Enhancing 2-Ketogluconate Production of Pseudomonas plecoglossicida JUIM01 by Maintaining the Carbon Catabolite Repression of 2-Ketogluconate Metabolism

Abstract
:1. Introduction
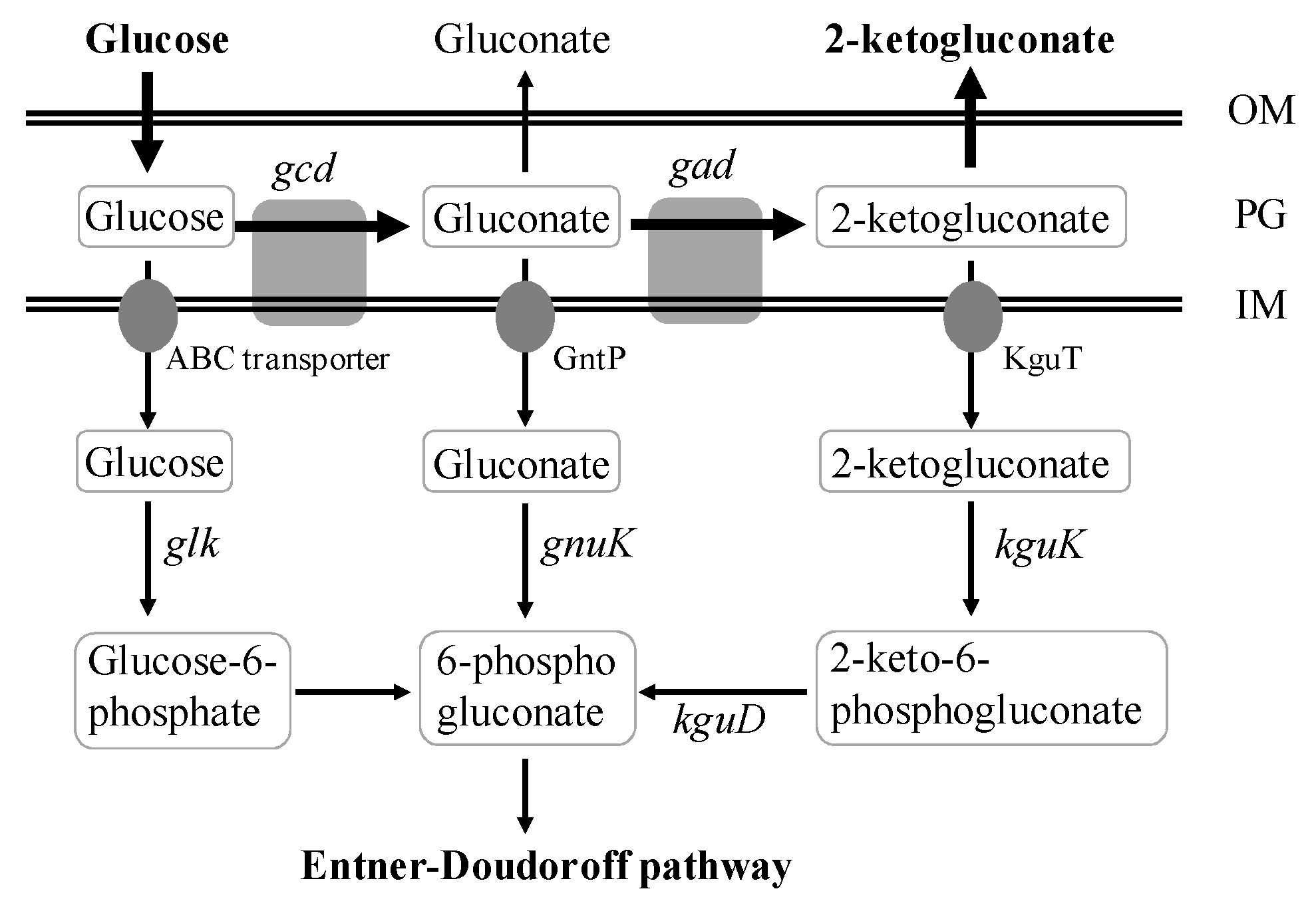
2. Results and Discussion
2.1. 2KGA Production and Consumption by P. plecoglossicida JUIM01 at Low Initial Glucose Concentration
2.2. Transcription and Proteomics Studies on 2KGA Consumption by P. plecoglossicida JUIM01
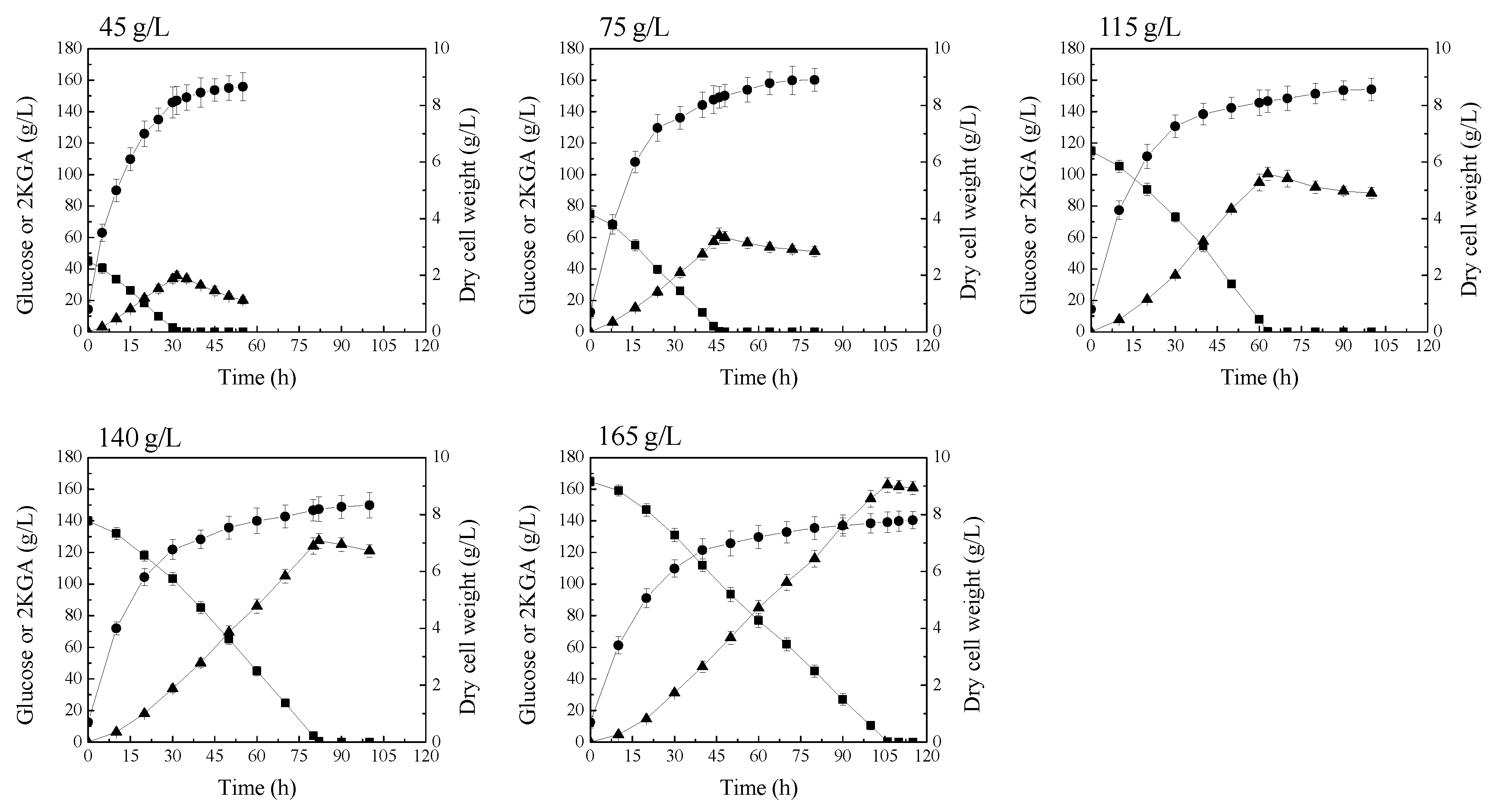
2.3. Attenuation of 2KGA Consumption by Increasing the Supply of Glucose
2.4. Fed-Batch Fermentation of P. plecoglossicida JUIM01 for 2KGA Production
3. Materials and Methods
3.1. Microorganism and Cultivation Conditions
3.2. Analytical Methods
3.3. Transcriptional Analysis
3.4. Whole-Cell Protein Extraction, Two-Dimensional Gel Electrophoresis, and Protein Identification
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sun, W.J.; Liu, C.F.; Yu, L.; Cui, F.J.; Zhou, Q.; Yu, S.L.; Sun, L. A novel bacteriophage KSL-1 of 2-Keto-gluconic acid producer Pseudomonas fluorescens K1005: Isolation, characterization and its remedial action. BMC Microbiol. 2012, 12, 127. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Xiao, F.; Wei, Z.; Cui, F.; Yu, L.; Yu, S.; Zhou, Q. Non-sterile and buffer-free bioconversion of glucose to 2-keto-gluconic acid by using Pseudomonas fluorescens AR4 free resting cells. Process Biochem. 2015, 50, 493–499. [Google Scholar] [CrossRef]
- Sun, W.J.; Zhou, Y.Z.; Zhou, Q.; Cui, F.J.; Yu, S.L.; Sun, L. Semi-continuous production of 2-keto-gluconic acid by Pseudomonas fluorescens AR4 from rice starch hydrolysate. Bioresour. Technol. 2012, 110, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Kiefler, I.; Bringer, S.; Bott, M. Metabolic engineering of Gluconobacter oxydans 621H for increased biomass yield. Appl. Microbiol. Biotechnol. 2017, 101, 5453–5467. [Google Scholar] [CrossRef] [PubMed]
- Krajewski, V.; Simic, P.; Mouncey, N.J.; Bringer, S.; Sahm, H.; Bott, M. Metabolic engineering of Gluconobacter oxydans for improved growth rate and growth yield on glucose by elimination of gluconate formation. Appl. Environ. Microbiol. 2010, 76, 4369–4376. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wei, D.; Shi, J.; Mojović, L.; Han, Z.; Hao, J. Two-stage fermentation for 2-Ketogluconic acid production by Klebsiella pneumoniae. J. Microbiol. Biotechnol. 