2.1. The Lsc3 Mutants Used in This Study
Lsc3 variants addressed in the study include the wild-type Lsc3 and its thirty six mutants. Twenty two of them were previously characterized by us. Among those are the inactive mutants of the catalytic triad Asp62Ala, Asp219Ala and Glu303Ala [
23], mutants with significantly decreased polymerizing ability (Thr302Pro, Gln301Ala and substitution mutants of His321 with Arg, Leu, Lys or Ser) [
21,
23], mutants Trp61Ala, Trp61Asn, Arg304Cys and Arg304Ala which are strongly hampered in sucrose-splitting as well as polymerization abilities [
23] and some others exhibiting moderate changes compared to the wild-type Lsc3. Fourteen mutants of Lsc3 are described here for the first time. Among those are so-called “Yanase mutants”. Inspired by a superior paper by Yanase
et al. [
6] on mutational analysis of the
Zymomonas mobilis levansucrase (LevU), we constructed homologues of Trp80Arg, Glu117Gln, Glu211Gln, Val223Ala and Asp308Asn mutants of
Z. mobilis enzyme. Corresponding mutants of Lsc3 are Trp109Arg, Glu146Gln, Glu236Gln, Val248Ala and Asp333Asn. To gain more information on the functions of these positions, some additional substitutions were made. Therefore, Trp109 in Lsc3 was also replaced with Ala and Phe, and Asp333 with Ala. Several novel mutants of Lsc3 studied here originate from random mutagenesis, the method of which was described by us earlier [
21]. All Lsc3 variants addressed in this work and respective mutagenic oligonucleotides are listed in
Supplementary Table S1. Mutant variants are also designated in
Figure 1 above the alignment of the levansucrases. The mutant
lsc3 genes were cloned into pURI3 vector for overexpression in
E. coli as N-terminally His
6-tagged fusion proteins ([
24,
25]; see
Section 3.1 and
Section 3.2).
Figure 1.
Lsc3 mutants with substituted positions designated as # above the MUSCLE [
26] alignment of levansucrases from
Pseudomonas syringae pv. tomato (Lsc3),
Erwinia amylovora,
Rahnella aquatilis,
Zymomonas mobilis,
Pseudomonas chrororaphis subsp.
aurantiaca, Gluconacetobacter diazotrophicus and
Bacillus subtilis.
Figure 1.
Lsc3 mutants with substituted positions designated as # above the MUSCLE [
26] alignment of levansucrases from
Pseudomonas syringae pv. tomato (Lsc3),
Erwinia amylovora,
Rahnella aquatilis,
Zymomonas mobilis,
Pseudomonas chrororaphis subsp.
aurantiaca, Gluconacetobacter diazotrophicus and
Bacillus subtilis.
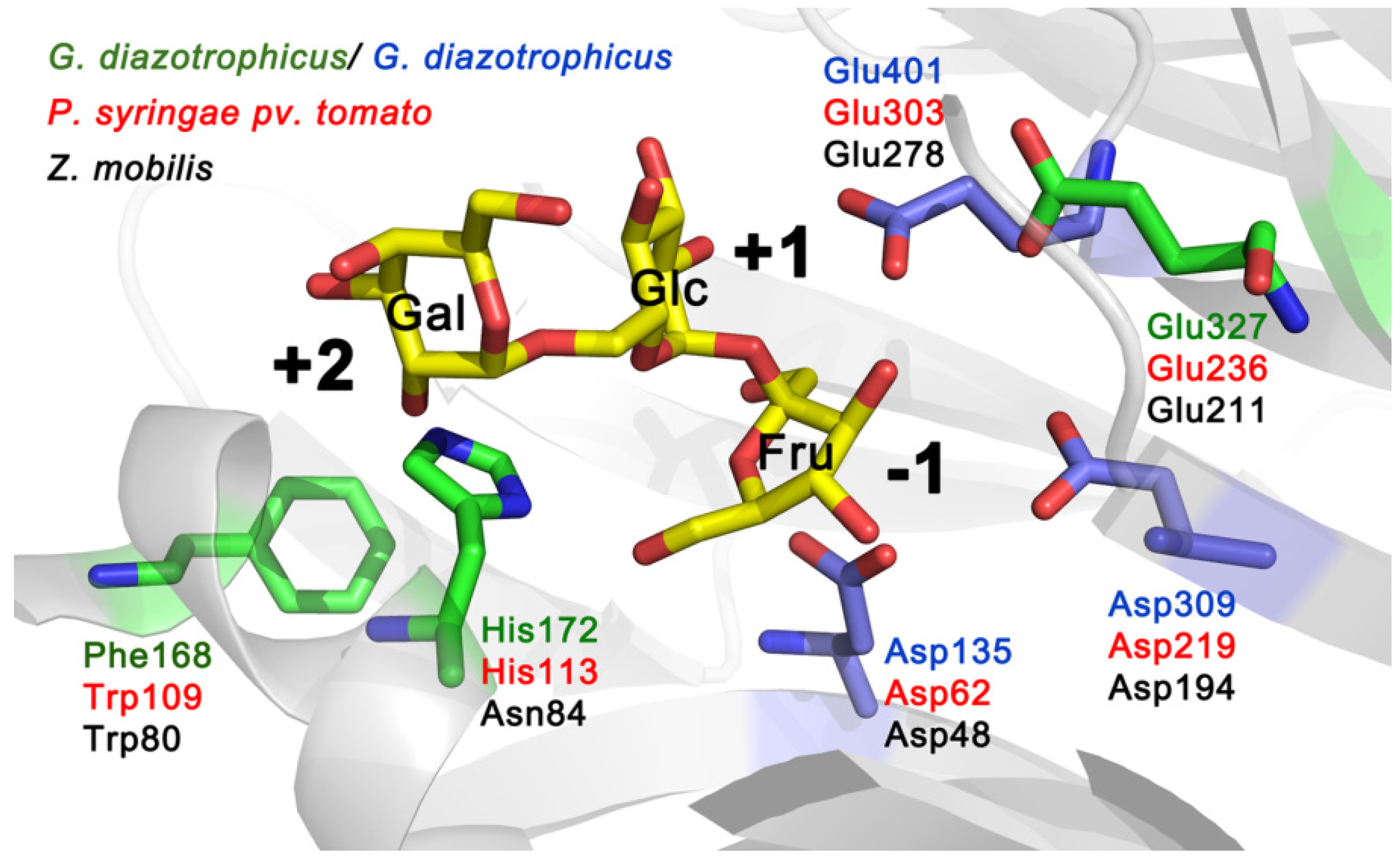
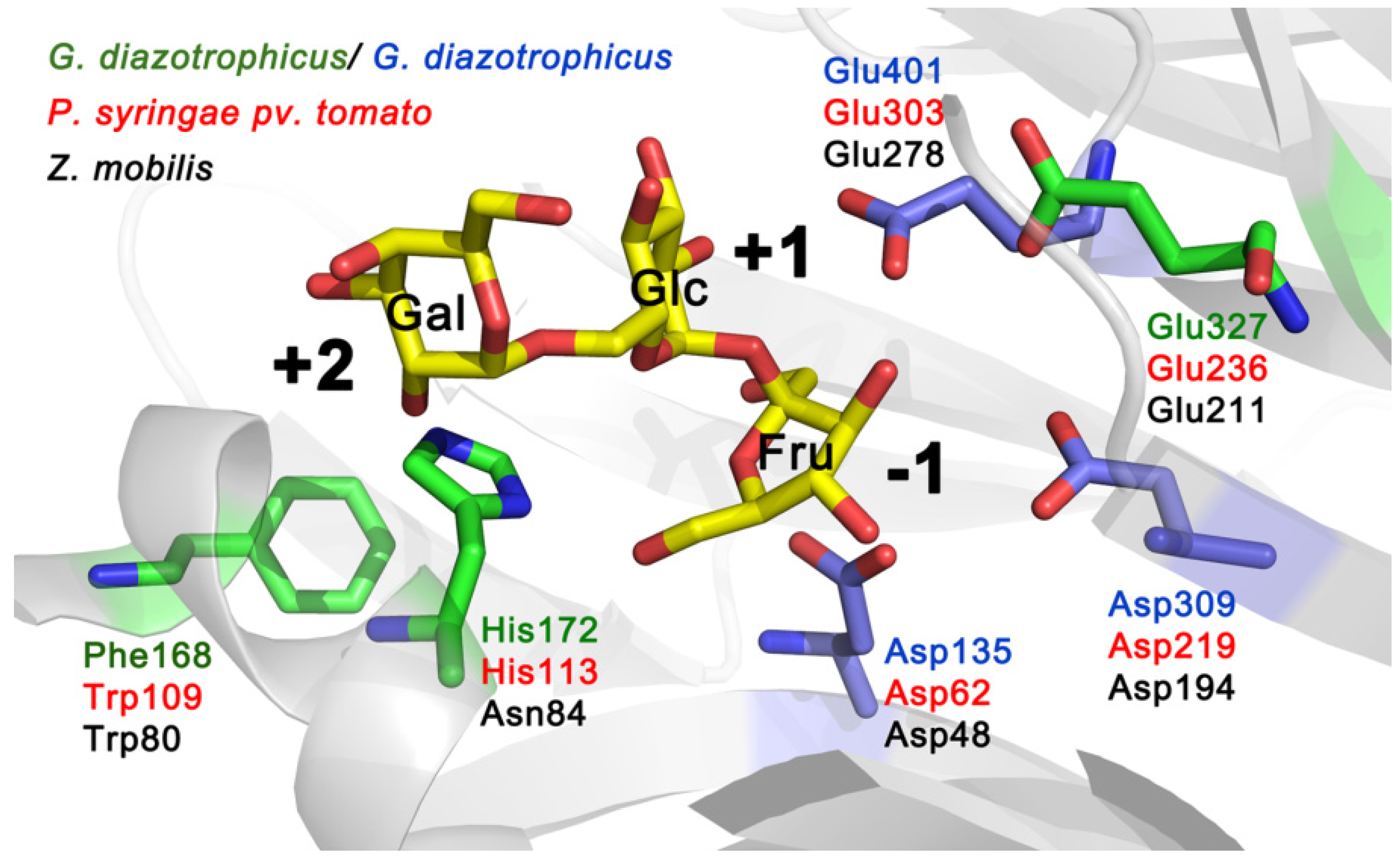
Figure 2 shows the location of amino acids equivalent to Trp109, His113 and Glu236 of Lsc3 on a 3D model of
G. diazotrophicus enzyme LsdA. Trp109 and Glu236 correspond to Trp80 and Glu211 in
Z. mobilis levansucrase (
Figure 1). The Glu211Gln mutant showed vastly reduced polymerizing ability and Trp80Arg mutant was unable to synthesize polymeric levan [
6]. As we show in this study, mutation of His113 in Lsc3 (we studied His113Gln and His113Ala substitutions) has a strong negative effect on the catalysis (
Section 2.3; for further discussion see
Section 2.7). The importance of the above-mentioned positions can be predicted from the location of respective residues in the substrate-binding pocket (
Figure 2).
