Reiter, R.J. Superior cervical ganglionectomy results in the loss of pineal concretions in the adult male gerbil (Meriones unguiculatus). Anat. Rec. 1985, 211, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Welsh, M.G.; Vaughan, M.K. Age-related changes in the intact and sympathetically denervated gerbil pineal gland. Am. J. Anat. 1976, 146, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Lewinski, A.; Waughan, M.K.; Champney, T.H.; Reiter, R.J.; Smith, N.K.R. Dark-exposure increases the number of pineal concretions in male gerbils (Meriones unguiculatus). IRCS Med. Sci. 1983, 11, 977–978. [Google Scholar]
- Welsh, M.G. Effects of superior cervical gangalinectomy, constant light and bliding on the gerbil pineal gland: An ultrastructural analysis. Anat. Rec. 1977, 187, 746. [Google Scholar]
- Sandyk, R. The association of pineal calcification with drug-induced dystonic movements. Int. J. Neurosci. 1990, 53, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Sandyk, R. Pineal calcification and subtypes of tardive dyskinesia. Int. J. Neurosci. 1990, 53, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Sandyk, R.; Kay, S.R. The relationship of pineal calcification and melatonin secretion to the pathophysiology of tardive dyskinesia and Tourette’s syndrome. Int. J. Neurosci. 1991, 58, 215–247. [Google Scholar] [CrossRef] [PubMed]
- Hinterberger, H.; Pickering, J. Catecholamine, indolealkylamine and calcium levels of human pineal glands in various clinical conditions. Pathology 1976, 8, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Mahlberg, R.; Walther, S.; Kalus, P.; Bohner, G.; Haedel, S.; Reischies, F.M.; Kuhl, K.P.; Hellweg, R.; Kunz, D. Pineal calcification in Alzheimer’s disease: An in vivo study using computed tomography. Neurobiol. Aging 2008, 29, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Berkiks, I.; Benmhammed, H.; Mesfioui, A.; Ouichou, A.; El Hasnaoui, A.; Mouden, S.; Touil, T.; Bahbiti, Y.; Nakache, R.; El Hessni, A. Postnatal melatonin treatment protects against affective disorders induced by early-life immune stimulation by reducing the microglia cell activation and oxidative stress. Int. J. Neurosci. 2017, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Chen, L.; Poeggeler, B.; Manchester, L.C.; Reiter, R.J. Melatonin: A potent, endogenous hydroxyl radical scavenger. Endocr. J. 1993, 1, 57–63. [Google Scholar]
- Liu, Z.; Gan, L.; Xu, Y.; Luo, D.; Ren, Q.; Wu, S.; Sun, C. Melatonin alleviates inflammasome-induced pyroptosis through inhibiting NF-κB/GSDMD signal in mice adipose tissue. J. Pineal Res. 2017, 63, e12414. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ahmed, M.M.; Jiang, Q.; Lu, N.; Tan, C.; Gao, Y.; Mahmood, Q.; Chen, D.; Fukunaga, K.; Li, M.; et al. Melatonin ameliorates hypoglycemic stress-induced brain endothelial tight junction injury by inhibiting protein nitration of TP53-induced glycolysis and apoptosis regulator. J. Pineal Res. 2017, 63, e12440. [Google Scholar] [CrossRef] [PubMed]
- Leeboonngam, T.; Pramong, R.; Sae-Ung, K.; Govitrapong, P.; Phansuwan-Pujito, P. Neuroprotective effects of melatonin on amphetamine-induced dopaminergic fiber degeneration in the hippocampus of postnatal rats. J. Pineal Res. 2017, e12456. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-N.; Liu, R.-Y.; Kamphorst, W.; Hofman, M.A.; Swaab, D.F. Early neuropathological Alzheimer’s changes in aged individuals are accompanied by decreased cerebrospinal fluid melatonin levels. J. Pineal Res. 2003, 35, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.Y.; Zhou, J.N.; van Heerikhuize, J.; Hofman, M.A.; Swaab, D.F. Decreased melatonin levels in postmortem cerebrospinal fluid in relation to aging, Alzheimer’s disease, and apolipoprotein E-epsilon4/4 genotype. J. Clin. Endocrinol. Metab. 1999, 84, 323–327. [Google Scholar] [PubMed]
- Lahiri, D.K. Melatonin affects the metabolism of the beta-amyloid precursor protein in different cell types. J. Pineal Res. 1999, 26, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Pappolla, M.; Bozner, P.; Soto, C.; Shao, H.; Robakis, N.K.; Zagorski, M.; Frangione, B.; Ghiso, J. Inhibition of Alzheimer beta-fibrillogenesis by melatonin. J. Biol. Chem. 1998, 273, 7185–7188. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Xu, G.; Duan, P.; Zhang, Q.; Wang, J. Effects of melatonin on wortmannin-induced tau hyperphosphorylation. Acta Pharmacol. Sin. 2005, 26, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-C.; Zhang, J.; Yu, X.; Han, L.; Zhou, Z.-T.; Zhang, Y.; Wang, J.-Z. Prevention of isoproterenol-induced tau hyperphosphorylation by melatonin in the rat. Sheng Li Xue Bao 2005, 57, 7–12. [Google Scholar] [PubMed]
