
Targeting N-Glycan Cryptic Sugar Moieties for Broad-Spectrum Virus Neutralization: Progress in Identifying Conserved Molecular Targets in Viruses of Distinct Phylogenetic Origins

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
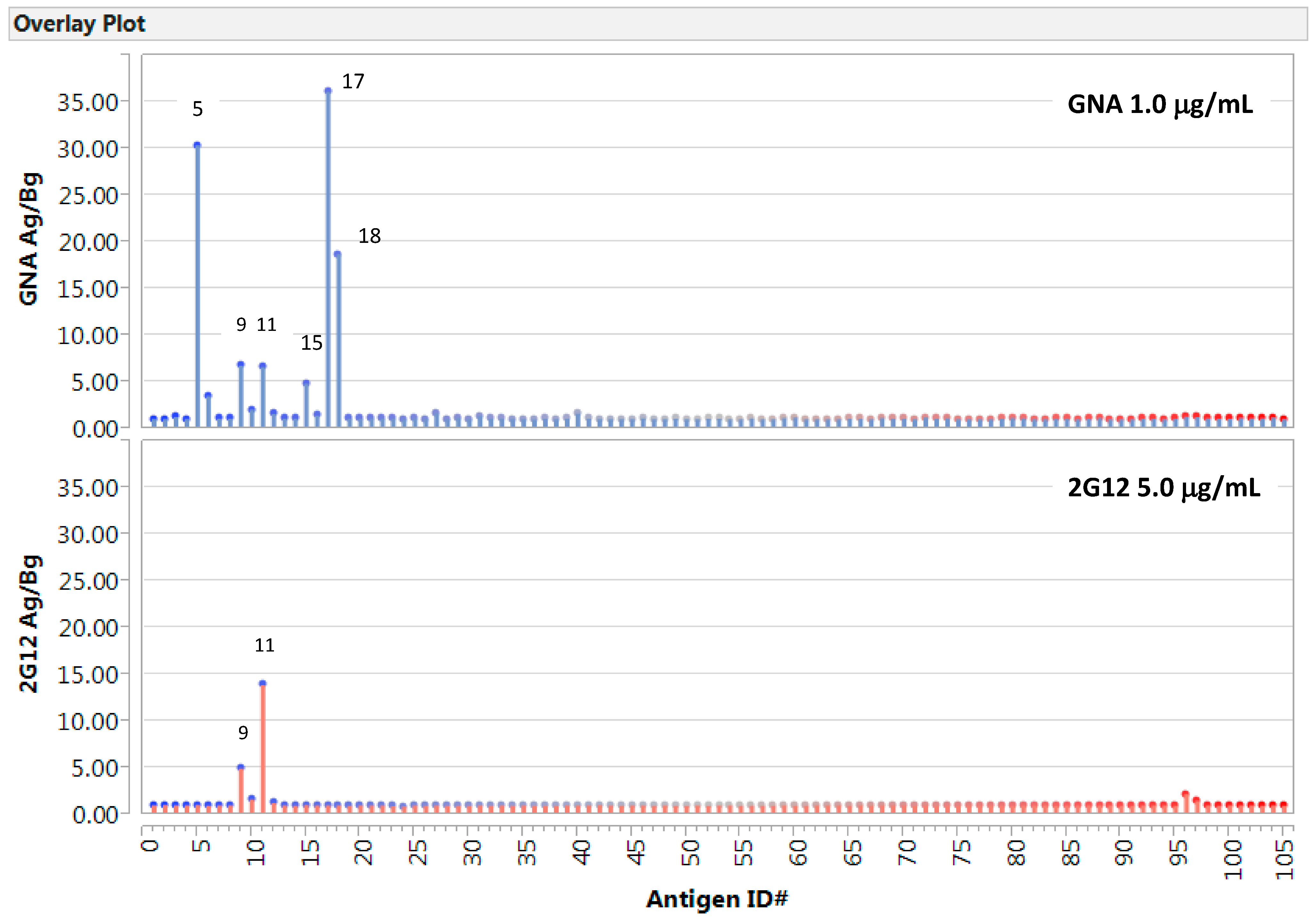
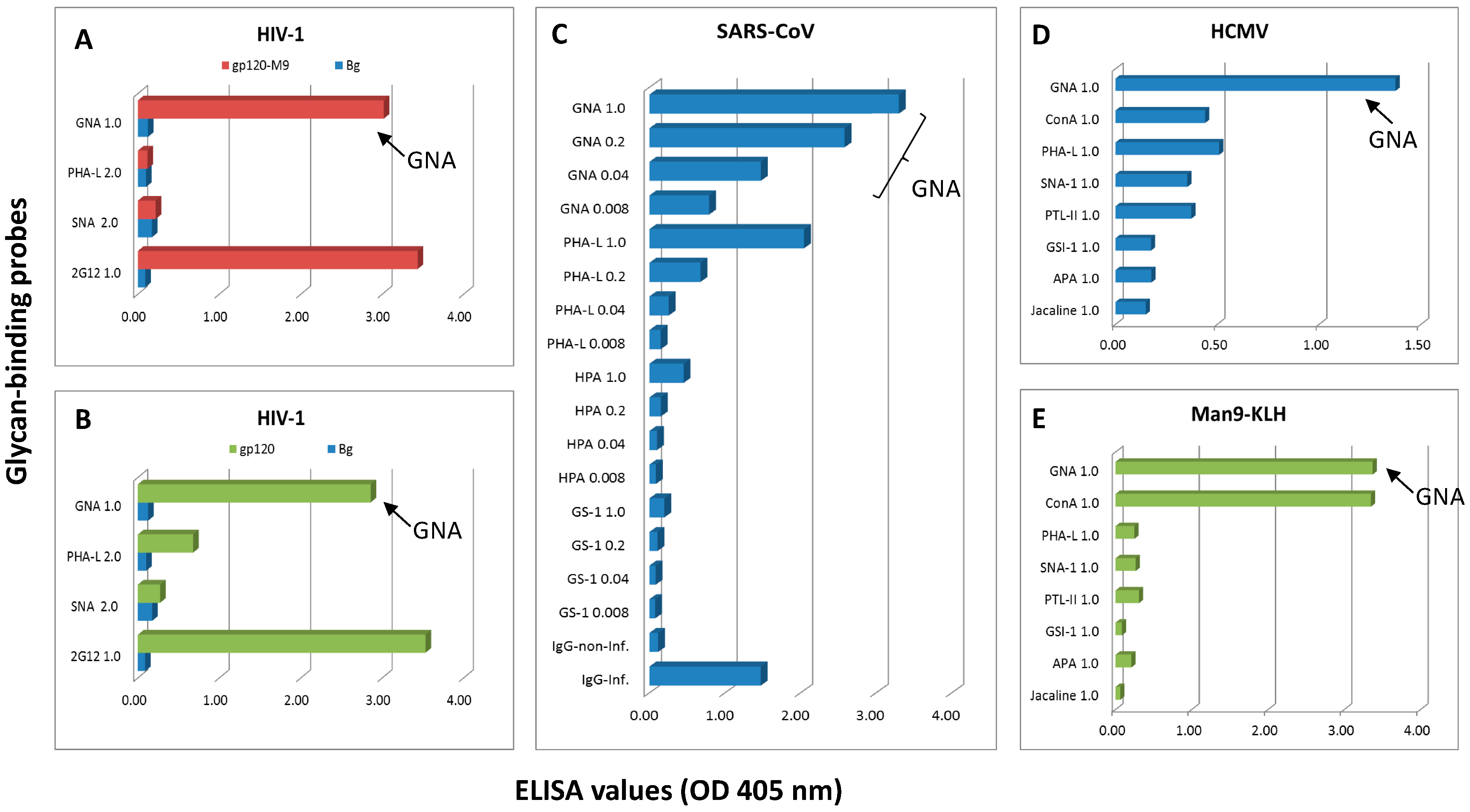
