Natural Dibenzo-α-Pyrones and Their Bioactivities



Abstract
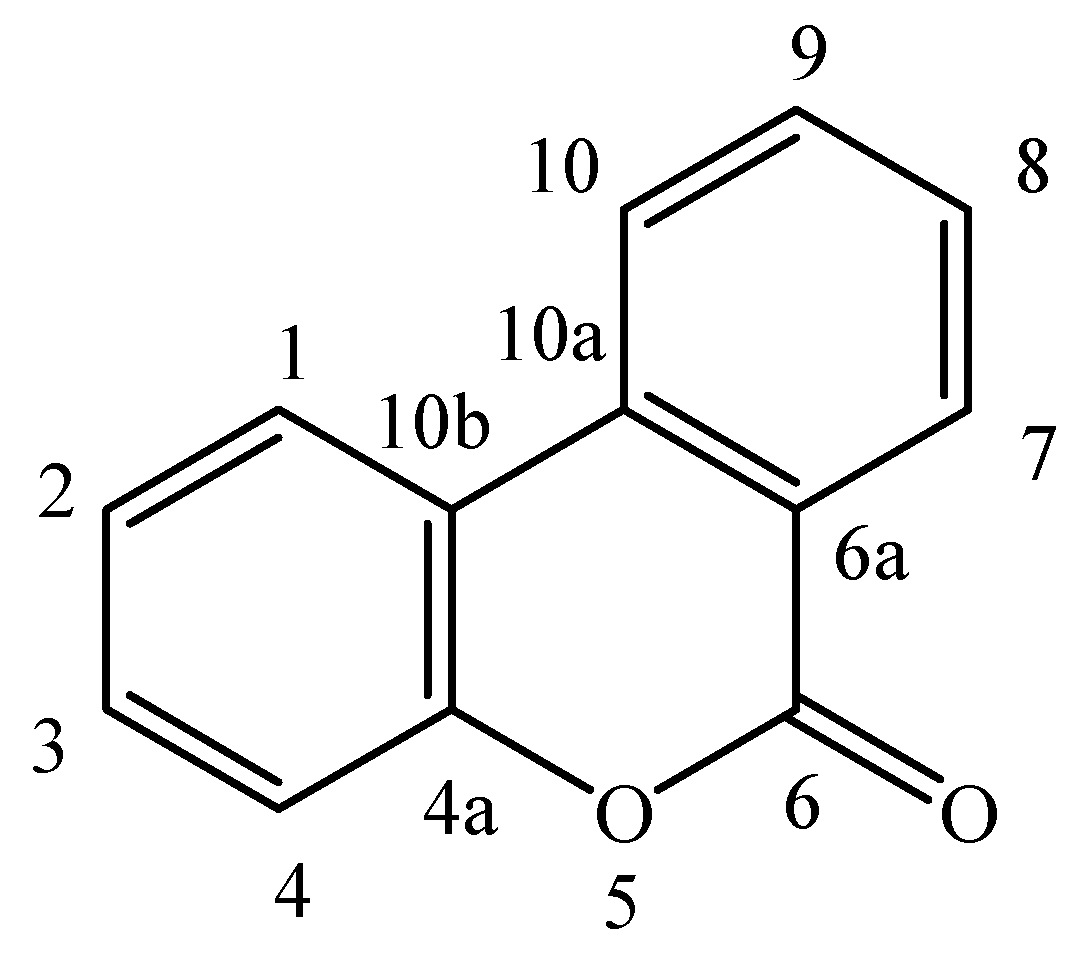
:1. Introduction
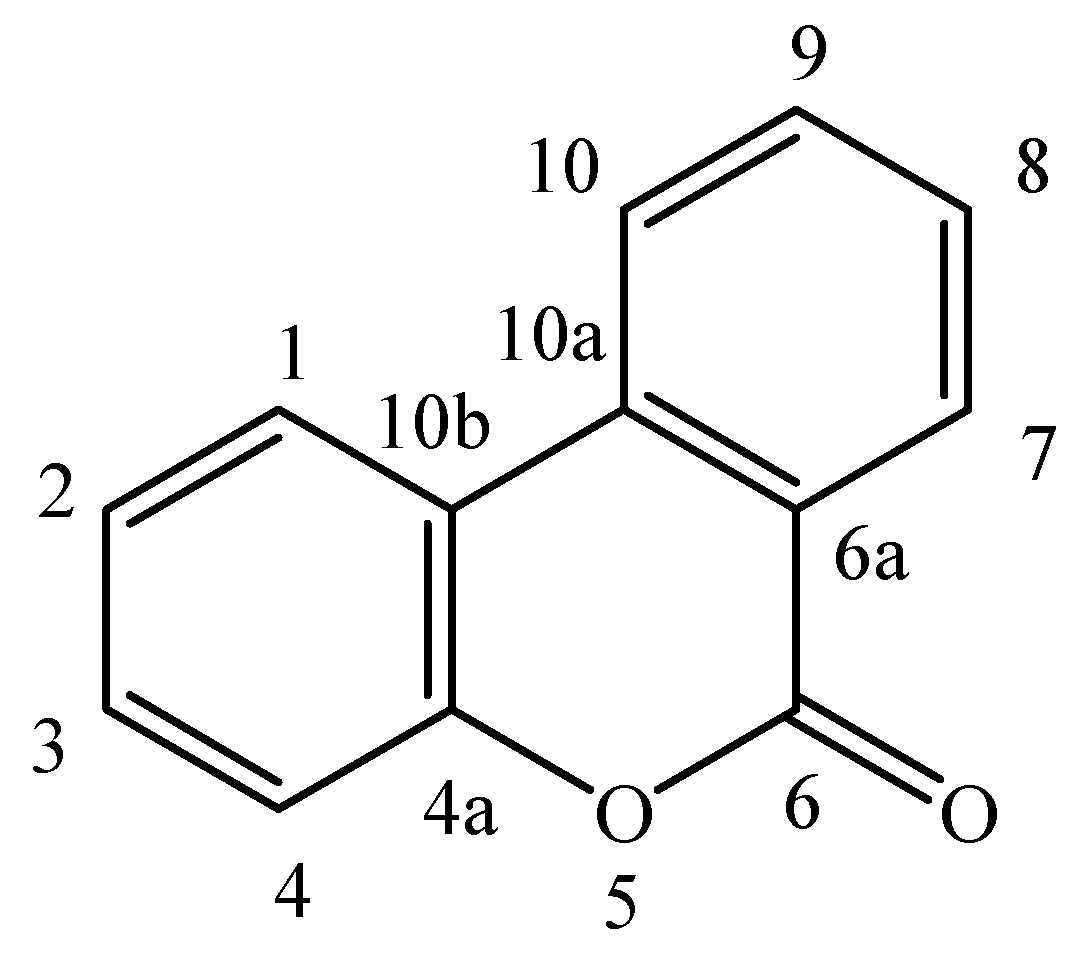
2. Occurrence
2.1. Dibenzo-α-pyrones from Fungi

| Dibenzo-α-pyrone | Fungal Species | Reference |
|---|---|---|
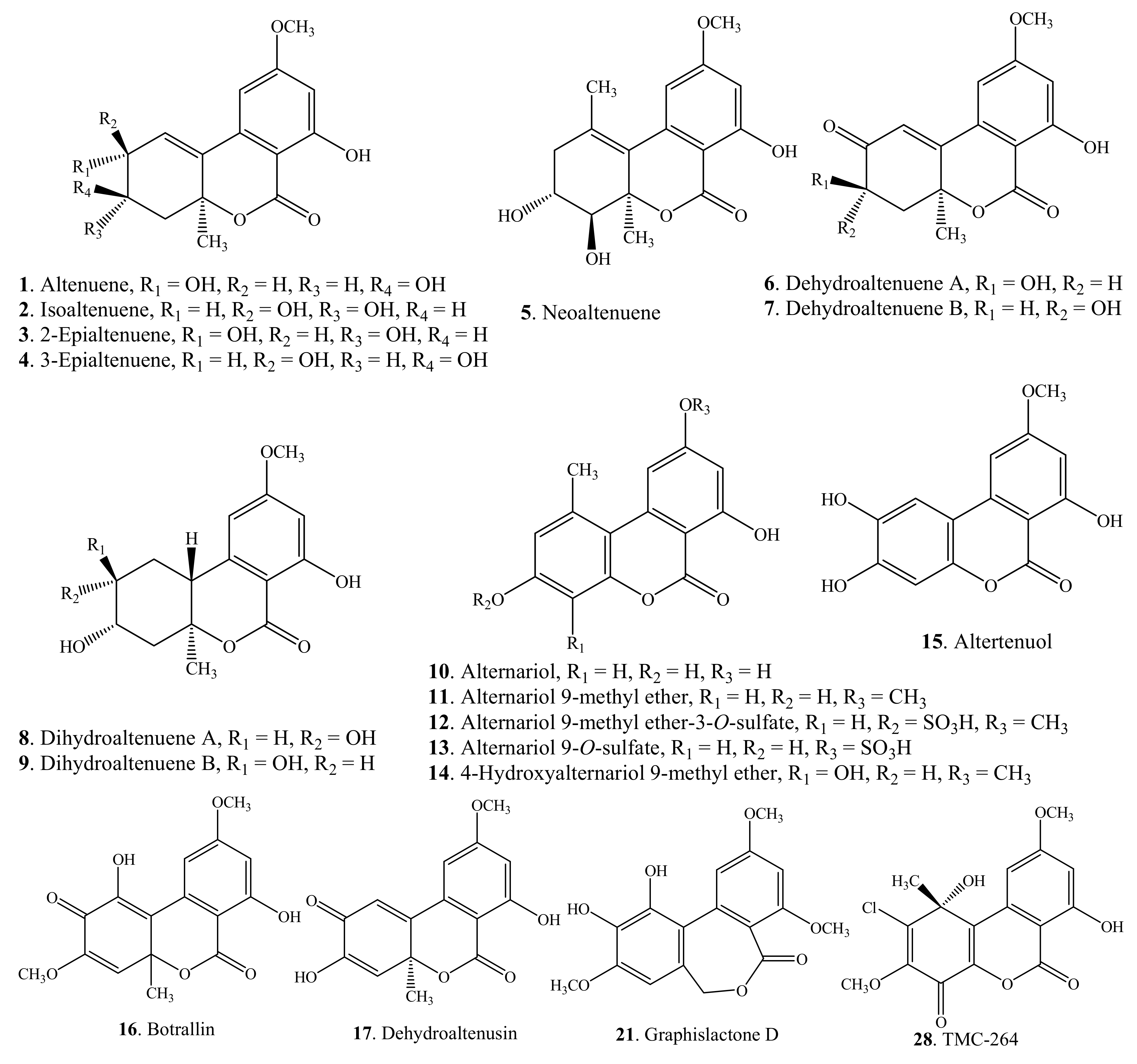
| Altenuene = ATL (1) | Endophytic Alternaria sp. isolated from Polygonum senegalense | [9] |
| Alternaria alternata | [10] | |
| Unidentified freshwater fungus belong to Tubeufiaceae | [19] | |
| Isoaltenuene (2) | Alternaria alternata | [20] |
| Unidentified freshwater fungus belong to Tubeufiaceae | [19] | |
| 2-Epialtenuene (3) | Alternaria alternata | [21] |
| Unidentified freshwater fungus belong to Tubeufiaceae | [19] | |
| 3-Epialtenuene (4) | Endophytic Alternaria sp. isolated from Polygonum senegalense | [9] |
| Neoaltenuene (5) | Alternaria alternata | [21] |
| Dehydroaltenuene A (6) | Unidentified freshwater fungus belong to Tubeufiaceae | [19] |
| Dehydroaltenuene B (7) | Unidentified freshwater fungus belong to Tubeufiaceae | [19] |
| Dihydroaltenuene A (8) | Unidentified freshwater fungus belong to Tubeufiaceae | [19] |
| Dihydroaltenuene B (9) | Unidentified freshwater fungus belong to Tubeufiaceae | [19] |
| Alternariol = AOH (10) | Endophytic Acremonium sp. isolated from Plantago lanceolata | [22] |
| Endophytic Alternaria sp. isolated from Polygonum senegalense | [9] | |
| Endophytic Alternaria no.28 isolated from Ginkgo biloba | [23] | |
| Endophytic Alternaria sp. PR-14 isolated from Paeonia delavayi | [24] | |
| Endophytic Alternaria sp. isolated from Datura stramonium | [25] | |
| Endophytic Alternaria sp. N.SBA10 isolated from Scutellaria baicalensis | [26] | |
| Alternaria alternata | [27] | |
| Alternaria alternata | [10] | |
| Endophytic Alternaria brassicicola ML-P08 isolated from Malus halliana | [28] | |
| Endophytic Alternaria tenuissima EN-192 isolated from Rhizophora stylosa | [29] | |
| Endophytic Colletotrichum sp. isolated from Aristolochia sp. | [30] | |
| Alternariol 9-methyl ether = AME = Djalonensone (11) | Endophytic Acremoniumsp. isolated from Plantago lanceolata | [22] |
| Endophytic Alternaria sp. isolated from Polygonum senegalense | [9] | |
| Endophytic Alternaria sp. PR-14 isolated from Paeonia delavayi | [24] | |
| Endophytic Alternaria sp. isolated from Datura stramonium | [25] | |
| Endophytic Alternaria sp. N.SBA10 isolated from Scutellaria baicalensis | [26] | |
| Alternaria alternata | [27] | |
| Alternaria alternata | [31] | |
| Alternaria alternata | [10] | |
| Endophytic Alternaria no.28 isolated from Ginkgo biloba | [23] | |
| Endophytic Alternaria brassicicola ML-P08 isolated from Malus halliana | [28] | |
| Endophytic Alternaria linicola isolated from Linum ustiatissimum | [32] | |
| Alternaria tenuis | [33] | |
| Alternariol 9-methyl ether = AME = Djalonensone (11) | Endophytic Alternaria tenuissima isolated from Acacia mangium | [34] |
| Endophytic Alternaria tenuissima EN-192 isolated from Rhizophora stylosa | [29] | |
| Endophytic Cephalosporium acremonium IFB-E007 isolated from Trachelospermum jasminoides | [35] | |
| Endophytic Colletotrhichum sp. isolated from Aristolochia sp. | [30] | |
| Endophytic Hyalodendriella sp. Ponipodef12 isolated from the hybrid ‘Neva’ of Populus deltoides × P. nigra | [12] | |
| Lachmum palmae | [36] | |
| Alternariol 9-methyl ether-3-O-sulfate (12) | Endophytic Alternaria sp. isolated from Polygonum senegalense | [9] |
| Alternariol 9-O-sulfate (13) | Endophytic Alternaria sp. isolated from Polygonum senegalense | [9] |
| 4-Hydroxyalternariol 9-methyl ether (14) | Endophytic Alternaria sp. isolated from Polygonum senegalense | [9] |
| Endophytic Alternaria sp. isolated from Datura stramonium | [25] | |
| Altertenuol = Altenuisol = Alternuisol (15) | Alternaria sp. | [37] |
| Alternaris tenuis | [38] | |
| Alternaris tenuis | [39,40] | |
| Alternaris tenuis | [41] | |
| Botrallin (16) | Botrytis allii | [42] |