2014, 24, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wang, Q.; Luan, F.; Man, Z.; Cui, F.; Qi, X. The role of kguT gene in 2-ketogluconate-producing Pseudomonas plecoglossicida JUIM01. Appl. Biochem. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Li, T.; Wang, B.; Liu, J.; Du, H.; Feng, H. Production of 2-Keto-D-gluconic acid by metabolically engineered Gluconobacter suboxydans. China Biotechnol. 2014, 34, 97–106. [Google Scholar]
- Li, K.; Mao, X.; Liu, L.; Lin, J.; Sun, M.; Wei, D.; Yang, S. Overexpression of membrane-bound gluconate-2-dehydrogenase to enhance the production of 2-keto-d-gluconic acid by Gluconobacter oxydans. Microb. Cell Fact. 2016, 15, 121. [Google Scholar] [CrossRef] [PubMed]
- del Castillo, T.; Ramos, J.L.; Rodríguez-Herva, J.J.; Fuhrer, T.; Sauer, U.; Duque, E. Convergent peripheral pathways catalyze initial glucose catabolism in Pseudomonas putida: Genomic and flux analysis. J. Bacteriol. 2007, 189, 5142–5152. [Google Scholar] [CrossRef] [PubMed]
- Nikel, P.I.; Chavarría, M.; Fuhrer, T.; Sauer, U.; de Lorenzo, V. Pseudomonas putida KT2440 strain metabolizes glucose through a cycle formed by enzymes of the Entner-Doudoroff, Embden-Meyerhof-Parnas, and Pentose Phosphate Pathways. J. Biol. Chem. 2015, 290, 25920–25932. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jeon, C.O.; Park, W. Dual regulation of zwf-1 by both 2-keto-3-deoxy-6-phosphogluconate and oxidative stress in Pseudomonas putida. Microbiology 2008, 154, 3905–3916. [Google Scholar] [CrossRef] [PubMed]
- Fuhrer, T.; Fischer, E.; Sauer, U. Experimental identification and quantification of glucose metabolism in seven bacterial species. J. Bacteriol. 2005, 187, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- del Castillo, T.; Ramos, J.L. Simultaneous catabolite repression between glucose and toluene metabolism in Pseudomonas putida is channeled through different signaling pathways. J. Bacteriol. 2007, 189, 6602–6610. [Google Scholar] [CrossRef] [PubMed]
- Rojo, F. Carbon catabolite repression in Pseudomonas: Optimizing metabolic versatility and interactions with the environment. FEMS Microbiol. Rev. 2010, 34, 658–684. [Google Scholar] [CrossRef] [PubMed]
