Action of Phytochemicals on Insulin Signaling Pathways Accelerating Glucose Transporter (GLUT4) Protein Translocation

Abstract
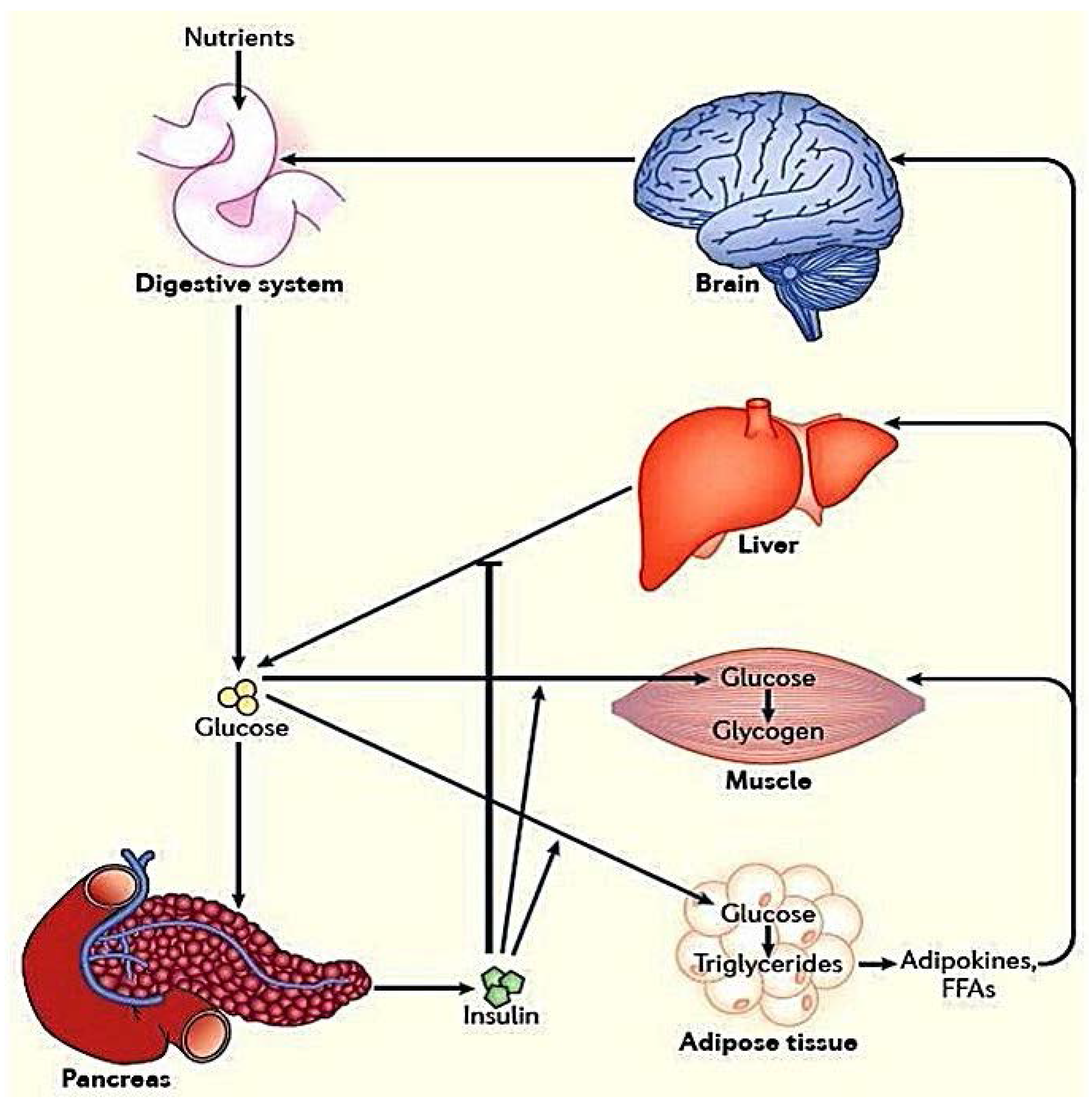
:1. Introduction
2. Phytochemicals
GLUT Proteins: Structure and Function
3. GLUT-4 Targets Insulin Signaling Pathway
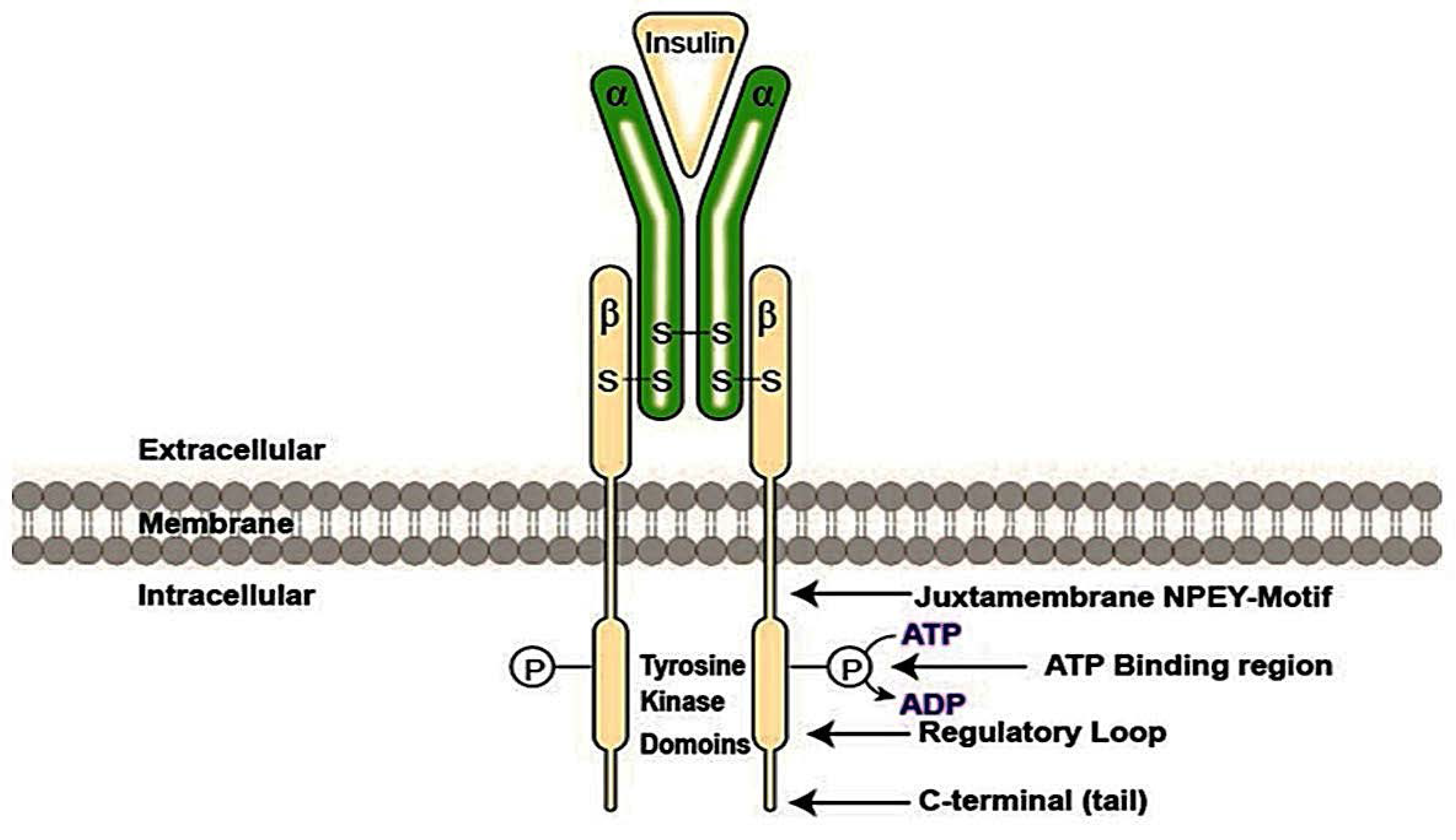
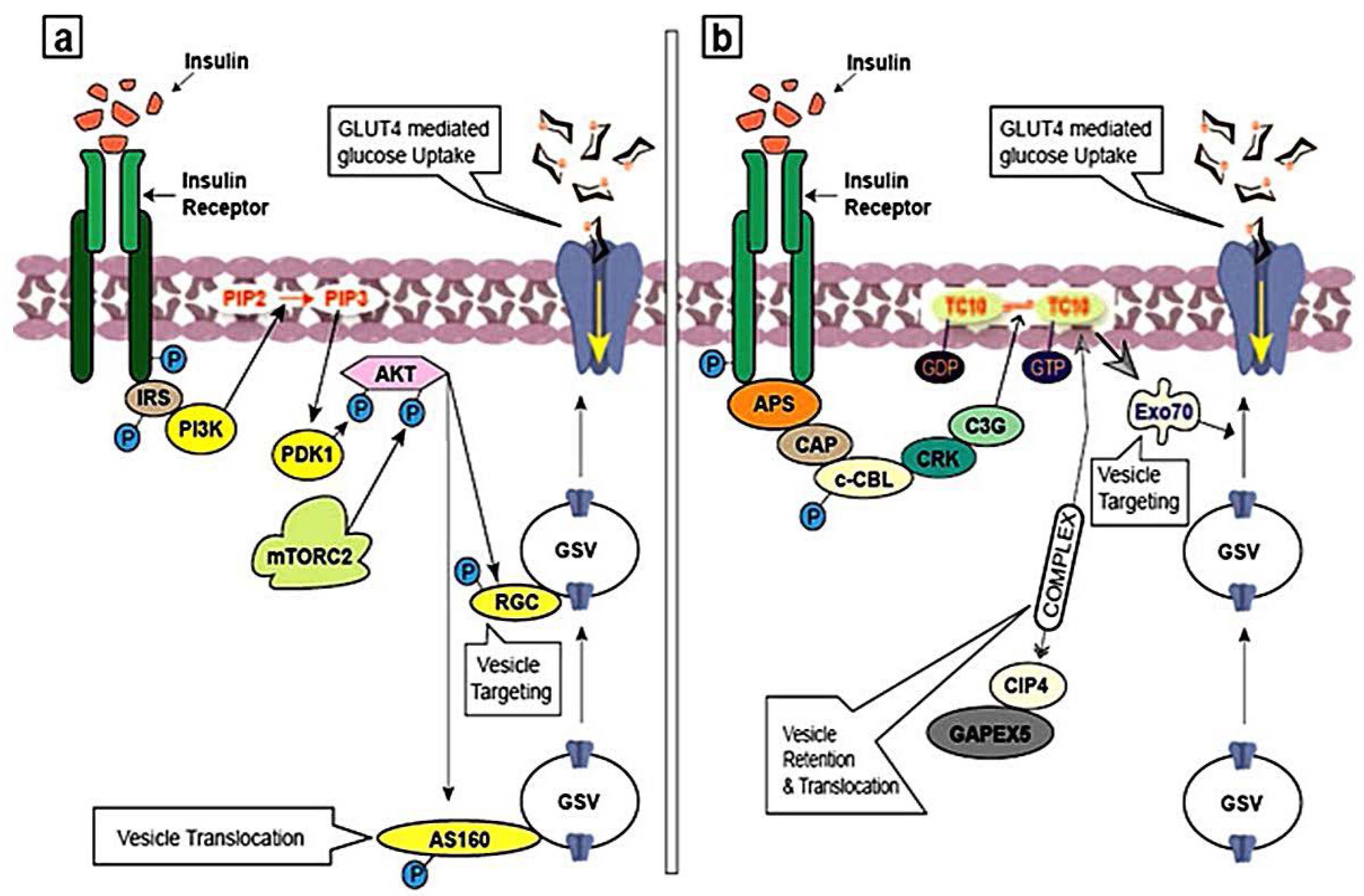
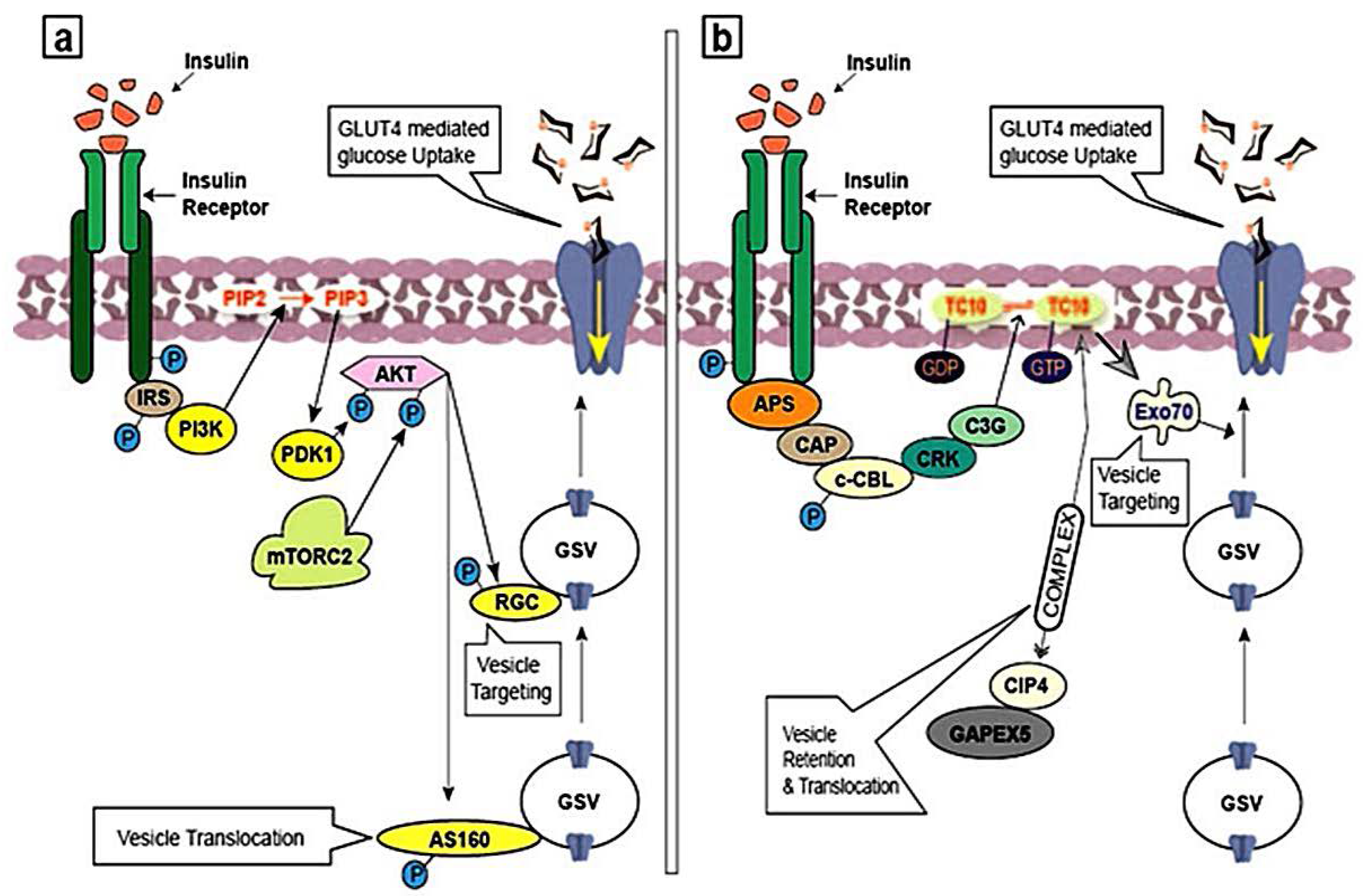
3.1. Insulin Signaling Pathway and GLUT4 Trafficking
3.2. Activation of PI3K Pathway
3.3. Activation of APS Pathway
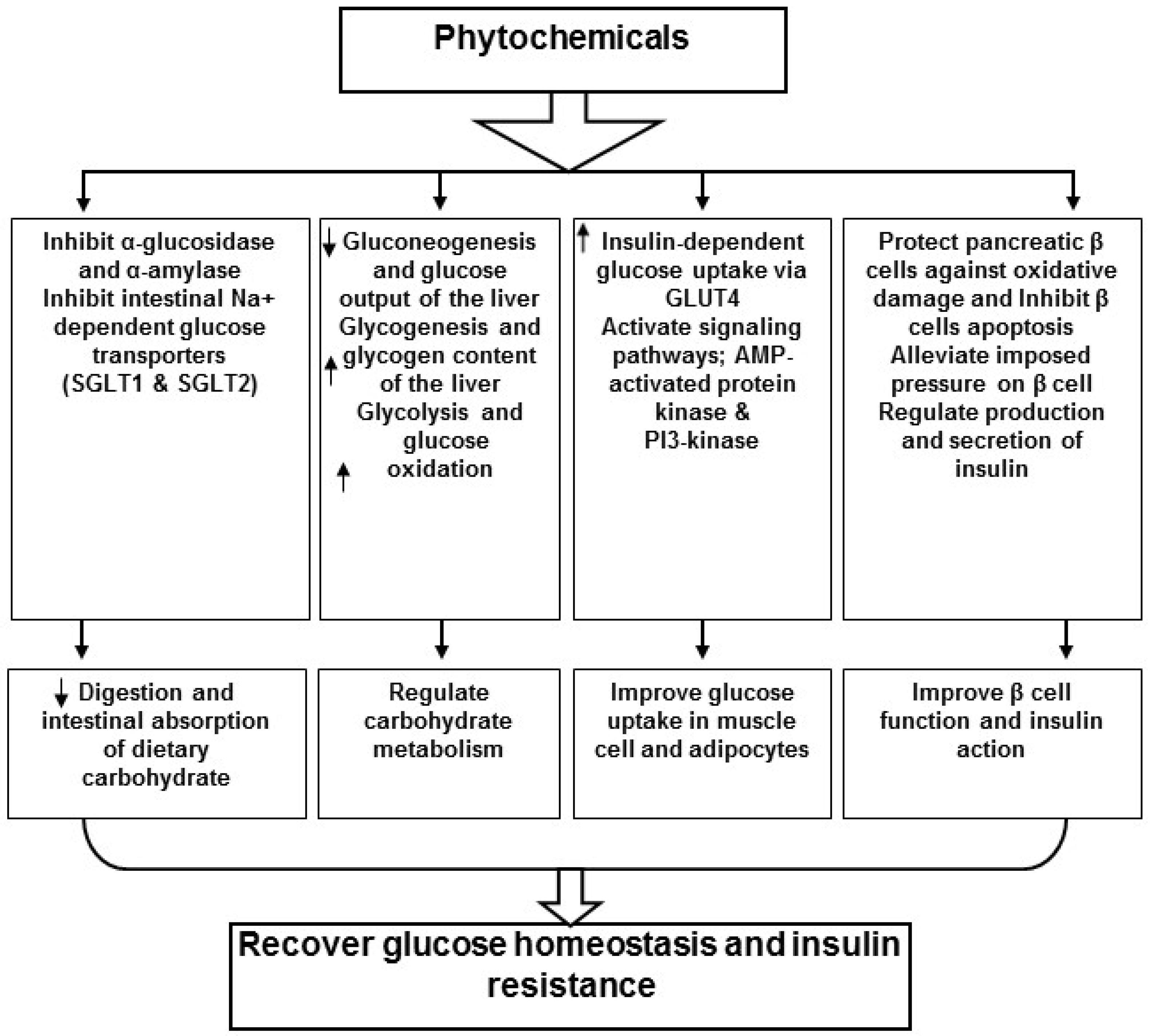
4. Phytochemicals and Insulin Signaling Pathway
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Ahn, M.Y.; Katsanakis, K.D.; Bheda, F.; Pillay, T.S. Primary and essential role of the adaptor protein APS for recruitment of both c-Cbl and its associated protein CAP in insulin signaling. J. Biol. Chem. 2004, 279, 21526–21532. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, M.; Levine, R.; Klein, S.; Huddlestun, B. The action of insulin on the distribution of galactose in eviscerated nephrectomized dogs. J. Biol. Chem. 1949, 179, 985. [Google Scholar]
- Morgan, H.E.; Henderson, M.J.; Regen, D.M.; Park, C.R. Regulation of glucose uptake in muscle. I. The effects of insulin and anoxia on glucose transport and phosphorylation in the isolated, perfused heart of normal rats. J. Biol. Chem. 1961, 236, 253–261. [Google Scholar] [PubMed]
- Suzuki, K.; Kono, T. Evidence that insulin causes translocation of glucose transport activity to the plasma membrane from an intracellular storage site. Proc. Natl. Acad. Sci. USA 1980, 77, 2542–2545. [Google Scholar] [CrossRef] [PubMed]
- Cushman, S.W.; Wardzala, L.J. Potential mechanism of insulin action on glucose transport in the isolated rat adipose cell. Apparent translocation of intracellular transport systems to the plasma membrane. J. Biol. Chem. 1980, 255, 4758–4762. [Google Scholar] [PubMed]
- Joost, H.G.; Bell, G.I.; Best, J.D.; Birnbaum, M.J.; Charron, M.J.; Chen, Y.T.; Doege, H.; James, D.E.; Lodish, H.F.; Moley, K.H.; et al. Nomenclature of the GLUT/SLC2A family of sugar/polyol transport facilitators. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 974–976. [Google Scholar] [CrossRef] [PubMed]
- Bouche, C.; Serdy, S.; Kahn, C.R.; Goldfine, A.B. The cellular fate of glucose and its relevance in type 2 diabetes. Endocr. Rev. 2004, 25, 807–830. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M.; Caruso, C.; Baldwin, S.A.; Panico, M.; Blench, I. Sequence and structure of a human glucose transporter. Science 1985, 229, 941–945. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Aspects Med. 2013, 34, 121–138. [Google Scholar] [CrossRef] [PubMed]
- James, D.E.; Strube, M.; Mueckler, M. Molecular cloning and characterization of an insulin-regulatable glucose transporter. Nature 1989, 338, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, M.J. Identification of a novel gene encoding an insulin-responsive glucose transporter protein. Cell 1989, 57, 305–315. [Google Scholar] [CrossRef]