- Li, X.-C.; Wang, Z.-F.; Zhang, J.-X.; Wang, Q.; Wang, J.-Z. Effect of melatonin on calyculin A-induced tau hyperphosphorylation. Eur. J. Pharmacol. 2005, 510, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Wade, A.G.; Farmer, M.; Harari, G.; Fund, N.; Laudon, M.; Nir, T.; Frydman-Marom, A.; Zisapel, N. Add-on prolonged-release melatonin for cognitive function and sleep in mild to moderate Alzheimer’s disease: A 6-month, randomized, placebo-controlled, multicenter trial. Clin. Interv. Aging 2014, 18, 947–961. [Google Scholar]
- Asayama, K.; Yamadera, H.; Ito, T.; Suzuki, H.; Kudo, Y.; Endo, S. Double blind study of melatonin effects on the sleep-wake rhythm, cognitive and non-cognitive functions in Alzheimer type dementia. J. Nippon Med. Sch. 2003, 70, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P.; Brusco, L.I.; Liberczuk, C.; Furio, A.M. The use of melatonin in Alzheimer’s disease. Neuro Endocrinol. Lett. 2002, 23 (Suppl. 1), 20–23. [Google Scholar] [PubMed]
- Brusco, L.I.; Márquez, M.; Cardinali, D.P. Monozygotic twins with Alzheimer’s disease treated with melatonin: Case report. J. Pineal Res. 1998, 25, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Tractenberg, R.E.; Singer, C.M.; Cummings, J.L.; Thal, L.J. The Sleep Disorders Inventory: An instrument for studies of sleep disturbance in persons with Alzheimer’s disease. J. Sleep Res. 2003, 12, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Singer, C.; Tractenberg, R.E.; Kaye, J.; Schafer, K.; Gamst, A.; Grundman, M.; Thomas, R.; Thal, L.J. A multicenter, placebo-controlled trial of melatonin for sleep disturbance in Alzheimer’s disease. Sleep 2003, 26, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Mahlberg, R.; Kunz, D.; Sutej, I.; Kühl, K.-P.; Hellweg, R. Melatonin treatment of day-night rhythm disturbances and sundowning in Alzheimer disease: An open-label pilot study using actigraphy. J. Clin. Psychopharmacol. 2004, 24, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Olcese, J.M.; Cao, C.; Mori, T.; Mamcarz, M.B.; Maxwell, A.; Runfeldt, M.J.; Wang, L.; Zhang, C.; Lin, X.; Zhang, G.; et al. Protection against cognitive deficits and markers of neurodegeneration by long-term oral administration of melatonin in a transgenic model of Alzheimer disease. J. Pineal Res. 2009, 47, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.; Wei, G.; Peng, S.; Qu, Z.; Yang, Y.; Yang, Q.; Huang, X.; Liu, J.; Zhuang, Z.; Yang, X. Melatonin ameliorates anxiety and depression-like behaviors and modulates proteomic changes in triple transgenic mice of Alzheimer’s disease. Biofactors 2017, 43, 593–611. [Google Scholar] [CrossRef] [PubMed]
- Baño Otalora, B.; Popovic, N.; Gambini, J.; Popovic, M.; Viña, J.; Bonet-Costa, V.; Reiter, R.J.; Camello, P.J.; Rol, M.Á.; Madrid, J.A. Circadian system functionality, hippocampal oxidative stress, and spatial memory in the APPswe/PS1dE9 transgenic model of Alzheimer disease: Effects of melatonin or ramelteon. Chronobiol. Int. 2012, 29, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Dragicevic, N.; Copes, N.; O’Neal-Moffitt, G.; Jin, J.; Buzzeo, R.; Mamcarz, M.; Tan, J.; Cao, C.; Olcese, J.M.; Arendash, G.W.; et al. Melatonin treatment restores mitochondrial function in Alzheimer’s mice: A mitochondrial protective role of melatonin membrane receptor signaling. J. Pineal Res. 2011, 51, 75–86. [Google Scholar] [CrossRef] [PubMed]
- García-Mesa, Y.; Giménez-Llort, L.; López, L.C.; Venegas, C.; Cristòfol, R.; Escames, G.; Acuña-Castroviejo, D.; Sanfeliu, C. Melatonin plus physical exercise are highly neuroprotective in the 3xTg-AD mouse. Neurobiol. Aging 2012, 33, 1124.e13–1124.e29. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, C.; Reverte, I.; Colomina, M.T.; Domingo, J.L.; Gómez, M. Chronic exposure to aluminum and melatonin through the diet: Neurobehavioral effects in a transgenic mouse model of Alzheimer disease. Food Chem. Toxicol. 2014, 69, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Stefanova, N.A.; Maksimova, K.Y.; Kiseleva, E.; Rudnitskaya, E.A.; Muraleva, N.A.; Kolosova, N.G. Melatonin attenuates impairments of structural hippocampal neuroplasticity in OXYS rats during active progression of Alzheimer’s disease-like pathology. J. Pineal Res. 2015, 59, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Perepechaeva, M.L.; Stefanova, N.A.; Grishanova, A.Y. Expression of genes for AhR and Nrf2 signal pathways in the retina of OXYS rats during the development of retinopathy and melatonin-induced changes in this process. Bull. Exp. Biol. Med. 2014, 157, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Rudnitskaya, E.A.; Maksimova, K.Y.; Muraleva, N.A.; Logvinov, S.V.; Yanshole, L.V.; Kolosova, N.G.; Stefanova, N.A. Beneficial effects of melatonin in a rat model of sporadic Alzheimer’s disease. Biogerontology 2015, 16, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Rudnitskaya, E.A.; Muraleva, N.A.; Maksimova, K.Y.; Kiseleva, E.; Kolosova, N.G.; Stefanova, N.A. Melatonin attenuates memory impairment, amyloid-β accumulation, and neurodegeneration in a rat model of sporadic alzheimer’s disease. J. Alzheimers Dis. 2015, 47, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Gehrman, P.R.; Connor, D.J.; Martin, J.L.; Shochat, T.; Corey-Bloom, J.; Ancoli-Israel, S. Melatonin fails to improve sleep or agitation in double-blind randomized placebo-controlled trial of institutionalized patients with alzheimer disease. Am. J. Geriatr. Psychiatry 2009, 17, 166–169. [Google Scholar] [CrossRef] [PubMed]
