MiR200 and miR302: Two Big Families Influencing Stem Cell Behavior †
by
 , , , , , and
, , , , , and
Francesca Balzano
1 ,
,
Sara Cruciani
1,2,
Valentina Basoli
1,2,
Sara Santaniello
1,2,
Federica Facchin
3 ,
,
Carlo Ventura
2,4 and
Margherita Maioli
1,2,5,6,* 1
Department of Biomedical Sciences, University of Sassari, Viale San Pietro 43/B, 07100 Sassari, Italy
2
Laboratory of Molecular Biology and Stem Cell Engineering, National Institute of Biostructures and Biosystems, Innovation Accelerator, CNR, Via Piero Gobetti 101, 40129 Bologna, Italy
3
Department of Experimental, Diagnostic and Specialty Medicine (DIMES), University of Bologna, Via Massarenti 9, 40138 Bologna, Italy
4
GUNA ATTRE (Advanced Therapies and Tissue REgeneration), Innovation Accelerator, CNR, Via Piero Gobetti 101, 40129 Bologna, Italy
5
Istituto di RicercaGenetica e Biomedica, Consiglio Nazionaledelle Ricerche (CNR), Monserrato, 09042 Cagliari, Italy
6
Center for Developmental Biology and Reprogramming (CEDEBIOR), Department of Biomedical Sciences, University of Sassari, Viale San Pietro 43/B, 07100 Sassari, Italy
*
Author to whom correspondence should be addressed.
†
miRNA and epigenetic changes in stem cell fate.
Molecules 2018, 23(2), 282; https://doi.org/10.3390/molecules23020282
Submission received: 9 January 2018
/
Revised: 25 January 2018
/
Accepted: 27 January 2018
/
Published: 30 January 2018
(This article belongs to the Section Chemical Biology)
Abstract
:In this review, we described different factors that modulate pluripotency in stem cells, in particular we aimed at following the steps of two large families of miRNAs: the miR-200 family and the miR-302 family. We analyzed some factors tuning stem cells behavior as TGF-β, which plays a pivotal role in pluripotency inhibition together with specific miRNAs, reactive oxygen species (ROS), but also hypoxia, and physical stimuli, such as ad hoc conveyed electromagnetic fields. TGF-β plays a crucial role in the suppression of pluripotency thus influencing the achievement of a specific phenotype. ROS concentration can modulate TGF-β activation that in turns down regulates miR-200 and miR-302. These two miRNAs are usually requested to maintain pluripotency, while they are down-regulated during the acquirement of a specific cellular phenotype. Moreover, also physical stimuli, such as extremely-low frequency electromagnetic fields or high-frequency electromagnetic fields conveyed with a radioelectric asymmetric conveyer (REAC), and hypoxia can deeply influence stem cell behavior by inducing the appearance of specific phenotypes, as well as a direct reprogramming of somatic cells. Unraveling the molecular mechanisms underlying the complex interplay between externally applied stimuli and epigenetic events could disclose novel target molecules to commit stem cell fate.
1. Introduction
Epigenetics is a physiological phenomenon of heritable changes in gene function that do not involve changes in the DNA sequence [1]. These changes of cellular and physiological phenotypic traits may result from external or environmental factors, or they may be part of a normal developmental program. In the last years, researchers have focused on the intertwining of epigenetic changes and stem cell dynamics. One of the most extraordinary mechanism that has been described to influence epigenetic processes is the involvement of non-coding RNA transcripts [1,2,3,4,5], in particular microRNAs (miRNAs). Epigenetic regulation by miRNAs can influence some functional aspects and the differentiation of stem cells towards different cell lineages. Several studies confirm the important role of miRNAs involved in cell life during differentiation, growth, expansion, and apoptotic processes. Moreover, miRNAs have arisen as critical molecular regulators for maintaining the functions of stem cells by finely tuning the levels of different signaling proteins [4,5]. In stem cells, this requires a massive and rapid transformation in the cellular phenotype, and prompt important changes in the proteomic network. miRNAs are able to suppress the translation of many target mRNAs, thus inducing fluctuations in gene expression [6]. Approximately, thousands of miRNAs have been identified so far, making miRNAs one of the most abundant classes of gene regulatory molecules in animals [7]. Nevertheless, understanding the mechanisms accounting for their function is still a remarkable challenge. In fact, owing to the important role and functions of miRNAs in regulating many cellular decisions related to pathological processes, they could be evaluated as new therapeutic targets for drug development in the treatment of many diseases. Understanding miRNA biogenesis, regulation, and mechanism in controlling stem cell differentiation will enhance the likelihood for success in stem cell-based therapeutic approaches, including tissue regeneration and engineering. Within this context, an interesting pathway is the relationship between miRNAs and TGF-β signaling, which has been extensively investigated. Studies by different authors suggest that TGF-β-related signals can either inhibit or enhance miRNA maturation [8,9], being themselves regulated by these small molecules. The aim of the present review is to present the role of specific miRNAs in regulating stem cell patterning, by influencing their pluripotency and differentiation capability. A section will be dedicated to the influence of specific physical stimuli, as electromagnetic fields, on stem cell epigenetic fate.
2. Stem Cells
Stem cells are undifferentiated cells that can differentiate into specialized cells or divide to produce more stem cells. In mammals, there are two broad types of stem cells: embryonic stem cells, which are isolated from the inner cell mass of blastocysts, and adult stem cells, which are found in various tissues. In adult organisms, stem cells and progenitor cells act as a repair system for the body, replenishing impaired tissues, having the remarkable potential to develop into many different cell types.
The key characteristics of stem cells are:
- Asymmetric divisions with the formation of stem cells (self-regeneration or self-renewal) and daughter cells with reduced differentiation potential, which transiently replicate and thus differentiate after a specific number of divisions;
- The persistence of the replicative capacity for the entire life of the individual;
- The maintenance a source of stem cells due to a specific microenvironment (stem cell niche) formed by other cells.
Self-regeneration is the ability to create an undifferentiated cell perfectly identical to the original stem cell (SC). Their cellular power (plasticity or stemness) is indeed the ability to develop into highly specialized cell types for specific biological functions. Depending upon the pluripotency, we can distinguish different types of stem cells. Totipotency is the ability to give rise to all embryonic and extraembryonic tissues, and is essentially restricted to the earliest stages of embryonic life, when the embryo is formed by only four or eight cells. Pluripotency is the ability to differentiate in all embryonic tissues, typical of the embryonic SC (ESCs) and of the induced pluripotent SC (IPSCs). Multipotency the ability of one of the three germinative layers (endoderm, mesoderm, or ectoderm), or a specific tissue, to differentiate into different cell types and is associated with adult SC. Finally, unipotency is the ability to differentiate into a single cell type [10,11].
3. Embryonic and Adult Stem Cells. Cellular Reprogramming and Generation of Pluripotent Stem Cells
Human Embryonic stem Cells (hESCs) were described for the first time in 1998 after being isolated from the internal cellular masses of human blastocysts which were donated for research purposes [12]. These cells were characterized, in vivo, through the formation of teratomas and, in vitro, through the expression analysis of specific enzymatic markers and assays, and through differentiation studies [12]. Unlike animal models, it is not possible, for ethical reasons, to perform chimerism studies and transmission to the germ line on hESCs. It is known that stem cells (both human and murine) have the ability to replicate indefinitely in vitro, although there are differences both at the gene expression level and at the growth medium conditions (e.g., the use of the Leukemia Inhibitory Factor—LIF—is indispensable for murine stem cells, but it is not necessary for human ones) [13].
4. Induced Pluripotent Stem Cells (iPSCs)
In 2006, Yamanaka, Nobel Prize for Medicine or Physiology in 2012, and his Collaborators at the University of Kyoto identified conditions that could “genetically reprogram” specialized adult cells to assume a state similar to that of stem cells. These adult cells, called induced pluripotent stem cells (iPSCs), have been reprogrammed by introducing important genes able to induce self-renewal and essential differentiation capability, like in ESCs. The strategy used was the nuclear reprogramming, a technique that experimentally induced a stable change in the nucleus of a mature cell, which can be maintained during the following cellular divisions. These changes are most frequently associated with the reacquisition of a pluripotent state, thereby providing the cells with a developmental potential [14]. This approach uses mature “somatic” cells from the adult and introduces the genes encoding critical transcription factors, which in turn regulate other key genes for the early steps in embryonic development. Four transcription factors (Oct4, Sox2, Klf4, and c-Myc) were required to reprogram mouse fibroblasts (cells found in the skin and other connective tissues) to acquire an embryonic-like state [8]. These factors were known to be involved in the maintenance of pluripotency. The newly-created iPSCs were found to be highly similar to ESCs and could be produced after several weeks in culture [8,9]. In 2007, two different research groups reached a new milestone by deriving iPSCs from human cells, using either the original four genes [10] or a different combination containing Oct4, Sox2, NANOG and Lin28 [11,15]. Since then, Researchers have generated iPSCs from somatic tissues of monkeys [14] and rats [16,17]. However, these cells can retain methylation patterns from their original cells, a phenomenon called epigenetic memory, which has been shown to influence the differentiation potential [18]. Several studies analyzed the operating system in iPSCs in the context of reprogramming yield. Armstrong et al. [19] compared iPSCs with ESCs, showing that reactive oxygen species (ROS) levels and mitochondrial mass are similar in these cells and significantly less than in human dermal fibroblasts from which iPSCs were first generated. These cells are expected to reproduce the behavior of ESCs in response to oxidative stress (OS). Armstrong et al. [20] also found a reduced number of mitochondria in pluripotent stem cells, as compared to fibroblasts. The loss of mitochondria appears to be a key step in the reprogramming process, being linked to a switch from an oxidative to a glycolytic-dependent metabolism [21]. It has been shown that to support their high proliferation rates, pluripotent stem cells are predominantly anabolic and tend to inactivate catabolic pathways. Consequently, energy production is obtained through glycolysis rather than oxidative phosphorylation, which is related to the decrease in the number of mitochondria [22,23]. A recent study suggests that some miRNAs play a fundamental role in regulating differentiation and maintaining pluripotency/multipotency in stem cells [24].
5. miRNAs
miRNAs are a type of non-coding RNA, 21–24 nucleotides in length, that function in the post-transcriptional regulation of gene expression. They can be found as isolated transcript units or clustered and co-transcribed as polycistronic primary transcripts [25]. Typically a miRNA interacts, through its specific sequence named Seed, with specific mRNAs through complementary base-pairing, to influence the translation or stability of the target mRNA molecule. MiRNA life is subjected to multipurpose regulation [26], beginning with the transcription of its own genes. The resulting transcribed RNA sequence is folded into a hairpin structure and then processed in the nucleus by Drosha (a class 2 ribonuclease enzyme III) in a new structure called pre-miRNA. Exportin-5 exports the hairpin pre-RNA from the nucleus to the cytoplasm. Subsequently, another endonuclease, called Dicer, catalyzes pre-miRNA deployment into mature miRNA. Argonaut and helix proteins are co-recruited to form the RNA-induced silencing complex (RISC) that allows the control of miRNA with consequent translational repression, and destabilization cleavage for post-transcriptional protein regulation. Most targeted miRNA sites are found in the 3′untranslated regions (3′UTR) of mRNA transcripts [27]. However, other studies have shown that targeted miRNA sites may reside beyond 3’UTR in mRNA protein encoding sequences [28]. Some pre-miRNAs can be secreted from the cells in the extracellular environment, becoming circulating miRNAs [29], to exert their role in other cells within the tissue or in different tissues. MiRNAs regulate genes and define cell fate and behavior. One of the main miRNA control and regulation mechanisms is represented by transforming growth factor beta (TGF-β) signaling [8,9].