Figure 2.
Catalytic centre of
G. diazotrophicus levansucrase LsdA (PDB ID: 1W18; [
5]) with side chains of catalytic triad residues (Asp135, Asp309 and Glu401) indicated in blue. The structure of the protein was visualized using PyMol [
27]. Raffinose (consisting of galactose, glucose and fructose residues) shown in substrate-binding pocket originates from the structure of the
B. subtilis levansucrase SacB in complex with raffinose (PDB ID: 3BYN) that was superimposed with the LsdA structure. Positions of LsdA equivalent to Trp109, His113 and Glu236 of Lsc3 of
P. syringae pv. tomato are shown on the model. Respective amino acids of levansucrases of different bacteria are shown in different colour: LevU of
Z. mobilis (
black), Lsc3 of
P. syringae pv. tomato (
red) and LsdA of
G. diazotrophicus (
green and
blue).
Figure 2.
Catalytic centre of
G. diazotrophicus levansucrase LsdA (PDB ID: 1W18; [
5]) with side chains of catalytic triad residues (Asp135, Asp309 and Glu401) indicated in blue. The structure of the protein was visualized using PyMol [
27]. Raffinose (consisting of galactose, glucose and fructose residues) shown in substrate-binding pocket originates from the structure of the
B. subtilis levansucrase SacB in complex with raffinose (PDB ID: 3BYN) that was superimposed with the LsdA structure. Positions of LsdA equivalent to Trp109, His113 and Glu236 of Lsc3 of
P. syringae pv. tomato are shown on the model. Respective amino acids of levansucrases of different bacteria are shown in different colour: LevU of
Z. mobilis (
black), Lsc3 of
P. syringae pv. tomato (
red) and LsdA of
G. diazotrophicus (
green and
blue).
2.3. Total Levansucrase Activity Can Be Reliably Measured on Microplates not only Using Purified Proteins, but also Implementing Permeabilized E. coli Transformants as Catalysts
Most often, enzyme activities are measured by performing reactions in a volume of 1–2 mL. This approach needs a high amount of protein and chemicals and is usually quite time-consuming and costly. Microplate-aided assays save time, money and can be robotized. Total activity of levansucrases is traditionally evaluated according to the velocity of sucrose-splitting reaction. This reaction is required for both sucrose hydrolysis as well as transfructosylation reaction. Each act of sucrose splitting releases a glucose molecule that can be quantified in several ways. We have been using the Glucose liquicolor assay. As described by the manufacturer (Human GmbH, Wiesbaden, Germany), this procedure relies on oxidation of glucose by glucose oxidase, yielding hydrogen peroxide which reacts under catalysis of peroxidase with phenol and 4-aminophenazone to yield a reddish quinoneimine product, concentration of which is measured at 500 nm. We routinely determine kinetic constants of sucrose-splitting reaction of levansucrases by performing reaction in Eppendorf tubes with 1 mL volume of reaction mixture. At fixed time points, small samples are withdrawn for the estimation of liberated glucose. We have earlier used this approach to determine the K
m and V
max values of sucrose-splitting reaction of levansucrases [
4,
21,
23] and applied the same method here (see
Section 3.8). Using a set of 36 Lsc3 mutants with largely varied catalytic activity, we show here that sucrose-splitting activity of the levansucrases can be reliably measured in a high-throughput way,
i.e., on microplates. Moreover, we demonstrate that sucrose splitting by a levansucrase can also be evaluated on the permeabilized
E. coli transformants expressing respective protein.
Figure 4 compares the results obtained by these two methods. One can see that they correlate fairly well. Our levansucrase assay on permeabilized
E. coli transformants presumes similar expression level for different Lsc3 variants in the host. Therefore, all induced
E. coli cultures subjected to levansucrase activity assay were studied for levansucrase expression and similar expression levels for different levansucrase variants were detected. From every induced culture, equal amounts were analysed by SDS-PAGE and the intensity of the bands corresponding to levansucrase protein was compared after staining of the gels with Coomassie Brilliant Blue (data not shown).
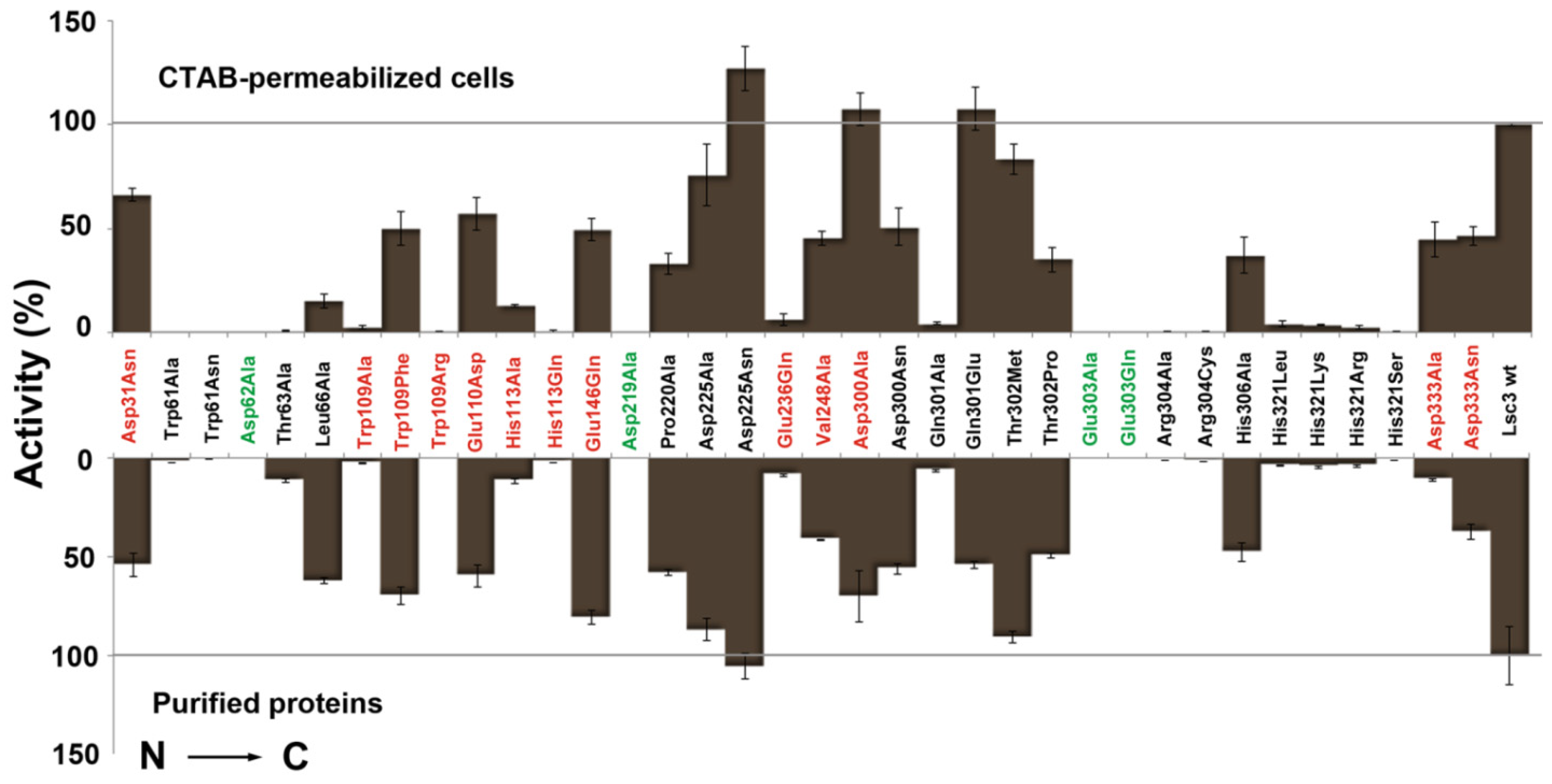
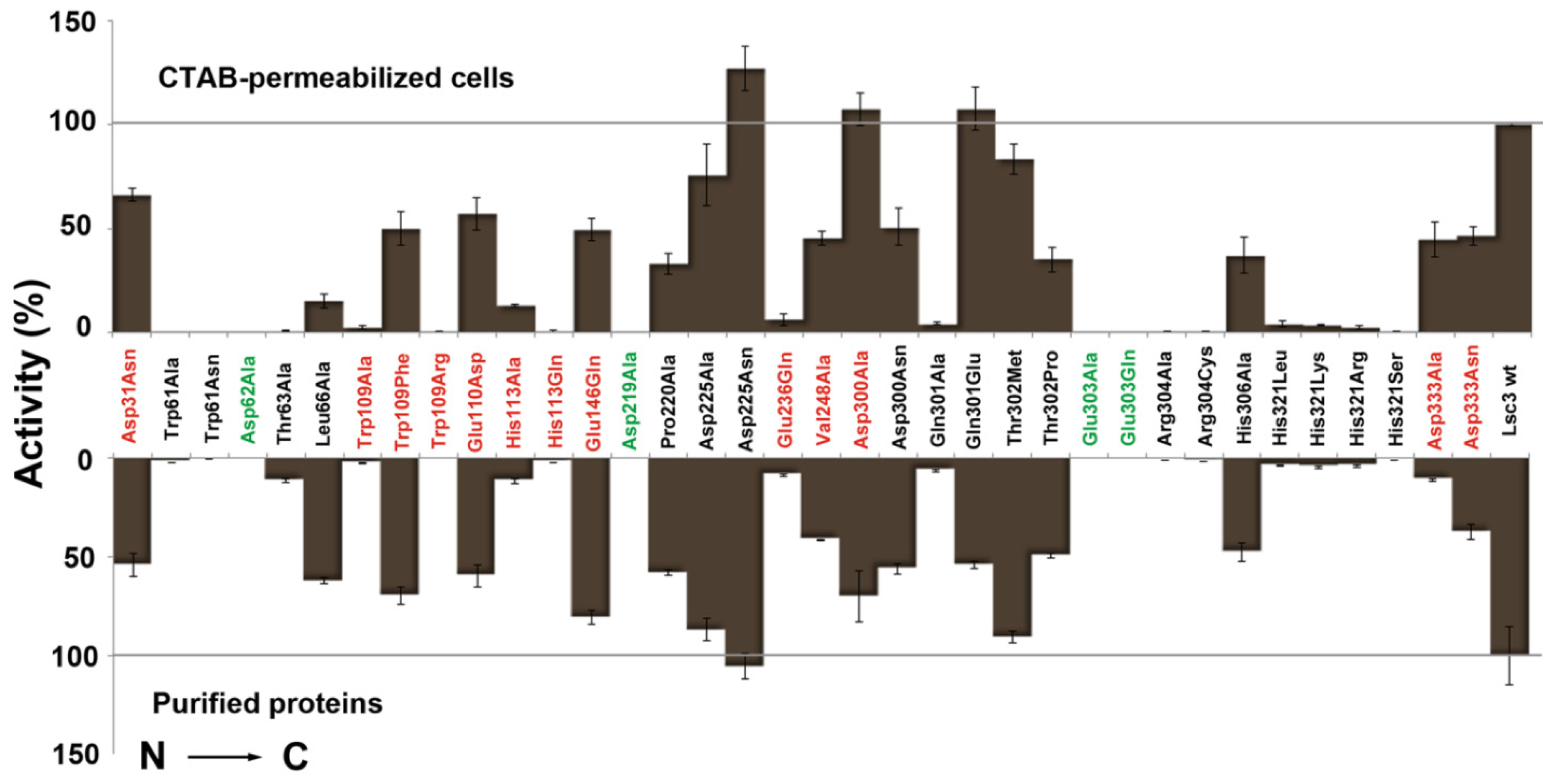
Figure 4.
Quantitative evaluation of the sucrose-splitting (total) levansucrase activity on microplates. Either CTAB-permeabilized