| Endophytic Hyalodendriella sp. Ponipodef12 isolated from the hybrid ‘Neva’ of Populus deltoides × P. nigra | [12,43] | |
| Endophytic Microsphaeropsis olivacea isolated from Pilgerodendron uviferum | [13] | |
| Dehydroaltenusin (17) | Acremonium sp. | [44] |
| Alternaria tenuis | [45] | |
| Penicillium verruculosum | [46] | |
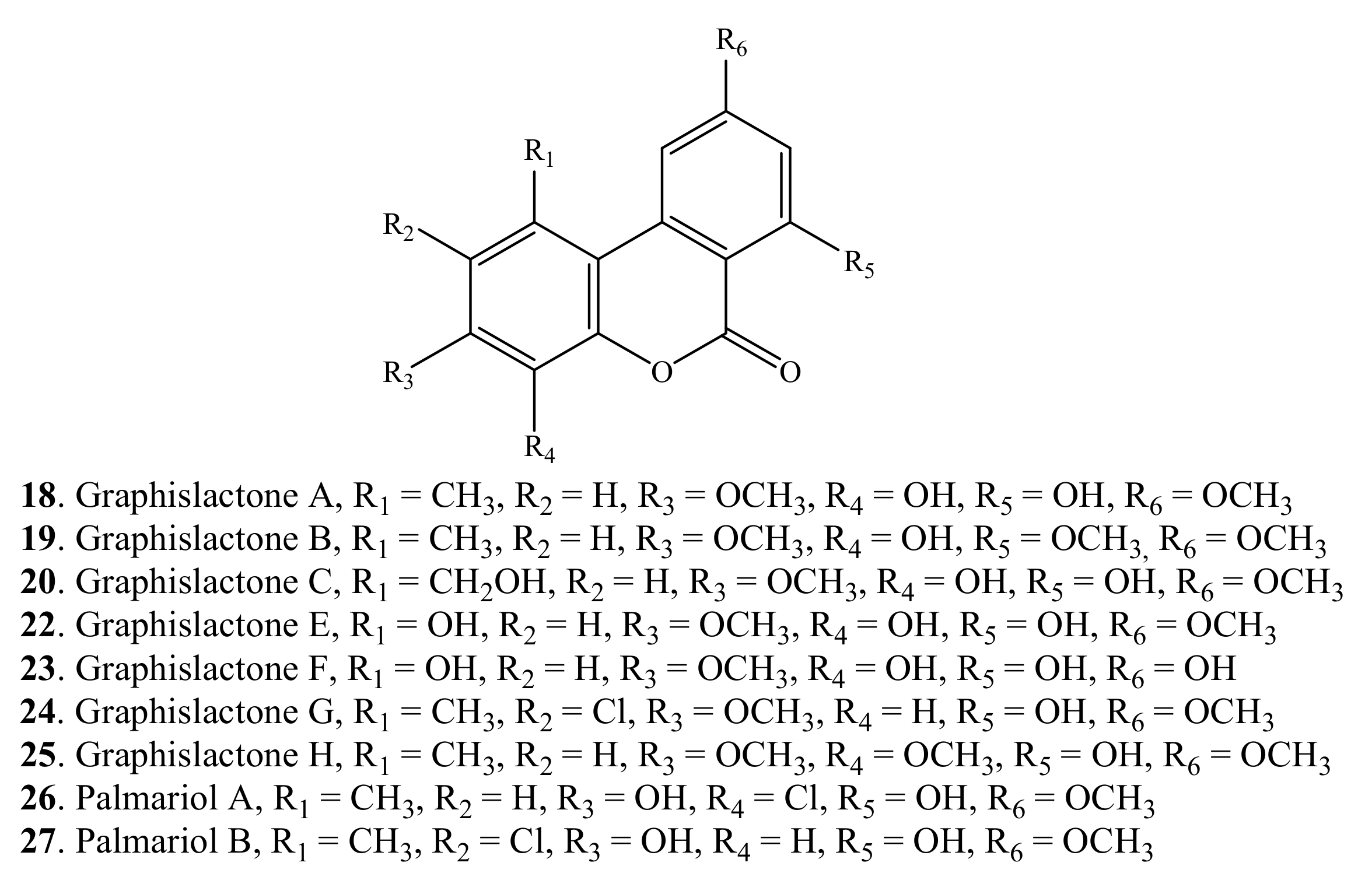
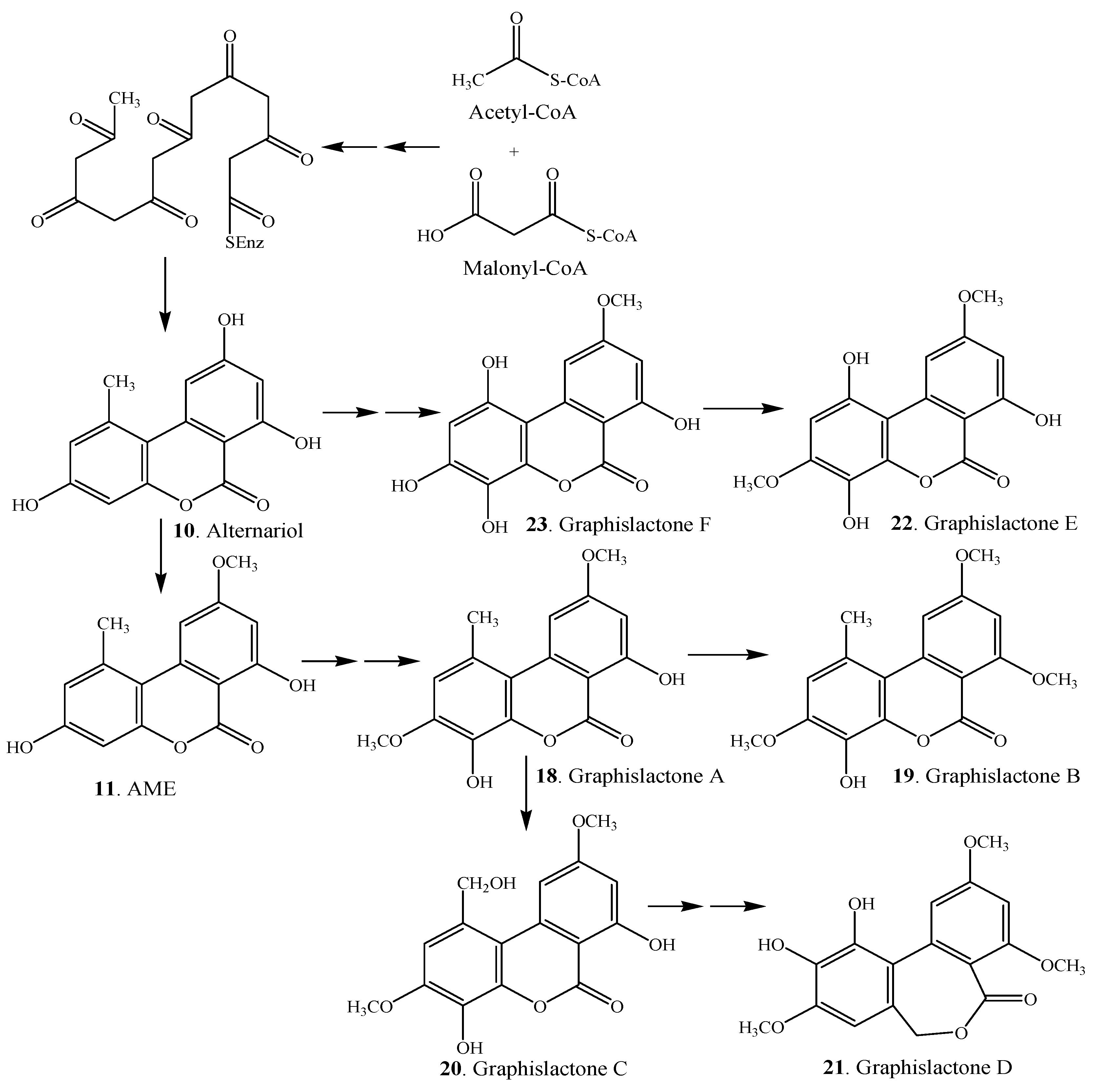
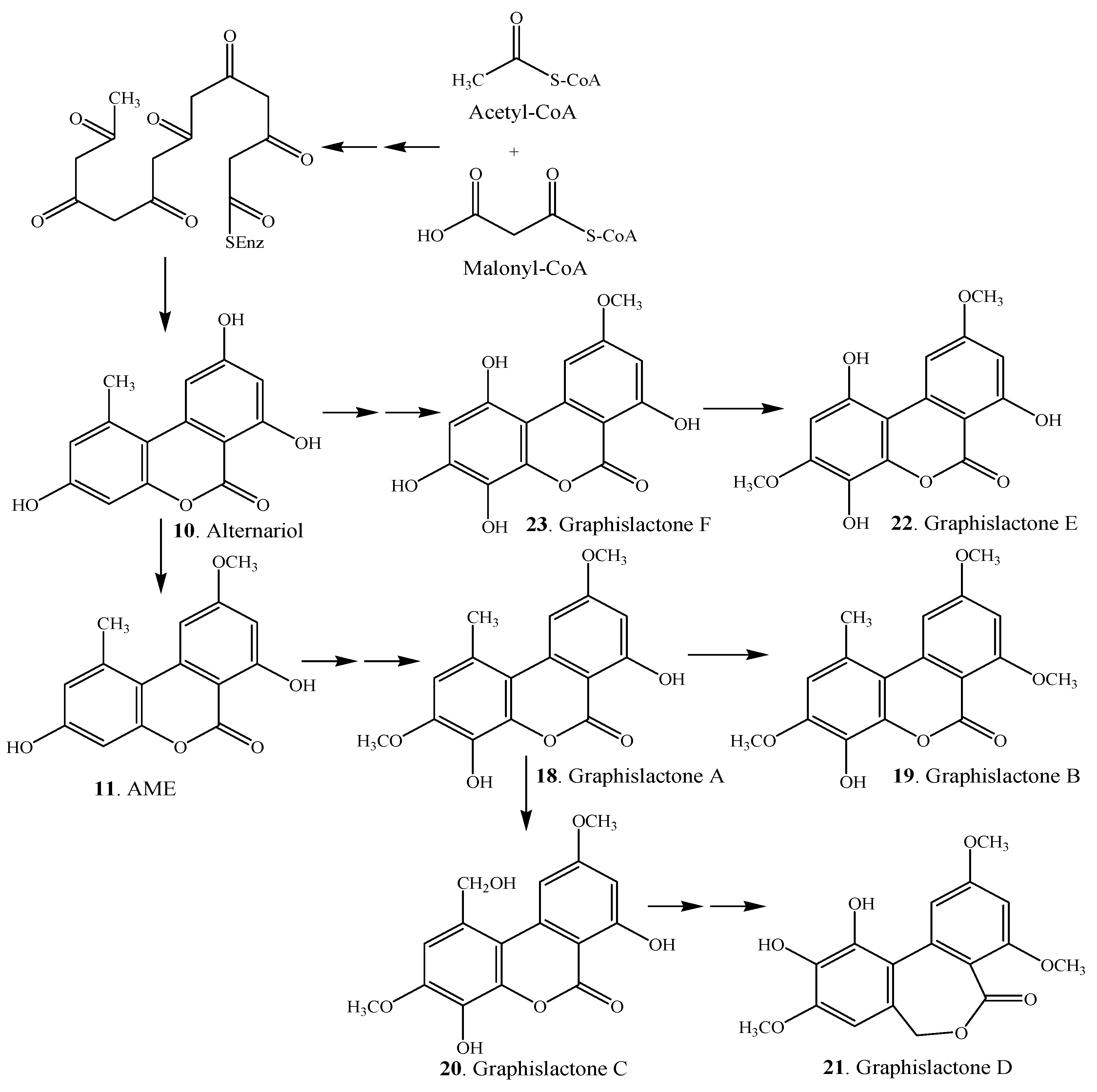
| Graphislactone A (18) | Mycobiont of Graphis scripta var. pulverulenta | [2] |
| Endophytic Cephalosporium acremonium IFB-E007 isolated from Trachelospermum jasminoides | [35] | |
| Endophytic Microsphaeropsis olivacea isolated from Pilgerodendron uviferum | [13] | |
| Graphislactone B (19) | Mycobiont of Graphis scripta var. pulverulenta | [2] |
| Graphislactone C (20) | Mycobiont of Graphis scripta var. pulverulenta | [2] |
| Graphislactone D (21) | Mycobiont of Graphis scripta var. pulverulenta | [2] |
| Graphislactone E (22) | Mycobiont of Graphis scripta | [3] |
| Mycobiont of Graphis prunicola | [3] | |
| Graphislactone F (23) | Mycobiont of Graphis prunicola | [3] |
| Graphislactone G (24) | Endophytic Cephalosporium acremonium IFB-E007 isolated from Trachelospermum jasminoides | [35] |
| Graphislactone H (25) | Endophytic Cephalosporium acremonium IFB-E007 isolated from Trachelospermum jasminoides | [35] |
| Palamriol A (26) | Lachmum palmae | [36] |
| Palmariol B (27) | Lachmum palmae | [36] |
| Endophytic Hyalodendriella sp. Ponipodef12 isolated from the hybrid ‘Neva’ of Populus deltoides × P. nigra | [12] | |
| TMC-264 (28) | Phoma sp. TC 1674 | [47] |


2.2. Dibenzo-α-pyrones from Plants
| Dibenzo-α-pyrone | Plant species (Family) | Reference |
|---|---|---|
| Alternariol 9-methyl ether (11) | Anthocleista djalonensis (Loganiaceae) | [48] |
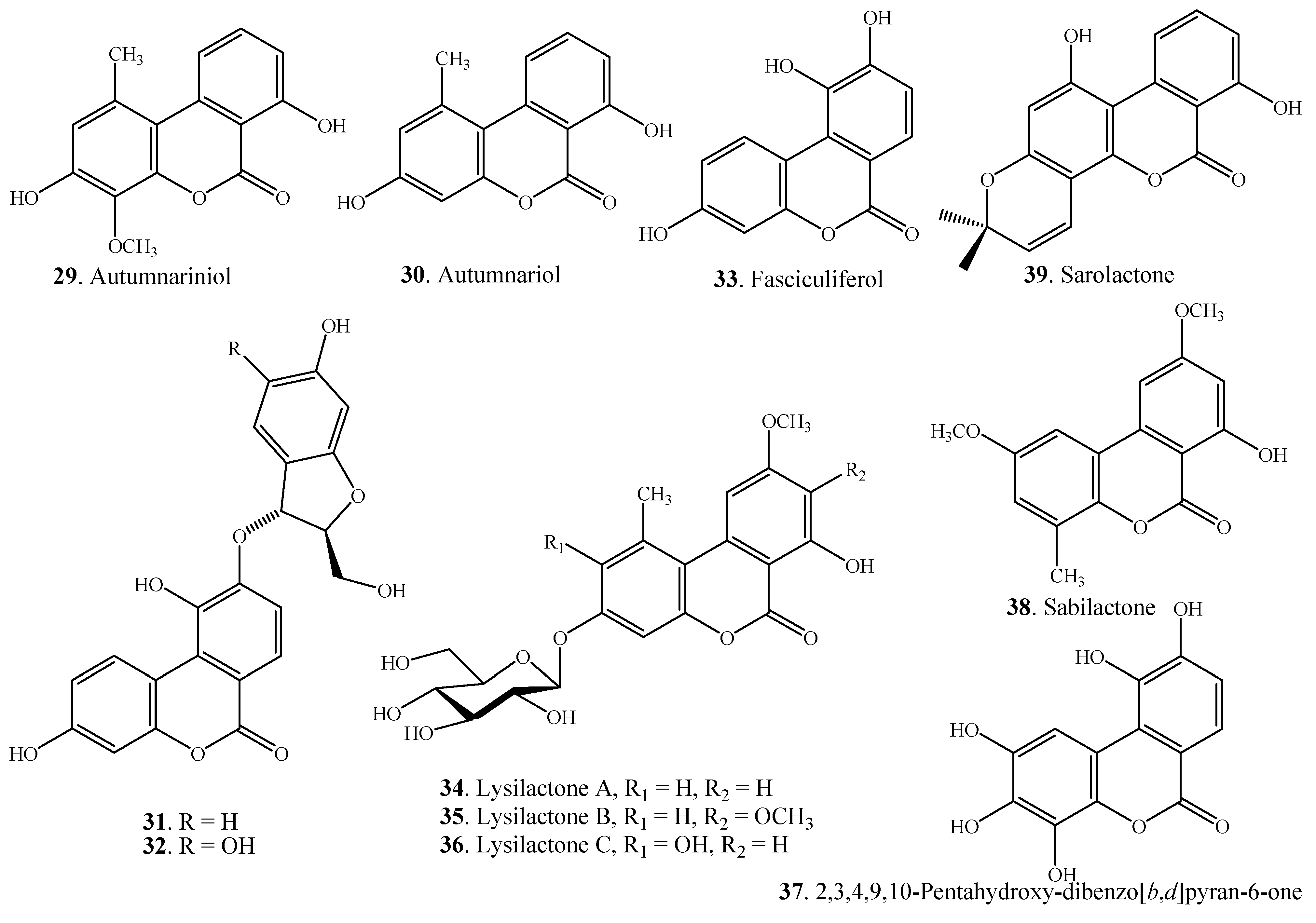
| Autumnariniol (29) | Eucomis autumnalis Graeb (Liliaceae) | [52] |
| Autumnariol (30) | Eucomis autumnalis Graeb (Liliaceae) | [52] |
| (2' S,3'R)-3,10-Dihydroxy-9-O-(6'- hydroxy-2'-hydroxymethyldihydrofuran-3-yl)-dibenzo[b,d]pyran-6-one (31) | Umtiza listerana (Caesalpiniaceae) | [50] |
| (2' S,3'R)-3,10-Dihydroxy-9-O-(5',6'-dihydroxy-2'-hydroxymethyldihydrofuran-3-yl)-dibenzo[b,d]pyran-6-one (32) | Umtiza listerana (Caesalpiniaceae) | [50] |
| Fasciculiferol (33) | Acacia fasciculifera (Mimosaceae) | [51] |
| Umtiza listerana (Caesalpiniaceae) | [50] | |
| Lysilactone A (34) | Lysimachia clethroides (Primulaceae) | [5] |
| Lysilactone B (35) | Lysimachia clethroides (Primulaceae) | [5] |
| Lysilactone C (36) | Lysimachia clethroides (Primulaceae) | [5] |
| 2,3,4,9,10-Pentahydroxy-6 H-dibenzo[b,d]pyran-6-one (37) | Chrozophora senegalensis (Euphorbiaceae) | [56] |
| Polygonum chinense (Polygonaceae) | [57] | |
| Sebastiania chamaelea (Euphorbiaceae) | [56] | |
| Tamarix nilotica (Tamaricaceae) | [58] | |
| Sabilactone (38) | Sabina vulgaris (Cupressaceae) | [54] |
| Sarolactone (39) | Hypericum japonicum (Guttiferae) | [55] |
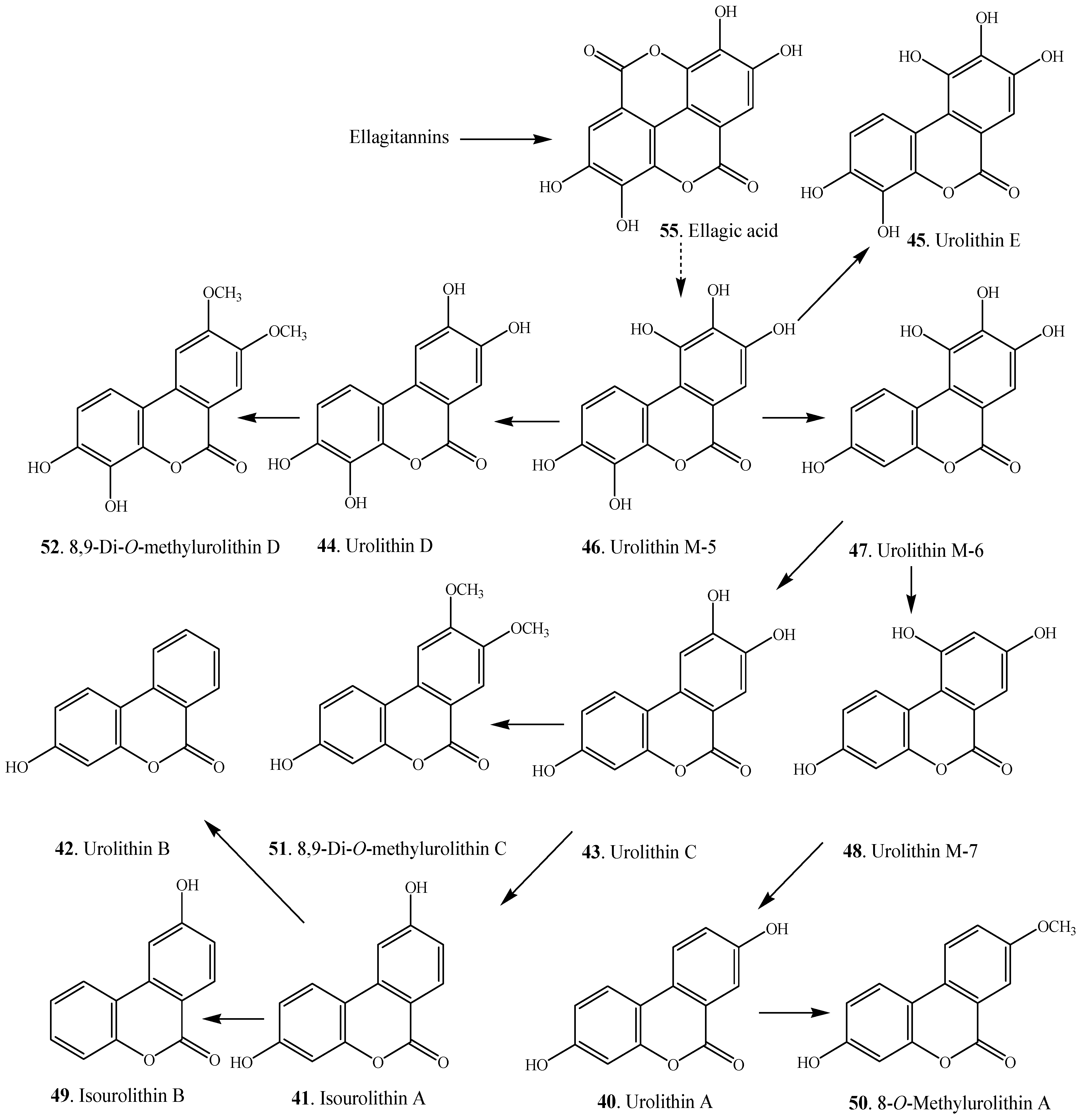
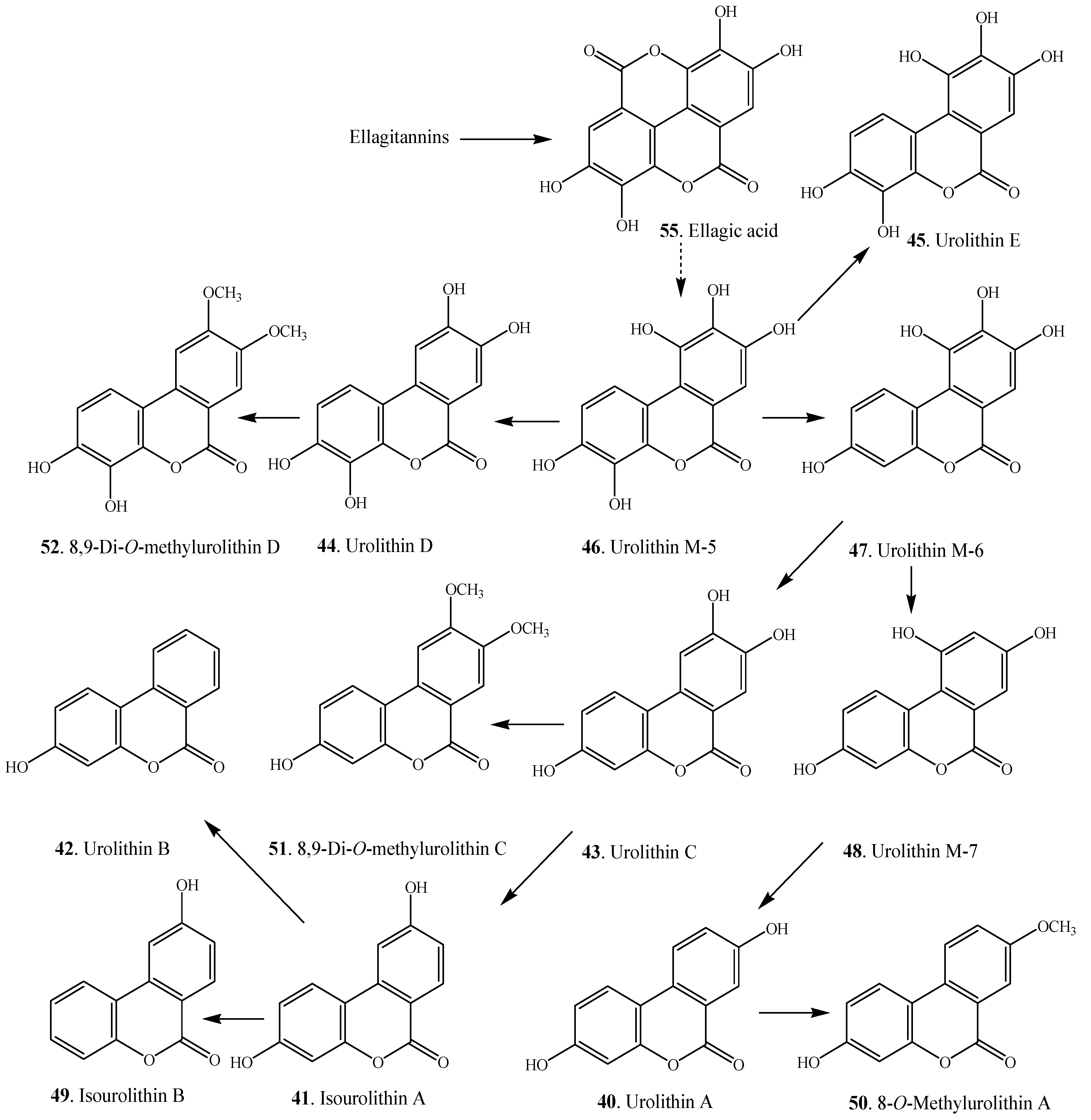
| Urolithin A (40) | Trapa natans (Trapaceae) | [4] |
| Isourolithin A (41) | Trapa natans (Trapaceae) | [4] |
| Urolithin B (42) | Trapa natans (Trapaceae) | [4] |
| Urolithin C (43) | Caesalpinia sappan (Caesalpiniaceae) | [53] |

| Dibenzo-α-pyrone | R1 | R2 | R3 | R4 | R5 | Reference |
|---|---|---|---|---|---|---|
| Urolithin A (40) | OH | H | OH | H | H | [6] |
| Urolithin B (42) | OH | H | H | H | H | [6] |
| Urolithin C (43) | OH | H | OH | OH | H | [6] |
| Urolithin D (44) | OH | OH | OH | OH | H | [59] |
| Urolithin E (45) | OH | OH | OH | H | OH | [59] |
| Urolithin M-5 (46) | OH | OH | OH | OH | OH | [7] |
| Urolithin M-6 (47) | OH | H | OH | OH | OH | [7,59] |
| Urolithin M-7 (48) | OH | H | OH | H | OH | [7,11] |
| Isourolithin A (41) | OH | H | H | OH | H | [7] |
| Isourolithin B (49) | H | H | H | OH | H | [7] |
| 8-O-Methylurolithin A (50) | OH | H | OCH3 | H | H | [6] |