6. Stem Cells, TGF-β, and Epigenetics
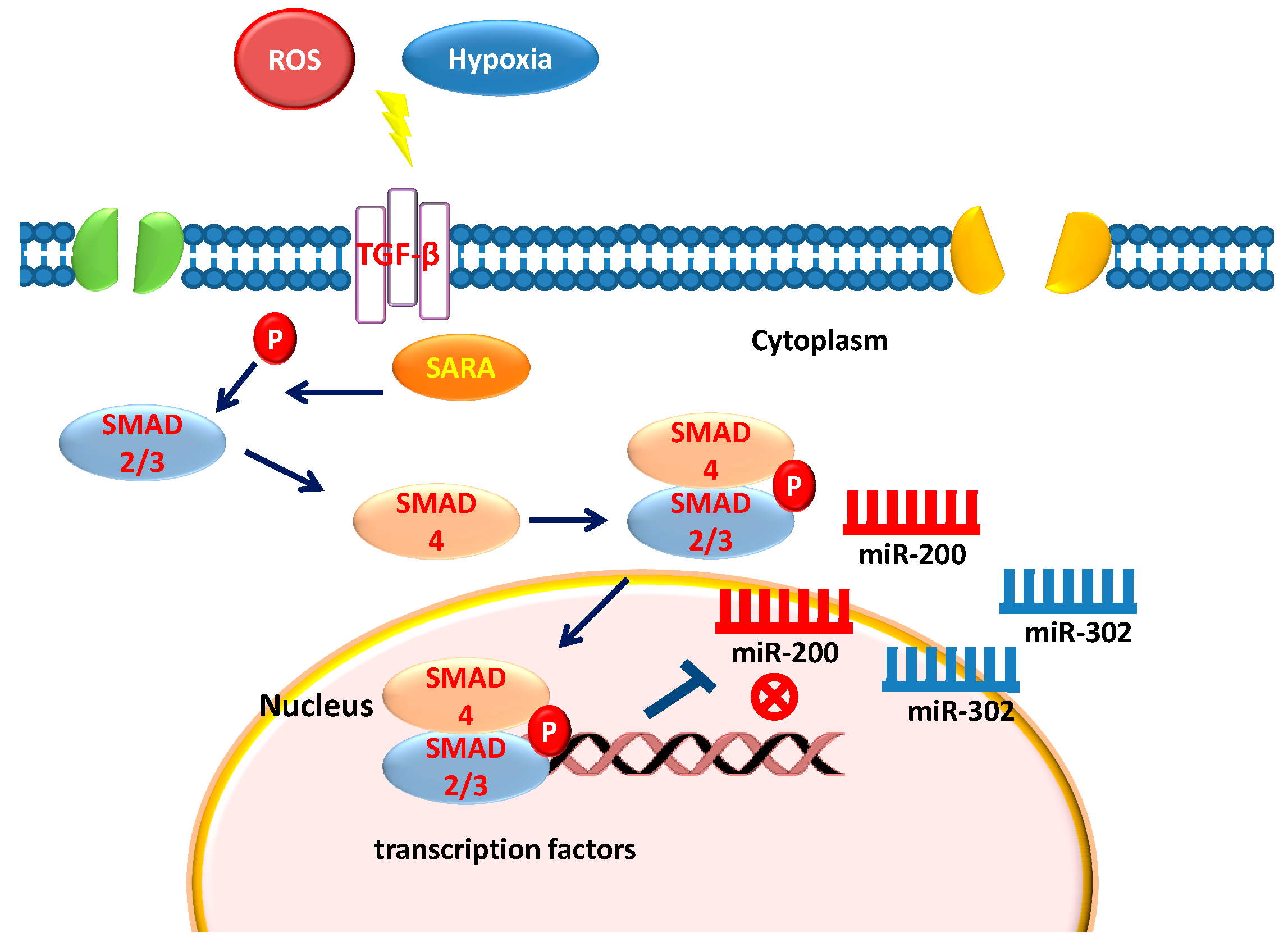
TGF-β is a multifunctional cytokine belonging to the transforming growth factor superfamily that includes four different isoforms (TGFB1, TGFB2, TGFB3, and TGFB4) and many other signaling proteins produced by all white blood cell lineages [8,9]. Activated TGF-β recruits other factors to form a serine/threonine kinase complex that binds to the TGF-β receptors, which comprises both type 1 and type 2 receptor subunits. After binding TGF-β, the two receptor subunits undergo a cross phosphorylation, then the activated receptor recruits the signaling cascade of small mother against decapentaplegic (SMAD) proteins [30,31]. This leads to the activation of different downstream targets and regulatory proteins, inducing the transcription of different target genes involved in differentiation, chemotaxis, proliferation, and activation of many immune cells [32]. A defect in the growth inhibitory response to TGF-β often correlates with the malignancy of many types of cancers [33]. Among the TGF-β protein signaling, Activin/Nodal branch is essential for maintaining pluripotency in hESCs [34,35,36]. Components of the hESC core transcriptional regulatory circuitry [37], such as NANOG represent one of the target genes that are activated by Smad2/3 [38,39]. One of the key events in acquiring stem cell properties of both breast cancer and normal mammary stem cells is an epithelial-mesenchymal transition (EMT) induced by TGF-β. TGF-β family signaling regulates stemness of normal and neoplastic stem cells by exerting different effects depending upon the cell type, the microenvironment and physiological state of the cells [40,41]. Recent evidence indicates an association between miRNAs and TGF-β signaling, providing a new insight into the deepness of these interactions that are also the basis of pluripotency and cell differentiation [42,43]. The relationship between TGF-β and miRNA signaling has been extensively studied, unraveling both an inhibitory or stimulatory role of the TGF-β pathway on the maturation of miRNAs [44]. Moreover, Smad proteins have been shown to control the transcription of miRNA coding genes by binding their gene promoter [45]. Smads control miRNAs synthesis by a Smad2-3 complex binding to Smad4 [44]. The Smad2-3-4 complex is moved to the nucleus where it binds to specific Smad binding elements in the DNA, while the Smad3 is recruited from the Drosha/DGCR8 microprocessor complex and promotes miRNA maturation [44,46,47]. The mechanisms that lie at the basis of the suggested activation pathways have not been completely elucidated. Some of the known activation pathways are cell or tissue-specific; others have been seen in response to many cytotoxic agents [48,49]. Some of the factors that can trigger the TGF-β are: proteases, integrins, pH, and ROS (reactive oxygen species) [50,51]. The perturbation of one of these activating factors can lead to altered TGF-β levels which in turn bring several outcomes such as: inflammation, autoimmune reactions, fibrosis, and cancer [52].
7. MicroRNAs and Cell Reprogramming
As reported above, Yamanaka and his Co-workers showed the feasibility to obtain pluripotent progenitors from fibroblasts by forcing the expression of four genes: OCT3/4, Sox2, Klf4, and C-MYC, the so called Yamanaka factors or OSKM [53]. This discovery is considered one of the most important of the third millennium since it proved the chance of reversing life trajectories [54,55,56]. Interestingly, some of those stem cell core regulators are able to activate the promoters of several miRNAs in ESCs, including miR-290–295, miR-302-367, and miR-92clusters [57]. MiRNA analysis has defined that ESCs and iPSCs have a distinct miRNA expression patterns, compared to differentiated somatic cells [58]. The miRNA-mediated reprogramming technique has been shown to be more efficient than the standard OSKM over-expression method [59]. Furthermore, cell reprogramming mediated by miRNA may provide a safer approach than traditional DNA reprogramming methods.
8. Stem Cell Function and miRNA Transcription
The mechanisms by which cells control the production and function of miRNAs have been recently described [60]. MiRNAs transcription is regulated like protein coding genes, for example by DNA methylation or through DNA promoter binding factors such as p53 or C-MYC [61]. As previously stated, the factors Oct4, Sox2, and NANOG play a central role in acquiring and maintaining “stemness”: these factors define the future of stem cells through several stages of self and mutual regulation. Recent studies have revealed the transcription of many other actors in the Pluripotency Regulation network, including LIN28, CMYC, Klf4, TCF3, or STAT3 [62]. Some of these nuclear stem cell regulators are able to activate the promoters of different miRNAs in ESCs, including miR-290–295, miR-302–367 and miR-92 clusters [63]. MiRNAs are central players in the process of ESC self-renewal/differentiation. The loss of mir-145 compromises differentiation and induces the expression of Oct4, Sox2 and Klf4 [45]. It also controls the differentiation of ESCs directly by targeting stem cell factors, thus silencing the self-renewal program [45]. MiRNAs involved in maintaining the pluripotency of stem cells are miR-200C and other miRNAs belonging to miR-203 family [46].
9. MiRNAs Orchestrating Stem Cell Pluripotency and Differentiation
Studies on mice, revealed that the miR-290–295 cluster and miR-296 were expressed in ESCs and that their levels decrease during stem cell differentiation. This proved that the miR-290–295 cluster had specific roles in maintaining pluripotency [64]. Interestingly, some nuclear stem cell regulators like Oct4 are able to activate promoters of several miRNAs in ESCs, including miR-290–295, miR-302–367, miR-371–373 and miR-92 clusters [63]. While miR-290–295 cluster is highly expressed in mESCs, miR-302–367 and miR-371–373 clusters are mainly regulators in hESCs. Actually, the miR-290–295 cluster represents the mouse homologue of human miR-371–373 [65,66,67,68,69]. The human ESCs express many members of the miR-302 family, as miR-302a, miR-302a*, miR-302b, miR-302b*, miR-302c, miR-302c*, and miR-302d [70]. The overexpression of miR-302 in human cancer cell lines resulted in their conversion into pluripotent cells that express key ESC markers such as Oct3/4, SSEA-3, SSEA-4, Sox2, and NANOG [71]. These miRNAs (miR-302s, miR-369s and miR-200c) are highly expressed in iPSCs and ESCs and are capable of reprogramming human somatic cells to acquire the pluripotent state [72]. Some miRNAs are known to facilitate cellular reprogramming for the generation of iPSCs, while other miRNAs can inhibit this process. This is achieved by reducing directly or indirectly the expression of the pluripotent genes involved in cellular reprogramming. In this regard, miR-34 (miR-34a, b, c) was found to suppress reprogramming of somatic cells by deleting the expression of NANOG, Sox2, and N-Myc [73]. Other differentiation-related miRNAs, as miR-296 and miR-470 were found to target coding regions of NANOG, SOX2 and OCT4 to promote differentiation [74]. The transcriptional effects of miR-21 itself are regulated in ESCs by a transcriptional repressor called the RE1-silencing transcription factor (REST). REST directly interacts with an upstream cis-element of the miR-21-dependent transcription factors, thus promoting differentiation [75]. A recent study demonstrates that miR-21, which is induced following the commitment of mouse ESCs to differentiation, has a potential binding sites in the 3′ UTRs of the mRNAs encoding for NANOG, SOX2, and OCT4, thus modulating the corresponding protein levels [76]. MiRNAs regulation defines ESC fate and behavior (Table 1).