- Furtado, L.M.; Somwar, R.; Sweeney, G.; Niu, W.; Klip, A. Activation of the glucose transporter GLUT4 by insulin. Biochem. Cell Biol. 2002, 80, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.T.; Kanzaki, M.; Pessin, J.E. Regulated membrane trafficking of the insulin-responsive glucose transporter 4 in adipocytes. Endocr. Rev. 2004, 25, 177–204. [Google Scholar] [CrossRef] [PubMed]
- Simpson, F.; Jonathan, P.W.; David, E.J. GLUT4—At the cross roads between membrane trafficking and signal transduction. Traffic 2001, 2, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signaling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Lazar, D.F.; Wiese, R.J.; Brady, M.J.; Mastick, C.C.; Waters, S.B.; Yamauchi, K.; Pessin, J.E.; Cuatrecasas, P.; Saltiel, A.R. Mitogen-activated protein kinase kinase inhibition does not block the stimulation of glucose utilization by insulin. J. Biol. Chem. 1995, 270, 20801–20807. [Google Scholar] [CrossRef] [PubMed]
- Polak, P.; Cybulski, N.; Feige, J.N.; Auwerx, J.; Rüegg, M.A.; Hall, M.N. Adipose-specific knockout of raptor results in lean mice with enhanced mitochondrial respiration. Cell Metab. 2008, 8, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Larsen, C.J. The Nobel Prize in physiology and medicine 1989. J. Michael Bishop and Harold E. Varmus. Pathol. Biol. (Paris) 1989, 37, 1077–1078. [Google Scholar]
- Mishra, R.K.; Wei, C.; Hresko, R.C.; Bajpai, R.; Heitmeier, M.; Matulis, S.M.; Nooka, A.K.; Rosen, S.T.; Hruz, P.W.; Schiltz, G.E.; et al. In Silico Modeling-based Identification of Glucose Transporter 4 (GLUT4)-selective Inhibitors for Cancer Therapy. J. Biol. Chem. 2015, 290, 14441–14453. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Bajpai, R.; Sharma, H.; Heitmeier, M.; Jain, A.D.; Matulis, S.M.; Nooka, A.K.; Mishra, R.K.; Hruz, P.W.; Schiltz, G.E.; et al. Development of GLUT4-selective antagonists for multiple myeloma therapy. Eur. J. Med. Chem. 2017, 139, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Sesti, G. Pathophysiology of insulin resistance. Best Practice. Res. Clin. Endocrinol. Metab. 2006, 20, 665–679. [Google Scholar]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry, 5th ed.; W H Freeman: New York, NY, USA, 2002. [Google Scholar]
- Soumyanath, A. Traditional Medicines for Modern Times—Antidiabetic Plants; CRC Press: Boca Raton, Floria, USA, 2005. [Google Scholar]
- Borris, R.P. Natural products research: Perspectives from a major pharmaceutical company. J. Ethnopharmacol. 1996, 51, 29–38. [Google Scholar] [CrossRef]
- Das, U.N. GLUT-4, tumor necrosis factor, essential fatty acids and daf-genes and their role in insulin resistance and non-insulin dependent diabetes mellitus. Prostaglandins Leukot. Essent. Fat. Acids 1999, 60, 13–20. [Google Scholar] [CrossRef]
- Kanaujia, A.; Duggar, R.; Pannakal, S.T.; Yadav, S.S.; Katiyar, C.K.; Bansal, V.; Anand, S.; Sujatha, S.; Lakshmi, B.S. Insulinomimetic activity of two new gallotannins from the fruits of Capparis moonii. Bioorg. Med. Chem. 2010, 18, 3940–3945. [Google Scholar] [CrossRef] [PubMed]