| 8,9-Di-O-methylurolithin C (51) | OH | H | OCH3 | OCH3 | H | [6] |
| 8,9-Di-O-methylurolithin D (52) | OH | OH | OCH3 | OCH3 | H | [6] |
2.3. Dibenzo-α-pyrones Produced by Transformation of Intestinal Bacteria
2.4. Dibenzo-α-pyrones from Bacteria
3. Biosynthesis and Biotransformation
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

4. Biological Activities and Functions
4.1. Toxicity on Human and Animals
4.2. Cytotoxic Activity
4.3. Phytotoxicity
4.4. Antioxidant Activity
4.5. Antiallergic Activity
4.6. Other Bioactivities
5. Conclusions and Future Perspectives
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Lou, J.; Fu, L.; Peng, Y.; Zhou, L. Metabolites from Alternaria fungi and their bioactivities. Molecules 2013, 18, 5891–5935. [Google Scholar] [CrossRef]
- Tanahashi, T.; Kuroishi, M.; Kuwahara, A.; Nagakura, N.; Hamada, N. Four phenolics from the cultured lichen mycobiont of Graphis scripta var. pulverulenta. Chem. Pharm. Bull. 1997, 45, 1183–1185. [Google Scholar] [CrossRef]
- Tanahashi, T.; Takenaka, Y.; Nagakura, N.; Hamada, N. 6H-Dibenzo[b,d]pyran-6-one derivatives from the cultured lichen mycobionts of Graphis spp. and their biosynthetic origin. Phytochemistry 2003, 62, 71–75. [Google Scholar] [CrossRef]
- Shirataki, Y.; Toda, S. Antioxidative effects of dibenzo-α-pyrones in fruits of Trapa natans on lipid peroxidation. Nat. Med. 2001, 55, 247–250. [Google Scholar]
- Liang, D.; Luo, H.; Liu, Y.-F.; Hao, Z.-Y.; Wang, Y.; Zhang, C.-L.; Zhang, Q.-J.; Chen, R.-Y.; Yu, D.-Q. Lysilactones A-C, three 6H-dibenzo[b,d]pyran-6-one glycosides from Lysimachia clethroides, total synthesis of lysilactone A. Tetrahedron 2013, 69, 2093–2097. [Google Scholar] [CrossRef]
- Bialonska, D.; Kasimsetty, S.G.; Khan, S.I.; Ferreira, D. Urolithins, intestinal microbial metabolites of pomegranate ellagitannins, exhibit potent antioxidant activity in a cell-based assay. J. Agric. Food Chem. 2009, 57, 10181–10186. [Google Scholar] [CrossRef]
- Gonzalez-Barrio, R.; Truchado, P.; Ito, H.; Espin, J.C.; Tomas-Barberan, F.A. UV and MS identification of urolithins and nasutins, and bioavailable metabolites of ellagic acid in different mammals. J. Agric. Food Chem. 2011, 59, 1152–1162. [Google Scholar] [CrossRef]
- Moreno, M.A.P.; Alonso, I.G.; Martin de Santos, R.; Lacarra, T.G. The role of the genus Alternaria. in mycotoxin production and human diseases. Nutr. Hosp. 2012, 27, 1772–1781. [Google Scholar]
- Aly, A.H.; Edrada-Ebel, R.; Indriani, I.D.; Wray, V.; Muller, W.E.G.; Totzke, F.; Zirrgiebel, U.; Schachtele, C.; Kubbutat, M.H.G.; Lin, W.H.; et al. Cytotoxic metabolites from the fungal endophyte Alternaria sp. and their subsequent detection in its host plant Polygonum senegalense. J. Nat. Prod. 2008, 71, 972–980. [Google Scholar] [CrossRef]
- Watanabe, I.; Kakishima, M.; Adachi, Y.; Nakajima, H. Potential mycotoxin productivity of Alternaria alternata isolated from garden trees. Mycotoxins 2007, 57, 3–9. [Google Scholar] [CrossRef]
- Jeong, S.-J.; Kim, N.-Y.; Kim, D.-H.; Kang, T.-H.; Ahn, N.-H.; Miyamoto, T.; Higuchi, R.; Kim, Y.-C. Hyaluronidase inhibitory active 6H-dibenzo[b,d]pyran-6-ones from the feces of Trogopterus xanthipes. Planta Med. 2000, 66, 76–77. [Google Scholar]
- Meng, X.; Mao, Z.; Lou, J.; Xu, L.; Zhong, L.; Peng, Y.; Zhou, L.; Wang, M. Benzopyranones from the endophytic fungus Hyalodendriella sp. Ponipodef12 and their bioactivities. Molecules 2012, 17, 11303–11314. [Google Scholar] [CrossRef]
- Hormazabal, E.; Schmeda-Hirschmann, G.; Astudillo, L.; Rodriguez, J.; Theoduloz, C. Metabolites from Microsphaeropsis olicacea, an endophytic fungus of Pilgerodendron ufiferum. Z. Naturforsch. C 2005, 60, 11–21. [Google Scholar]
- Teske, J.; Deiters, A. A cyclotrimerization route to cannabinois. Org. Lett. 2008, 10, 2195–2198. [Google Scholar] [CrossRef]
- Edwards, J.P.; West, S.J.; Marschke, K.B.; Mais, D.E.; Gottardis, M.M.; Jones, T.K. 5-Aryl-1,2-dihydro-5H-chromeno[3,4-f]quinolines as potent, orally active, nonsteroidal progesterone receptor agonists: The effect of D-ring substituents. J. Med. Chem. 1998, 41, 303–310. [Google Scholar] [CrossRef]