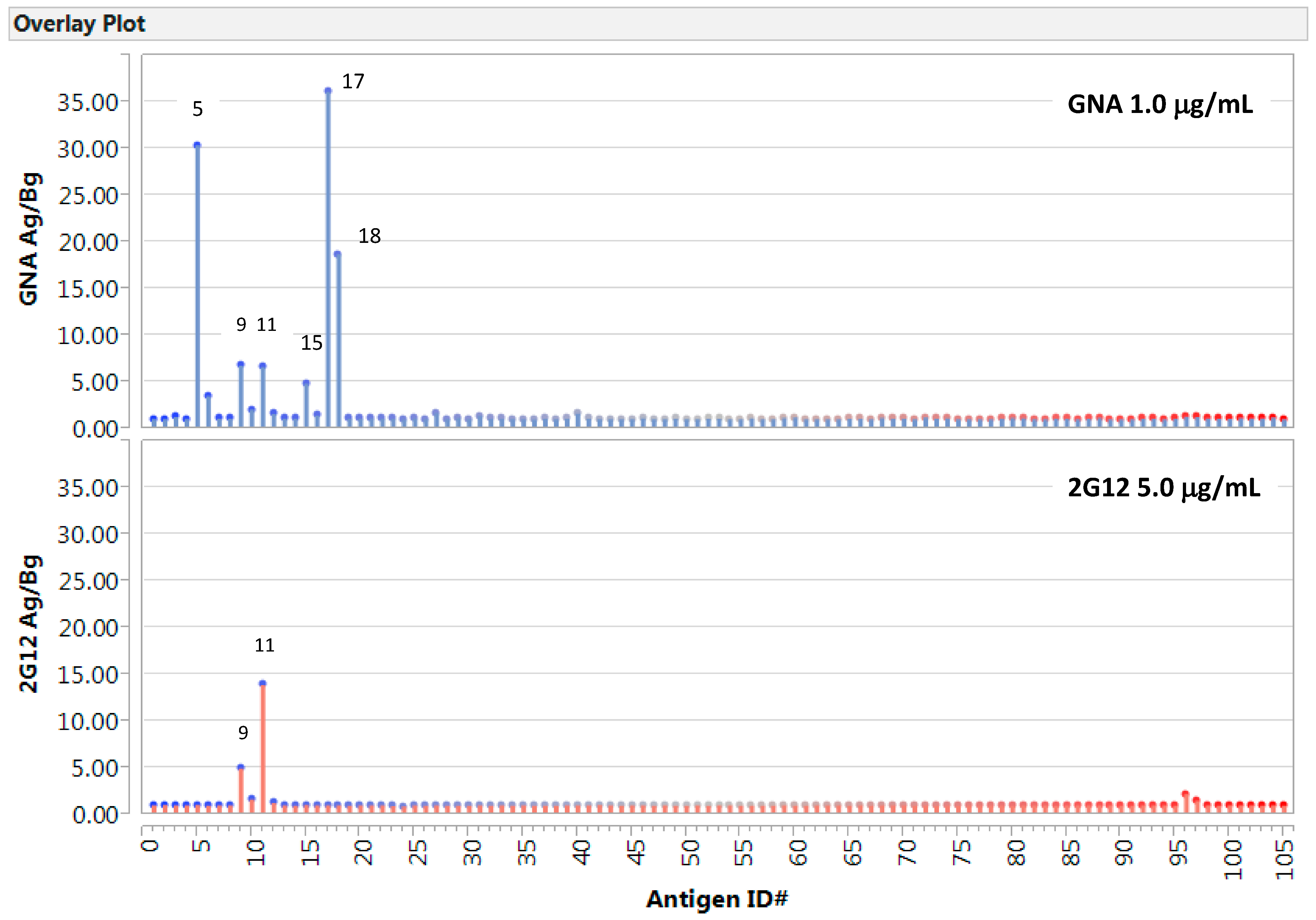
2.1. Detection of GNA- or 2G12-Epitopes in the Native Viral Antigen Preparations
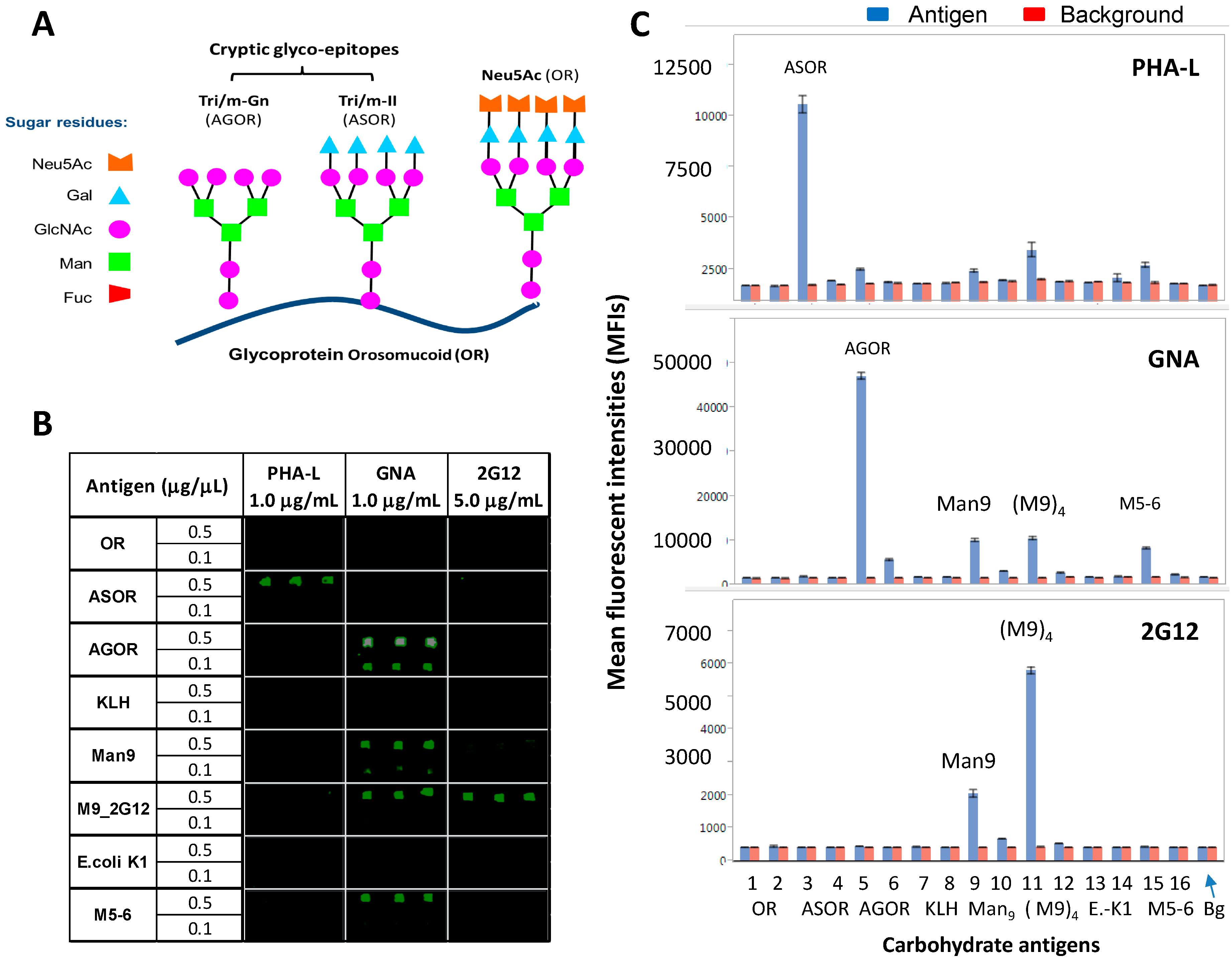
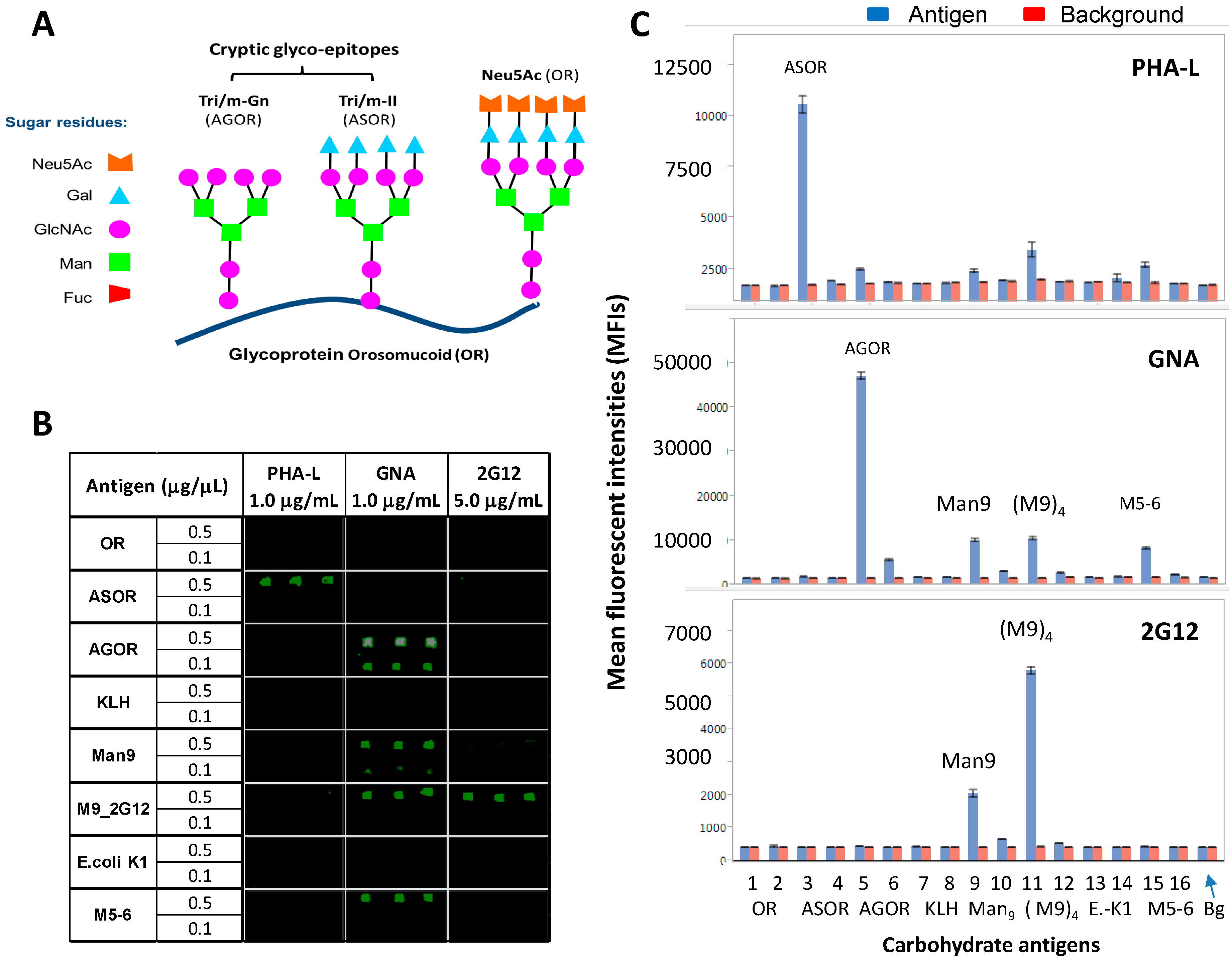
2.2. Carbohydrate Microarrays to Explore the Potential Glyco-Epitopes of GNA
- (1)
- Orosomucoid (OR) (Neu5Ac), ASOR (Tri/m-II), and Agalacto-OR (AGOR) (Tri/m-Gn) (Figure 2A). These autoantigens display distinct glyco-epitopes with identical protein carriers. ASOR is an asialo-derivative of OR, and AGOR is an agalacto-derivative of ASOR. They are crucial for defining binding-specificities for N-glycan cryptic epitopes, Tri/m-II, and Tri/m-Gn.
- (2)
- Thiolated keyhole limpet hemocyanin (KLH-SH), (Man9GlcNAc2Asn)n-KLH (Man9), and [(Man9GlcNAc2Asn)4]n-KLH (M9_2G12). The two Man9-KLH conjugates display Man9 moieties in two defined cluster configurations. M9_2G12 is highly specific for 2G12.
- (3)
- Ribonuclease B (RB) with Man5-6GlcNAc2Asn (M5-6) as the main glycans.
- (4)
- Phosphomannan (P-Man), a yeast polysaccharide. Both M5-6-RB and P-Man are known to be positive with GNA; the former but not the latter also binds to Con A. Using these microarrays, we characterized GNA, PHA-L, and 2G12 for their epitope-binding profiles. Results are summarized in Figure 2 and Figure 3.