10. The Two Big miRNA Families of Pluripotency: miR-200 and miR-302/367
10.1. MiRNA-200 Family
The miR-200 family comprises five members (miR-200a, miR-200b, miR-200c, miR-141, and miR-429), located within two clusters on two different chromosomes: chromosomes 1 and 12 in humans [77]. Members of the miR-200 family, directly activated by Oct4 and Sox2 able to bind to their promoter regions, help fibroblasts to overcome the MET (Mesenchymal Epithelial transition) barrier and facilitate iPSC generation. All members of the miR-200 family (miR-200a, miR-200b, miR-200c, miR-141, and miR-429) are significantly more expressed in iPSCs and ESCs than in MEF cells, suggesting that miR-200 family may correlate with pluripotency and that its activation may promote the emergence of iPSCs. Members of the miR-200 family are specific and direct targets for the key pluripotency-associated transcription factors Oct4/Sox2 in iPSC generation. However, very little is known about the role and intrinsic regulation of the miR-200 family during iPSC generation. Oct4 and Sox2 could bind the promoter region of miR-141/200c and miR-200a/b/429, respectively, and activate the transcription of miR-200s. MiR-200 family members have been found to inhibit ESC differentiation directly targeting Cadherin11 and Neuropilin1 [69]. Recently, it has been demonstrated that the MET process is an important early event in iPSC generation [78], and that the activation of EMT is associated with the maintenance of stem-cell properties [79]. Noteworthy, TGF-β induces reversible DNA methylation of miR-200 promoters [80]. Moreover, miR-200b and miR-200c loci can be reversibly de-methylated during prolonged signaling of TGF-β. Changes in the degree of methylation of the miR-200 promoter are closely related to miR-200 expression, and repression. The mechanism through which TGF-β controls the DNA methylation of miR-200 is not clear, but it may involve the presence of active DNA Methyltransferases (DNAMTs). Even invasive mesenchymal breast cancer cell lines show miR-200 methylated promoters, in contrast to epithelial cells where miR-200 promoters were de-methylated. Similar observations have recently been yielded from primary mesenchymal cells and invasive bladder tumors [81,82,83]. Furthermore, Gregorya et al. have identified a central role for an autocrine TGF-β/ZEB/miR-200 signaling network in controlling the transition between epithelial and mesenchymal states. Prolonged activation of this pathway leads to epigenetic changes in miR-200 and may contribute to invasive breast cancer progression [84]. Considering these findings, a remarkable connection between EMT and breast cancer stem cells was recently demonstrated, where TGF-β treatment was shown to initiate EMT-associated embryonic differentiation programs of epithelial–mesenchymal, with EMT and MET representing a basic mechanism for epithelial cell plasticity [79]. MET has been shown to be an important early event in somatic cell reprogramming [76,77,78,79,80,81,82,83,84,85]. The miR-200 family and ZEB1 have been shown to be key regulators of these processes [85,86]. A link among the TGF-β–related factors, the bone morphogenetic proteins, and the miR-200 family have recently been described in somatic cell reprogramming [87]. It is well known that the miR-200 family suppresses EMT. It has also been reported that the knockdown of miR-200c inhibits NANOG and up regulates GATA4, thereby inhibiting self-renewal and promoting hESC differentiation [88]. Transfection with a group of miRNAs including miR-200c, miR-302s, and miR-376s family successfully coaxed somatic cells into an iPSC fate [89]. As stated above, TGF-β plays an important role in repressing the miR-200 [90].
10.2. The miR-302/367 Family
The miR-302/367 big family, comprises miR-367, miR-302d, miR-302a, miR-302c, miR-302b, and miR-371/373. The miR-302 cluster is the most abundant miRNA family in hESCs, and can promote somatic cell reprogramming, while miR-371–373, are less expressed [91]. The structure of the gene coding for the human miR-302/367 cluster has been well characterized [92]. MiR-371, miR-372, and miR-373 are gene clusters located in the region of the human chromosome 19q13.4. They are the human equivalent of miR-290/295 in the mouse [93]. MiR-302/367 family is ubiquitously distributed in vertebrates as a cluster spanning the La-related protein 7 (LARP7) intron, in the 4q25 region of human chromosome 4 [92]. The cluster was initially identified to be specifically expressed in undifferentiated hESCs, and their malignant counterpart human embryonic carcinoma cells (hECCs), and was found to play a role in pluripotency of stem cells and in cancer formation [92]. This cluster is highly expressed in, and therefore constitutes a signature miRNA for stem cells [93,94]. It is also highly expressed in iPSCs while progressively declining during differentiation [93]. The miR-290–295/miR-371–373 family members directly control the G1-S cell cycle transition and inhibit apoptosis. This family also plays a significant role in regulating cell proliferation, differentiation and reprogramming [95,96]. It was demonstrated that Oct3/4, NANOG, Rex1, and Sox2 act as transcriptional activators of the miR-302/367 cluster [97,98]. This cluster is specifically expressed in embryonic stem cells, iPSCs or tumor cells, to coordinate proliferation, differentiation, pluripotency, maintenance, and reprogramming [99,100]. MiR-302 members repressed lysine-specific histone demethylase 1 and 2 (AOF1 and AOF2) and methyl-CpG binding proteins (MECP1 and MECP2), leading to destabilization of DNA methyltransferase 1, thus causing a genome-wide demethylation promoting reprogramming and iPSC development [101]. Overexpression of the miR-302-367 cluster significantly increased the conversion of reprogrammed iPSCs by repressing MBD2 (methyl-CpG binding domain) expression, thereby inducing NANOG expression [102].
Another study found that the miR-302/TGF-β/Nodal/Smad-2/3 pathway was also involved in EMT. In EMT, epithelial cells form monolayers and display elongated morphology and enhanced motility [103]. MiR-302 members directly repressed the expression of the transforming growth factor beta receptor 2 (TGFBR2), and the member C (RHOC) genes of the Ras family, thus allowing the EMT process [92]. In addition, miR-302 members could negatively regulate the level of the TGF-β related protein Lefty1 and Lefty2, thus becoming an upstream modulator of the TGF-β/Nodal signaling pathway, finely tuning pluripotency and differentiation [104]. The main core promoter for this process has been shown to include the embryonic stem cell factors NANOG, OCT3/4, SOX2, and REX1 [105]. This is an additional cue to consider the miR-302/367 cluster an important stem cell signature for stem cells [92]. This cluster is also highly expressed in iPSCs where its expression decreases during differentiation [105].
11. Epithelial–Mesenchymal Transition (EMT) in Stem Cells
TGF-β plays an important role during cell reprogramming in iPSC generation and pluripotency. Fibroblasts undergoing MET adopt the epithelial state of pluripotent cells and conversely human pluripotent stem cells resemble cells obtained from the epiblast [106]. The occurrence of EMT in human pluripotent stem cells and embryoid bodies during cell commitment reflects the EMT observed during gastrulation in human development [107]. By contrast, suppression of EMT by inhibiting TGF-β signaling improves reprogramming efficiency [108]. The TGF-β signaling pathway promotes EMT, and miR-302 promotes reprogramming partly decreasing the expression of TGF-β receptor 2, and the related SMAD2 and/or SMAD3 phosphorylation and activation [109]. Furthermore, as stated above, TGF-β plays an important role in repressing the miR-200. Members of the miR-200 family are specific and direct targets for the key pluripotency-associated transcription factors Oct4/Sox2 in iPSC generation. These miRNAs act as regulators of EMT in many cell types. For example, snail promotes EMT through the repression of miRNA 200 family members and induces mesoderm differentiation during the epiblast stem cell stage, suggesting that miR-200 family members might suppress EMT and the differentiation of ESCs at the epiblast stem cell stage [67]. These two major miRNAs families are the basis of the pluripotency induction by interacting with the TGF-β, thus demonstrating that TGF-β plays a crucial role in suppressing pluripotency by acting on the EMT [106]. Among the other TGF-β may be triggered by ROS (reactive oxygen species) [50].
12. Hypoxia Exerts Opposite Effect on miR-200 and miR-302 Family
Hypoxia is broadly recognized as a mechanism that drives angiogenesis. Several possible mechanisms have been proposed for down-regulation of miRNAs in a low oxygen environment [110]. MiR-200b down regulation is crucial in inducing angiogenesis via Ets-1 direct targeting. These findings consolidate the notion that under hypoxic conditions miR-200b down-regulation is required to relieve Ets-1 repression resulting in successful angiogenic outcomes. Furthermore, hypoxia-induced angiogenesis, as well as Ets-1 up-regulation was rescued by delivery of the miR-200b mimic [110]. On the other hand, miR-302 expression was induced by hypoxia. Recent findings by other Authors highlighted a central role of miR-302 overexpression during hypoxia/re oxygenation-mediated cardiomyocyte apoptosis. In particular, this proapoptotic effect of miR-302 was mediated by inhibiting the expression of the antiapoptotic molecule myeloid leukemia cell-differentiation protein-1 (Mcl-1) [111].
13. Physical Stimuli: Ad Hoc Conveyance of Electromagnetic Fields to Drive Stem Cell Fate
Previous studies revealed a role of reactive oxygen species (ROS) in transducing mechanically or electrically-induced cardiovascular differentiation in mouse ESCs [112]. Moreover, fibroblast direct reprogramming toward cardiogenic, neurogenic, and myogenic lineages occurred through an upregulation of the NADPH oxidase isoforms Nox2 and Nox4 [113]. In the same paper, Maioli et al. described an interesting biphasic effect on the expression of certain stemness related genes, elicited by the Radio Electric Asymmetric Conveyer (REAC), an innovative device conveying radio electric fields of 2.4 GHz. In particular, REAC-stimulation was able to induce an increase in the expression of Oct4, Sox2, cMyc, NANOG, and Klf4 within 6–20 h, while downregulating the same genes following prolonged times of exposure [113,114]. This study demonstrates the feasibility of using a physical stimulus to afford the expression of stemness in human adult somatic cells up to the attainment of three major target lineages (cardiogenic, neurogenic, and skeletal myogenic) (Figure 1). So far, the acquirement of a pluripotent state in somatic cells, including fibroblast reprogramming to iPSCs or direct reprogramming to cardiac or neuronal lineages without an iPSC intermediate, has only been achieved by targeted genetic engineering through viral vector-mediated technologies or cumbersome and expensive protein transduction methods or by combinatorial approaches encompassing both viral-mediated gene delivery and chemical stimulation or gene-delivery and physical stimulation of the target cells [114,115,116]. Electromagnetic fields (EL-EMFs) are also able to affect biological processes, including stem cell development proliferation and differentiation [117].