- Sangeetha, K.N.; Sujatha, S.; Muthusamy, V.S.; Anand, S.; Nithya, N.; Velmurugan, D.; Balakrishnan, A.; Lakshmi, B.S. 3β-taraxerol of Mangifera indica, a PI3K dependent dual activator of glucose transport and glycogen synthesis in 3T3-L1 adipocytes. Biochim. Biophys. Acta 2010, 1800, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wu, K.; Mao, X.; Wu, Y.; Ouyang, J. Astragalus polysaccharide improves insulin sensitivity in KKAy mice: Regulation of PKB/GLUT4 signaling in skeletal muscle. J. Ethnopharmacol. 2010, 127, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Scazzocchio, B.; Varì, R.; Filesi, C.; D’Archivio, M.; Santangelo, C.; Giovannini, C.; Iacovelli, A.; Silecchia, G.; Volti, G.; Galvano, F.; et al. Cyanidin-3-O-b-Glucoside and Protocatechuic Acid Exert Insulin-Like Effects by Upregulating PPARg Activity in Human Omental Adipocytes. Diabetes 2011, 6, 2234–2244. [Google Scholar] [CrossRef] [PubMed]
- Cheong, S.H.; Furuhashi, K.; Ito, K.; Nagaoka, M.; Yonezawa, T.; Miura, Y.; Yagasaki, K. Daidzein promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and improves glucose homeostasis in Type 2 diabetic model mice. J. Nutr. Biochem. 2014, 25, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.F.; Dixit, P.; Jaiswal, N.; Tamrakar, A.K.; Srivastava, A.K.; Maurya, R. Chemical constituents of Kigelia pinnata twigs and their GLUT4 translocation modulatory effect in skeletal muscle cells. Fitoterapia 2012, 83, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Latha, R.C.R.; Daisy, P. Insulin-secretagogue, antihyperlipidemic and other protective effects of gallic acid isolated from Terminalia bellerica Roxb. in streptozotocin-induced diabetic rats. Chem. Biol. Interact. 2011, 189, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, P.K.; Doble, M. Effect of Natural Products on Commercial Oral Antidiabetic Drugs in Enhancing 2-Deoxyglucose Uptake by 3T3-L1 Adipocytes. Ther. Adv. Endocrinol. Metab. 2011, 3, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Girón, M.D.; Sevillano, N.; Salto, R.; Haidour, A.; Manzano, M.; Jiménez, M.L.; Rueda, R.; López-Pedrosa, J.M. Salacia oblonga extract increases glucose transporter 4-mediated glucose uptake in L6 rat myotubes: Role of mangiferin. Clin. Nutr. 2009, 28, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K.; Kim, M.Y.; Onodera, A.; Matsue, H. α-Glucosidase inhibitory and antihyperglycemic effects of polyphenols in the fruit of Viburnum dilatatum Thunb. Agric. Food Chem. 2006, 54, 4588–4592. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K. Antidiabetic and antioxidant effects of polyphenols in brown alga Ecklonia stolonifera in genetically diabetic KK-Ay mice. Plant Foods Hum. Nutr. 2008, 63, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, C.; Artacho, R.; Giménez, R. Beneficial effects of green tea—A review. J. Am. Coll. Nutr. 2006, 25, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Thirunavukkarasu, M.; Penumathsa, S.V.; Koneru, S.; Juhasz, B.; Zhan, L.; Otani, H.; Bagchi, D.; Das, D.K.; Maulik, N. Resveratrol alleviates cardiac dysfunction in streptozotocin-induced diabetes: Role of nitric oxide, thioredoxin, and heme oxygenase. Free Radic. Biol. Med. 2007, 43, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Moskaug, J.Ø.; Carlsen, H.; Myhrstad, M.; Blomhoff, R. Molecular imaging of the biological effects of quercetin and quercetin-rich foods. Mech. Ageing Dev. 2004, 125, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Thorens, B.; Mueckler, M. Glucose transporters in the 21st Century. Am. J. Physiol. Endocrinol. Metab. 2010, 298, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Pao, S.S.; Paulsen, I.T.; Saier, M.H. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar] [PubMed]