E. coli transformants expressing a certain Lsc3 variant (upper panel) or respective purified proteins (lower panel) were used as catalysts. Respective methods are described in
Section 3.3 and
Section 3.4. Mutants of catalytic triad positions are designated in green and Lsc3 mutants studied for the first time in red. Mutant Glu303Gln falls into both categories. The activities of mutant enzymes are expressed as percentage of respective value of wild-type Lsc3 taken as 100% (grey lines). Total activity of purified wild-type Lsc3 protein was 461.6 ± 95.5 U/mg. In the assay on permeabilized cells, the absorption values at 500 nm indicating glucose concentration were normalized to cell suspension densities and the value obtained for wild-type Lsc3 was taken as 100%. At least three parallel measurements for each levansucrase variant were made and mean values with respective standard deviations are shown.
Figure 4.
Quantitative evaluation of the sucrose-splitting (total) levansucrase activity on microplates. Either CTAB-permeabilized
E. coli transformants expressing a certain Lsc3 variant (upper panel) or respective purified proteins (lower panel) were used as catalysts. Respective methods are described in
Section 3.3 and
Section 3.4. Mutants of catalytic triad positions are designated in green and Lsc3 mutants studied for the first time in red. Mutant Glu303Gln falls into both categories. The activities of mutant enzymes are expressed as percentage of respective value of wild-type Lsc3 taken as 100% (grey lines). Total activity of purified wild-type Lsc3 protein was 461.6 ± 95.5 U/mg. In the assay on permeabilized cells, the absorption values at 500 nm indicating glucose concentration were normalized to cell suspension densities and the value obtained for wild-type Lsc3 was taken as 100%. At least three parallel measurements for each levansucrase variant were made and mean values with respective standard deviations are shown.
![Molecules 19 08434 g004]()
To permeabilize the
E. coli cells, we used 0.1% cetyltrimethylammonium bromide (CTAB) (
Section 3.3) that has only minimal activity-reducing effect on levansucrases. Notably, we have earlier used this surfactant not only for levansucrase activity assay on recombinant
E. coli [
23], but also to evaluate activities of maltase, β-glucuronidase and enzymes of methanol oxidation in yeast cell suspensions [
29,
30]. Earlier, de Abreu
et al. [
31] described a high-throughput microplate assay of
Schwanniomyces occidentalis β-fructofuranosidase mutants. Gene library of respective genes was expressed in an invertase-negative mutant of
Saccharomyces cerevisiae to reveal the protein variants with enhanced transfructosylation ability [
31]. In that case, permeabilization of the yeast cells was not required as invertase is a cell surface-bound protein.
Figure 4 indicates that even a simple levansucrase activity assay on permeabilized
E. coli clearly discloses catalytically disabled mutants Trp61Ala, Trp61Asn, Asp62Ala, Asp219Ala, Glu303Ala, Glu303Gln, Arg304Ala, Arg304Cys, His321Arg, His321Leu, His321Lys and His321Ser [
21,
23]. Concerning novel mutants, Trp109Ala, Trp109Arg, His113Gln and Glu236Gln stick out as the variants with significantly reduced sucrose-splitting ability. For comparison, V
max values of the sucrose-splitting reaction determined with purified proteins are presented in
Figure 5. Some Lsc3 mutants, for example Glu236Gln and substitution mutants of His321, have highly reduced affinity for sucrose (
Table 1). In consequence of that, the sucrose-splitting ability of these mutants is somewhat underestimated in microplate assay which was conducted with 100 mM sucrose (
Figure 4,
Table 1).
Table 1.
Affinity (K
m) for sucrose and polymerizing properties of Lsc3 and its mutants. K
m and TA were determined as described in
Section 3.8 and the reaction products were analysed by HPLC (
Section 3.9). The degree of polymerization (DP) of detected FOS is also presented. Reactions were performed either with purified proteins (see
Section 3.6) or permeabilized cells of levansucrase-expressing
E. coli (see
Section 3.5). In the TA assay, 100% corresponds to the hypothetic situation where no free fructose is produced as a reaction product and all fructose residues of reacted sucrose molecules are incorporated into polymerization products FOS and levan. The mean values and standard deviation values of at least two independent experiments are shown.
Table 1.