14. Electromagnetic Fields
Baek et al. showed that EMF exposure induces epigenetic changes that promote efficient somatic cell reprogramming to pluripotency [118]. There is a general agreement on the effects of EMF on biological systems. It was indicated that low-frequency electromagnetic fields affect cell migration, cellular differentiation, apoptosis, and stress response [119,120]. EMF can also affect many stages of embryonic development, which are of interest for morphology and migration of embryonic cells [120,121]. Moreover, it was reported that EMFs of selected frequencies promote osteogenic and neurogenic differentiation, a finding that has been clinically applied in repairing bone fractures and promoting wound healing [122,123,124]. These studies indicate that EMFs can be involved in managing the conversion of cell destiny. Stem cells respond to EMFs differently, depending on their state of differentiation. It is possible that EMFs modulate the activity of transcription factors and the level of cell cycle regulatory genes [125,126,127]. It is believed that one of the possible mechanisms of EMF biological activity involves the generation of ROS within the cell. Excessive concentration of ROS, such as superoxide anions (O2−) and hydrogen peroxide (H2O2), is considered to be cell destructive and results in inhibition of gene expression. In contrast, small amounts of ROS function as intracellular second messengers and activate signaling cascades involved in growth and differentiation of many cell types. The high level of ROS modifies signaling pathways by phosphorylation mechanisms [128]. However, the energy of a weak EMF is not sufficient to directly break a chemical bond in DNA [128]. Therefore, it can be concluded that genotoxic effects are mediated by indirect mechanisms as microthermal processes, generation of ROS, or disturbance of DNA repair processes [128]. Although ROS were originally thought to be merely a harmful product of metabolism, accumulating evidence demonstrates a role of ROS in cell fate signaling [128]. H2O2 is thought to be the main ROS species involved in intracellular signaling, and in specific contexts can act directly as a second messenger, integrating environmental cues and passing them to downstream signal transduction cascades [128,129]. This is due mostly to the longer half-life of H2O2 and its ability to diffuse easier through membranes than other types of ROS [129,130]. Slight variations in ROS content may have deep effects on stem cell fate [130,131]. iPSCs and ESCs exhibit similar ROS levels and mitochondrial mass, but significantly less than in human dermal fibroblasts from which iPSCs were generated. As stated before, the loss of mitochondria appears to be a key step in the reprogramming process, and is linked to a switch from an oxidative metabolism to a glycolytic-dependent state [132,133]. It has been recently shown that maintaining low (physiological) O2 (2–5% O2) in cultures inhibits spontaneous differentiation and supports pluripotency [132,133,134]. These studies provided evidence that molecular processes that mediate pluripotency and suppress differentiation are supported under 4%-O2 conditions and that the cells retain a “memory” of their prior environment. Hypoxia induces the expression of Sox2 and Oct4 genes that are related to stem cell function [135,136]. In particular, Sox2, together with Sox4, was recently shown to play a pivotal role in the maintenance of stemness in ESCs. Hypoxia has been demonstrated to induce EMT, which prompts invasion and metastasis from cancer cells [135]. During EMT, epithelial cells undergo several biochemical alterations that allow the acquisition of the mesenchymal phenotype enabling cancer cells to evade their “homeland” and to colonize remote locations [135,136]. EMT-inducers, including TGF-β and hypoxia, trigger changes in gene expression by complex signaling pathways [136]. The loss of polarity and gain of motile characteristics of mesenchymal cells during embryonic development have suggested analogies with metastatic cancer cells during malignant progression [135]. Notably, recent data on several cancer types have demonstrated that EMT is involved in generating cells with properties of stem cells [136]. This implies that hypoxia-induced EMT may affect cancer stem cells (CSCs) or induce stem-like cells from more differentiated progenitors determining an increase of CSC population responsible for early systemic cancer dissemination and metastasis formation. Many studies have demonstrated a functional connection between low oxygen level, ROS production, and EMT [134,137]. Indeed, in contrast to cancer cells in which ROS levels are increased, CSCs generally maintain low ROS, exhibiting redox patterns that are similar to the corresponding normal stem cells [137]. Diehn et al. reported that ROS levels are lower in human and murine breast CSCs than in non-stem breast cancer cells [135].
15. ROS and miRNA
The miR-200 family has been extensively studied in the EMT of cancer cells [30]. In EMT, miR-200 family down-modulation enhances cancer aggressiveness and metastases, whereas reintroduction of miR-200 family in some tumors inhibits their growth. ROS induces miR-200c and other miR-200 family members; the following down-modulation of ZEB1 has a key role in ROS-induced apoptosis and senescence [138]. Another study demonstrated that H2O2 and other oxidant agents increase the expression of miR-200c and induce growth arrest, apoptosis, and senescence in HUVEC cells by inhibition of ZEB1 expression [138]. Overall, these findings indicate a potential role of miR-200 family in the regulation of ROS homeostasis [138]. The effect of miR-302s on cell survival under oxidative stress has been shown to determine the effects of miR-302d on ROS, with a protective role against oxidative stress [139]. Accordingly, miR-302d transfection significantly decreased the generation of ROS, indicating that the protective action of miR-302 on oxidant-induced cell death may be related to the inhibition of ROS generation [140].
As mentioned above, the interaction of miR-200 and miR-302 families with ROS is different. Mir-200 family members increase after ROS stimulation, while miR-302 family act by lowering the level of ROS. There are therefore opposite mechanisms that maintain cellular homeostasis and that can be used to modulate pluripotency and differentiation in stem cells [138,139,140,141]. The mechanisms that induce pluripotency and differentiation are complex, and not fully clarified. The interest of the scientific world has increased since Yamanaka created iPSCs, paving a novel way to regenerative medicine. To date, the limits of using iPSCs for regenerative purposes are caused by the use of viruses to transport OCT4, SOX2, c-Myc, NANOG, or other pluripotency factors within the target. This can cause teratoma and possibly cancer.
On the whole, our interest in writing this review was to understand how miRNAs work in stem cells, cancer stem cells, or iPSCs. We also attempted in drawing the readers’ attention on transcription factors, and physical stimuli that can be used to maintain pluripotency, or to induce differentiation.
16. Conclusions
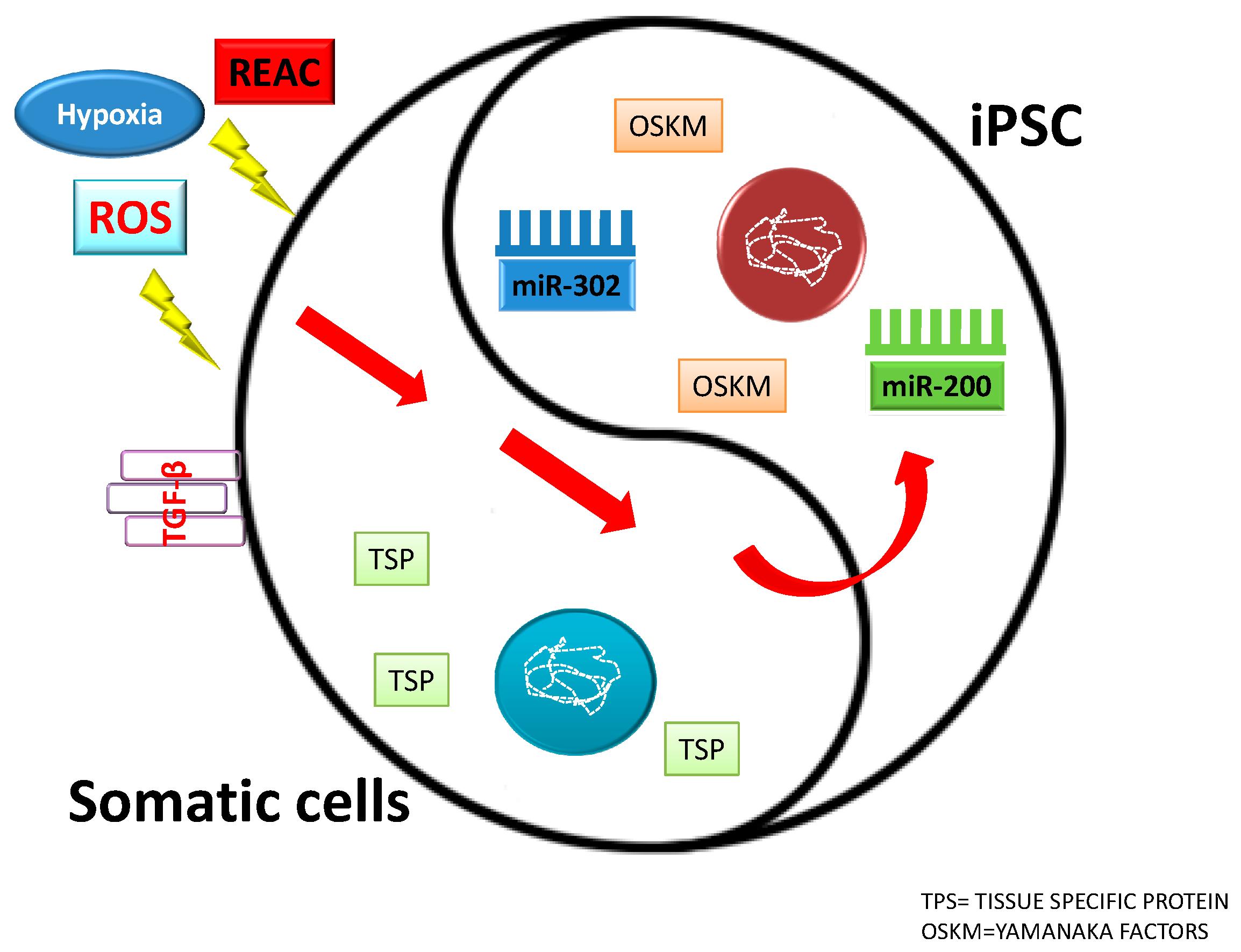
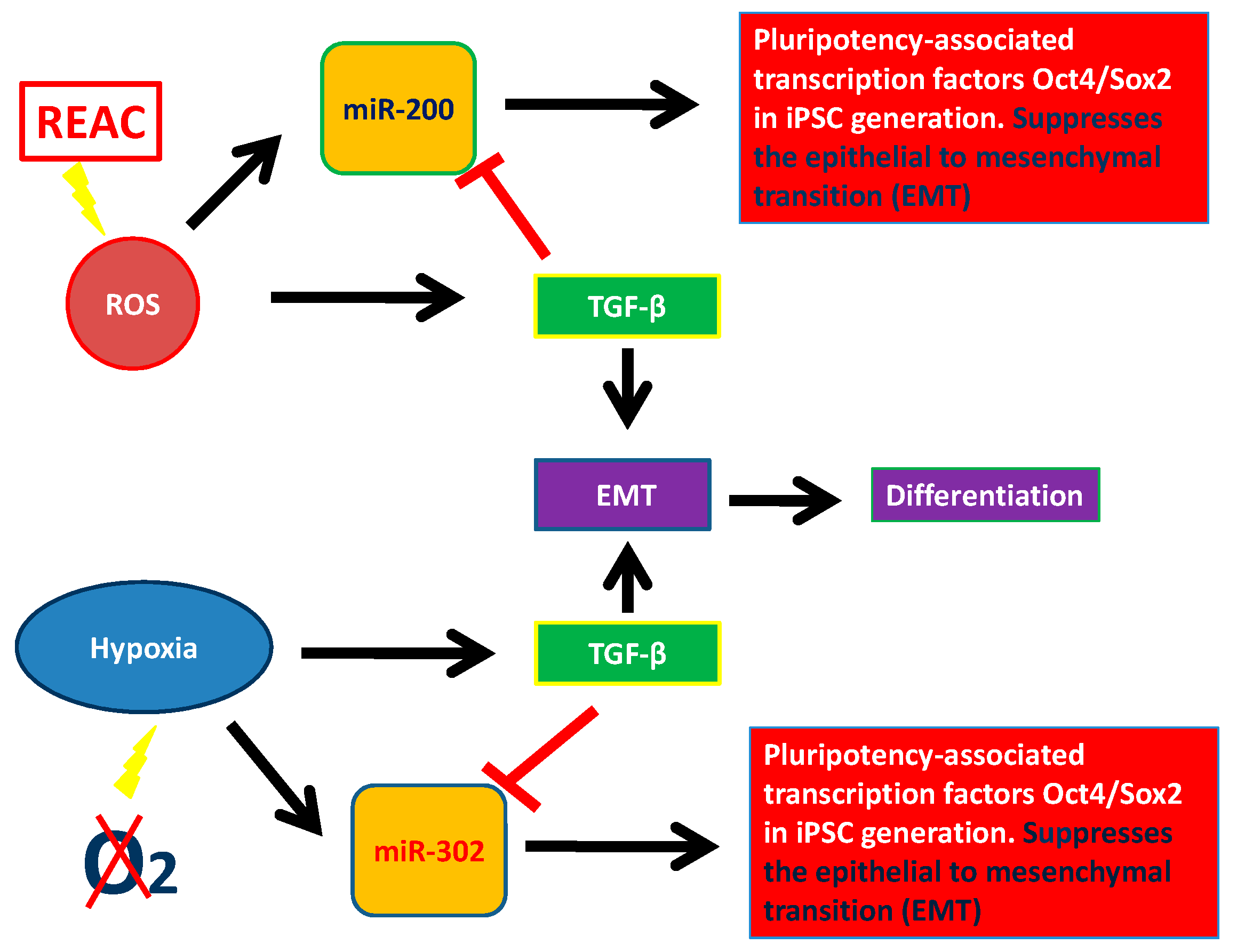
In this review, we described epigenetic factors modulating pluripotency and stem cell behavior, particularly the role of miRNAs. We considered two large miRNA families involved in plutipotency: miR-200 family and miR-302 family. We analyzed some features of stem cells which encompass miRNA recruitment, biophysical stimuli, such as electromagnetic fields and REAC conveyed radioelectric fields, and chemistry, like ROS and hypoxia. The REAC action bypassed a persistent reprogramming toward an induced pluripotent stem cell-like state and involved the transcriptional induction of the NADPH oxidase subunit Nox4 [112,113,114,115]. This finding demonstrates the feasibility of using a physical stimulus to afford the expression of pluripotentiality in human adult somatic cells up to the attainment of major complex lineages. It is believed that one possible mechanism involves the generation of ROS within the cell. Excessive concentration of ROS is considered to be cell destructive and results in inhibition of gene expression. In contrast, small amounts of ROS function as intracellular second messengers and activate signaling cascades involved in growth and differentiation of many cell types. This complicated regulatory mechanism is called “hormesis”. Hormesis can be considered an adaptive function characterized by a dose-dependent biphasic response, which occurs as a result of exposure to a very wide range of stimuli [142,143,144]. This intricate mechanism of activation and inhibition of miRNAs and their targets leads to a fine stem cell homeostasis. MiR-200 and miR-302 are pluripotency activators which trigger pluripotency genes (SOX2, OCT4, and NANOG). These two miRNAs families can be modulated by physical and chemical factors. As showed in Figure 2, ROS induction leads to TGF-β activation which acts blocking miR-200 and miR-302. Figure 2 shows the complex interplay occurring between ROS, miR-200, and TGF-β and between hypoxia, miR302, and TGF-β. This factor, is directly modulated by both of the above mentioned miRNAs, and acts itself by modulating cell differentiation and reprogramming, in turn downregulating both miRNA 200 and miRNA 302. These two major miRNA families are the basis of pluripotency induction by interacting with TGF-β, thus demonstrating that TGF-β, with or without the involvement of ROS, plays a crucial role in suppressing pluripotency, thus influencing stem cell behavior (Figure 3) [112]. Unravelling the hormetic factors, as for example ROS, and their role in influencing stem cells pluripotency and differentiation capabilities could pave the way to novel therapeutic approaches in regenerative medicine.