- García, T.; Ribes, D.; Colomina, M.T.; Cabré, M.; Domingo, J.L.; Gómez, M. Evaluation of the protective role of melatonin on the behavioral effects of aluminum in a mouse model of Alzheimer’s disease. Toxicology 2009, 265, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Casado, M.E.; Lima, E.; García, J.A.; Doerrier, C.; Aranda, P.; Sayed, R.K.; Guerra-Librero, A.; Escames, G.; López, L.C.; Acuña-Castroviejo, D. Melatonin rescues zebrafish embryos from the parkinsonian phenotype restoring the parkin/PINK1/DJ-1/MUL1 network. J. Pineal Res. 2016, 61, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, G.G.; Moráles-Sánchez, E.W.; Pacheco-Moisés, F.P.; Jiménez-Gil, F.J.; Macías-Islas, M.A.; Mireles-Ramírez, M.A.; González-Usigli, H. Effect of melatonin administration on cyclooxygenase-2 activity, serum levels of nitric oxide metabolites, lipoperoxides and glutathione peroxidase activity in patients with Parkinson’s disease. Gac. Med. Mex. 2017, 153, S72–S81. [Google Scholar] [PubMed]
- Wang, X.; Sirianni, A.; Pei, Z.; Cormier, K.; Smith, K.; Jiang, J.; Zhou, S.; Wang, H.; Zhao, R.; Yano, H.; et al. The melatonin mt1 receptor axis modulates mutant huntingtin-mediated toxicity. J. Neurosci. 2011, 31, 14496–14507. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, J.; Nthenge-Ngumbau, D.N.; Rajamma, U.; Mohanakumar, K.P. Melatonin protects against behavioural dysfunctions and dendritic spine damage in 3-nitropropionic acid-induced rat model of Huntington’s disease. Behav. Brain Res. 2014, 264, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Weishaupt, J.H.; Bartels, C.; Pölking, E.; Dietrich, J.; Rohde, G.; Poeggeler, B.; Mertens, N.; Sperling, S.; Bohn, M.; Hüther, G.; et al. Reduced oxidative damage in ALS by high-dose enteral melatonin treatment. J. Pineal Res. 2006, 41, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Barcelo, E.J.; Revilla, N.; Mediavilla, M.D.; Martinez-Cue, C.; Reiter, R.J. Clinical uses of melatonin in neurological diseases and mental and behavioural disorders. Curr. Med. Chem. 2017, 24, 3851–3878. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Sánchez, N.; Cruz-Chamorro, I.; Díaz-Sánchez, M.; Sarmiento-Soto, H.; Medrano-Campillo, P.; Martínez-López, A.; Lardone, P.J.; Guerrero, J.M.; Carrillo-Vico, A. Melatonin reduces inflammatory response in peripheral T helper lymphocytes from relapsing-remitting multiple sclerosis patients. J. Pineal Res. 2017, 63, e12442. [Google Scholar] [CrossRef] [PubMed]
- Kashani, I.R.; Rajabi, Z.; Akbari, M.; Hassanzadeh, G.; Mohseni, A.; Eramsadati, M.K.; Rafiee, K.; Beyer, C.; Kipp, M.; Zendedel, A. Protective effects of melatonin against mitochondrial injury in a mouse model of multiple sclerosis. Exp. Brain Res. 2014, 232, 2835–2846. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk-Sowa, M.; Pierzchala, K.; Sowa, P.; Polaniak, R.; Kukla, M.; Hartel, M. Influence of melatonin supplementation on serum antioxidative properties and impact of the quality of life in multiple sclerosis patients. J. Physiol. Pharmacol. 2014, 65, 543–550. [Google Scholar] [PubMed]
- Ghareghani, M.; Scavo, L.; Arnoult, D.; Zibara, K.; Farhadi, N. Melatonin therapy reduces the risk of osteoporosis and normalizes bone formation in multiple sclerosis. Fundam. Clin. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk-Sowa, M.; Sowa, P.; Adamczyk, J.; Niedziela, N.; Misiolek, H.; Owczarek, M.; Zwirska-Korczala, K. Effect of melatonin supplementation on plasma lipid hydroperoxides, homocysteine concentration and chronic fatigue syndrome in multiple sclerosis patients treated with interferons-beta and mitoxantrone. J. Physiol. Pharmacol. 2016, 67, 235–242. [Google Scholar] [PubMed]