Affinity (Km) for sucrose and polymerizing properties of Lsc3 and its mutants. Km and TA were determined as described in Section 3.8 and the reaction products were analysed by HPLC (Section 3.9). The degree of polymerization (DP) of detected FOS is also presented. Reactions were performed either with purified proteins (see Section 3.6) or permeabilized cells of levansucrase-expressing E. coli (see Section 3.5). In the TA assay, 100% corresponds to the hypothetic situation where no free fructose is produced as a reaction product and all fructose residues of reacted sucrose molecules are incorporated into polymerization products FOS and levan. The mean values and standard deviation values of at least two independent experiments are shown.
| Levansucrase | Km (mM) | Transfructosylation Activity (TA; %) Permeabilized Cells/Purified Protein | Degree of Polymerization (DP) Permeabilized Cells/Purified Protein |
|---|
| Lsc3 wt | 18.5 ± 2.5 | 71 ± 1/74 ± 1 a | 3–7/3–7 a |
| Asp31Asn | 14.9 ± 4.6 | 65 ± 4/70 ± 3 | 3–6/3–6 |
| Trp61Ala | 143.4 ± 6.1 | ND/69 ± 1 a | ND/3–6 a |
| Trp61Asn | 869.3 ± 104.0 | ND/51 ± 1 a | ND/3–4 a |
| Asp62Ala | 20.7 ± 2.1 | ND/ND | ND/ND |
| Thr63Ala | 15.9 ± 1.8 | 75 ± 3/71 ± 3 a | 3–6/3–7 a |
| Leu66Ala | 27.3 ± 4.2 | 69 ± 4/73 ± 2 a | 3–6/3–7 a |
| Trp109Ala | 29.8 ± 3.3 | 72 ± 6/77 ± 1 | 3–5/3–7 |
| Trp109Phe | 9.6 ± 0.6 | 62 ± 3/74 ± 1 | 3–7/3–7 |
| Trp109Arg | 249.1 ± 38.5 | ND/40 ± 1 | ND/3 |
| Glu110Asp | 57.6 ± 9.7 | 57 ± 4/70 ± 1 | 3–7/3–7 |
| His113Ala | 170.4 ± 17.0 | 43 ± 1/41 ± 6 | 3–5/3–6 |
| His113Gln | 190.1 ± 28.3 | 65 ± 5/51 ± 3 | 3–5/3–6 |
| Glu146Gln | 40.6 ± 5.0 | 63 ± 5/76 ± 1 | 3–6/3–6 |
| Asp219Ala | 43.4 ± 10.2 | ND/ND | ND/ND |
| Pro220Ala | 23.9 ± 2.3 | 71 ± 2/75 ± 2 a | 3–6/3–6 a |
| Asp225Ala | 13.7 ± 1.2 | 61 ± 1/71 ± 2 a | 3–6/3–7 a |
| Asp225Asn | 18.8 ± 0.8 | 57 ± 2/71 ± 2 a | 3–6/3–7 a |
| Glu236Gln | 267.1 ± 40.2 | 46 ± 1/50 ± 4 | 3–5/3–6 |
| Val248Ala | 14.1 ± 1.1 | 63 ± 2/72 ± 1 | 3–7/3–6 |
| Asp300Ala | 19.4 ± 1.3 | 53 ± 4/58 ± 1 | 3–7/3–8 |
| Asp300Asn | 50.7 ± 5.4 | 52 ± 2/60 ± 1 a | 3–9/3–10 a |
| Gln301Ala | 313.7 ± 30.0 | 24 ± 5/24 ± 1 a | 3–4/3–4 a |
| Gln301Glu | 23.6 ± 1.7 | 34 ± 8/45 ± 1 a | 3–5/3–5 a |
| Thr302Met | 15.1 ± 1.8 | 56 ± 1/70 ± 1 a | 3–6/3–6 a |
| Thr302Pro | 42.5 ± 6.7 | 39 ± 5/52 ± 5 a | 3–6/3–6 a |
| Glu303Ala | 27.1 ± 5.5 | ND/ND | ND/ND |
| Glu303Gln | 129.6 ± 9.6 | ND/ND | ND/ND |
| Arg304Ala | 66.0 ± 9.8 | ND/70 ± 2 a | ND/3–6 a |
| Arg304Cys | 12.5 ± 1.1 | ND/69 ± 1 a | ND/3–4 a |
| His306Ala | 21.2 ± 2.4 | 70 ± 5/72 ± 1 a | 3–7/3–7 a |
| His321Leu | 352.1 ± 41.7 | 17 ± 1/20 ± 5 | 3/3 |
| His321Lys | 529.5 ± 68.3 | 30 ± 1/27 ± 2 | 3–4/3–4 |
| His321Arg | 451.0 ± 36.8 | 36 ± 1/25 ± 1 | 3/3–4 |
| His321Ser | 503.3 ± 87.8 | 31 ± 4/23 ± 3 | 3–4/3–4 |
| Asp333Ala | 27.0 ± 2.9 | 65 ± 3/68 ± 1 | 3–5/3–5 |
| Asp333Asn | 41.3 ± 4.2 | 67 ± 3/80 ± 1 | 3–7/3–7 |
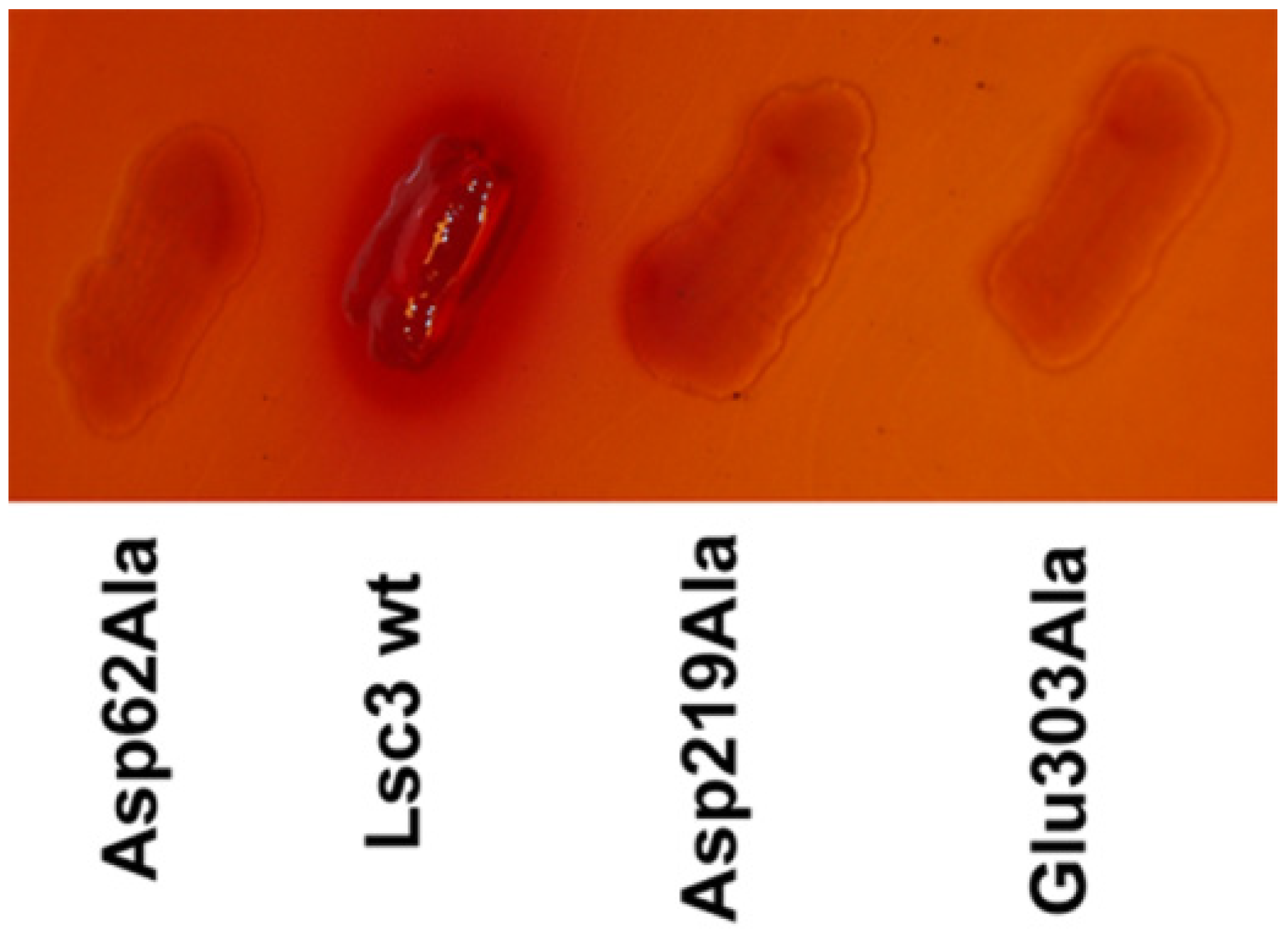
2.4. The Ability of FOS Production by Levansucrases Can be Evaluated on Microplates
Production of β-2,6 linked FOS is important because of their potential biological (prebiotic) effects. They are not produced commercially and are therefore practically impossible to purchase. The β-2,6 linked FOS have been produced for prebiotic efficiency studies in small amounts by controlled chemical hydrolysis of bacterial levan and isolation of the oligosaccharide fractions [
17]. However, some levansucrases, for example LsdA of
G. diazotrophicus, produce mostly short-chain products from sucrose and only a low amount of levan [
32,
33,
34]. Levansucrases with this ability are potential catalysts for large-scale synthesis of β-2,6 linked FOS. We showed that Lsc3 protein of
P. syringae pv. tomato is capable of FOS synthesis from sucrose, raffinose and sugar beet molasses. A high substrate concentration (600 mM and higher) and prolonged reaction time are required for FOS synthesis [
22,
23]. In the search of FOS-producing levansucrases, high-throughput methods for the analysis are preferred. We show here that the ability of levansucrases to produce FOS can be evaluated on microplates. Moreover, even permeabilized
E. coli culture expressing a levansucrase can be reliably applied for FOS production assay (see
Supplementary Figure S1). Indeed, if a levansucrase-expressing
E. coli was permeabilized by 0.1% CTAB and incubated on a microplate in buffer with 1 M of sucrose for 20 h, FOS were produced. The FOS spectrum characteristic for Lsc3 variants of our panel was determined by HPLC (see
Section 3.9) and results are presented in
Table 1. Respective spectra from reactions with purified proteins are given for comparison.