Acknowledgments
This work was funded by Banco di Sardegna Foundation, Project “Myrtus 2.0: from waste to resource (Bioactivity of myrtle by-products)”, and GUNA ATTRE (Advanced Therapies and Tissue REgeneration), Bologna, Italy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dupont, C.; Armant, D.R.; Brenner, C.A. Epigenetics: Definition, mechanisms and clinical perspective. Semin. Reprod. Med. 2009, 27, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Marson, A.; Levine, S.S.; Cole, M.F.; Frampton, G.M.; Brambrink, T.; Johnstone, S.; Guenther, M.G.; Johnston, W.K.; Wernig, M.; Newman, J.; et al. Connecting microRNA genes to the core transcriptional regulatory circuitry of embryonic stem cells. Cell 2008, 134, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Houbaviy, H.B.; Murray, M.F.; Sharp, P.A. Embryonic stem cell-specific MicroRNAs. Dev. Cell 2003, 5, 351–358. [Google Scholar] [CrossRef]
- Bar, M.; Wyman, S.K.; Fritz, B.R.; Qi, J.; Garg, K.S.; Parkin, R.K.; Kroh, E.M.; Bendoraite, A.; Mitchell, P.S.; Nelson, A.M.; et al. MicroRNA discovery and profiling in human embryonic stem cells by deep sequencing of small RNA libraries. Stem Cells 2008, 26, 2496–2505. [Google Scholar] [CrossRef] [PubMed]
- Morin, R.D.; O’Connor, M.D.; Griffith, M.; Kuchenbauer, F.; Delaney, A.; Prabhu, A.L.; Zhao, Y.; McDonald, H.; Zeng, T.; Hirst, M.; et al. Application of massively parallel sequencing to microRNA profiling and discovery in human embryonic stem cells. Genome Res. 2008, 18, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Selbach, M.; Schwanhausser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Berezikov, E.; Guryev, V.; van de Belt, J.; Wienholds, E.; Plasterk, R.H.; Cuppen, E. Phylogenetic shadowing and computational identification of human microRNA genes. Cell 2005, 120, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Blahna, M.T.; Hata, A. Smad-mediated regulation of microRNA biosynthesis. FEBS Lett. 2012, 586, 1906–1912. [Google Scholar] [CrossRef] [PubMed]
- Elston, R.; Inman, G.J. Crosstalk between p53 and tgf-beta signaling. J. Signal Transduct. 2012, 2012, 294097. [Google Scholar] [PubMed]
- Chang, K.C.; Wang, C.; Wang, H. Balancing self-renewal and differentiation by asymmetric division: Insights from brain tumor suppressors in Drosophila neural stem cells. Bioessays 2012, 34, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Shen, D.; Shen, J.; Gao, S.M.; Li, B.; Wong, C.; Feng, W.; Song, Y. The Super Elongation Complex Drives Neural Stem Cell Fate Commitment. Dev. Cell 2017, 40, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [PubMed]
- Hirai, H.; Karian, P.; Kikyo, N. Regulation of embryonic stem cell self-renewal and pluripotency by leukaemia inhibitory factor. Biochem. J. 2011, 438, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bi, Y.; Gao, S. Epigenetic regulation of somatic cell reprogramming. Curr. Opin. Genet. Dev. 2017, 46, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Butz, H.; Rácz, K.; Hunyady, L.; Patócs, A. Crosstalk between TGF-β signaling and the microRNA machinery. Trends Pharmacol. Sci. 2012, 33, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 2008, 455, 1124–1128. [Google Scholar] [CrossRef] [PubMed]
- Creemers, E.E.; Tijsen, A.J.; Pinto, Y.M. Circulating microRNAs: Novel biomarkers and extracellular communicators in cardiovascular disease? Circ. Res. 2012, 110, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Bar-Nur, O.; Russ, H.A.; Efrat, S.; Benvenisty, N. Epigenetic memory and preferential lineage-specific differentiation in induced pluripotent stemcells derived from human pancreatic islet beta cells. Cell Stem Cell 2011, 9, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.; Tilgner, K.; Saretzki, G.; Atkinson, S.P.; Stojkovic, M.; Moreno, R.; Przyborski, S.; Lako, M. Human induced pluripotent stem cell lines show stress defense mechanisms and mitochondrial regulation similar to those of human embryonic stem cells. Stem Cells 2010, 28, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Prigione, A.; Fauler, B.; Lurz, R.; Lehrach, H.; Adjaye, J. The senescence-related mitochondrial/oxidative stress pathway is repressed in human induced pluripotent stem cells. Stem Cells 2010, 28, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Folmes, C.D.; Nelson, T.J.; Martinez-Fernandez, A.; Arrell, D.K.; Lindor, J.Z.; Dzeja, P.P.; Ikeda, Y.; Perez-Terzic, C.; Terzic, A. Somatic oxidative bioenergetics transitions into pluripotency-dependent glycolysis to facilitate nuclear reprogramming. Cell Metab. 2011, 14, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Folmes, C.D.; Dzeja, P.P.; Nelson, T.J.; Terzic, A. Metabolic plasticity in stem cell homeostasis and differentiation. Cell Stem Cell 2012, 11, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Nuebel, E.; Daley, G.Q.; Koehler, C.M.; Teitell, M.A. Metabolic regulation in pluripotent stem cells during reprogramming and self-renewal. Cell Stem Cell 2012, 11, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Berardi, E.; Pues, M.; Thorrez, L.; Sampaolesi, M. miRNAs in ESC differentiation. Am. J. Physiol. Heart Circ. Physiol. 2012, 15, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Davis-Dusenbery, B.N.; Hata, A. Mechanisms of control of microRNA biogenesis. J. Biochem. 2010, 148, 381–392. [Google Scholar] [PubMed]
- Baek, D.; Villén, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The impact of microRNAs on protein output. Nature 2008, 455, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Lüningschrö, P.; Hauser, S.; Kaltschmidtab, B.; Kaltschmidta, C. MicroRNAs in pluripotency, reprogramming and cell fate induction. BBA-Mol. Cell Res. 2013, 8, 1894–1903. [Google Scholar]
- Min, P.K.; Chan, S.Y. The biology of circulating microRNAs in cardiovascular disease. Eur. J. Clin. Investig. 2015, 45, 860–874. [Google Scholar] [CrossRef] [PubMed]
- Maioli, M.; Santaniello, S.; Montella, A.; Bandiera, P.; Cantoni, S.; Bianchi, C.C.F.; Lionetti, V.; Rizzolio, F.; Marchesi, I.; Bagella, L.; et al. Hyaluronan esters drive Smad gene expression and signaling enhancing cardiogenesis in mouse embryonic and human mesenchymal stem cells. PLoS ONE 2010, 5, e15151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massagué, J. TGFβ signaling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Nakao, A.; Afrakhte, M.; Morén, A.; Nakayama, T.; Christian, J.L.; Heuchel, R.; Itoh, S.; Kawabata, M.; Heldin, N.E.; Heldin, C.H.; et al. Identification of Smad7, a TGFbeta-inducible antagonist of TGF-beta signaling. Nature 1997, 389, 631–635. [Google Scholar] [PubMed]
- Massagué, J.; Blain, S.W.; Lo, R.S. TGFbeta signaling in growth control, cancer, and heritable disorders. Cell 2000, 103, 295–309. [Google Scholar] [CrossRef]
- Sakaki-Yumoto, M.; Katsuno, Y.; Derynck, R. TGF-β family signaling in stem cells. Biochim. Biophys. Acta 2013, 1830, 2280–2296. [Google Scholar] [CrossRef] [PubMed]
- Pera, M.F.; Tam, P.P.L. Extrinsic regulation of pluripotent stem cells. Nature 2010, 465, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Warmflash, A.; Arduini, B.L.; Brivanlou, A.H. The molecular circuitry underlying pluripotency in embryonic stem cells. Wiley Interdiscip. Rev. Syst. Biol. Med. 2012, 4, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core Transcriptional Regulatory Circuitry in Human Embryonic Stem Cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Vallier, L.; Mendjan, S.; Brown, S.; Chng, Z.; Teo, A.; Smithers, L.E.; Trotter, M.W.B.; Cho, C.H.; Martinez, A.; Rugg-Gunn, P.; et al. Activin/Nodal signaling maintains pluripotency by controlling Nanog expression. Development 2009, 136, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.M.; Reynolds, D.; Cliff, T.; Ohtsuka, S.; Mattheyses, A.L.; Sun, Y.; Menendez, L.; Kulik, M.; Dalton, S. Signaling Network Crosstalk in Human Pluripotent Cells: A Smad2/3-Regulated Switch that Controls the Balance between Self-Renewal and Differentiation. Cell Stem Cell 2012, 10, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Fedele, M.; Cerchia, L.; Chiappetta, G. The Epithelial-to-Mesenchymal Transition in Breast Cancer: Focus on Basal-Like Carcinomas. Cancers 2017, 9, 134. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.F.; Nofech-Mozes, S.; Bayani, J.; Bartlett, J.M. EMT in breast carcinoma—A Review. J. Clin. Med. 2016, 5, 65. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 12, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridharan, N.M.R.; Xie, W.; Utikal, J.; Eminli, S.; Arnold, K.; Stadtfeld, M.; Yachechko, R.; Tchieu, J.; Jaenisch, R.; Plath, K.; et al. Directly reprogrammed fibroblasts show global epigenetic remodeling and widespread tissue contribution. Cell Stem Cell 2007, 1, 55–70. [Google Scholar]
- Guo, L.; Zhang, Y.; Zhang, L.; Huang, F.; Li, J.; Wang, S. MicroRNAs, TGF-β signaling, and the inflammatory microenvironment in cancer. Tumor Biol. 2016, 37, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Papagiannakopoulos, T.; Pan, G.; Thomson, J.A.; Kosik, K.S. MicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human embryonic stem cells. Cell 2009, 137, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Wellner, U.; Schubert, J.; Burk, U.C.; Schmalhofer, O.; Zhu, F.; Sonntag, A.; Waldvogel, B.; Vannier, C.; Darling, D.; zur Hausen, A.; et al. The EMT-activator ZEB1 promotes tum-origenicity by repressing stemness-inhibiting microRNAs. Nat. Cell Biol. 2009, 11, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Triboulet, R.; Chang, H.; LaPierre, R.J.; Gregory, R.I. Post-transcriptional control of DGCR8 expression by the Microprocessor. RNA 2009, 15, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Annes, J.P.; Munger, J.S.; Rifkin, D.B. Making sense of latent TGFβ activation. J. Cell Sci. 2003, 116, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Dijke, P.T.; Hill, C.S. New insights into TGF-β-Smadsignalling. Trends Biochem. Sci. 2004, 29, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.; Kietzmann, T. Reactive oxygen species and fibrosis: Further evidence of a significant liaison. Cell Tissue Res. 2016, 365, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J.; Gomis, R.R. The logic of TGFβ signaling. FEBS Lett. 2006, 580, 2811–2820. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.R.; Erler, J.T. Remodeling and homeostasis of the extracellular matrix: Implications for fibrotic diseases and cancer Disease. Models Mech. 2011, 4, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, S.; Takahashi, K. Induction of pluripotent stem cells from mouse fibroblast cultures. Tanpakushitsu Kakusan Koso 2006, 51, 2346–2351. [Google Scholar] [PubMed]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanley, J.; Rastegarlari, G.; Nathwani, A.C. An introduction to induced pluripotent stem cells. Br. J. Haematol. 2010, 151, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Anokye-Danso, F.; Snitow, M.; Morrisey, E.E. How microRNAs facilitate reprogramming to pluripotency. J. Cell Sci. 2012, 125, 4179–4787. [Google Scholar] [CrossRef] [PubMed]