- López-González, A.; Álvarez-Sánchez, N.; Lardone, P.J.; Cruz-Chamorro, I.; Martínez-López, A.; Guerrero, J.M.; Reiter, R.J.; Carrillo-Vico, A. Melatonin treatment improves primary progressive multiple sclerosis: A case report. J. Pineal Res. 2015, 58, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Henden, T.; Stokkan, K.A.; Reiter, R.J.; Nonaka, K.O.; Lerchl, A.; Jones, D.J. Age-associated reduction in pineal beta-adrenergic receptor density is prevented by life-long food restriction in rats. Biol. Signals 1992, 1, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Dax, E.M.; Sugden, D. Age-associated changes in pineal adrenergic receptors and melatonin synthesizing enzymes in the Wistar rat. J. Neurochem. 1988, 50, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Paltsev, M.A.; Polyakova, V.O.; Kvetnoy, I.M.; Anderson, G.; Kvetnaia, T.V.; Linkova, N.S.; Paltseva, E.M.; Rubino, R.; De Cosmo, S.; De Cata, A.; et al. Morphofunctional and signaling molecules overlap of the pineal gland and thymus: Role and significance in aging. Oncotarget 2016, 7, 11972–11983. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, K.; Saeki, K.; Kurumatani, N. Association between urinary 6-sulfatoxymelatonin excretion and arterial stiffness in the general elderly population: The HEIJO-KYO cohort. J. Clin. Endocrinol. Metab. 2014, 99, 3233–3239. [Google Scholar] [CrossRef] [PubMed]
- Brazão, V.; Santello, F.H.; Colato, R.P.; Mazotti, T.T.; Tazinafo, L.F.; Toldo, M.P.A.; do Vale, G.T.; Tirapelli, C.R.; do Prado, J.C. Melatonin: Antioxidant and modulatory properties in age-related changes during Trypanosoma cruzi infection. J. Pineal Res. 2017, 63, e12409. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Palacios-Bois, J.; Schwartz, G.; Iskandar, H.; Thakur, M.; Quirion, R.; Nair, N.P. Circadian rhythms of melatonin and cortisol in aging. Biol. Psychiatry 1989, 25, 305–319. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.; Kim, S.J.; Manchester, L.C.; Qi, W.; Garcia, J.J.; Cabrera, J.C.; El-Sokkary, G.; Rouvier-Garay, V. Augmentation of indices of oxidative damage in life-long melatonin-deficient rats. Mech. Ageing Dev. 1999, 110, 157–173. [Google Scholar] [CrossRef]
- Pierpaoli, W.; Regelson, W. Pineal control of aging: Effect of melatonin and pineal grafting on aging mice. Proc. Natl. Acad. Sci. USA 1994, 91, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.C.; Böker, S.M.; Bender, Y.Y.; Diederichs, G.; Fallenberg, E.M.; Wagner, M.; Hamm, B.; Makowski, M.R. Diagnostic accuracy of susceptibility-weighted magnetic resonance imaging for the evaluation of pineal gland calcification. PLoS ONE 2017, 12, e0172764. [Google Scholar] [CrossRef] [PubMed]
- Daghighi, M.H.; Rezaei, V.; Zarrintan, S.; Pourfathi, H. Intracranial physiological calcifications in adults on computed tomography in Tabriz, Iran. Folia Morphol. 2007, 66, 115–119. [Google Scholar]
- Admassie, D.; Mekonnen, A. Incidence of normal pineal and chroids plexus calcification on brain CT (computerized tomography) at Tikur Anbessa Teaching Hospital Addis Ababa, Ethiopia. Ethiop. Med. J. 2009, 47, 55–60. [Google Scholar] [PubMed]
- Fan, K.J. Pineal calcification among black patients. J. Natl. Med. Assoc. 1983. [Google Scholar]
- Daramola, G.F.; Olowu, A.O. Physiological and radiological implications of a low incidence of pineal calcification in Nigeria. Neuroendocrinology 1972, 9, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Tuntapakul, S.; Kitkhuandee, A.; Kanpittaya, J.; Johns, J.; Johns, N.P. Pineal calcification is associated with pediatric primary brain tumor. Asia Pac. J. Clin. Oncol. 2016, 12, e405–e410. [Google Scholar] [CrossRef] [PubMed]
- Kitkhuandee, A.; Sawanyawisuth, K.; Johns, N.P.; Kanpittaya, J.; Johns, J. Pineal calcification is associated with symptomatic cerebral infarction. J. Stroke Cerebrovasc. Dis. 2014, 23, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Kunz, D.; Bes, F.; Schlattmann, P.; Herrmann, W.M. On pineal calcification and its relation to subjective sleep perception: A hypothesis-driven pilot study. Psychiatry Res. 1998, 82, 187–191. [Google Scholar] [CrossRef]
- Sandyk, R.; Pardeshi, R. The relationship between ECT nonresponsiveness and calcification of the pineal gland in bipolar patients. Int. J. Neurosci. 1990, 54, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Liebrich, L.-S.; Schredl, M.; Findeisen, P.; Groden, C.; Bumb, J.M.; Nölte, I.S. Morphology and function: MR pineal volume and melatonin level in human saliva are correlated. J. Magn. Reson. Imaging 2014, 40, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Turgut, M.; Yenisey, C.; Akyüz, O.; Ozsunar, Y.; Erkus, M.; Biçakçi, T. Correlation of serum trace elements and melatonin levels to radiological, biochemical, and histological assessment of degeneration in patients with intervertebral disc herniation. Biol. Trace Elem. Res. 2006, 109, 123–134. [Google Scholar] [CrossRef]