- Sonnleitner, E.; Abdou, L.; Haas, D. Small RNA as global regulator of carbon catabolite repression in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2009, 106, 21866–21871. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yeom, J.; Jeon, C.O.; Park, W. Intracellular 2-keto-3-deoxy-6-phosphogluconate is the signal for carbon catabolite repression of phenylacetic acid metabolism in Pseudomonas putida KT2440. Microbiology 2009, 155, 2420–2428. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Phale, P.S. Inducible uptake and metabolism of glucose by the phosphorylative pathway in Pseudomonas putida CSV86. FEMS Microbiol. Lett. 2006, 259, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.R.M.Y.; Leow, C.K.; Neo, J.C.H.; Tan, G.S.; Lo, S.L.; Lim, J.W.E.; Seow, T.K.; Lai, P.B.S.; Chung, M.C.M. Proteome analysis of human hepatocellular carcinoma tissues by two-dimensional difference gel electrophoresis and mass spectrometry. Proteomics 2005, 5, 2258–2271. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Sun, X.; Zhao, Y.; Guo, X.; Jiang, H.; Li, H.; Gu, Z. Evolution of gene regulation during transcription and translation. Genome Biol. Evol. 2015, 7, 1155–1167. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Shao, M.; Chen, L.; Wang, J.; Wu, G.; Tian, X.; Liu, J.; Huang, S.; Zhang, W. Systematic characterization of hypothetical proteins in Synechocystis sp. PCC 6803 reveals proteins functionally relevant to stress responses. Gene 2013, 512, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Wang, J.; Chen, L.; Tian, X.; Huang, S.; Ren, X.; Zhang, W. Quantitative iTRAQ LC-MS/MS proteomics reveals metabolic responses to biofuel ethanol in cyanobacterial Synechocystis sp. PCC 6803. J. Proteome Res. 2012, 11, 5286–5300. [Google Scholar] [CrossRef] [PubMed]
- Man, Z.; Rao, Z.; Xu, M.; Guo, J.; Yang, T.; Zhang, X.; Xu, Z. Improvement of the intracellular environment for enhancing L-arginine production of Corynebacterium glutamicum by inactivation of H2O2-forming flavin reductases and optimization of ATP supply. Metab. Eng. 2016, 38, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, R.; Bao, T.; Rao, Z.; Yang, T.; Xu, M.; Xu, Z.; Li, H.; Yang, S. The rebalanced pathway significantly enhances acetoin production by disruption of acetoin reductase gene and moderate-expression of a new water-forming NADH oxidase in Bacillus subtilis. Metab. Eng. 2014, 23, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Man, Z.; Xu, M.; Rao, Z.; Guo, J.; Yang, T.; Zhang, X.; Xu, Z. Systems pathway engineering of Corynebacterium crenatum for improved L-arginine production. Sci. Rep. 2016, 6, 28629. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Man, Z.; Rao, Z.; Xu, M.; Yang, T.; Zhang, X.; Xu, Z. Improvement of the ammonia assimilation for enhancing L-arginine production of Corynebacterium crenatum. J. Ind. Microbiol. Biotechnol. 2017, 44, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Merfort, M.; Herrmann, U.; Ha, S.W.; Elfari, M.; Bringer-Meyer, S.; Görisch, H.; Sahm, H. Modification of the membrane-bound glucose oxidation system in Gluconobacter oxydans significantly increases gluconate and 5-keto-D-gluconic acid accumulation. Biotechnol. J. 2006, 1, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, R.; Bao, T.; Yang, T.; Xu, M.; Li, H.; Xu, Z.; Rao, Z. Moderate expression of the transcriptional regulator ALsR enhances acetoin production by Bacillus subtilis. J. Ind. Microbiol. Biotechnol. 2013, 40, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Kremmydas, G.F.; Tampakaki, A.P.; Georgakopoulos, D.G. Characterization of the biocontrol activity of Pseudomonas fluorescens strain X reveals novel genes regulated by glucose. PLoS ONE 2013, 8, e61808. [Google Scholar] [CrossRef] [PubMed]
- Gnoni, A.; Lippolis, R.; Zanotti, F.; Papa, S.; Palese, L.L. A two-dimensional electrophoresis and mass spectrometry protein analysis of the antibiotic producer Nonomuraea sp. ATCC 39727 in different growth conditions. FEMS Microbiol. Lett. 2007, 274, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, K.; Xu, S.; Wu, J.; Liu, P.; Du, G.; Li, J.; Chen, J. Identification of membrane proteins associated with phenylpropanoid tolerance and transport in Escherichia coli BL21. J. Proteomics 2015, 113, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Du, B.; Chi, Z.; Ma, L.; Wang, S.; Zhang, X.; Wu, W.; Wang, X.; Xu, G.; Guo, C. Preliminary explorations of the role of mitochondrial proteins in refractory epilepsy: Some findings from comparative proteomics. J. Neurosci. Res. 2007, 85, 3160–3170. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | Relative Transcriptional Levels | |||||