3. Experimental Section
3.1. Printing Protein, Carbohydrate and Lipid/Liposome Microarrays
3.2. Staining and Scanning of Microarrays
3.3. Microarray Data-Processing and Statistical Analysis
3.4. Viral Antigen Preparations and Antigen-Specific ELISA
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Abbreviations
| OR | (orosomucoid) |
| ASOR | (asialo-orosomucoid) |
| AGOR | (agalacto-orosomucoid) |
| Tri/m-II | (Tri-antennary and multivalent type II (Galβ1→4GlcNAc) chain epitopes) |
| Tri/m-Gn | (Tri-antennary or multi-valent GlcNAc-terminating epitopes) |
| GNA | (Galanthus nivalis agglutinin) |
| PHA-L | (Phaseolus vulgaris-L lectin) |
| SNA-I | (Sambucus nigra I agglutinin) |
| HCMV | (human cytomegalovirus) |
| HIV-1 | (human immunodeficiency virus-1) |
| SARS-CoV | (severe acute respiratory syndrome coronavirus) |
Conflicts of Interest
References
- Calarese, D.A.; Scanlan, C.N.; Zwick, M.B.; Deechongkit, S.; Mimura, Y.; Kunert, R.; Zhu, P.; Wormald, M.R.; Stanfield, R.L.; Roux, K.H.; et al. Antibody domain exchange is an immunological solution to carbohydrate cluster recognition. Science 2003, 300, 2065–2071. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.M.; Burton, D.R. Rational antibody-based HIV-1 vaccine design: Current approaches and future directions. Curr. Opin. Immunol. 2010, 22, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, C.N.; Pantophlet, R.; Wormald, M.R.; Ollmann Saphire, E.; Stanfield, R.; Wilson, I.A.; Katinger, H.; Dwek, R.A.; Rudd, P.M.; Burton, D.R. The broadly neutralizing anti-human immunodeficiency virus type 1 antibody 2G12 recognizes a cluster of alpha1-->2 mannose residues on the outer face of gp120. J. Virol. 2002, 76, 7306–7321. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.W.; Venturi, M.; Schiffner, L.; Kalyanaraman, R.; Katinger, H.; Lloyd, K.O.; Kwong, P.D.; Moore, J.P. The mannose-dependent epitope for neutralizing antibody 2G12 on human immunodeficiency virus type 1 glycoprotein gp120. J. Virol. 2002, 76, 7293–7305. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.M.; Simek, M.D.; Priddy, F.; Gach, J.S.; Wagner, D.; Zwick, M.B.; Phogat, S.K.; Poignard, P.; Burton, D.R. A limited number of antibody specificities mediate broad and potent serum neutralization in selected HIV-1 infected individuals. PLoS Pathog. 2010, 6, e1001028. [Google Scholar] [CrossRef] [PubMed]
- Pejchal, R.; Walker, L.M.; Stanfield, R.L.; Phogat, S.K.; Koff, W.C.; Poignard, P.; Burton, D.R.; Wilson, I.A. Structure and function of broadly reactive antibody PG16 reveal an H3 subdomain that mediates potent neutralization of HIV-1. Proc. Natl. Acad. Sci. USA 2010, 107, 11483–11488. [Google Scholar] [CrossRef] [PubMed]
- Falkowska, E.; Ramos, A.; Feng, Y.; Zhou, T.; Moquin, S.; Walker, L.M.; Wu, X.; Seaman, M.S.; Wrin, T.; Kwong, P.D.; et al. PGV04, an HIV-1 gp120 CD4 binding site antibody, is broad and potent in neutralization but does not induce conformational changes characteristic of cd4. J. Virol. 2012, 86, 4394–4403. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.M.; Sok, D.; Nishimura, Y.; Donau, O.; Sadjadpour, R.; Gautam, R.; Shingai, M.; Pejchal, R.; Ramos, A.; Simek, M.D.; et al. Rapid development of glycan-specific, broad, and potent anti-HIV-1 gp120 neutralizing antibodies in an R5 SIV/HIV chimeric virus infected macaque. Proc. Natl. Acad. Sci. USA 2011, 108, 20125–20129. [Google Scholar] [CrossRef] [PubMed]
- Pejchal, R.; Doores, K.J.; Walker, L.M.; Khayat, R.; Huang, P.S.; Wang, S.K.; Stanfield, R.L.; Julien, J.P.; Ramos, A.; Crispin, M.; et al. A potent and broad neutralizing antibody recognizes and penetrates the HIV glycan shield. Science 2011, 334, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.X. Synthetic carbohydrate antigens for HIV vaccine design. Curr. Opin. Chem. Biol. 2013, 17, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Trkola, A.; Purtscher, M.; Muster, T.; Ballaun, C.; Buchacher, A.; Sullivan, N.; Srinivasan, K.; Sodroski, J.; Moore, J.P.; Katinger, H. Human monoclonal antibody 2G12 defines a distinctive neutralization epitope on the gp120 glycoprotein of human immunodeficiency virus type 1. J. Virol. 1996, 70, 1100–1108. [Google Scholar] [PubMed]