- Sigurdardottir, L.G.; Markt, S.C.; Sigurdsson, S.; Aspelund, T.; Fall, K.; Schernhammer, E.; Rider, J.R.; Launer, L.; Harris, T.; Stampfer, M.J.; et al. Pineal gland volume assessed by mri and its correlation with 6-sulfatoxymelatonin levels among older men. J. Biol. Rhythms 2016, 31, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Fore, S. Evidence of pineal gland calcification on cbct is not insignificant: what else you might discover about your patient. Int. J. Orthod Milwaukee 2016, 27, 11–13. [Google Scholar] [PubMed]
- Mori, R.; Kodaka, T.; Sano, T. Preliminary report on the correlations among pineal concretions, prostatic calculi and age in human adult males. Anat. Sci. Int. 2003, 78, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Swietoslawski, J. The age-related quantitative ultrastructural changes in pinealocytes of gerbils. Neuro. Endocrinol. Lett. 1999, 20, 391–396. [Google Scholar] [PubMed]
- Macpherson, P.; Matheson, M.S. Comparison of calcification of pineal, habenular commissure and choroid plexus on plain films and computed tomography. Neuroradiology 1979, 18, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Schmid, H.A.; Raykhtsaum, G. Age-related differences in the structure of human pineal calcium deposits: Results of transmission electron microscopy and mineralographic microanalysis. J. Pineal Res. 1995, 18, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Quay, W.B. Pineal Chemistry; Charles C. Thomas: Springfield, IL, USA, 1974; pp. 1–430. [Google Scholar]
- Jung, D.; Vollrath, L. Structural dissimilarities in different regions of the pineal gland of Pirbright white guinea-pigs. J. Neural Transm. 1982, 54, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Diehl, B.J. Occurrence and regional distribution of calcareous concretions in the rat pineal gland. Cell Tissue Res. 1978, 195, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Lukaszyk, A.; Reiter, R.J. Morphological basis for polypeptide hormone secretion by the pineal body of macacus rhesus. Endokrynol. Pol. 1975, 26, 603–611. [Google Scholar] [PubMed]
- Chau, Y.P.; Liao, K.K.; Kao, M.H.; Huang, B.N.; Kao, Y.S.; Lu, K.S. Ultrastructure, ZIO-staining and chromaffinity of gerbil pinealocytes. Gaoxiong Yi Xue Ke Xue Za Zhi 1994, 10, 613–623. [Google Scholar] [PubMed]
- Lewczuk, B.; Przybylska, B.; Wyrzykowski, Z. Distribution of calcified concretions and calcium ions in the pig pineal gland. Folia Histochem. Cytobiol. 1994, 32, 243–249. [Google Scholar] [PubMed]
- Krstic, R. Human pineal acervuli: Ultrastructural and X-ray energy dispensive microprobe analysis. Experientia 1976, 32, 800. [Google Scholar]
- Krstić, R. A combined scanning and transmission electron microscopic study and electron probe microanalysis of human pineal acervuli. Cell Tissue Res. 1976, 174, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Krstić, R. Pineal calcification: Its mechanism and significance. J. Neural Transm. Suppl. 1986, 21, 415–432. [Google Scholar] [PubMed]
- Khan, S.R.; Pearle, M.S.; Robertson, W.G.; Gambaro, G.; Canales, B.K.; Doizi, S.; Traxer, O.; Tiselius, H.-G. Kidney stones. Nat. Rev. Dis. Prim. 2016, 2, 16008. [Google Scholar] [CrossRef] [PubMed]
- Galliani, I.; Falcieri, E.; Giangaspero, F.; Valdrè, G.; Mongiorgi, R. A preliminary study of human pineal gland concretions: Structural and chemical analysis. Bollettino Della Societa Italiana di Biologia Sperimentale 1990, 66, 615–622. [Google Scholar] [PubMed]
- Kodaka, T.; Mori, R.; Debari, K.; Yamada, M. Scanning electron microscopy and electron probe microanalysis studies of human pineal concretions. Microscopy 1994, 43, 307–317. [Google Scholar]
- Bocchi, G.; Valdre, G. Physical, chemical, and mineralogical characterization of carbonate-hydroxyapatite concretions of the human pineal gland. J. Inorg. Biochem. 1993, 49, 209–220. [Google Scholar] [CrossRef]
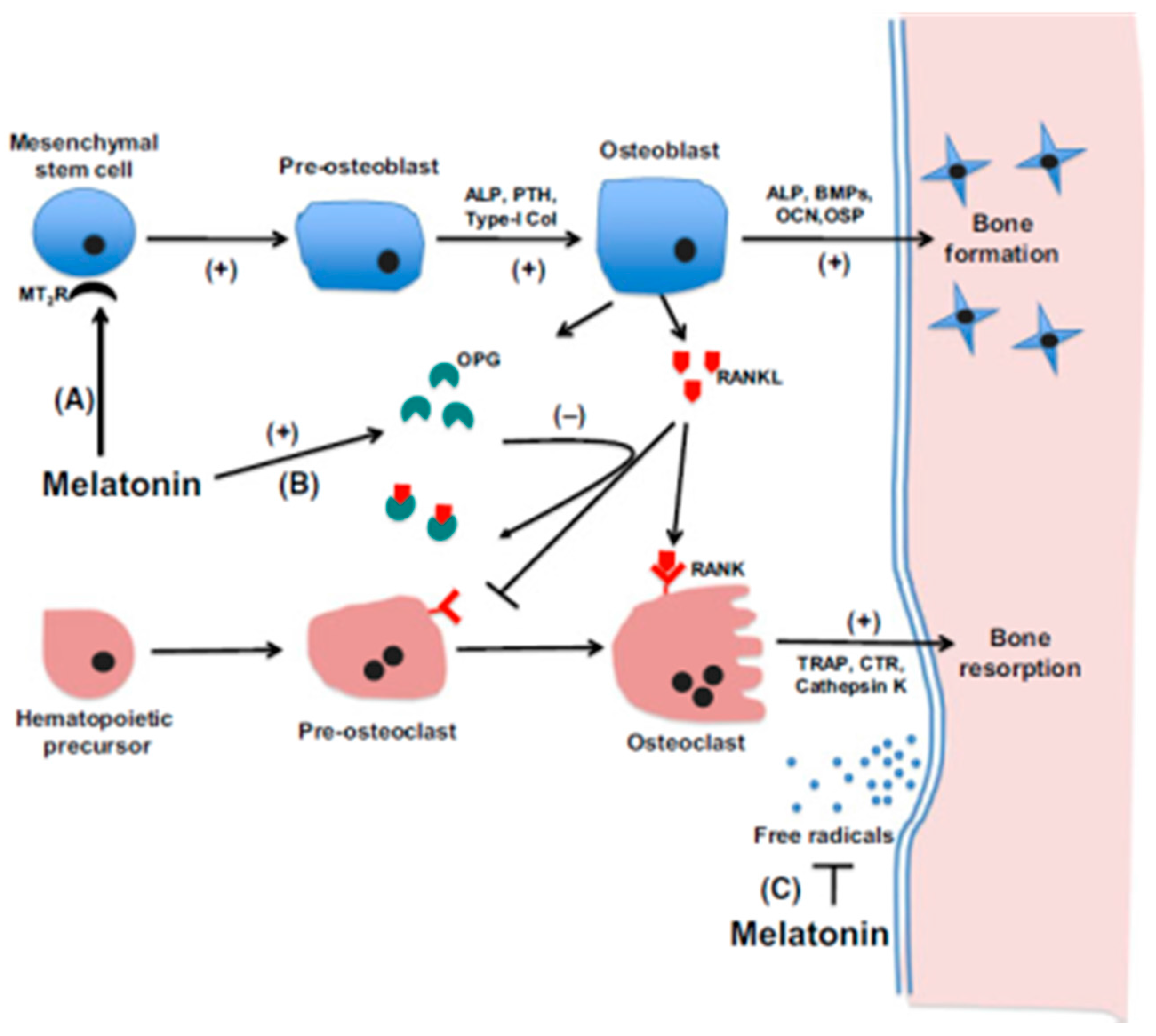
- Maria, S.; Swanson, M.H.; Enderby, L.T.; D’Amico, F.; Enderby, B.; Samsonraj, R.M.; Dudakovic, A.; van Wijnen, A.J.; Witt-Enderby, P.A. Melatonin-micronutrients Osteopenia Treatment Study (MOTS): a translational study assessing melatonin, strontium (citrate), vitamin D3 and vitamin K2 (MK7) on bone density, bone marker turnover and health related quality of life in postmenopausal osteopenic women following a one-year double-blind RCT and on osteoblast-osteoclast co-cultures. Aging 2017, 26, 256–285. [Google Scholar]