Table 1 also shows transfructosylating activity (TA) of the levansucrases.
The liberation of glucose from a sucrose-splitting reaction shows the total levansucrase activity because glucose molecules are by-products of the reaction and are not polymerized. Fructosyl residues can be transferred to water, resulting in free fructose (hydrolytic activity) or to acceptor molecules other than water (sucrose and fructans), resulting in polymerization products such as FOS and levan (transfructosylating activity). The amount of free glucose and fructose in the reaction mixture is measured and TA is calculated according to the formula: ([Glc] − [Fru
F])/[Glc]) × 100, where [Glc] indicates the content of free glucose and [Fru
F] the content of free fructose in the reaction mixture. Therefore TA indicates the percentage of fructosyl residues that levansucrase protein is using for transfructosylation [
21]. Low TA value means that most of the fructosyl residues from splitted sucrose are transferred to water.
Table 1 shows that the size range of FOS produced can be detected by both high-throughput methods. For example, the Asp300Asn mutant stands out in both assays as the producer of FOS with extended chain length, up to degree of polymerization (DP) 9–10. The mutants synthesizing only short-chain FOS, with DP up to 4 (e.g., Gln301Ala and substitution mutants of His321) were also disclosed in both assays. In several cases, FOS production could not be detected in the assay on permeabilized transformants. In case of mutants with largely decreased catalytic activity, such as Trp61Ala, Trp61Asn, Trp109Arg, Arg304Ala and Arg304Cys (see activity data on
Figure 4 and V
max data in
Figure 5), FOS were detected only in reactions that were performed with an increased amount (100 µg/mL) of purified protein.
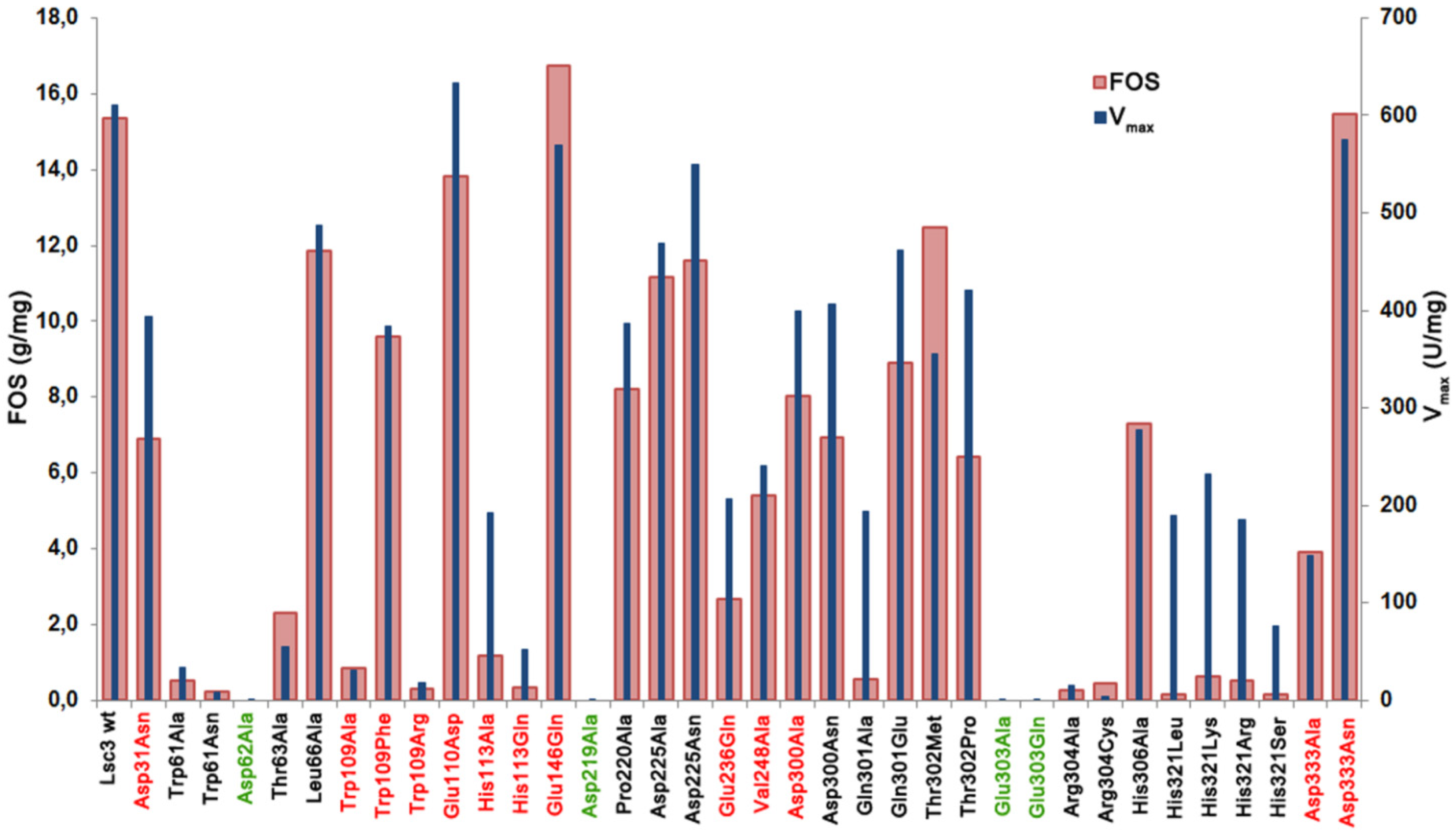
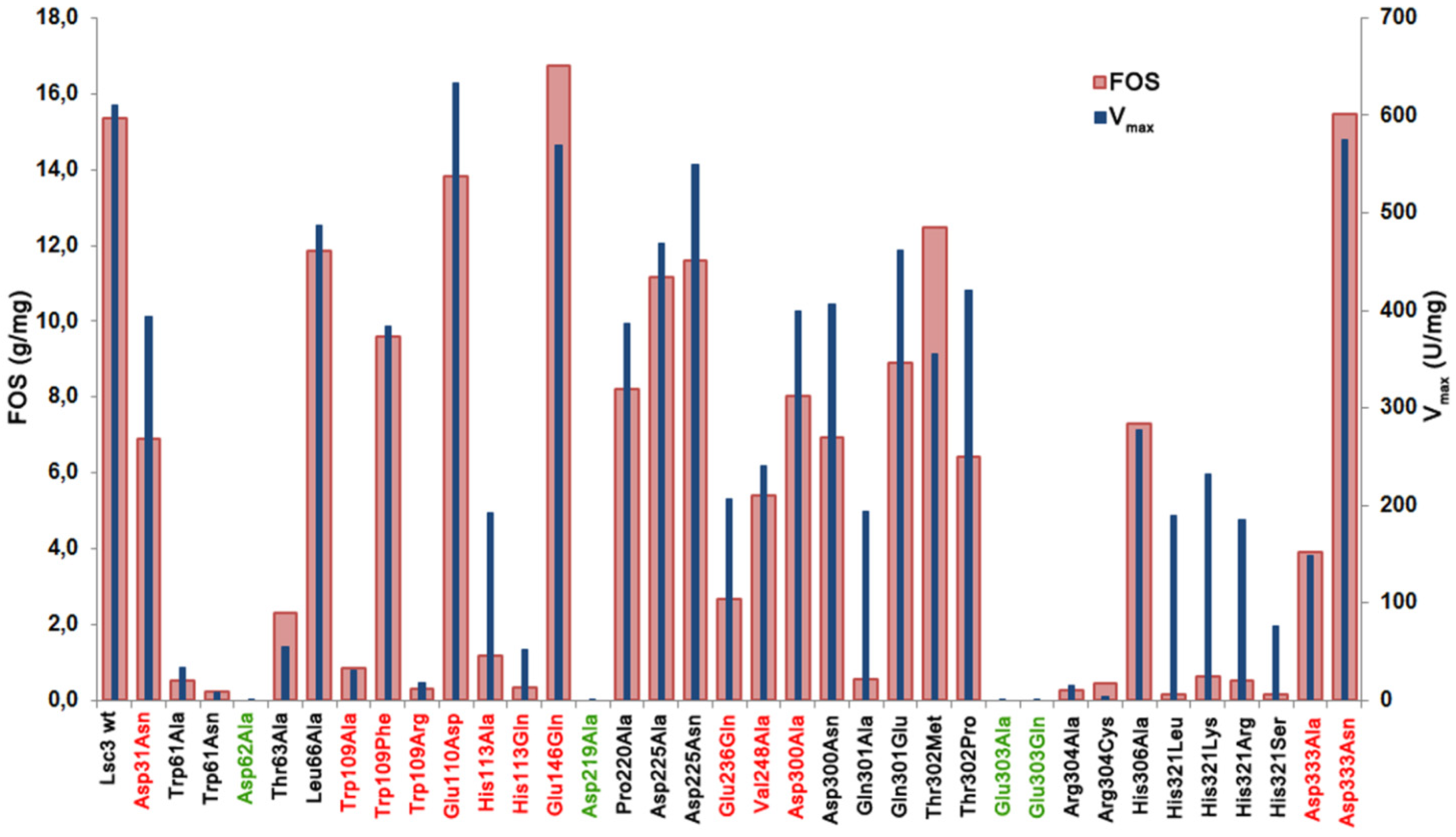
Figure 5 shows data on total FOS production by Lsc3 variants per 1 mg of pure protein.