- Coghlan, M.J.; Kym, P.; Elmore, S.W.; Wang, A.X.; Luly, J.R.; Wilcox, D.; Stashko, M.; Lin, C.-W.; Miner, J.; Tyree, C.; et al. Synthesis and characterization of non-steroidal ligands for the glucocorticoid receptor: Selective quinoline derivatives with prednisolone-equivalent functional activity. J. Med. Chem. 2001, 44, 2879–2885. [Google Scholar] [CrossRef]
- Schmidt, J.M.; Tremblay, G.B.; Page, M.; Mercure, J.; Feher, M.; Dunn-Dufault, R.; Peter, M.G.; Redden, P.R. Synthesis and evaluation of a novel nonsteroidal-specific endothelial cell proliferation inhibitor. J. Med. Chem. 2003, 46, 1289–1292. [Google Scholar] [CrossRef]
- Sashidhara, K.V.; Kumar, A.; Kumar, M.; Sonkar, R.; Bhatia, G.; Khanna, A.K. Novel coumarin derivatives as potential antidyslipidemic agents. Bioorg. Med. Chem. Lett. 2010, 20, 4248–4251. [Google Scholar] [CrossRef]
- Jiao, P.; Gloer, J.B.; Campbell, J.; Shearer, C.A. Altenuene derivatives from an unidentified freshwater fungus in the family Tubeufiaceae. J. Nat. Prod. 2006, 69, 612–615. [Google Scholar] [CrossRef]
- Visconti, A.; Bottalico, A.; Solfrizzo, M.; Palmisano, F. Isolation and structure elucidation of isoaltenuene, a new metabolite of Alternaria alternata. Mycotoxin Res. 1989, 5, 69–76. [Google Scholar]
- Bradburn, N.; Coker, R.D.; Blunden, G.; Turner, C.H.; Crabb, T.A. 5'-Epialtenuene and neoaltenuene, dibenzo-α-pyrones from Alternaria alternata cultured on rice. Phytochemistry 1994, 35, 665–669. [Google Scholar] [CrossRef]
- Hussain, H.; Krohn, K.; Ullah, Z.; Draeger, S.; Schulz, B. Bioactive chemical constituents of two endophytic fungi. Biochem. Syst. Ecol. 2007, 35, 898–900. [Google Scholar] [CrossRef]
- Qin, J.-C.; Zhang, Y.-M.; Hu, L.; Ma, Y.-T.; Gao, J.-M. Cytotoxic metabolites produced by Alternaria no.28, an endophytic fungus isolated from Ginkgo biloba. Nat. Prod. Commun. 2009, 4, 1473–1476. [Google Scholar]
- Wu, S.; Chen, Y.; Li, Z.; Yang, L.; Li, S. Metabolites of the endophytic fungus Alternaria sp. PR-14 of Paeonia delavayi. Nat. Prod. Res. Dev. 2011, 23, 850–852. [Google Scholar]
- Sun, J.; Awakawa, T.; Noguchi, H.; Abe, I. Induced production of mycotoxins in an endophytic fungus from the medicinal plant Datura stramonium L. Bioorg. Med. Chem. Lett. 2012, 22, 6397–6400. [Google Scholar] [CrossRef]
- Zheng, Z.; Liu, Y.; Fan, L. Antioxidant activity and structure identification of metabolites of an endophytic fungus Alternaria. sp. N.SBA10 isolated from Scutellaria baicalensis. Mycosystema 2012, 31, 917–923. [Google Scholar]
- Griffin, G.F.; Chu, F.S. Toxicity of the Alternaria metabolites alternariol, alternariol methyl ether, altenuene, and tenuazonic acid in the chicken embryo assay. Appl. Environ. Microbiol. 1983, 46, 1420–1422. [Google Scholar]
- Gu, W. Bioactive metabolites from Alternaria brassicicola ML-P08, an endophytic fungus residing in Malus halliana. World J. Microbiol. Biotechnol. 2009, 25, 1677–1683. [Google Scholar] [CrossRef]
- Sun, H.; Gao, S.; Li, X.; Li, C.; Wang, B. Chemical constituents of marine mangrove-derived endophytic fungus Alternaria tenuissima EN-192. Chin. J. Oceanol. Limnol. 2013, 31, 464–470. [Google Scholar] [CrossRef]
- Yang, Z.; Yin, Y.; Wang, Z.; Yang, T.; Chen, D. Cytotoxic metabolites of endophytic fungus Colletotrichum sp. from Aristolochia sp. Nat. Prod. Res. Dev. 2012, 24, 329–332. [Google Scholar]
- An, Y.-H.; Zhao, T.-Z.; Miao, J.; Liu, G.-T.; Zheng, Y.-Z.; Xu, Y.-M.; van Etten, R.L. Isolation, identification, and mutagenicity of alternariol monomethyl ether. J. Agric. Food Chem. 1989, 37, 1341–1343. [Google Scholar] [CrossRef]
- Evans, N.; Mcroberts, N.; Hill, R.A.; Marshall, G. Phytotoxin production by Alternaria linicola and phytoalexin production by the linseed host. Ann. Appl. Biol. 1996, 129, 415–431. [Google Scholar] [CrossRef]