- Bryant, N.J.; Goovers, R.; James, D.E. Regulated transport of the glucose transporter GLUT4. Nat. Rev. Mol. Cell Biol. 2002, 3, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Zisman, A.; Peroni, O.D.; Abel, E.D.; Michael, M.D.; Mauvais-Jarvis, F.; Lowell, B.B.; Wojtaszewski, J.F.; Hirshman, M.F.; Virkamaki, A.; Goodyear, L.J.; et al. Targeted disruption of the glucose transporter 4 selectively in muscle causes insulin resistance and glucose intolerance. Nat. Med. 2000, 6, 924–928. [Google Scholar] [PubMed]
- Zhao, F.Q.; Keating, A.F. Expression and regulation of glucose transporters in the bovine mammary gland. J. Dairy Sci. 2007, 90, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Hatakeyama, H.; Watanabe, T.M.; Sato, M.; Higuchi, H.; Kanzaki, M. Identification of three distinct functional sites of insulin-mediated GLUT4 trafficking in adipocytes using quantitative single molecule imaging. Mol. Biol. Cell 2010, 21, 2721–2731. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Reinisch, K.; Ferro-Novick, S. Coats, tethers, Rabs, and SNAREs work together to mediate the intracellular destination of a transport vesicle. Dev. Cell 2007, 12, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Leto, D.; Saltiel, A.R. Regulation of glucose transport by insulin: Traffic control of GLUT4. Nat. Rev. Mol. Cell Biol. 2012, 13, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Nolan, J.H.; Jeffrey, S.E. Signaling, cytoskeletal and membrane mechanisms regulating GLUT4 exocytosis. Trends Endocrinol. Metab. 2011, 22, 110–116. [Google Scholar]
- Feener, E.P.; Backer, J.M.; King, G.L.; Wilden, P.A.; Sun, X.J.; Kahn, C.R.; White, M.F. Insulin stimulates serine and tyrosine phosphorylation in the juxtamembrane region of the insulin receptor. J. Biol. Chem. 1993, 268, 11256–11264. [Google Scholar] [PubMed]
- White, M.F.; Kahn, C.R. The insulin signaling system. J. Biol. Chem. 1994, 269, 1–5. [Google Scholar] [PubMed]
- Saltiel, A.R.; Pessin, J.E. Insulin signaling in microdomains of the plasma membrane. Traffic 2003, 4, 711–716. [Google Scholar] [CrossRef] [PubMed]
- McClain, D.A.; Maegawa, H.; Lee, J.; Dull, T.J.; Ulrich, A.; Olefsky, J.M. A mutant insulin receptor with defective tyrosine kinase displays no biological activity and does not undergo endocytosis. J. Biol. Chem. 1987, 262, 14663–14671. [Google Scholar] [PubMed]
- Whiteman, E.L.; Cho, H.; Birnbaun, M.J. Role of Akt/protein kinase B in metabolism. Trends Endocrinol. Metab. 2002, 13, 444–451. [Google Scholar] [CrossRef]