Figure 5.
The amount of total FOS (g/mg of protein) produced from 1.2 M sucrose in a 20 h reaction by purified wild-type Lsc3 and thirty six mutant proteins. Mutants of catalytic triad positions are shown in green and the mutants studied in this work for the first time are in red. Vmax (U/mg) of sucrose-splitting reaction is plotted for comparison. Mean values of at least two independent experiments are shown. Standard deviation was less than 15%.
Figure 5.
The amount of total FOS (g/mg of protein) produced from 1.2 M sucrose in a 20 h reaction by purified wild-type Lsc3 and thirty six mutant proteins. Mutants of catalytic triad positions are shown in green and the mutants studied in this work for the first time are in red. Vmax (U/mg) of sucrose-splitting reaction is plotted for comparison. Mean values of at least two independent experiments are shown. Standard deviation was less than 15%.
V
max values of sucrose splitting for respective mutants are also included. Comparison of these two characteristics indicates that in case of mutants Asp31Asn, Glu110Asp, His113Ala, Glu236Gln, Asp300Ala, Asp300Asn, Gln301Ala, Thr302Pro and substitution mutants of His321 (His321Arg, His321Leu, His321Lys, His321Ser), FOS production is substantially more affected that the ability to split sucrose, meaning that these positions are specifically important for polymerization reaction. The mutants Glu146Gln, Thr302Met and Asp333Asn with fairly good sucrose-splitting activity and slightly enhanced FOS production (
Figure 5) can be considered as promising candidates for enzymatic synthesis of FOS for biotechnological applications.
2.6. Thermostability Assay of Levansucrases Using Two Different Approaches
Enzymes applied in biotechnology should possess a high catalytic activity and long shelf-life. Bacterial extracellular proteins (including levansucrases) are suited for commercial applications as in nature they have to withstand harsh environmental conditions. We have shown earlier that Lsc3 protein has very high catalytic activity and long-term stability [
21]. Previously we evaluated stability of the Lsc3 preparation during 30 days of storage at 37 °C in McIlvaine’s buffer (pH 6.0) with no loss of catalytic activity [
21]. In our current study we further extended the storage time and report that during 140 days of incubation at 37 °C, Lsc3 retained 50% of initial sucrose-splitting activity. If the same preparation was kept at 50 °C, total catalytic activity dropped more rapidly—after 32 days the residual activity was only 6% of the initial activity. In current experiments, we kept the protein in 100 mM MES buffer (pH 6.5) supplemented with 150 mM NaCl.
Thermofluor is a high-throughput method for protein characterization based on differential scanning fluorimetry [
35]. In case of levansucrases, it has earlier been used for optimization of crystallization conditions of
Erwinia amylovora levansucrase [
8]. After having determined the optimal buffer conditions for wild-type Lsc3 we performed a Thermofluor assay on the panel of thirty seven levansucrase variants. The Thermofluor assay allows the determination of the melting temperature (T
m) of the protein. Increase of temperature promotes unfolding of the protein and T
m is defined as the midpoint of the unfolding transition. A shift in T
m indicates a change in stability of the protein. The T
m for Lsc3 according to Thermofluor assay was 65.4 °C. The corresponding value of
E. amylovora levansucrase was 57 °C and unfolding of this protein started at temperature above 45 °C [
8]. In the assay, buffer conditions (100 mM HEPES buffer; pH 7.5 with 100 mM NaCl) were slightly different than those of our assay. When we conducted the Thermofluor assay of Lsc3 in 100 mM HEPES buffer (pH 7.5), the T
m value of it was only one degree lower than in 100 mM MES buffer (pH 6.5). Based on that we assume Lsc3 is substantially more thermostable than levansucrase of
E. amylovora.
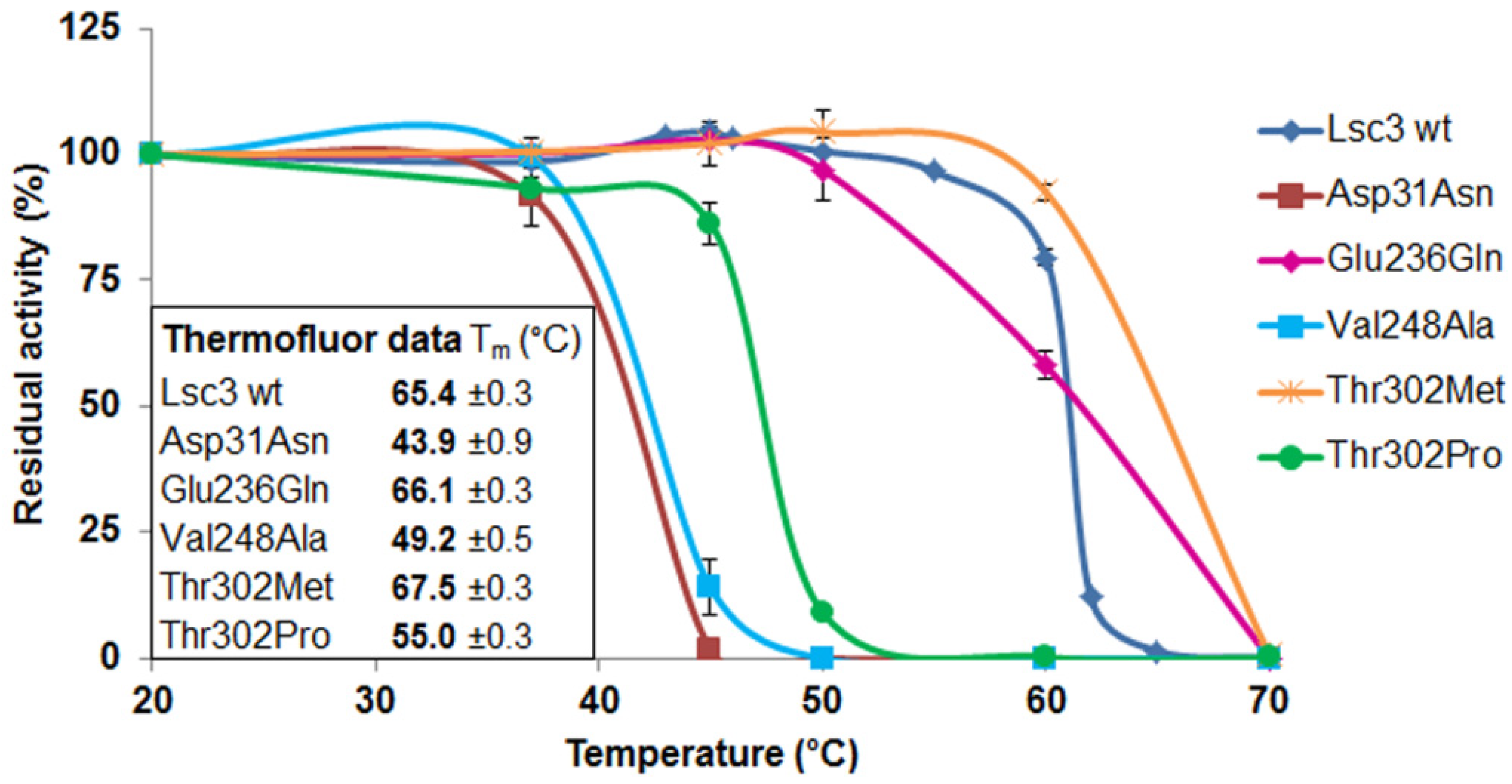
Most Lsc3 mutants had T
m values close to the wild-type protein, ranging from 62 to 66 °C. The mutants Asp31Asn, Val248Ala and Thr302Pro had much lower T
m than the wild-type Lsc3, whereas the mutant Thr302Met had enhanced stability—its T
m was 67.5 °C. We then performed a traditional thermal inactivation assay with wild-type Lsc3 and the mutants Asp31Asn, Glu236Gln, Val248Ala, Thr302Met and Thr302Pro. The proteins were incubated in McIllvaine’s buffer (pH 6.0) for 30 min at a temperature ranging from 20 to 70 °C, cooled on ice and residual total catalytic activity was determined by measuring release of glucose from 100 mM sucrose at 37 °C (see
Section 3.10). As a result, mutants with reduced T
m values according to the Thermofluor assay also showed decreased thermostability in a traditional thermal inactivation study (
Figure 7). We therefore conclude that Thermofluor can be used as a high-throughput tool to evaluate thermostability of levansucrases and their mutant variants.