|---|---|---|---|---|---|---|
| gcd | gad | kguK | kguD | glk | gnuK | |
| 4 | 1.0 ± 0.2 | 1.0 ± 0.3 | 1.0 ± 0.2 | 1.0 ± 0.2 | 1.0 ± 0.1 | 1.0 ± 0.2 |
| 8 | 1.1 ± 0.3 | 1.2 ± 0.2 | 1.1 ± 0.1 | 1.6 ± 0.3 | 0.5 ± 0.2 | 0.7 ± 0.2 |
| 12 | 6.2 ± 1.5 | 1.7 ± 0.4 | 2.0 ± 0.6 | 4.5 ± 1.4 | 3.0 ± 0.9 | 0.9 ± 0.4 |
| 16 | 5.0 ± 2.1 | 1.5 ± 0.3 | 14.2 ± 2.5 | 11.8 ± 2.0 | 1.7 ± 0.4 | 1.6 ± 0.5 |
| Glucose (g/L) | 2KGA Production (g/L) | 2KGA Yield on Glucose (g/g) | Productivity (g/L/h) | Biomass (g/L) | Biomass Yield on Glucose (g/g) |
|---|---|---|---|---|---|
| 45 | 35.6 ± 3.0 | 0.79 ± 0.07 | 1.13 ± 0.09 | 8.2 | 0.18 |
| 75 | 61.2 ± 4.8 | 0.82 ± 0.06 | 1.33 ± 0.10 | 8.2 | 0.11 |
| 115 | 100.4 ± 4.3 | 0.87 ± 0.04 | 1.58 ± 0.07 | 8.1 | 0.07 |
| 140 | 127.3 ± 4.6 | 0.91 ± 0.03 | 1.55 ± 0.06 | 8.0 | 0.06 |
| 165 | 162.7 ± 4.8 | 0.99 ± 0.03 | 1.53 ± 0.04 | 7.7 | 0.05 |
| Name | Sequence (5′→3′) |
|---|---|
| rpoDF | GATTCGTCAGGCGATCAC |
| rpoDR | AATACGGTTGAGTTGTTGA |
| gcdF | ACCAGTACCTGCGTGCCTAT |
| gcdR | CCTTGCCGGTGTAGGTCAT |
| gadF | TTTCATGGATTGGGTGGAAC |
| gadR | CGCATCGACTTTCTTCATCA |
| kguKF | GCGACCCGCAAGTGGAATAC |
| kguKR | GAAGGAGATGCTGCGACCGT |
| kguDF | CCGAAACCACTGCCGACACC |
| kguDR | ACGATGCCCAGCGTCTTGC |
| glkF | CTGCATGAGCGGGTATTTC |
| glkR | ATGATCCAACGCCTGCTG |
| gnuKF | GTTCGGACTGGCTACTGATACC |
| gnuKR | ACCGCCAAAGCCGTCCT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, W.; Alexander, T.; Man, Z.; Xiao, F.; Cui, F.; Qi, X. Enhancing 2-Ketogluconate Production of Pseudomonas plecoglossicida JUIM01 by Maintaining the Carbon Catabolite Repression of 2-Ketogluconate Metabolism. Molecules 2018, 23, 2629. https://doi.org/10.3390/molecules23102629
Sun W, Alexander T, Man Z, Xiao F, Cui F, Qi X. Enhancing 2-Ketogluconate Production of Pseudomonas plecoglossicida JUIM01 by Maintaining the Carbon Catabolite Repression of 2-Ketogluconate Metabolism. Molecules. 2018; 23(10):2629. https://doi.org/10.3390/molecules23102629
Chicago/Turabian StyleSun, Wenjing, Tjahjasari Alexander, Zaiwei Man, Fangfang Xiao, Fengjie Cui, and Xianghui Qi. 2018. "Enhancing 2-Ketogluconate Production of Pseudomonas plecoglossicida JUIM01 by Maintaining the Carbon Catabolite Repression of 2-Ketogluconate Metabolism" Molecules 23, no. 10: 2629. https://doi.org/10.3390/molecules23102629