- Gonzalez, E.; McGraw, T.E. Insulin signaling diverges into Akt-dependent and -independent signals to regulate the recruitment/docking and the fusion of GLUT4 vesicles to the plasma membrane. Mol. Biol. Cell 2006, 17, 4484–4493. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.W.; Leto, D.; Xiong, T.; Yu, G.; Cheng, A.; Decker, S.; Saltiel, A.R. A Ral GAP complex links PI 3-kinase/Akt signaling to RalA activation in insulin action. Mol. Biol. Cell 2011, 22, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Okada, S.; Kanzaki, M.; Elmendorf, J.S.; Coker, K.J.; Ceresa, B.P.; Syu, L.J.; Noda, Y.; Saltiel, A.R.; Pessin, J.E. Synip: A novel insulin-regulated syntaxin 4-binding protein mediating GLUT4 translocation in adipocytes. Mol. Cell. 1999, 3, 751–760. [Google Scholar] [CrossRef]
- Katome, T.; Obata, T.; Matsushima, R.; Masuyama, N.; Cantley, L.C.; Gotoh, Y.; Kishi, K.; Shiota, H.; Ebina, Y. Use of RNA interference-mediated gene silencing and adenoviral overexpression to elucidate the roles of AKT/protein kinase B isoforms in insulin actions. J. Biol. Chem. 2003, 278, 28312–28323. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.F.; Young, P.W.; Yonezawa, K.; Kasuga, M.; Holman, G.D. Inhibition of the translocation of GLUT1 and GLUT4 in 3T3-L1 cells by the phosphatidylinositol 3-kinase inhibitor, wortmannin. Biochem. J. 1994, 300, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Kohn, A.D.; Summers, S.A.; Birnbaum, M.J.; Roth, R.A. Expression of a constitutively active Akt Ser/Thr kinase in 3T3-L1 adipocytes stimulates glucose uptake and glucose transporter 4 translocation. J. Biol. Chem. 1996, 271, 31372–31378. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.; Ramm, G.; Lopez, J.A.; James, D.E. Rapid activation of Akt2 is sufficient to stimulate GLUT4 translocation in 3T3-L1 adipocytes. Cell Metab. 2008, 7, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Wang, Y.; Fan, J.; Chen, Y.; Ji, W.; Qu, A.; Xu, P.; James, D.E.; Xu, T. Dissecting multiple steps of GLUT4 trafficking and identifying the sites of insulin action. Cell Metab. 2007, 5, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Yamada, E.; Okada, S.; Saito, T.; Ohshima, K.; Sato, M.; Tsuchiya, T.; Uehara, Y.; Shimizu, H.; Mori, M. Akt2 phosphorylates Synip to regulate docking and fusion of GLUT4-containing vesicles. J. Cell Biol. 2005, 168, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Ribon, V.; Printen, J.A.; Hoffman, N.G.; Kay, B.K.; Saltiel, A.R. A novel, multifuntional c-Cbl binding protein in insulin receptor signaling in 3T3-L1 adipocytes. Mol. Cell. Biol. 1998, 18, 872–879. [Google Scholar] [CrossRef]
- Liu, J.; Kimura, A.; Baumann, C.A.; Saltiel, A.R. APS facilitates c-Cbl tyrosine phosphorylation and GLUT4 translocation in response to insulin in 3T3-L1 adipocytes. Mol. Cell. Biol. 2002, 22, 3599–3609. [Google Scholar] [CrossRef] [PubMed]
- Ribon, V.; Hubbell, S.; Herrera, R.; Saltiel, A.R. The product of the cbl oncogene forms stable complexes in vivo with endogenous Crk in a tyrosine phosphorylation-dependent manner. Mol. Cell. Biol. 1996, 16, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, B.S.; Feller, S.M.; Hanafusa, H. Four proline-rich sequences of the guanine-nucleotide exchange factor C3G bind with unique specificity to the first Src homology 3 domain of Crk. J. Biol. Chem. 1994, 269, 32781–32787. [Google Scholar] [PubMed]