Figure 7.
Thermal inactivation assay of purified wild-type Lsc3 and the mutant proteins Asp31Asn, Glu236Gln, Val248Ala, Thr302Met and Thr302Pro. After incubation of 5 μM of a levansucrase in McIlvaine’s buffer (pH 6.0) at a selected temperature, its residual activity was measured according to release of glucose from 100 mM sucrose [
4]. The activity of a preparation kept at 20 °C was used as reference for 100% of activity. Inner panel shows T
m values obtained by the Thermofluor assay. The mean values of at least two parallel experiments and respective standard deviations are shown.
Figure 7.
Thermal inactivation assay of purified wild-type Lsc3 and the mutant proteins Asp31Asn, Glu236Gln, Val248Ala, Thr302Met and Thr302Pro. After incubation of 5 μM of a levansucrase in McIlvaine’s buffer (pH 6.0) at a selected temperature, its residual activity was measured according to release of glucose from 100 mM sucrose [
4]. The activity of a preparation kept at 20 °C was used as reference for 100% of activity. Inner panel shows T
m values obtained by the Thermofluor assay. The mean values of at least two parallel experiments and respective standard deviations are shown.
2.7. Structure-Function Analysis of Novel Lsc3 Mutants
In the current work we introduce fourteen novel mutants of the Lsc3 protein of P. syringae pv. tomato DC3000. Phenotype of these mutants will be discussed according to the location of mutated residue, starting from N-terminus of the protein.
The Asp31Asn substitution originates from a random multiple mutant of Lsc3, MutG.
E. coli expressing MutG from a plasmid had nonmucoid colonies on sucrose-containing agar medium. This mutant Lsc3 variant had four substitutions: Asp31Asn, Glu252Gly, Asp300Asn and Cys371Trp [
21]. Based on MutG, two single mutants, Asp300Asn [
21] and Asp31Asn were designed. Position corresponding to Asp31 is not strictly conserved—levansucrases of different bacteria and archaea have also His, Ala, Asn or Glu at this position (see
Figure S2 in Supplementary materials). The Asp31Asn mutant has the lowest thermostability from mutants of the current work (
Figure 7). It suggests that N-terminal domain of Lsc3 plays a role in the stability of the whole protein. The Asp31Asn substitution probably interferes with proper folding of the protein. According to crystal structure data of
G. diazotrophicus and
B. subtilis levansucrases, stable folding of levansucrases is provided through coordinated packing of N- and C-termini of the polypeptide. Similarly to
B. subtilis levansucrase, the N-terminus of the
G. diazotrophicus LsdA protein runs along the perimeter of blade IV forming a clamp-like loop that adds a fifth β-strand to blade III [
5]. Notably, the amino acids of LsdA that comprise additional stabilizing β-strand to blade III are in close proximity of the “RA
DAL” motif [
5] that harbours aspartate (underlined) corresponding to Asp31 of Lsc3.
The Trp109Arg mutant of Lsc3 is a counterpart of Trp109Arg mutant of
Z. mobilis levansucrase, which displayed largely reduced levan synthesis [
6]. We also made additional Trp109 substitutions with Ala and Phe.
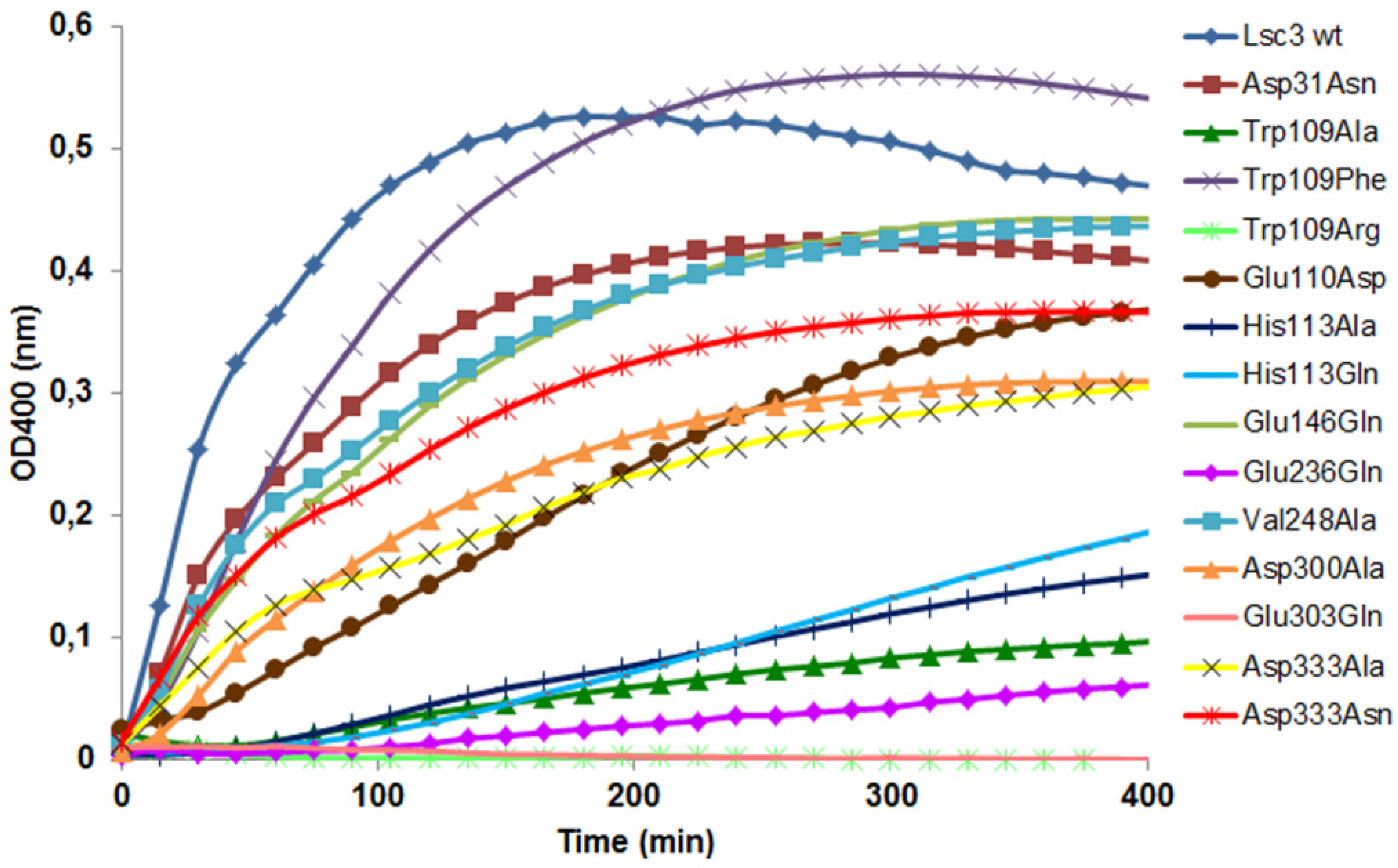
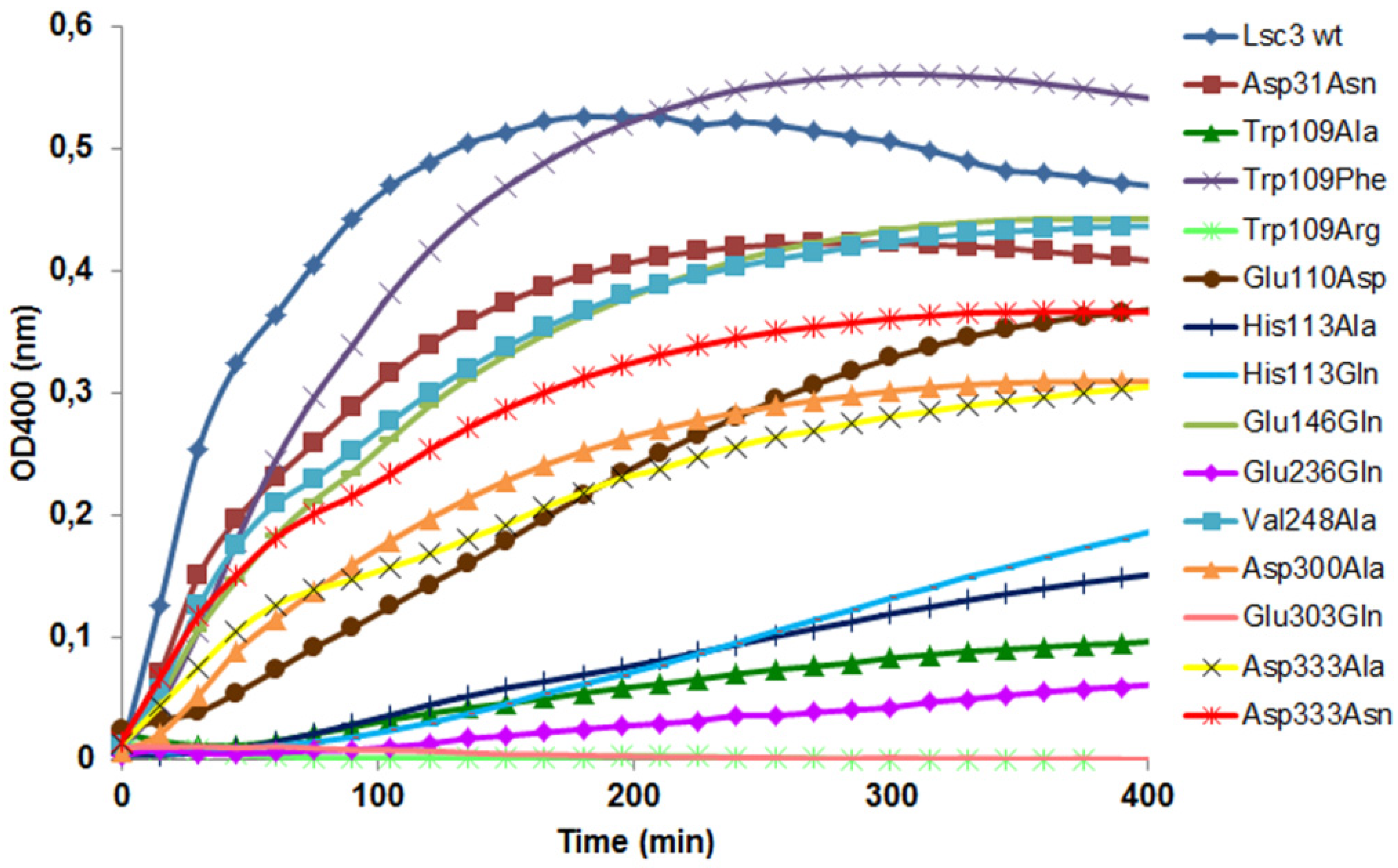
Figure 5 and
Figure 6 show that the Trp109Arg and Trp109Ala mutants display a drastic reduction of levan and FOS synthesis. Some levansucrases, such as the G.