- Razak, S.R.; Ueno, K.; Takayama, N.; Nariai, N.; Nagasaki, M.; Saito, R.; Koso, H.; Lai, C.; Murakami, M.; Tsuji, K.; et al. Profiling of microRNA in human and mouse ES and iPS cells reveals overlapping but distinct microRNA expression patterns. PLoS ONE 2013, 8, 73532. [Google Scholar] [CrossRef] [PubMed]
- Anokye-Danso, F.; Trivedi, C.M.; Juhr, D.; Gupta, M.; Cui, Z.; Tian, Y.; Zhang, Y.; Yang, W.; Gruber, P.J.; Epstein, J.A.; et al. Highly efficient miRNA-mediated reprogramming of mouse and human somatic cells to pluripotency. Cell Stem Cell 2011, 8, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Saint-André, V.; Federation, A.J. Models of human core transcriptional regulatory circuitries. Genome Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Duan, S.; Yi, F.; Yi, F.; Ocampo, A.; Liu, L.; Belmonte, J.C. Mitochondrial Regulation in Pluripotent Stem Cells. Cell Metab. 2013, 18, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Kaspi, H.; Chapnik, E.; Levy, M.; Beck, G.; Hornstein, E.; Soen, Y. Brief Report: miR-290–295 Regulate Embryonic Stem Cell Differentiation Propensities by Repressing Pax6. Stem Cells 2013, 31, 2266–2272. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Blelloch, R. Cell Cycle Regulation by MicroRNAs in Embryonic Stem Cells. Cancer Res. 2009, 15, 4093–4096. [Google Scholar] [CrossRef] [PubMed]
- Gambardella, G.; Carissimo, A.; Chen, A.; Cutillo, L.; Nowakowski, T.J.; di Bernardo, D.; Blelloch, R. The impact of microRNAs on transcriptional heterogeneity and gene co-expression across single embryonic stem cells. Nat. Commun. 2017, 8, 14126. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.M.; Cahan, P.; Shalek, A.K.; Satija, R.; DaleyKeyser, A.J.; Li, H.; Zhang, J.; Pardee, K.; Gennert, D.; Trombetta, J.J.; et al. Deconstructing transcriptional heterogeneity in pluripotent stem cells. Nature 2014, 516, 56–61. [Google Scholar] [CrossRef] [PubMed]
- DeCastro, A.J.; DiRenzo, J. miRNA function and modulation in stem cells and cancer stem cells. microRNA Diagn. Ther. 2014, 12–17. [Google Scholar] [CrossRef]
- Ambros, V.; Chen, X. The regulation of genes and genomes by small RNAs. Development 2007, 134, 1635–1641. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, L.A.; Dennis, L.M.; Gill, M.E.; Houbaviy, H.; Markoulaki, S.; Fu, D.; White, A.C.; Kirak, O.; Sharp, P.A.; Page, D.C.; et al. Mir-290-295 deficiency in mice results in partially penetrant embryonic lethality and germ cell defects. Proc. Natl. Acad. Sci. USA 2011, 108, 14163–14168. [Google Scholar] [CrossRef] [PubMed]
- Suh, M.R.; Lee, Y.; Kim, J.Y.; Kim, S.K.; Moon, S.H.; Lee, J.Y.; Cha, K.Y.; Chung, H.M.; Yoon, H.S.; Moon, S.Y.; et al. Human embryonic stem cells express a unique set of microRNAs. Dev. Biol. 2004, 270, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.L.; Chang, D.C.; Chang-Lin, S.; Lin, C.H.; Wu, D.T.; Chen, D.T.; Ying, S.Y. Mir-302 reprograms human skin cancer cells into a pluripotent ES-cell-like state. RNA 2008, 14, 2115–2124. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, N.; Ishii, H.; Nagano, H.; Haraguchi, N.; Dewi, D.L.; Kano, Y.; Nishikawa, S.; Tanemura, M.; Mimori, K.; Tanaka, F.; et al. Reprogramming of mouse and human cells to pluripotency using mature microRNAs. Cell Stem Cell 2011, 8, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Lin, C.P.; Ho, J.J.; He, X.; Okada, N.; Bu, P.; Zhong, Y.; Kim, S.Y.; Bennett, M.J.; Chen, C.; et al. miR-34 miRNAs provide a barrier for somatic cell reprogramming. Nat. Cell Biol. 2011, 13, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Z.; Jiang, J.; Xu, C.; Kang, J.; Xiao, L.; Wu, M.; Xiong, J.; Guo, X.; Liu, H. Endogenous miRNA Sponge lincRNA-RoR Regulates Oct4, Nanog, and Sox2 in Human Embryonic Stem Cell Self-Renewal. Dev. Cell 2013, 25, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Kagalwala, M.N.; Parker-Thornburg, J.; Adams, H.; Majumder, S. REST maintains selfrenewal and pluripotency of embryonic stem cells. Nature 2008, 453, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liang, J.; Ni, S.; Zhou, T.; Qing, X.; Li, H.; He, W.; Chen, J.; Li, F.; Zhuang, Q.; et al. A mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell 2010, 7, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Korpal, M.; Kang, Y. The emerging role of miR-200 family of microRNAs in epithelial-mesenchymal transition and cancer metastasis. RNA Biol. 2008, 5, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Samavarchi-Tehrani, P.; Golipour, A.; David, L.; Sung, H.K.; Beyer, T.A.; Datti, A.; Woltjen, K.; Nagy, A.; Wrana, J.L. Functional genomics reveals a BMP-driven mesenchymal-to-epithelial transition in the initiation of somatic cell reprogramming. Cell Stem Cell 2010, 7, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Dumont, N.; Wilson, M.B.; Crawford, Y.G.; Reynolds, P.A.; Sigaroudinia, M.; Tlsty, T.D. Sustained induction of epithelial to mesenchymal transition activates DNA methylation of genes silenced in basal-like breast cancers. Proc. Natl. Acad. Sci. USA 2008, 105, 14867–14872. [Google Scholar] [CrossRef] [PubMed]
- Vrba, L.; Jensen, T.J.; Garbe, J.C.; Heimark, R.L.; Cress, A.E.; Dickinson, S.; Stampfer, M.R.; Futscher, B.W. Role for DNA methylation in the regulation of miR-200c and miR-141 expression in normal and cancer cells. PLoS ONE 2010, 5, e8697. [Google Scholar] [CrossRef] [PubMed]
- Wiklund, E.D.; Gao, S.; Hulf, T.; Sibbritt, T.; Nair, S.; Costea, D.E.; Villadsen, S.B.; Bakholdt, V.; Bramsen, J.B.; Sørensen, J.A.; et al. MicroRNA alterations and associated aberrant DNA methylation patterns across multiple sample types in oral squamous cell carcinoma. PLoS ONE 2011, 6, e27840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiklund, E.D.; Bramsen, J.B.; Hulf, T.; Dyrskjøt, L.; Ramanathan, R.; Hansen, T.B.; Villadsen, S.B.; Gao, S.; Ostenfeld, M.S.; Borre, M.; et al. Coordinated epigenetic repression of the miR-200 family and miR-205 in invasive bladder cancer. Int. J. Cancer 2011, 128, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Gregory, P.A.; Bracken, C.P.; Smith, E.; Bert, A.G.; Wright, J.A.; Roslan, S.; Morris, M.; Wyatt, L.; Farshid, G.; Lim, Y.Y.; et al. An autocrine TGF-beta/ZEB/miR-200 signaling network regulates establishment and maintenance of epithelial-mesenchymal transition. Mol. Biol. Cell. 2011, 15, 1686–1698. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Han, Q.; Pei, D. EMT and MET as paradigms for cell fate switching. J. Mol Cell Biol. 2012, 4, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Lindahl-Allen, M.; Polytarchou, C.; Hirsch, H.A.; Tsichlis, P.N.; Struhl, K. Loss of miR-200 inhibition of Suz12 leads to polycomb-mediated repression required for the formation and maintenance of cancer stem cells. Mol. Cell 2010, 39, 761–772. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Zheng, Y.; Tenenhaus Dann, C. Mesenchymal to Epithelial Transition Mediated by CDH1 Promotes Spontaneous Reprogramming of Male Germline Stem Cells to Pluripotency. Stem Cell Rep. 2017, 8, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, N.; Park, S.-W.; Kim, H.; Park, H.-J.; Han, Y.-M. Lineage-specific Expression of miR-200 Family in Human Embryonic Stem Cells during In Vitro Differentiation. Int. J. Stem Cells 2017, 10, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, T.; Kuboki, T. Cellular Reprogramming Using Defined Factors and MicroRNAs. Stem Cells Int. 2016. [Google Scholar] [CrossRef] [PubMed]
- Hill, L.; Browne, G.; Tulchinsky, E. ZEB/miR-200 feedback loop: At the crossroads of signal transduction in cancer. Int. J. Cancer. 2013, 132, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Subramanyam, D.; Lamouille, S.; Judson, R.L.; Liu, J.Y.; Bucay, N.; Derynck, R.; Blelloch, R. Multiple targets of miR-302 and miR-372 promote reprogramming of human fibroblasts to induced pluripotent stem cells. Nat. Biotechnol. 2011, 29, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhu, X.; Dou, Y. The miR-302/367 cluster: A comprehensive update on its evolution and functions. Open Biol. 2015, 5, 150138. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Aksoy, M.; Shi, J.; Houbaviy, H. Evolution of the miR-/miR-371–373 Cluster Family Seed Repertoire. PLoS ONE 2014, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Roybon, L.; Lamas, N.J.; Garcia, A.D.; Yang, E.J.; Sattler, R.; Lewis, V.J.; Kim, Y.A.; Kachel, C.A.; Rothstein, J.D.; Przedborski, S.; et al. Human stem cell-derived spinal cord astrocytes with defined mature or reactive phenotypes. Cell Rep. 2013, 4, 1035–1048. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Baskerville, S.; Shenoy, A.; Babiarz, J.E.; Baehner, L.; Blelloch, R. Embryonic stem cell-specific microRNAs regulate the G1-S transition and promote rapid proliferation. Nat. Genet. 2008, 40, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.X.; Ravi, A.; Calabrese, J.M.; Medeiros, L.A.; Kirak, O.; Dennis, L.M.; Jaenisch, R.; Burge, C.B.; Sharp, P.A. A latent pro-survival function for the mir-290–295 cluster in mouse embryonic stem cells. PLoS Genet. 2011, 7, e1002054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesus, A.B.D.; Lucena-Aguilar, G.; Menendez, P. The miR-302–367 cluster as a potential stemness regulator in ESCs. Cell Cycle 2009, 8, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Card, D.A.; Hebbar, P.B.; Li, L.; Trotter, K.W.; Komatsu, Y.; Mishina, Y.; Archer, T.K. Oct4/Sox2-regulated miR-302 targets cyclin D1 in human embryonic stem cells. Mol. Cell. Biol. 2008, 28, 6426–6438. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Huttner, W.B.; Calegari, F. Cdk4/cyclinD1 overexpression in neural stem cells shortens G1, delays neurogenesis, and promotes the generation and expansion of basal progenitors. Cell Stem Cell 2009, 5, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.C.; Weng, S.C.; Chou, C.J.; Chang, T.T.; Tsai, W.J. Activation and proliferation signals in primary human T lymphocytes inhibited by ergosterol peroxide isolated from Cordyceps cicadae. Br. J. Pharmacol. 2003, 140, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.L.; Chang, D.C.; Lin, C.H.; Ying, S.Y.; Leu, D.; Wu, D.T.S. Regulation of somatic cell reprogramming through inducible mir-302 expression. Nucleic Acids Res. 2011, 39, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Prasain, N.; Chae, H.D.; Kim, Y.J.; Mantel, C.; Yoder, M.C.; Broxmeyer, H.E. Epigenetic regulation of NANOG by miR-302 cluster-MBD2 completes induced pluripotent stem cell reprogramming. Stem Cells 2013, 31, 666–681. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.W.; Gomez, E.W. Biomechanics of TGFbeta-induced epithelial–mesenchymal transition: Implications for fibrosis and cancer. Clin. Transl. Med. 2014, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Barroso-delJesus, A.; Lucena-Aguilar, G.; Sanchez, L.; Ligero, G.; Gutierrez-Aranda, I.; Menendez, P. The Nodal inhibitor Lefty is negatively modulated by the microRNA miR-302 in human embryonic stem cells. FASEB J. 2011, 25, 1497–1508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cui, W. Sox2, a key factor in the regulation of pluripotency and neural differentiation. World J. Stem Cells 2014, 6, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-S.; Yi, B.-R.; Kim, N.-H.; Choi, K.-C. Role of the epithelial–mesenchymal transition and its effects on embryonic stem cells. Exp. Mol. Med. 2014, 46, e108. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hutchins, A.P.; Chen, Y.; Li, S.; Shan, Y.; Liao, B.; Zheng, D.; Shi, X.; Li, Y.; Chan, W.-Y.; et al. A sequential EMT-MET mechanism drives the differentiation of human embryonic stem cells towards hepatocytes. Nat. Commun. 2017, 8, 15166. [Google Scholar] [CrossRef] [PubMed]