- Chiang, S.H.; Baumann, C.A.; Kanzaki, M.; Thurmond, D.C.; Watson, R.T.; Neudauer, C.L.; Macara, I.G.; Pessin, J.E.; Saltiel, A.R. Insulin-stimulated GLUT4 translocation requires the CAP-dependent activation of TC10. Nature 2001, 410, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Chiang, S.H.; Saltiel, A.R. TC10α is required for insulin-stimulated glucose uptake in adipocytes. Endocrinology 2007, 148, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Ribon, V.; Johnson, J.H.; Camp, H.S.; Saltiel, A.R. Thiazolidinediones and insulin resistance: Peroxisome proliferator activated receptor gamma activation stimulates expression of the CAP gene. Proc. Natl. Acad. Sci. USA 1998, 95, 14751–14756. [Google Scholar] [CrossRef]
- Mitra, P.; Zheng, X.; Czech, M.P. RNAi-based analysis of CAP, Cbl, and CrkII function in the regulation of GLUT4 by insulin. J. Biol. Chem. 2004, 279, 37431–37435. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.Á.; Park, J.G.; Jiang, Z.Y.; Holik, J.J.; Mitra, P.; Semiz, S.; Guilherme, A.; Powelka, A.M.; Tang, X.; Virbasius, J.; et al. Analysis of insulin signalling by RNAi-based gene silencing. Biochem. Soc. Trans. 2004, 32, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.Y.; Bai, X.; Chen, X.H.; Fang, J.B.; Liu, S.H.; Chen, J.C. Anti-diabetic effect of methylswertianin and bellidifolin from Swertia punicea Hemsl. and its potential mechanism. Phytomedicine 2010, 17, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Naowaboot, J.; Pannangpetch, P.; Kukongviriyapan, V.; Prawan, A.; Kukongviriyapan, U.; Itharat, A. Mulberry leaf extract stimulates glucose uptake and GLUT4 translocation in rat adipocytes. Am. J. Chin. Med. 2012, 40, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Baldea, L.A.N.; Martineau, L.C.; Benhaddou-Andaloussi, A.; Arnason, J.T.; Lévy, É.; Haddad, P.S. Inhibition of intestinal glucose absorption by anti-diabetic medicinal plants derived from the James Bay Cree traditional pharmacopeia. J. Ethnopharmacol. 2010, 132, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Elchebly, M.; Payette, P.; Michaliszyn, E.; Cromlish, W.; Collins, S.; Loy, A.L.; Normandin, D.; Cheng, A.; Himms-Hagen, J.; Chan, C.C.; et al. Increased insulin sensitivity and obesity resistance in mice lacking the protein tyrosine phosphatase-1B gene. Science 1999, 283, 1544–1548. [Google Scholar] [CrossRef] [PubMed]
- Arya, A.; Looi, C.Y.; Wong, W.F.; Noordin, M.I.; Nyamathulla, S.; Mustafa, M.R.; Mohd, M.A. In vitro antioxidant, PTP-1B inhibitory effects and in vivo hypoglycemic potential of selected medicinal plants. Int. J. Pharmacol. 2013. [Google Scholar] [CrossRef]
- Zhang, W.; Hong, D.; Zhou, Y.; Zhang, Y.; Shen, Q.; Li, J.Y.; Hu, L.H.; Li, J. Ursolic acid and its derivative inhibit protein tyrosine phosphatase 1B, enhancing insulin receptor phosphorylation and stimulating glucose uptake. Biochim. Biophys. Acta 2006, 1760, 1505–1512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Zhang, W.; Hong, D.; Shi, L.; Shen, Q.; Li, J.Y.; Li, J.; Hu, L.H. Oleanolic acid and its derivatives: New inhibitor of protein tyrosine phosphatase 1B with cellular activities. Bioorg. Med. Chem. 2008, 16, 8697–8705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Salituro, G.; Szalkowski, D.; Li, Z.; Zhang, Y.; Royo, I.; Vilella, D.; Dı́ez, M.T.; Pelaez, F.; Ruby, C.; et al. Discovery of a small molecule insulin mimetic with antidiabetic activity in mice. Science 1999, 284, 974–977. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Li, Z.; Park, K.; Deng, L.; Pai, A.; Zhong, L.; Pirrung, M.C.; Webster, N.J. Identification of novel orally available small molecule insulin mimetics. J. Pharmacol. Exp. Ther. 2007, 323, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Arya, A.; Al-Obaidi, M.M.J.; Shahid, N.; Noordin, M.I.B.; Looi, C.Y.; Wong, W.F.; Khaing, S.L.; Mustafa, M.R. Synergistic effect of quercetin and quinic acid by alleviating structural degeneration in the liver, kidney and pancreas tissues of STZ-induced diabetic rats: A mechanistic study. Food Chem. Toxicol. 2014, 71, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Taha, H.; Arya, A.; Paydar, M.; Looi, C.Y.; Wong, W.F.; Murthy, C.V.; Noordin, M.I.; Ali, H.M.; Mustafa, A.M.; Hadi, A.H.A. Upregulation of insulin secretion and downregulation of pro-inflammatory cytokines, oxidative stress and hyperglycemia in STZ-nicotinamide-induced type 2 diabetic rats by Pseuduvaria monticola bark extract. Food Chem. Toxicol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Arya, A.; Al-Obaidi, M.M.J.; Karim, R.B.; Taha, H.; Khan, A.K.; Shahid, N.; Sayem, A.S.; Looi, C.Y.; Mustafa, M.R.; Mohd, M.A.; Ali, H.M. Extract of Woodfordia fruticosa flowers ameliorates hyperglycemia, oxidative stress and improves β-cell function in streptozotocin-nicotinamide induced diabetic rat. J. Ethnopharmacol. 2015, 175, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Coman, C.; Rugina, O.D.; Socaciu, C. Plants and natural compounds with antidiabetic action. Not. Bot. Horti Agrobot. Cluj-Napoca 2012, 40, 314. [Google Scholar]
- Kim, K.J.; Lee, M.S.; Jo, K.; Hwang, J.K. Piperidine alkaloids from Piper retrofractum Vahl. protect against high-fat diet-induced obesity by regulating lipid metabolism and activating AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2011, 411, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Gunawan-Puteri, M.D.; Kawabata, J. Novel α-glucosidase inhibitors from Macaranga tanarius leaves. Food Chem. 2010, 123, 384–389. [Google Scholar] [CrossRef]
- Xie, W.; Tanabe, G.; Matsuoka, K.; Amer, M.F.; Minematsu, T.; Wu, X.; Yoshikawa, M.; Muraoka, O. Role of the side chain stereochemistry in the α-glucosidase inhibitory activity of kotalanol, a potent natural α-glucosidase inhibitor. Bioorg. Med. Chem. 2011, 19, 2252–2262. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, O.; Morikawa, T.; Miyake, S.; Akaki, J.; Ninomiya, K.; Yoshikawa, M. Quantitative determination of potent α-glucosidase inhibitors, salacinol and kotalanol, in Salacia species using liquid chromatography-mass spectrometry. J. Pharmaceut. Biomed. 2010, 52, 770–773. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemicals | Plants | Effects on Insulin Signaling Pathways Accelerating GLUT4 | References |
|---|---|---|---|