diazotrophicus LsdA (see
Figure 1), have a Phe at the corresponding position. This explains why substitution of Trp109 with Phe restored the wild-type phenotype to Lsc3. Replacement of Trp109 with Phe even slightly enhanced levan synthesis (
Figure 6). We hypothesize that Phe in this position slightly changes the architecture of catalytic centre causing the increase in donor and acceptor binding. Therefore fructan chains are elongated more efficiently. There is no evidence based on crystal structures of levansucrases that Trp or Phe in the respective position directly interact with the substrate [
1,
5]. To make further conclusions, additional evidence from crystal structures and data on product spectra of levansucrases and their mutants are needed. According to Betts and Russell [
36], change of Trp to Phe is favoured in proteins, whereas change to Ala or Arg is not. We assume that Trp109 participates in forming of +2 subsite of the substrate-binding pocket of Lsc3 (see
Figure 2). As sucrose-splitting activities of Trp109Arg and Trp109Ala mutants are also strongly reduced (
Figure 4 and
Figure 5), Trp109 is probably also involved in binding of the substrate at +1 and/or −1 subsites or contributing to substrate cleavage reaction. This assumption is supported by the fact that the Trp109Arg mutant has strongly reduced affinity for sucrose—the K
m is increased from 18.5 to 249 mM (
Table 1).
The Glu110Asp mutant was designed by us to study catalytic significance of a neighbouring position of Trp109. As shown in
Figure 1, this position is not conserved in levansucrases. For example, respective residue in LsdA of
G. diazotrophicus is Asp. Our data on the catalytic properties of Glu110Asp mutant shows no significant changes compared to the wild-type protein. Yet, velocity of levan synthesis was reduced and K
m for sucrose was increased about by 3 times (
Figure 6,
Table 1). Importantly, when Glu110 was changed to Ala, the protein completely precipitated and could not be purified by Ni
2+-affinity chromatography (data not shown) suggesting the importance of the position.
The His113Gln substitution was detected by us in a random double mutant (MutE) of Lsc3. This mutant carried an additional substitution, Val195Ile, and
E. coli expressing this protein had nonmucoid colonies on a sucrose-containing agar plate. We chose to design the His113Gln mutant due to its proximity to a nucleophile Asp62 on the 3D model and a high conservation score.
Figure 2 shows that homologues of His113 residue in levansucrases reside close to fructose residue (binds at −1 subsite) and galactose residue (binds at +2 subsite) of the raffinose molecule bound to the enzyme. Crystal structure of
Arthrobacter sp. K-1 fructosyltransferase in complex with fructose (PDB: 3VSS; [
37]) also shows that His147 in
Arthrobacter sp. K-1 enzyme (a homologue of His113 of Lsc3) is at hydrogen-bondage distance from bound fructose. Notably, the
Arthrobacter sp. K-1 enzyme is structurally similar to levansucrases [
23,
37]. To verify the importance of His113 in the catalysis, we additionally constructed the His113Ala mutant. Like His113Gln substitution, it had a very strong effect on sucrose-splitting reaction and levan synthesis (
Figure 4 and
Figure 6). Notably, the Gln substitution had an even stronger negative effect on sucrose cleavage and FOS synthesis (
Figure 5). We predict that His113 participates in the binding of both fructosyl donor (at −1 subsite) and acceptor (at +2 subsite) to Lsc3. Importantly, this is the first report of catalytic importance of this position in levansucrases.
The Glu146Gln variant of Lsc3 was constructed to mimic the Glu117Gln mutant (see
Figure 1) of
Z. mobilis levansucrase. The Glu117Gln mutant had decreased affinity for sucrose and increased transfructosylating activity [
6]. According to our results, the Glu146Gln mutant of Lsc3 displayed a two-fold increase in K
m towards sucrose, whereas significantly enhanced transfructosylation was not detected (
Table 1;
Figure 6). Still, our assay showed that the Glu146Gln mutant produced more FOS per mg of protein than the wild-type Lsc3 (
Figure 5), whereas levan production was reduced (
Figure 6). Typically, levansucrases have a Glu at position corresponding to Glu146 of Lsc3, but the
Arthrobacter sp. K-1 β-fructofuranosidase which has a high transfrucosylating activity, has a Gln [
37]. We conclude that substitution of Glu146 in Lsc3 with Gln favours synthesis of FOS at the expense of levan.
The Glu236Gln mutant of Lsc3 corresponds to Glu211Gln mutant of
Z. mobilis levansucrase which retained only 28% of hydrolyzing activity and 17% of transfructosylating activity of the wild-type protein [
6]. The Glu236Gln mutant of Lsc3 showed decreased transfructosylation and ability to bind sucrose—the K
m was increased 14.4-fold (see
Table 1 and
Figure 5 and
Figure 6). The position corresponding to Glu236 in Lsc3 is completely conserved among levansucrases (see
Figure 1 and
Figure 2). In-depth analysis of crystal structures of
B. subtilis levansucrase SacB complexed with substrates (PDB IDs: 1PT2 in complex with sucrose and 3BYN with raffinose [
10,
38]) revealed that Glu262 locates close to Arg246 of the RDP motif that participates in binding of the substrate at both the −1 and +1 subsites. According to both crystal structures, Glu262 forms hydrogen bonds with Arg246 and also with fructose and glucose residues of the substrate over a water molecule. We assume that Glu236 contributes to stabilization of the substrate in the active site.
The Val248Ala mutant of Lsc3 is a counterpart of Val223Ala mutant of
Z. mobilis levansucrase that displayed a tenfold decrease of the k
cat whereas the TA value remained the same [
6]. The V
max of sucrose-splitting reaction of Val248Ala mutant of Lsc3 was 39% of the wild-type level when measured at 37 °C (
Figure 5). However, the T
m value of the Val248Ala mutant was 16 degrees lower than that of the wild-type enzyme and its reduced thermostability became also evident in a traditional thermal inactivation assay (
Figure 7). We assume that Val248Ala substitution affects the folding of Lsc3 protein and thereby its stability and catalytic activity.
We have previously described the Asp300Asn mutant originating from random mutagenesis [
21]. This substitution slightly reduces the affinity of the protein for sucrose. An interesting feature of the Asp300Asn mutant is the production of long-chain FOS, with DP up to 10, whereas the wild-type Lsc3 synthesizes FOS with DP up to 7 [
21,
23]. The Asp300Ala mutant addressed in the current work behaved similarly to wild-type Lsc3, producing FOS with DP up to 8 (
Table 1). Thus, for a yet unknown reason, the presence of Asn at position 300 of Lsc3 promotes synthesis of long-chain FOS.
The Asp333Asn mutant of Lsc3 was constructed to mimic the Asp308Asn mutant of
Z. mobilis levansucrase. The Asp308Asn mutant had slightly decreased transfructosylation ability and about four-fold decreased affinity for sucrose [
6]. This position is highly conserved in levansucrases (
Figure 1). Yet, in
Arthrobacter sp. K1 enzyme, a Glu, and in
B. subtilis SacB an Asn (Asn372) is found at the same position (
Figure 1). Aside of making the Asp333Asn mutant, we also constructed respective Ala substitution variant. V
max of sucrose-splitting reaction of the Asp333Ala mutant was four-fold lower compared to wild-type enzyme and levan synthesis was also significantly reduced. The Asp333Asn mutant behaved much more similarly to wild-type Lsc3 than the Asp333Ala (
Figure 5 and
Figure 6). We suppose that Asp333 can be involved in substrate binding and elongation of the fructan chain.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}