- Pero, R.W.; Main, C.E. Chlorosis of tobacco induced by alternariol monomethyl ether produced by Alternaria tenuis. Phytopathology 1970, 60, 1570–1573. [Google Scholar] [CrossRef]
- Jeon, Y.-T.; Ryu, K.-H.; Kang, M.-K.; Park, S.-H.; Yun, H.; Qt, P.; Kim, S.-U. Alternariol monomethyl ether and α,β-dehydrocurvularin from endophytic fungi Alternaria spp. inhibit appressorium formation of Magnaporthe grisea. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 39–42. [Google Scholar]
- Zhang, H.-W.; Huang, W.-Y.; Song, Y.-C.; Chen, J.-R.; Tan, R.-X. Four 6H-dibenzo[b,d]pyran-6-one derivatives produced by the endophyte Cephalosporium acremonium IFB-E007. Helv. Chim. Acta 2005, 88, 2861–2864. [Google Scholar] [CrossRef]
- Matumoto, T.; Hosoya, T.; Shigemori, H. Palmariols A and B, two new chlorinated dibenzo-α-pyrones from discomycete Lachnum palmae. Heterocycles 2010, 81, 1231–1237. [Google Scholar] [CrossRef]
- Chu, F.S. Isolation of altenuisol and altertoxins I and II, minor mycotoxins elaborated by Alternaria. J. Am. Oil Chem. Soc. 1981, 58, 1006–1008. [Google Scholar] [CrossRef]
- Rosett, T.; Sankhala, R.H.; Stickings, C.E.; Taylor, M.E.U.; Thomas, R. Studies in the biochemistry of micro-organisms. 103. Metabolites of Alternaria tenuis Auct.: Culture filtrate products. Biochem. J. 1957, 67, 390–400. [Google Scholar]
- Thomas, R. Studies in the biosynthesis of fungal metabolites IV. Alternariol monomethyl ether and its relation to other phenolic metabolites of Alternaria tenuis. Biochem. J. 1961, 80, 234–240. [Google Scholar]
- Thomas, R.; Nemecek, G.; Podlech, J. Structure confirmation of altertenuol. Nat. Prod. Res. 2013, 27, 2053–2054. [Google Scholar] [CrossRef]
- Pero, R.W.; Harvan, D.; Blois, M.C. Isolation of the toxin, altenuisol, from the fungus, Alternaria tenuis Auct. Tetrahedron Lett. 1973, 12, 945–948. [Google Scholar] [CrossRef]
- Kameda, K.; Aoki, H.; Namiki, M. An alternative structure for botrallin a metabolite of Botrytis allii. Tetrahedron Lett. 1974, 1, 103–106. [Google Scholar] [CrossRef]
- Zhong, L.; Xu, L.; Meng, X.; Peng, Y.; Chen, Y.; Sui, P.; Wang, M.; Zhou, L. Botrallin from the endophytic fungus Hyalodendriella sp. Ponipodef12 and its antimicrobial activity. Afr. J. Biotechnol. 2011, 10, 18174–18178. [Google Scholar]
- Murakami-Nakai, C.; Maida, N.; Yonezawa, Y.; Kuriyama, I.; Kamisuki, S.; Takahashi, S.; Sugawara, F.; Yoshida, H.; Sakaguchi, K.; Mizushina, Y. The effects of dehydroaltenusin, a novel mammalian DNA polymerase α inhibitor, on cell proliferation and cell cycle progression. Biochim. Biophys. Acta 2004, 1674, 193–199. [Google Scholar]
- Maeda, N.; Kokai, Y.; Ohtani, S.; Sahara, H.; Kuriyama, I.; Kamisuki, S.; Takahashi, S.; Sakaguchi, K.; Sugawara, F.; Yoshida, H. Anti-tumor effects of dehydroaltenusin, a specific inhibitor of mammalian DNA polymerase α. Biochem. Biophys. Res. Commun. 2007, 352, 390–396. [Google Scholar] [CrossRef]
- Nakanishi, S.; Toki, S.; Saitoh, Y.; Tsukuda, E.; Kawahara, K.; Ando, K.; Matsuda, Y. Isolation of myosin light chain kinase inhibitors from microorganisms: Dehydroaltenusin, altenusin, atrovenetinone, and cyclooctasulfur. Biosci. Biotech. Biochem. 1995, 59, 1333–1335. [Google Scholar] [CrossRef]
- Sakurai, M.; Nishio, M.; Yamamoto, K.; Okuda, T.; Kawano, K.; Ohnuki, T. TMC-264, a novel antiallergic heptaketide produced by the fungus Phoma sp. TC 1674. Org. Lett. 2003, 5, 1083–1085. [Google Scholar] [CrossRef]
- Onocha, P.; Okopie, D.A.; Connolly, J.D.; Roycroft, D.S. Monoterpene diol, iridoid glucoside and dibenzo-α-pyrone from Anthocleista djalonensis. Phytochemistry 1995, 40, 1183–1189. [Google Scholar] [CrossRef]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-derived bioactive compounds produced by endophytic fungi. Mini-Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef]
- Burger, A.P.N.; Brandt, E.V.; Roux, D.G. O-(Dihydrobenzofuranyl)-dibenzo-α-pyrones from Umtiza listerana. Phytochemistry 1983, 22, 2813–2817. [Google Scholar] [CrossRef]