- McLellan, J.S.; Pancera, M.; Carrico, C.; Gorman, J.; Julien, J.P.; Khayat, R.; Louder, R.; Pejchal, R.; Sastry, M.; Dai, K.; et al. Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature 2011, 480, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lu, J. Glycan arrays lead to the discovery of autoimmunogenic activity of SARS-CoV. Physiol. Genomics 2004, 18, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Eickmann, M.; Becker, S.; Klenk, H.D.; Doerr, H.W.; Stadler, K.; Censini, S.; Guidotti, S.; Masignani, V.; Scarselli, M.; Mora, M.; et al. Phylogeny of the SARS coronavirus. Science 2003, 302, 1504–1505. [Google Scholar] [CrossRef] [PubMed]
- Marra, M.A.; Jones, S.J.; Astell, C.R.; Holt, R.A.; Brooks-Wilson, A.; Butterfield, Y.S.; Khattra, J.; Asano, J.K.; Barber, S.A.; Chan, S.Y.; et al. The genome sequence of the SARS-associated coronavirus. Science 2003, 300, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Rota, P.A.; Oberste, M.S.; Monroe, S.S.; Nix, W.A.; Campagnoli, R.; Icenogle, J.P.; Penaranda, S.; Bankamp, B.; Maher, K.; Chen, M.H.; et al. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science 2003, 300, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Chakraborti, S.; Dimitrov, A.S.; Gramatikoff, K.; Dimitrov, D.S. The SARS-COV S glycoprotein: Expression and functional characterization. Biochem. Biophys. Res. Commun. 2003, 312, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, G.; Harvey, D.J.; Feldmann, F.; Stroeher, U.; Feldmann, H.; Royle, L.; Dwek, R.A.; Rudd, P.M. Identification of N-linked carbohydrates from severe acute respiratory syndrome (SARS) spike glycoprotein. Virology 2010, 399, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Schols, D.; Neyts, J.; Van Damme, E.; Peumans, W.; de Clercq, E. Alpha-(1–3)- and alpha-(1–6)-d-mannose-specific plant lectins are markedly inhibitory to human immunodeficiency virus and cytomegalovirus infections in vitro. Antimicrob. Agents Chemother. 1991, 35, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; van Laethem, K.; Hatse, S.; Froeyen, M.; van Damme, E.; Bolmstedt, A.; Peumans, W.; de Clercq, E.; Schols, D. Marked depletion of glycosylation sites in HIV-1 gp120 under selection pressure by the mannose-specific plant lectins of Hippeastrum hybrid and Galanthus nivalis. Mol. Pharmacol. 2005, 67, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; van Laethem, K.; Hatse, S.; Vermeire, K.; de Clercq, E.; Peumans, W.; van Damme, E.; Vandamme, A.M.; Bolmstedt, A.; Schols, D. Profile of resistance of human immunodeficiency virus to mannose-specific plant lectins. J. Virol. 2004, 78, 10617–10627. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Hatse, S.; Vermeire, K.; Princen, K.; Aquaro, S.; Perno, C.F.; de Clercq, E.; Egberink, H.; Vanden Mooter, G.; Peumans, W.; et al. Mannose-specific plant lectins from the amaryllidaceae family qualify as efficient microbicides for prevention of human immunodeficiency virus infection. Antimicrob. Agents Chemother. 2004, 48, 3858–3870. [Google Scholar] [CrossRef] [PubMed]
- Keyaerts, E.; Vijgen, L.; Pannecouque, C.; van Damme, E.; Peumans, W.; Egberink, H.; Balzarini, J.; van Ranst, M. Plant lectins are potent inhibitors of coronaviruses by interfering with two targets in the viral replication cycle. Antivir. Res. 2007, 75, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, C.; Wei, Y.; Huang, W.; Wang, L.X. Expression, glycoform characterization, and antibody-binding of HIV-1 V3 glycopeptide domain fused with human IgG1-Fc. Bioconjugate Chem. 2010, 21, 875–883. [Google Scholar] [CrossRef]