- Maria, S.; Samsonraj, R.M.; Munmun, F.; Glas, J.; Silvestros, M.; Kotlarczyk, M.P.; Rylands, R.; Dudakovic, A.; van Wijnen, A.J.; Enderby, L.T.; et al. Biological effects of melatonin on osteoblast/osteoclast cocultures, bone, and quality of life: Implications of a role for MT2 melatonin receptors, MEK1/2, and MEK5 in melatonin-mediated osteoblastogenesis. J. Pineal Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Su, P.; Xu, C.; Chen, C.; Liang, A.; Du, K.; Peng, Y.; Huang, D. Melatonin inhibits adipogenesis and enhances osteogenesis of human mesenchymal stem cells by suppressing PPARγ expression and enhancing Runx2 expression. J. Pineal Res. 2010, 49, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, J.; Ling, Y.; Chen, C.; Liang, A.; Peng, Y.; Chang, H.; Su, P.; Huang, D. Sustained release of melatonin from poly (lactic-co-glycolic acid) (PLGA) microspheres to induce osteogenesis of human mesenchymal stem cells in vitro. J. Pineal Res. 2013, 54, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.-M.; Li, H.-B.; Sun, S.-X.; Jiang, Y.-C.; Wang, B.; Dong, Y.-F. Melatonin promotes osteoblast differentiation of bone marrow mesenchymal stem cells in aged rats. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 4446–4456. [Google Scholar] [PubMed]
- Shimauchi, Y.; Yahata, T.; Matsubara, S.; Araki, M. Role of tissue interaction between pineal primordium and neighboring tissues in avian pineal morphogenesis studied by intraocular transplantation. Dev. Genes Evol. 2002, 212, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Calvo, J.; Boya, J. Ultrastructural study of the embryonic development in the rat pineal gland. Anat. Rec. 1981, 199, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Hayano, M.; Sung, J.H.; Mastri, A.R.; Hill, E.G. Striated muscle in the pineal gland of swine. J. Neuropathol. Exp. Neurol. 1976, 35, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Diehl, B.J. Occurrence and regional distribution of striated muscle fibers in the rat pineal gland. Cell Tissue Res. 1978, 190, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Maria, S.; Witt-Enderby, P.A. Melatonin effects on bone: Potential use for the prevention and treatment for osteopenia, osteoporosis, and periodontal disease and for use in bone-grafting procedures. J. Pineal Res. 2014, 56, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Radio, N.M.; Doctor, J.S.; Witt-Enderby, P.A. Melatonin enhances alkaline phosphatase activity in differentiating human adult mesenchymal stem cells grown in osteogenic medium via MT2 melatonin receptors and the MEK/ERK (1/2) signaling cascade. J. Pineal Res. 2006, 40, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Jung, Y.H.; Oh, S.Y.; Yun, S.P.; Han, H.J. Melatonin enhances the human mesenchymal stem cells motility via melatonin receptor 2 coupling with Gαq in skin wound healing. J. Pineal Res. 2014, 57, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Sharan, K.; Lewis, K.; Furukawa, T.; Yadav, V.K. Regulation of bone mass through pineal-derived melatonin-MT2 receptor pathway. J. Pineal Res. 2017, 63, e12423. [Google Scholar] [CrossRef] [PubMed]
- Ciavarella, C.; Gallitto, E.; Ricci, F.; Buzzi, M.; Stella, A.; Pasquinelli, G. The crosstalk between vascular MSCs and inflammatory mediators determines the pro-calcific remodelling of human atherosclerotic aneurysm. Stem Cell Res. Ther. 2017, 8, 99. [Google Scholar] [CrossRef] [PubMed]
- El-Zayadi, A.A.; Jones, E.A.; Churchman, S.M.; Baboolal, T.G.; Cuthbert, R.J.; El-Jawhari, J.J.; Badawy, A.M.; Alase, A.A.; El-Sherbiny, Y.M.; McGonagle, D. Interleukin-22 drives the proliferation, migration and osteogenic differentiation of mesenchymal stem cells: A novel cytokine that could contribute to new bone formation in spondyloarthropathies. Rheumatology 2017, 56, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jaiswal, P.K.; Makhoul, G.; Jurakhan, R.; Selvasandran, K.; Ridwan, K.; Cecere, R. Hypoxia modulates cell migration and proliferation in placenta-derived mesenchymal stem cells. J. Thorac. Cardiovasc. Surg. 2017, 154, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Yoon, Y.; Lee, S. Hypoxic preconditioning promotes the bioactivities of mesenchymal stem cells via the HIF-1α-GRP78-Akt axis. Int. J. Mol. Sci. 2017, 18, 1320. [Google Scholar] [CrossRef] [PubMed]
- Ciria, M.; García, N.A.; Ontoria-Oviedo, I.; González-King, H.; Carrero, R.; De La Pompa, J.L.; Montero, J.A.; Sepúlveda, P. Mesenchymal stem cell migration and proliferation are mediated by hypoxia-inducible factor-1α upstream of notch and SUMO pathways. Stem Cells Dev. 2017, 26, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ha, N.; Dai, Q.; Zhou, S.; Yu, C.; Jiang, L. Hypoxia suppresses osteogenesis of bone mesenchymal stem cells via the extracellular signal-regulated 1/2 and p38-mitogen activated protein kinase signaling pathways. Mol. Med. Rep. 2017, 16, 5515–5522. [Google Scholar] [CrossRef] [PubMed]