| Resveratrol | Numerous plants | Induces AKT and VEGF as well as the expression of GLUT4 in muscle of STZ-induced diabetic rats via PI3K-AKT pathways | [25] |
| Gallotannins | Capparis moon | Increases GLUT4 and PI3K mRNA expression in the L6 cells | [26] |
| 3β-taraxerol | Mangifera indica | Activates glucose transport through the translocation of GLUT4, mediating by PI3K dependent activation of AKT protein | [27] |
| Astragalus polysaccharide | Astragalus membranaceus | Regularizes insulin-stimulated PKB-Ser473 phosphorylation and GLUT4 translocation | [28] |
| Cyanidin-3-O-β-glucoside and protocatechuic acid | Numerous plants | Insulin-like activity enhancing GLUT4 translocation and adiponectin secretion | [29] |
| Daidzein | Glycine max | Activates AMPK followed by GLUT4 translocation and enhances glucose homeostasis | [30] |
| Iridoid, catalpol, specioside and verminoside | Kigelia pinnata | Stimulation of GLUT4 trafficking to cell surface | [31] |
| Gallic acid | Myriophyllum spicatum | Decreases blood glucose and also enhances glucose uptake through the compartmentalization of GLUT4 to the plasma membrane | [32] |
| Berberine and vanillic acid | Berberis aristata and numerous plants | Improves the translocation of GLUT4 via AMPK-dependent pathway | [33] |
| Mangiferin | Salacia oblonga | Enhances the GLUT4 proteins expression and translocation of this glucose transporter to the surface of L6-myocites and 3T3-adipocytes | [34] |
| Isoform | Gene | Chromosome | Tissue Distribution | Substrate | Function |
|---|---|---|---|---|---|
| GLUT1 | SLC2A1 | 1p34.2 | Ubiquitous, erythrocytes | Glucose, galactose, dehydroacetic acid | Glucose uptake of tissues, glucose transportation through blood barriers into tissues |
| GLUT2 | SLC2A2 | 3q26.2 | Brain, intestine, liver, beta-cells, kidney | Glucose, fructose, galactose, dehydroacetic acid, glucosamine | Facilitated transporter of glucose and fructose, low attraction with large capacity |
| GLUT3 | SLC2A3 | 12p13.31 | Brain | Glucose, galactose, dehydroacetic acid | Neuronal uptake, facilitated transporter of glucose with strong affinity |
| GLUT4 | SLC2A4 | 17p13 | Skeletal and heart muscles, adipocytes | Glucose, dehydroacetic acid | Insulin dependent transporters, responsible for insulin resistance |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayem, A.S.M.; Arya, A.; Karimian, H.; Krishnasamy, N.; Ashok Hasamnis, A.; Hossain, C.F. Action of Phytochemicals on Insulin Signaling Pathways Accelerating Glucose Transporter (GLUT4) Protein Translocation. Molecules 2018, 23, 258. https://doi.org/10.3390/molecules23020258
Sayem ASM, Arya A, Karimian H, Krishnasamy N, Ashok Hasamnis A, Hossain CF. Action of Phytochemicals on Insulin Signaling Pathways Accelerating Glucose Transporter (GLUT4) Protein Translocation. Molecules. 2018; 23(2):258. https://doi.org/10.3390/molecules23020258
Chicago/Turabian StyleSayem, Abu Sadat Md, Aditya Arya, Hamed Karimian, Narendiran Krishnasamy, Ameya Ashok Hasamnis, and Chowdhury Faiz Hossain. 2018. "Action of Phytochemicals on Insulin Signaling Pathways Accelerating Glucose Transporter (GLUT4) Protein Translocation" Molecules 23, no. 2: 258. https://doi.org/10.3390/molecules23020258