- Van Heerden, F.R.; Brandt, E.V.; Ferreira, D.; Roux, D.G. Metabolites from the purple heartwoods of the Mimosoideae. Part 4. Acacia fasciculifera F. Muell ex. Benth: Fasciculiferin, fasciculiferol, and the synthesis of 7-aryl- and 7-flavanylpeltogynoids. .J. Chem. Soc. Perkin Trans. 1 1981, 9, 2483–2490. [Google Scholar]
- Sidwell, W.T.L.; Fritz, H.; Tamm, C. Autumnariol and autumnariniol, two new dibenzo-α-pyrones from Eucomis autumnalis. Detection of long-range coupling over six bonds in the NMR spectra. Helv. Chim. Acta 1971, 54, 207–215. [Google Scholar] [CrossRef]
- Chen, Y.-P.; Liu, L.; Zhou, Y.-H.; Wen, J.; Jiang, Y.; Tu, P.-F. Chemical constituents from sappan lignum. J. Chin. Pharm. Sci. 2008, 17, 82–86. [Google Scholar]
- Fang, S.; Gu, Y.; Yu, H.; Musadillin, S. The antitumor constituents from Sabina vulgaris Ant. Acta. Bot. Sin. 1989, 31, 382–388. [Google Scholar]
- Ishiguro, K.; Yamaki, M.; Kashihara, M.; Takagi, S.; Isoi, K. A chromene from Hypericum japonicum. Phytochemistry 1990, 29, 1010–1011. [Google Scholar] [CrossRef]
- Garcia-Alvarez, M.-C.; Moussa, I.; Njomnang Soh, P.; Nongonierma, R.; Abdoulaye, A.; Nicolau-Travers, M.-L.; Fabre, A.; Wdzieczak-Bakala, J.; Ahond, A.; Poupat, C.; et al. Both plants Sebastiania chamaelea from Niger and Chrozophora senegalensis from Senegal used in African traditional medicine in malaria treatment share a same active principle. J. Ethnopharmacol. 2013, 149, 676–684. [Google Scholar] [CrossRef]
- Xie, X.; Wu, P.; Lin, L.; Liu, M.; Wei, X. Identification of chemical constituents from Polygonum chinense Linn. J. Trop. Subtrop. Bot. 2007, 15, 450–454. [Google Scholar]
- Nawwar, M.A.M.; Souleman, A.M.A. 3,4,8,9,10-Pentahydroxy-dibenzo[b,d]pyran-6-one from Tamarix nilotica. Phytochemistry 1984, 23, 2966–2967. [Google Scholar] [CrossRef]
- Gil, M.I.; Tomás-Barberán, F.A.; Hess-Pierce, B.; Holcroft, D.M.; Kader, A.A. Antioxidant activity of pomegranate juice and its relationship with phenolic composition and processing. J. Agric. Food Chem. 2000, 48, 4581–4589. [Google Scholar]
- Larrosa, M.; García-Conesa, M.T.; Espín, J.C.; Tomás-Barberán, F.A. Ellagitannins, ellagic acid and vascular health. Mol. Aspects Med. 2010, 31, 513–539. [Google Scholar]
- Landete, J.M. Ellagitannins, ellagic acid and their derived metabolites. A review about source, metabolism, function and health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Cerda, B.; Espin, J.C.; Parra, S.; Martinez, P.; Tomas-Barberan, F.A. The potent in vitro antioxidant ellagitannins from pomegranate juice are metabolized into bioavailable but poor antioxidant hydroxyl-6H-dibenzopyran-6-one derivatives by the colonic microflora of healthy humans. Eur. J. Nutr. 2004, 43, 205–220. [Google Scholar]
- Garcia-Villalba, R.; Beltran, D.; Espin, J.C.; Selma, M.V.; Tomas-Barberan, F.A. Time course production of urolithins from ellagic acid by human gut microbiota. J. Agric. Food Chem. 2013, 61, 8797–8806. [Google Scholar] [CrossRef]
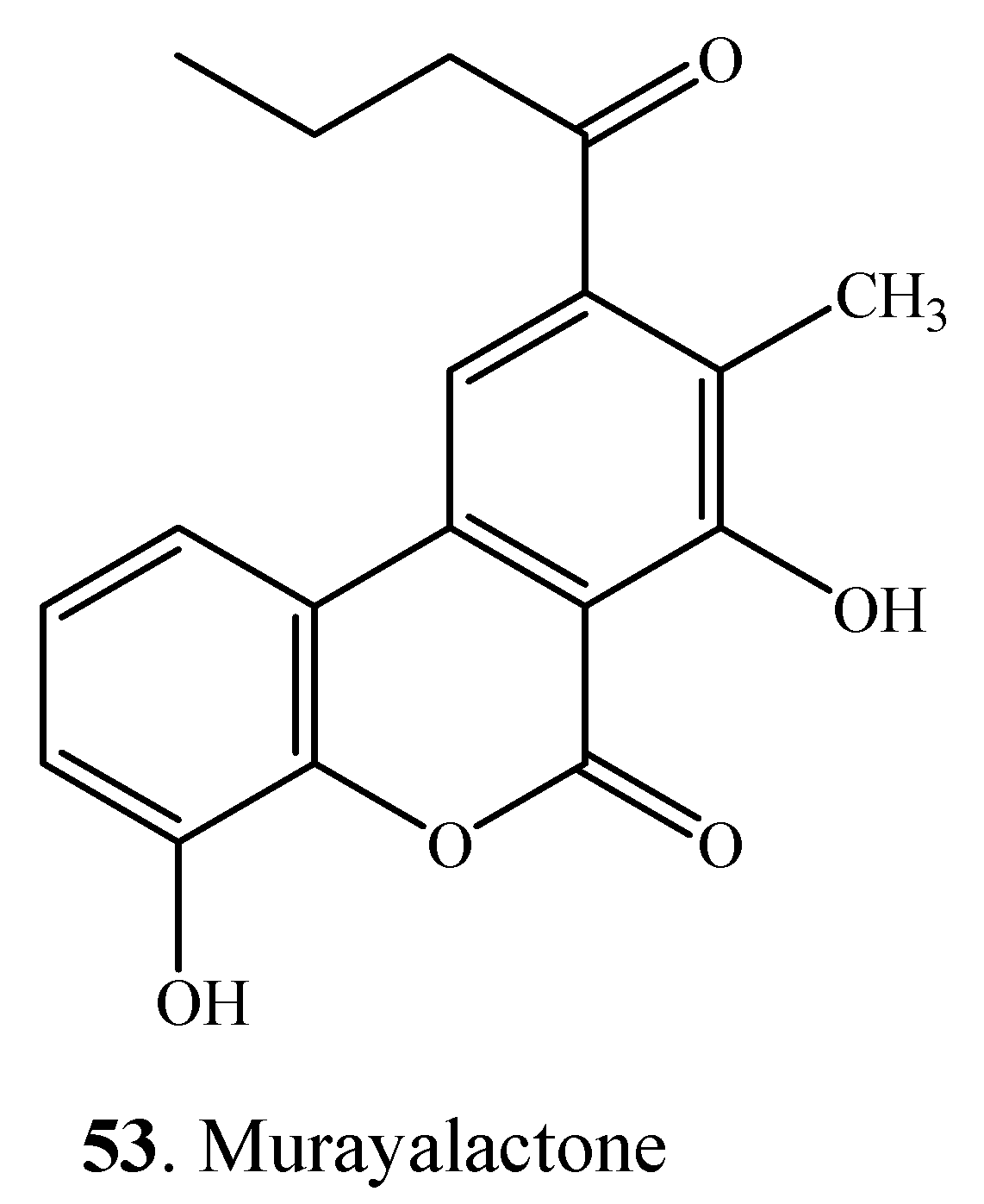
- Melville, C.; Gould, S.J. Murayalactone, a dibenzo-α-pyrone from Streptomyces murayamaensis. J. Nat. Prod. 1994, 57, 597–601. [Google Scholar] [CrossRef]
- Stinson, E.E.; Wise, W.B.; Moreau, M.; Jurewicz, A.J.; Pfeffer, P.E. Alternariol: Evidence for biosynthesis via norlichexanthone. Can. J. Chem. 1986, 64, 1590–1594. [Google Scholar] [CrossRef]
- Saha, D.; Fetzner, R.; Burkhardt, B.; Podlech, J.; Metzler, M.; Dang, H.; Lawrence, C.; Fischer, R. Identification of a polyketide synthase required for alternariol (AOH) and alternariol-9-methyl ether (AME) formation in Alternaria alternata. PLoS One 2012, 7, e40564. [Google Scholar]
- Werner, R.A.; Sossmann, A.; Schwarz, C.; Bacher, A.; Schmidt, H.-L.; Eisenreich, W. Biosynthesis of gallic acid in Rhus typhia: Discrimination between alternative pathways from natural oxygen isotope abundance. Phytochemistry 2004, 65, 2809–2813. [Google Scholar] [CrossRef]
- Niemetz, R.; Gross, G.G. Enzymology of gallotannin and ellagitannin biosynthesis. Phytochemistry 2005, 66, 2001–2011. [Google Scholar] [CrossRef]