- Knowles, H. Hypoxic regulation of osteoclast differentiation and bone resorption activity. Hypoxia 2015, 3, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Son, J.-H.; Cho, Y.-C.; Sung, I.-Y.; Kim, I.-R.; Park, B.-S.; Kim, Y.-D. Melatonin promotes osteoblast differentiation and mineralization of MC3T3-E1 cells under hypoxic conditions through activation of PKD/p38 pathways. J. Pineal Res. 2014, 57, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chen, X.; Yan, J.; Li, M.; Liu, T.; Zhu, C.; Pan, G.; Guo, Q.; Yang, H.; Pei, M.; et al. Melatonin at pharmacological concentrations suppresses osteoclastogenesis via the attenuation of intracellular ROS. Osteoporos. Int. 2017, 28, 3325–3337. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, H.J.; Bae, M.-K.; Kim, Y.-D. Suppression of osteoclastogenesis by melatonin: A melatonin receptor-independent action. Int. J. Mol. Sci. 2017, 18, 1142. [Google Scholar] [CrossRef] [PubMed]
- Ping, Z.; Wang, Z.; Shi, J.; Wang, L.; Guo, X.; Zhou, W.; Hu, X.; Wu, X.; Liu, Y.; Zhang, W.; et al. Inhibitory effects of melatonin on titanium particle-induced inflammatory bone resorption and osteoclastogenesis via suppression of NF-κB signaling. Acta Biomater. 2017, 62, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Hanson, J.M.; Nickles, R.J. Cerebral vasocapacitance in human aging. J. Neuroimaging 1994, 4, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Wostyn, P.; Audenaert, K.; De Deyn, P.P. Alzheimer’s disease-related changes in diseases characterized by elevation of intracranial or intraocular pressure. Clin. Neurol. Neurosurg. 2008, 110, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Bäckström, M.; Olson, L.; Seiger, A. N-acetyltransferase and hydroxyindole-O-methyltransferase activity in intraocular pineal transplants: Diurnal thythm as evidence for functional sympathetic adrenergic innervation. Acta Physiol. Scand. 1976, 96, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.Y. Pineal transplants to the anterior chamber of the eye: Evidence for functional reinnervation. Exp. Neurol. 1975, 49, 617–621. [Google Scholar] [CrossRef]
- Wragg, L.E.; Machado, C.R.; Snyder, S.H.; Axelrod, J. Anterior chamber pineal transplants: Their metabolic activity and independence of environmental lighting. Life Sci. 1967, 6, 31–38. [Google Scholar] [CrossRef]
- Aguado, L.I.; Benelbaz, G.A.; Gutierrez, L.S.; Rodriguez, E.M. Ultrastructure of the rat pineal gland grafted under the kidney capsule. Cell Tissue Res. 1977, 176, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Scott, D.E.; Reiter, R.J. No difference in day-night serum melatonin concentration after pineal grafting into the third cerebral ventricle of pinealectomized rats. J. Pineal Res. 1991, 11, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Welsh, M.G.; Li, K. Third ventricular pineal gland grafts in the gerbil: An electron microscopical and immunohistochemical investigation. Microsc. Res. Tech. 1992, 21, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Scott, D.E.; Reiter, R.J. Transplantation of the mammalian pineal gland: Studies of survival, revascularization, reinnervation, and recovery of function. Exp. Neurol. 1993, 122, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J. The effect of pinealectomy, pineal grafts and denervation of the pineal gland on the reproductive organs of male hamsters. Neuroendocrinology 1967, 2, 138–146. [Google Scholar] [CrossRef]
- Provinciali, M.; Di Stefano, G.; Bulian, D.; Tibaldi, A.; Fabris, N. Effect of melatonin and pineal grafting on thymocyte apoptosis in aging mice. Mech. Ageing Dev. 1996, 90, 1–19. [Google Scholar] [CrossRef]
- Moroni, F.; Marcheselli, F.; Recchioni, R.; Fattoretti, P.; Bertoni-Freddari, C. Pineal graft in old rats improves erythrocyte resistance to peroxyl radical-induced hemolysis. Biogerontology 2004, 5, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Vriend, J.; Reiter, R.J. Melatonin feedback on clock genes: A theory involving the proteasome. J. Pineal Res. 2015, 58, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kalisinska, E.; Bosiacka-Baranowska, I.; Lanocha, N.; Kosik-Bogacka, D.; Krolaczyk, K.; Wilk, A.; Kavetska, K.; Budis, H.; Gutowska, I.; Chlubek, D. Fluoride concentrations in the pineal gland, brain and bone of goosander (Mergus merganser) and its prey in Odra River estuary in Poland. Environ. Geochem. Health 2014, 36, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Luke, J. Fluoride deposition in the aged human pineal gland. Caries Res. 2001, 35, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Grases, F.; Costa-Bauzà, A.; Prieto, R.M. A potential role for crystallization inhibitors in treatment of Alzheimer’s disease. Med. Hypotheses 2010, 74, 118–119. [Google Scholar] [CrossRef] [PubMed]
- Grases, F.; Simonet, B.M.; Prieto, R.M.; March, J.G. Phytate levels in diverse rat tissues: Influence of dietary phytate. Br. J. Nutr. 2001, 86, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Grases, F.; Simonet, B.M.; Prieto, R.M.; March, J.G. Variation of InsP(4),InsP(5) and InsP(6) levels in tissues and biological fluids depending on dietary phytate. J. Nutr. Biochem. 2001, 12, 595–601. [Google Scholar] [CrossRef]
