Characterization of Natural Aryl Hydrocarbon Receptor Agonists from Cassia Seed and Rosemary

Abstract
:1. Introduction
2. Results and Discussion
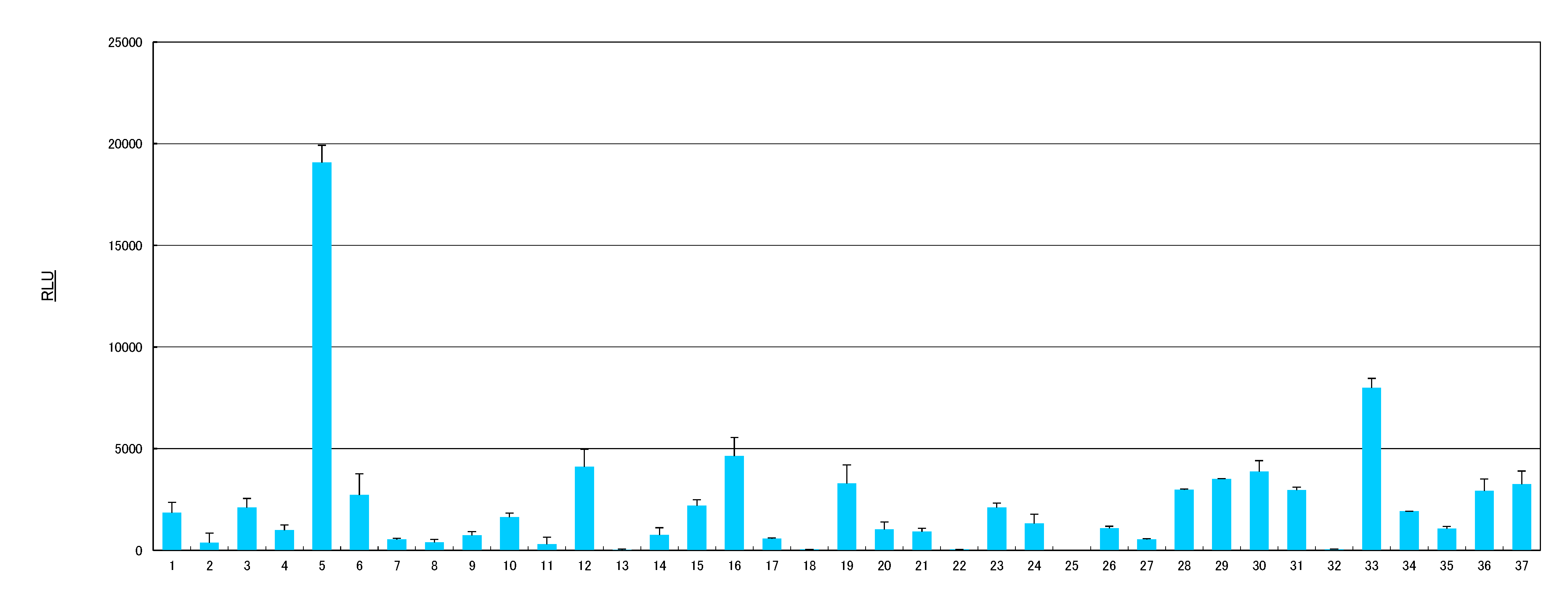
2.1. AhR Activities of Health Food Materials

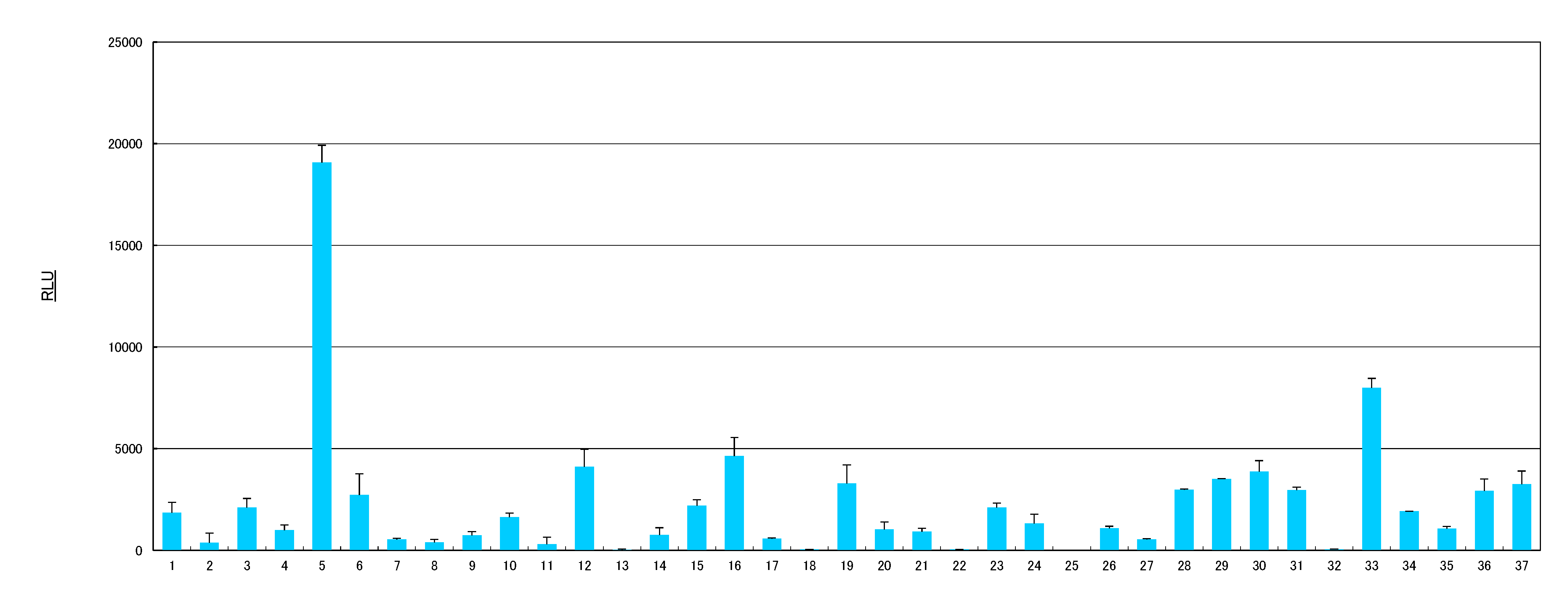{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Materials |
|---|---|
| 1 | Ashitaba (Japanese name) (Angelica keiskei) |
| 2 | Aloe (Aloe arborescens) |
| 3 | Amachazuru (Japanese name) (Gynostemma pentaphyllum) |
| 4 | Bitter melon (Momordica charantia) |
| 5 | Cassia seed (Cassia obtusifolia) |
| 6 | Celery (Apium graveolens) |
| 7 | Coix seed (Coix lacryma-jobi) |
| 8 | Cornus fruit (Cornus officinalis) |
| 9 | Crataegus fruit (Crataegus cuneata) |
| 10 | Echinacea (Echinacea purpurea) |
| 11 | Elder (Sambucus racemosa) |
| 12 | Eleutherococcus senticosus rhizome (Eleutherococcus senticosus) |
| 13 | Eucalyptus leaf (Eucalyptus globulus) |