- Dasenbrock, J.; Simpson, T.J. Alternariol is not biosynthesized via norlichexanthone. J. Chem. Sco. Chem. Commun. 1987, 1987, 1235–1236. [Google Scholar] [CrossRef]
- Fisch, K.M.; Gillaspy, A.F.; Gipson, M.; Henrikson, J.C.; Hoover, A.R.; Jackson, L.; Najar, F.Z.; Wagele, H.; Cichewicz, R.H. Chemical induction of silent biosynthetic pathway transcription in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2009, 36, 1199–1213. [Google Scholar] [CrossRef]
- Cerda, B.; Tomas-Barberan, F.A.; Espin, J.C. Metabolism of antioxidant and chemopreventive ellagitannins from strawberries, raspberries, walnuts, and oak-aged wine in humans: Identification of biomarkers and individual variability. J. Argic. Food Chem. 2005, 53, 227–235. [Google Scholar] [CrossRef]
- Espin, J.C.; Gonzalez-Barrio, R.; Cerda, B.; Lopez-Bote, C.; Tomas-Barberan, F.A. Iberian pig as a model to clarify obscure points in the bioavailability and metabolism of ellagitannins in humans. J. Agric. Food Chem. 2007, 55, 10476–10485. [Google Scholar] [CrossRef]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Zajkowski, P.; Grabarkiewicz-Szcesna, J.; Schmidt, R. Toxicity of mycotoxins produced by four Alternaria species to Artemia salina larvae. Mycotoxin Res. 1991, 7, 11–15. [Google Scholar] [CrossRef]
- Panigrahi, S.; Dallin, S. Toxicity of the Alternaria spp. metabolites, tenuazonic acid, alternariol, altertoxin-I, and alternariol monomethyl ether to brine shrimp (Artemia. salina L.) larvae. J. Sci. Food Agric. 1994, 66, 493–496. [Google Scholar] [CrossRef]
- Bensassi, F.; Gallerne, C.; Sharaf El Dein, O.; Hajlaoui, M.R.; Bacha, H.; Lemaire, C. Cell death induced by the Alternaria mycotoxin alternariol. Toxicol. In Vitro 2012, 26, 915–923. [Google Scholar] [CrossRef]
- Schreck, I.; Deigendesch, U.; Burkhardt, B.; Marko, D.; Weiss, C. The Alternaria mycotoxins alternariol and alternariol methyl ether induce cytochrome P450 1A1 and apoptosis in murine hepatoma cells dependent on the aryl hydrocarbon receptor. Arch. Toxicol. 2012, 86, 625–632. [Google Scholar] [CrossRef]
- Mizushina, Y.; Maeda, N.; Kuriyama, I.; Yoshida, H. Dehydroalternusin is a specific inhibitor of mammalian DNA polymerase α. Expert Opin. Inv. Drug. 2011, 20, 1523–1534. [Google Scholar] [CrossRef]
- Mizushina, Y.; Kamisuki, S.; Mizuno, T.; Takemura, M.; Asahara, H.; Linn, S.; Yamaguchi, T.; Matsukage, A.; Hanaoka, F.; Yoshida, S.; et al. Dehydroaltenusin, a mammalian DNA polymerase a inhibitor. J. Biol. Chem. 2000, 275, 33957–33961. [Google Scholar]
- Selma, M.V.; Espin, J.C.; Tomas-Barberan, F.A. Interaction between phenolics and gut microbiota: Role in human health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef]
- Kasimsetty, S.G.; Bialonska, D.; Reddy, M.K.; Thornton, C.; Willett, K.L.; Ferreira, D. Effects of pomergranate chemical constituents/intestinal microbial metabolites on CYP1B1 in 22Rv1 prostate cancer cells. J. Agric. Food Chem. 2009, 57, 10636–10644. [Google Scholar] [CrossRef]
- Qiu, Z.; Zhou, B.; Jin, L.; Yu, H.; Liu, L.; Liu, Y.; Qin, C.; Xie, S.; Zhu, F. In vitro antioxidant and antiproliferative effects of ellagic acid and its colonic metabolite, urolithins, on human baldder cancer T24 cells. Food Chem. Toxicol. 2013, 59, 428–437. [Google Scholar] [CrossRef]
- Graniti, A. Phytotoxins and their involvement in plant diseases. Experientia 1991, 47, 751–755. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Biesenthal, C.J.; Zaharia, I.L. Comparison of the phytotoxic activity of the phytotoxin destruxin B and four natural analogs. Plant Sci. 2000, 156, 185–192. [Google Scholar] [CrossRef]
- Montemurro, N.; Visconti, A. Alternaria Metabolites—Chemical and Biological Data. In Alternaria: Biology, Plant Disease and Metabolites; Chelkowski, J., Visconti, A., Eds.; Elsevier: Amsterdam, The Netherlands, 1992; pp. 449–557. [Google Scholar]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef]