- Herwig, A.; Pévet, P.; Bothorel, B.; Steinlechner, S.; Saboureau, M. Trans-pineal microdialysis in the Djungarian hamster (Phodopus sungorus): A tool to study seasonal changes of circadian clock activities. J. Pineal Res. 2006, 40, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Gunaratna, P.C.; Cadle, K.K.; Kissinger, C.B. An improved liquid chromatographic method with electrochemical detection for direct determination of serotonin in microdialysates from Caudate-putamen and pineal gland regions of rat brain. J. Neurosci. Methods 2006, 155, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Barker, S.A.; Borjigin, J.; Lomnicka, I.; Strassman, R. LC/MS/MS analysis of the endogenous dimethyltryptamine hallucinogens, their precursors, and major metabolites in rat pineal gland microdialysate. Biomed. Chromatogr. 2013, 27, 1690–1700. [Google Scholar] [CrossRef] [PubMed]
- Borjigin, J.; Liu, T. Application of long-term microdialysis in circadian rhythm research. Pharmacol. Biochem. Behav. 2008, 90, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-H.; Chang, C.-L.; Lin, K.-C.; Sung, P.-H.; Chai, H.-T.; Zhen, Y.-Y.; Chen, Y.-C.; Wu, Y.-C.; Leu, S.; Tsai, T.-H.; et al. Melatonin augments apoptotic adipose-derived mesenchymal stem cell treatment against sepsis-induced acute lung injury. Am. J. Transl. Res. 2014, 6, 439–458. [Google Scholar] [PubMed]
- Chang, C.-L.; Sung, P.-H.; Sun, C.-K.; Chen, C.-H.; Chiang, H.-J.; Huang, T.-H.; Chen, Y.-L.; Zhen, Y.-Y.; Chai, H.-T.; Chung, S.-Y.; et al. Protective effect of melatonin-supported adipose-derived mesenchymal stem cells against small bowel ischemia-reperfusion injury in rat. J. Pineal Res. 2015, 59, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Han, Y.-S.; Lee, S.H. Potentiation of biological effects of mesenchymal stem cells in ischemic conditions by melatonin via upregulation of cellular prion protein expression. J. Pineal Res. 2017, 62, e12385. [Google Scholar] [CrossRef] [PubMed]
- Mortezaee, K.; Khanlarkhani, N.; Sabbaghziarani, F.; Nekoonam, S.; Majidpoor, J.; Hosseini, A.; Pasbakhsh, P.; Kashani, I.R.; Zendedel, A. Preconditioning with melatonin improves therapeutic outcomes of bone marrow-derived mesenchymal stem cells in targeting liver fibrosis induced by CCl4. Cell Tissue Res. 2017, 369, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Mendivil-Perez, M.; Soto-Mercado, V.; Guerra-Librero, A.; Fernandez-Gil, B.I.; Florido, J.; Shen, Y.-Q.; Tejada, M.A.; Capilla-Gonzalez, V.; Rusanova, I.; Garcia-Verdugo, J.M.; et al. Melatonin enhances neural stem cell differentiation and engraftment by increasing mitochondrial function. J. Pineal Res. 2017, 63, e12415. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Huang, S.H.; Chen, J.W.; Wang, K.C.; Yang, Y.R.; Liu, P.F.; Lin, G.J.; Sytwu, H.K. Melatonin enhances interleukin-10 expression and suppresses chemotaxis to inhibit inflammation in situ and reduce the severity of experimental autoimmune encephalomyelitis. Int. Immunopharmacol. 2016, 3, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Forrestel, A.C.; Miedlich, S.U.; Yurcheshen, M.; Wittlin, S.D.; Sellix, M.T. Chronomedicine and type 2 diabetes: Shining some light on melatonin. Diabetologia 2017, 60, 808–822. [Google Scholar] [CrossRef] [PubMed]
- Kubatka, P.; Bojková, B.; Mciková-Kalická, K.; Mníchová-Chamilová, M.; Adámeková, E.; Ahlers, I.; Ahlersová, E.; Cermáková, M. Effects of tamoxifen and melatonin on mammary gland cancer induced by N-methyl-N-nitrosourea and by 7,12-dimethylbenz(a)anthracene, respectively, in female Sprague-Dawley rats. Folia Biol. 2001, 47, 5–10. [Google Scholar]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, D.X.; Xu, B.; Zhou, X.; Reiter, R.J. Pineal Calcification, Melatonin Production, Aging, Associated Health Consequences and Rejuvenation of the Pineal Gland. Molecules 2018, 23, 301. https://doi.org/10.3390/molecules23020301
Tan DX, Xu B, Zhou X, Reiter RJ. Pineal Calcification, Melatonin Production, Aging, Associated Health Consequences and Rejuvenation of the Pineal Gland. Molecules. 2018; 23(2):301. https://doi.org/10.3390/molecules23020301
Chicago/Turabian StyleTan, Dun Xian, Bing Xu, Xinjia Zhou, and Russel J. Reiter. 2018. "Pineal Calcification, Melatonin Production, Aging, Associated Health Consequences and Rejuvenation of the Pineal Gland" Molecules 23, no. 2: 301. https://doi.org/10.3390/molecules23020301
APA StyleTan, D. X., Xu, B., Zhou, X., & Reiter, R. J. (2018). Pineal Calcification, Melatonin Production, Aging, Associated Health Consequences and Rejuvenation of the Pineal Gland. Molecules, 23(2), 301. https://doi.org/10.3390/molecules23020301