- Scanlan, C.N.; Ritchie, G.E.; Baruah, K.; Crispin, M.; Harvey, D.J.; Singer, B.B.; Lucka, L.; Wormald, M.R.; Wentworth, P., Jr.; Zitzmann, N.; et al. Inhibition of mammalian glycan biosynthesis produces non-self antigens for a broadly neutralising, HIV-1 specific antibody. J. Mol. Biol. 2007, 372, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Doores, K.J.; Bonomelli, C.; Harvey, D.J.; Vasiljevic, S.; Dwek, R.A.; Burton, D.R.; Crispin, M.; Scanlan, C.N. Envelope glycans of immunodeficiency virions are almost entirely oligomannose antigens. Proc. Natl. Acad. Sci. USA 2010, 107, 13800–13805. [Google Scholar] [CrossRef] [PubMed]
- Rambaruth, N.D.; Greenwell, P.; Dwek, M.V. The lectin helix pomatia agglutinin recognizes O-GlcNAc containing glycoproteins in human breast cancer. Glycobiology 2012, 22, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Tempel, W.; Tschampel, S.; Woods, R.J. The xenograft antigen bound to Griffonia simplicifolia lectin 1-B4. X-ray crystal structure of the complex and molecular dynamics characterization of the binding site. J. Biol. Chem. 2002, 277, 6615–6621. [Google Scholar] [CrossRef] [PubMed]
- Galili, U.; Clark, M.R.; Shohet, S.B.; Buehler, J.; Macher, B.A. Evolutionary relationship between the natural anti-Gal antibody and the Gal alpha 1----3Gal epitope in primates. Proc. Natl. Acad. Sci. USA 1987, 84, 1369–1373. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Bhat, R.; Sobel, R.A.; Huang, W.; Wang, L.X.; Olsson, T.; Steinman, L. Uncovering cryptic glycan markers in multiple sclerosis (MS) and experimental autoimmune encephalomyelitis (EAE). Drug Dev. Res. 2014, 75, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Benko, D.M.; Gibson, W. Primate cytomegalovirus glycoproteins: Lectin-binding properties and sensitivities to glycosidases. J. Virol. 1986, 59, 703–713. [Google Scholar] [PubMed]
- Weekes, M.P.; Tomasec, P.; Huttlin, E.L.; Fielding, C.A.; Nusinow, D.; Stanton, R.J.; Wang, E.C.; Aicheler, R.; Murrell, I.; Wilkinson, G.W.; et al. Quantitative temporal viromics: An approach to investigate host-pathogen interaction. Cell 2014, 157, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Dafik, L.; Nolley, R.; Huang, W.; Wolfinger, R.D.; Wang, L.X.; Peehl, D.M. Anti-oligomannose antibodies as potential serum biomarkers of aggressive prostate cancer. Drug Dev. Res. 2013, 74, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, D. N-glycan cryptic antigens as active immunological targets in prostate cancer patients. J. Proteomics Bioinform. 2012, 5, 090–095. [Google Scholar] [PubMed]
- Wang, D. Carbohydrate antigen microarrays. Methods Mol. Biol. 2012, 808, 241–249. [Google Scholar] [PubMed]
- Wang, D.; Liu, S.; Trummer, B.J.; Deng, C.; Wang, A. Carbohydrate microarrays for the recognition of cross-reactive molecular markers of microbes and host cells. Nat. Biotechnol. 2002, 20, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not applicable.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Tang, J.; Tang, J.; Wang, L.-X. Targeting N-Glycan Cryptic Sugar Moieties for Broad-Spectrum Virus Neutralization: Progress in Identifying Conserved Molecular Targets in Viruses of Distinct Phylogenetic Origins. Molecules 2015, 20, 4610-4622. https://doi.org/10.3390/molecules20034610
Wang D, Tang J, Tang J, Wang L-X. Targeting N-Glycan Cryptic Sugar Moieties for Broad-Spectrum Virus Neutralization: Progress in Identifying Conserved Molecular Targets in Viruses of Distinct Phylogenetic Origins. Molecules. 2015; 20(3):4610-4622. https://doi.org/10.3390/molecules20034610
Chicago/Turabian StyleWang, Denong, Jin Tang, Jiulai Tang, and Lai-Xi Wang. 2015. "Targeting N-Glycan Cryptic Sugar Moieties for Broad-Spectrum Virus Neutralization: Progress in Identifying Conserved Molecular Targets in Viruses of Distinct Phylogenetic Origins" Molecules 20, no. 3: 4610-4622. https://doi.org/10.3390/molecules20034610