- Ishimoto, H.; Shibata, M.; Myojin, Y.; Ito, H.; Sugimoto, Y.; Tai, A.; Hatano, T. In vivo anti-inflammatoy and antioxidant properties of ellagitannin metabolite urolithin A. Bioorg. Med. Chem. Lett. 2011, 21, 5901–5904. [Google Scholar] [CrossRef]
- Kallio, T.; Kallio, J.; Jaakkola, M.; Maki, M.; Kilpelaimen, P.; Virtanen, V. Urolithins display both antioxidant and pro-oxidant activities depending on assay system and conditions. J. Agric. Food Chem. 2013, 61, 10720–10729. [Google Scholar] [CrossRef]
- Sakurai, M.; Nishio, M.; Yamamoto, K.; Okuda, T.; Kawano, K.; Ohnuki, T. TMC-264, a novel inhibitor of STAT6 activation produced by Phoma sp. TC 1674. J. Antibiot. 2003, 56, 513–519. [Google Scholar] [CrossRef]
- Tatsuta, K.; Furuyama, A.; Yano, T.; Suzuki, Y.; Ogura, T.; Hosokawa, S. The first total synthesis and structural determination of TMC-264. Tetrahedron Lett. 2008, 49, 4036–4039. [Google Scholar] [CrossRef]
- Larrosa, M.; Gonzalez-Sarrias, A.; Garcia-Conesa, M.T.; Tomas-Barberan, F.A.; Espin, J.C. Urolithins, ellagic acid-derived metabolites produced by human colonic microflora, exhibit estrogenic and antiestrogenic activities. J. Agric. Food Chem. 2006, 54, 1611–1620. [Google Scholar] [CrossRef]
- Beresteskiy, A.O. A review of fungal phytotoxins: From basic studies to practical use. Appl. Biochem. Microbiol. 2008, 44, 453–465. [Google Scholar] [CrossRef]
- Brase, S.; Encinas, A.; Keck, J.; Nising, C.F. Chemistry and biology of mycotoxins and related fungal metabolites. Chem. Rev. 2009, 109, 3903–3990. [Google Scholar] [CrossRef]
- Altemoller, M.; Podlech, J.; Fenske, D. Total synthesis of altenuene and isoaltenuene. Eur. J. Org. Chem. 2006, 2006, 1678–1684. [Google Scholar] [CrossRef]
- Altemoller, M.; Podlech, J. Total synthesis of neoaltenuene. Eur. J. Org. Chem. 2009, 2009, 2275–2282. [Google Scholar] [CrossRef]
- Koch, K.; Podlech, J.; Pfeiffer, E.; Metzler, M. Total synthesis of alternariol. J. Org. Chem. 2005, 70, 3275–3276. [Google Scholar] [CrossRef]
- Nemecek, G.; Cudaj, J.; Podlech, J. Revision of the structure and total of synthesis of altenuisol. Eur. J. Org. Chem. 2012, 2012, 3863–3870. [Google Scholar] [CrossRef]
- Kuramochi, K.; Kuriyama, I.; Mori, M.; Kamisuki, S.; Takahashi, S.; Tsubaki, K.; Sugawara, F.; Sakaguchi, K.; Yoshida, H.; Mizushina, Y. Syntheses of dehydroaltenusin, a selective inhibitor of mammalian DNA polymerase. Curr. Org. Synth. 2011, 8, 134–144. [Google Scholar] [CrossRef]
- Altemoller, M.; Gehring, T.; Cudaj, J.; Podlech, J.; Goesmann, H.; Feldmann, C.; Rothenberger, A. Total synthesis of graphislactones A, C, D, and H, of ulocladol, and of the originally proposed and revised structures of graphislactones E and F. Eur. J. Org. Chem. 2009, 2009, 2130–2140. [Google Scholar] [CrossRef]
- Nealmongkol, P.; Tangdenpaisal, K.; Sitthimonchai, S.; Ruchirawat, S.; Thasana, N. Cu(I)-mediated lactone formation in subcritical water: A benign synthesis of benzopyranones and urolithins A-C. Tetrahedron 2013, 69, 9277–9283. [Google Scholar] [CrossRef]
- Pottie, I.R.; Nandaluru, P.R.; Bodwell, G.J. An inverse electron-demand diels-alder-based total synthesis of urolithin M7. Synlett 2011, 2011, 2245–2247. [Google Scholar]
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mao, Z.; Sun, W.; Fu, L.; Luo, H.; Lai, D.; Zhou, L. Natural Dibenzo-α-Pyrones and Their Bioactivities. Molecules 2014, 19, 5088-5108. https://doi.org/10.3390/molecules19045088
Mao Z, Sun W, Fu L, Luo H, Lai D, Zhou L. Natural Dibenzo-α-Pyrones and Their Bioactivities. Molecules. 2014; 19(4):5088-5108. https://doi.org/10.3390/molecules19045088
Chicago/Turabian StyleMao, Ziling, Weibo Sun, Linyun Fu, Haiyu Luo, Daowan Lai, and Ligang Zhou. 2014. "Natural Dibenzo-α-Pyrones and Their Bioactivities" Molecules 19, no. 4: 5088-5108. https://doi.org/10.3390/molecules19045088
APA StyleMao, Z., Sun, W., Fu, L., Luo, H., Lai, D., & Zhou, L. (2014). Natural Dibenzo-α-Pyrones and Their Bioactivities. Molecules, 19(4), 5088-5108. https://doi.org/10.3390/molecules19045088