- Ichida, J.K.; Blanchard, J.; Lam, K.; Son, E.Y.; Chung, J.E.; Egli, D.; Loh, K.M.; Carter, A.C.; di Giorgio, F.P.; Koszka, K.; et al. A small-molecule inhibitor of TGF-Beta signaling replaces Sox2 in reprogramming by inducing nanog. Cell Stem Cell 2009, 5, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Young, J.L.; Suresh, R.; Chauhan, H.; Park, W.S.; Hong, S.-H.; Kim, K.-S. Dissecting microRNA-mediated regulation of stemness, reprogramming, and pluripotency. Cell Regen. 2016, 5, 2. [Google Scholar]
- Chan, Y.C.; Khanna, S.; Roy, S.; Sen, C.K. miR-200b Targets Ets-1 and Is Down-regulated by Hypoxia to Induce Angiogenic Response of Endothelial Cells. J. Biol. Chem. 2011, 286, 2047–2056. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.-C.; Yeh, C.-H. Inhibition of miR-302 Suppresses Hypoxia-Reoxygenation-Induced H9c2 Cardiomyocyte Death by Regulating Mcl-1 Expression. Oxid. Med. Cell. Longev. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ji, A.R.; Ku, S.Y.; Cho, M.S.; Kim, Y.Y.; Kim, Y.J.; Oh, S.K.; Kim, S.H.; Moon, S.Y.; Choi, Y.M. Reactive oxygen species enhance differentiation of human embryonic stem cells into mesendodermal lineage. Exp. Mol. Med. 2010, 42, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Maioli, M.; Rinaldi, S.; Santaniello, S.; Castagna, A.; Pigliaru, G.; Gualini, S.; Cavallini, C.; Fontani, V.; Ventura, C. Radio Electric Conveyed Fields Directly Reprogram Human Dermal Skin Fibroblasts Toward Cardiac, Neuronal, and Skeletal Muscle-Like Lineages. Cell Transplant. 2013, 22, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Maioli, M.; Rinaldi, S.; Pigliaru, G.; Santaniello, S.; Basoli, V.; Castagna, A.; Fontani, V.; Ventura, C. REAC technology and hyaluron synthase 2, an interesting network to slow down stem cell senescence. Sci. Rep. 2016, 6, 28682. [Google Scholar] [CrossRef] [PubMed]
- Maioli, M.; Rinaldi, S.; Santaniello, S.; Castagna, A.; Pigliaru, G.; Delitala, A.; Bianchi, F.; Tremolada, C.; Fontani, V.; Ventura, C. Radioelectric asymmetric conveyed fields and human adipose-derived stem cells obtained with a nonenzymatic method and device: A novel approach to multipotency. Cell Transplant. 2014, 23, 1489–1500. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.; Quan, X.; Kim, S.; Lengner, C.; Park, J.K.; Kim, J. Electromagnetic fields mediate efficient cell reprogramming into a pluripotent state. ACS Nano 2014, 8, 10125–10138. [Google Scholar] [CrossRef] [PubMed]
- Razavi, S.; Salimi, M.; Shahbazi-Gahrouei, D.; Karbasi, S.; Kermani, S. Extremely low-frequency electromagnetic field influences the survival and proliferation effect of human adipose derived stem cells. Adv. Biomed. Res. 2014, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.; Oh, J.; Song, J.; Choi, H.; Yoo, J.; Park, G.Y.; Han, J.; Chang, Y.; Park, H.; Kim, H.; et al. Generation of Integration-Free Induced Neurons Using Graphene Oxide-Polyethylenimine. Small 2017, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wu, W.; Chen, X.; He, C.; Liu, X. Effect of Pulsed Electromagnetic Field with Different Frequencies on the Proliferation, Apoptosis and Migration of Human Ovarian Cancer Cells. J. Biomed. Eng. 2012, 29, 291–295. [Google Scholar]
- Juutilainen, J. Developmental Effects of Electromagnetic Fields. Bioelectromagnetics 2005, 26, S107–S115. [Google Scholar] [CrossRef] [PubMed]
- Levin, M. Large-Scale Biophysics: Ion Flows and Regeneration. Trends Cell Biol. 2007, 17, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Hiraki, Y.; Endo, N.; Takigawa, M.; Asada, A.; Takahashi, H.; Suzuki, F. Enhanced Responsiveness to Parathyroid Hormoneand Induction of Functional Differentiation of Cultured Rabbit Costal Chondrocytes by a Pulsed Electromagnetic Field. Biochim. Biophys. Acta 1987, 931, 94–100. [Google Scholar] [CrossRef]
- Kang, K.S.; Hong, J.M.; Kang, J.A.; Rhie, J.W.; Jeong, Y.H.; Cho, D.W. Regulation of Osteogenic Differentiation of Human Adipose-Derived Stem Cells by Controlling Electromagnetic Field Conditions. Exp. Mol. Med. 2013, 45, e6. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Xu, T.; Guo, F.; Yin, W.; Peng, T. The Effects ofHigh-Intensity Pulsed Electromagnetic Field on Proliferationand Differentiation of Neural Stem Cells of Neonatal Rats in vitro. Med. Sci. 2009, 29, 732–736. [Google Scholar]
- Sun, L.Y.; Hsieh, D.K.; Lin, P.C.; Chiu, H.T.; Chiou, T.W. Pulsed electromagnetic fields accelerate proliferation and osteogenic gene expression in human bone marrow mesenchymal stem cells during osteogenic differentiation. Bioelectromagnetics 2010, 31, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, Z.; Fisher, M.; Lohmann, C.H.; Simon, B.J.; Boya, B.D. Osteoprotegerin (OPG) production by cells in the osteoblast lineage is regulated by pulsed electromagnetic fields in cultures grown on calcium phosphate substrates. Ann. Biomed. Eng. 2009, 37, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, Z.; Simon, B.J.; Duran, M.A.; Barabino, G.; Chaudhri, R.; Boyan, B.D. Pulsed electromagnetic fields enhance BMP-2 dependent osteoblastic differentiation of human mesenchymal stem cells. J. Orthop. Res. 2008, 26, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Janssen-Heininger, Y.M.; Mossman, B.T.; Heintz, N.H.; Forman, H.J.; Kalyanaraman, B.; Finkel, T.; Stamler, J.S.; Rhee, S.G.; van der Vliet, A. Redox-based regulation of signal transduction: Principles, pitfalls, and promises. Free Radic. Biol. Med. 2008, 45, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Maziarz, A.; Kocan, B.; Bester, M.; Budzik, S.; Cholewa, M.; Ochiya, T.; Banas, A. How electromagnetic fields can influence adult stem cells: Positive and negative impacts. Stem Cell Res. Ther. 2016, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signaling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Nagai, H.; Lin, T.M.; Peterson, R.E.; Tohyama, C.; Kobayashi, T.; Nohara, K. Organic Chemicals Adsorbed onto Diesel Exhaust Particles Directly Alter the Differentiation of Fetal Thymocytes Through Arylhydrocarbon Receptor but Not Oxidative Stress Responses. J. Immunotoxicol. 2006, 3, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Milhavet, O.; Lemaitre, J.M. Senescent-Derived Pluripotent Stem Cells Are Able to Redifferentiate into Fully Rejuvenated Cells. In Tumor Dormancy, Quiescence, and Senescence; Springer: Dordrecht, The Netherlands, 2014; Volume 2, pp. 265–276. [Google Scholar]
- Teslaa, T.; Teitell, M.A. Pluripotent stem cell energy metabolism: An update. EMBO J. 2015, 34, 138–153. [Google Scholar] [CrossRef] [PubMed]
- Ezashi, T.; Das, P.; Roberts, R.M. Low O2 tensions andthe prevention of differentiation of hES cells. Proc. Natl. Acad. Sci. USA 2005, 102, 4783–4788. [Google Scholar] [CrossRef] [PubMed]
- Westfall, S.D.; Sachdev, S.; Das, P.; Hearne, L.B.; Hannink, M.; Roberts, R.M.; Ezashi, T. Identification of Oxygen-Sensitive Transcriptional Programs in Human Embryonic Stem Cells. Stem Cells Dev. 2008, 17, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Dando, I.; Cordani, M.; DallaPozza, E.; Biondani, G.; Donadelli, M.; Palmieri, M. Antioxidant Mechanisms and ROS-Related MicroRNAs in Cancer Stem CellsOxid. Med. Cell Longev. 2015, 425708. [Google Scholar]
- Vaupel, P.; Mayer, A. Hypoxia in cancer: Significance and impact on clinical outcome. Cancer Metastasis Rev. 2007, 26, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Höckel, M.; Vaupel, P. Tumor hypoxia: Definitions and current clinical, biologic, and molecular aspects. J. Natl. Cancer Inst. 2001, 93, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.-M.; Desai, L.P. Reciprocal regulation of TGF-β and reactive oxygen species: A perverse cycle for fibrosis. Redox Biol. 2015, 6, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Magenta, A.; Cencioni, C.; Fasanaro, P.; Zaccagnini, G.; Greco, S.; Sarra-Ferraris, G.; Antonini, A.; Martelli, F.; Capogrossi, M.C. miR-200c is upregulated by oxidative stress and induces endothelial cell apoptosis and senescence via ZEB1 inhibition. Cell Death Differ. 2011, 18, 1628–1639. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.L.; Siriwardane, M.; Almeida-Poradaa, G.; Poradaa, C.D.; Brinkc, P.; Christa, G.J.; Harrisona, B.S. The effect of low-frequency electromagnetic field on human bone marrow stem/progenitor cell differentiation. Stem Cell Res. 2015, 15, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. Defining hormesis. Hum. Exp. Toxicol. 2002, 21, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Shin, K.K.; Lee, A.L.; Kim, Y.S.; Park, H.J.; Park, Y.K.; Bae, Y.C.; Jung, J.S. MicroRNA-302 induces proliferation and inhibits oxidant-induced cell death in human adipose tissue-derived mesenchymal stem cells. Cell Death Dis. 2014, 5, e1385. [Google Scholar] [CrossRef] [PubMed]
- Harnoss, J.M.; Strowitzki, M.J.; Radhakrishnan, P.; Platzer, L.K.; Harnoss, J.C.; Hank, T.; Cai, J.; Ulrich, A.; Schneider, M. Therapeutic inhibition of prolyl hydroxylase domain-containing enzymes in surgery: Putative applications and challenges. Hypoxia 2015, 3, 1–14. [Google Scholar] [PubMed]
Figure 1.
TGF-B activation and SMAD-mediated miRNA downregulation.
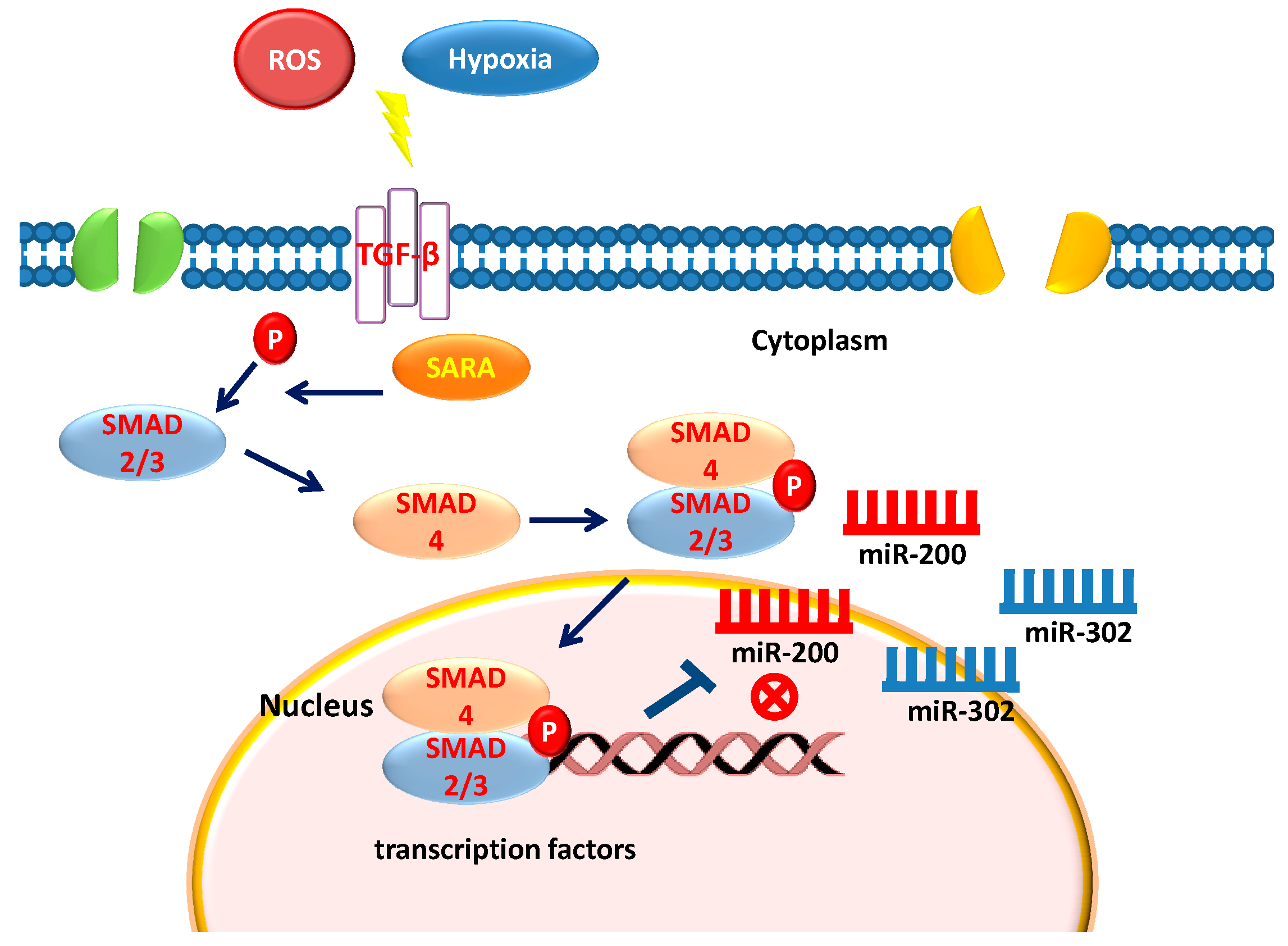
Figure 2.
Influence of different factors on somatic cells and induced pluripotent stem cells (IPSCs) fate.
Figure 2.
Influence of different factors on somatic cells and induced pluripotent stem cells (IPSCs) fate.
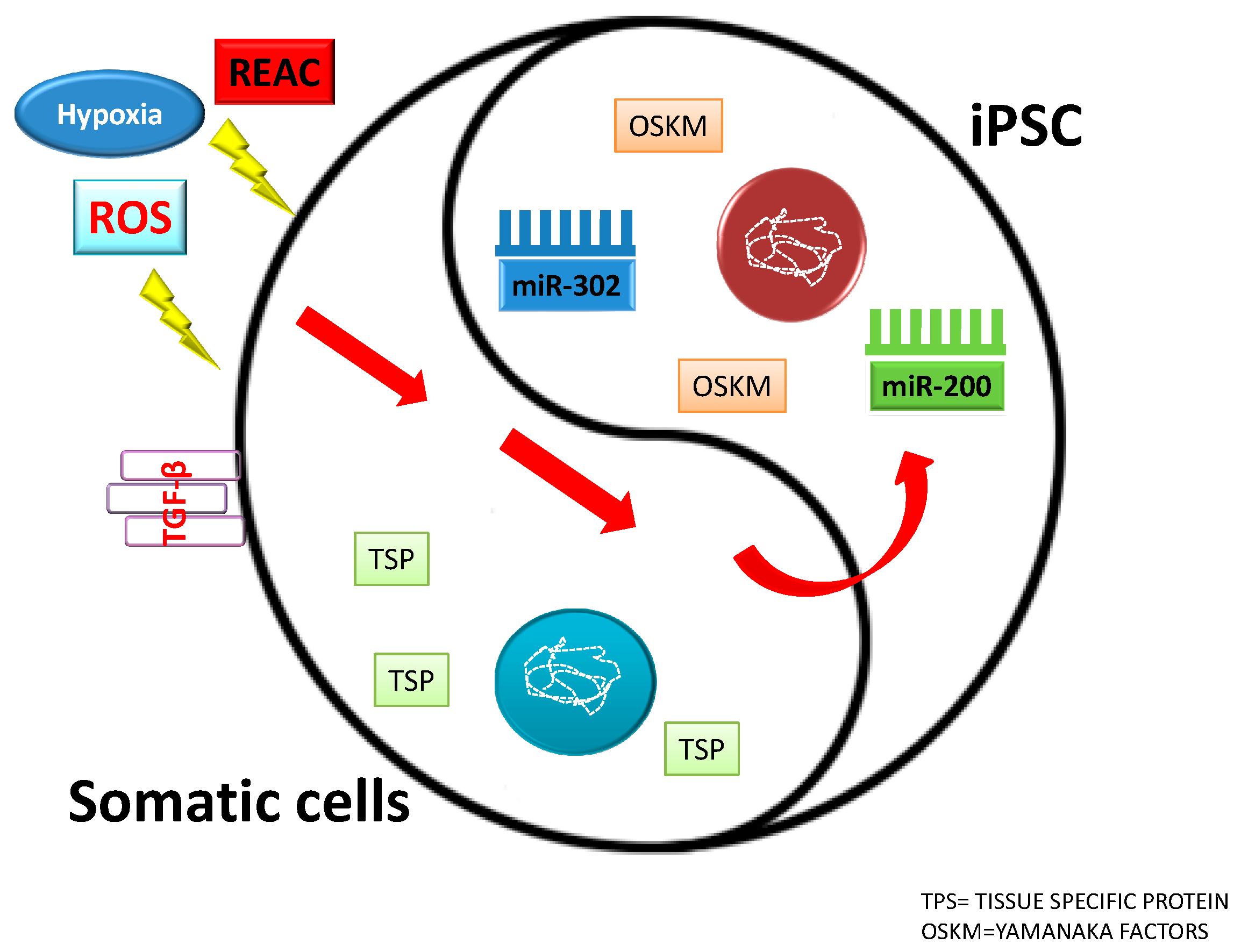
Figure 3.
Interaction between miR-200, miR-302, physical stimuli, TGF-β and pluripotency.


{kind=link}
{kind=link}
{kind=link}
Table 1.
MiRNA regulation on stemness.
| miRNA | Influence on Pluripontecy | Influence on Differentiation | Inhibition of EMT /MET |
|---|---|---|---|
| miR-200 family | yes | no | yes |
| miR-302 family | yes | no | yes |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Balzano, F.; Cruciani, S.; Basoli, V.; Santaniello, S.; Facchin, F.; Ventura, C.; Maioli, M. MiR200 and miR302: Two Big Families Influencing Stem Cell Behavior. Molecules 2018, 23, 282. https://doi.org/10.3390/molecules23020282
AMA Style
Balzano F, Cruciani S, Basoli V, Santaniello S, Facchin F, Ventura C, Maioli M. MiR200 and miR302: Two Big Families Influencing Stem Cell Behavior. Molecules. 2018; 23(2):282. https://doi.org/10.3390/molecules23020282
Chicago/Turabian StyleBalzano, Francesca, Sara Cruciani, Valentina Basoli, Sara Santaniello, Federica Facchin, Carlo Ventura, and Margherita Maioli. 2018. "MiR200 and miR302: Two Big Families Influencing Stem Cell Behavior" Molecules 23, no. 2: 282. https://doi.org/10.3390/molecules23020282