| 14 | Eucommia bark (Eucommia ulmoides) |
| 15 | Fennel (Foeniculum vulgare) |
| 16 | Fenugreek (Trigonella foenum-graecum) |
| 17 | Field horsetail (Equisetum arvense) |
| 18 | Garcinia (Garcinia verrucosa) |
| 19 | Giant crape-myrtle (Lagerstroemia speciosa) |
| 20 | Ginger (Zingiber officinale) |
| 21 | Ginkgo (Ginkgo biloba) |
| 22 | Gymnema (Gymnema sylvestre) |
| 23 | Kaki persimmon (Diospyros kaki) |
| 24 | Lemon balm (Melissa officinalis) |
| 25 | Lemon grass (Cymbopogon citratus) |
| 26 | Linden (Tilia europaea) |
| 27 | Maca (Lepidium meyenii) |
| 28 | Mugwort (Artemisia indica) |
| 29 | Parsley (Petroselinum crispum) |
| 30 | Perilla herb (Perilla frutescens) |
| 31 | Plantago herb (Plantago asiatica) |
| 32 | Rabdosia herba (Rabdosia japonica) |
| 33 | Rosemary (Rosmarinus officinalis) |
| 34 | Sesame (Sesamum indicum) |
| 35 | Star anise (Illicium verum) |
| 36 | Sweet hydrangea leaf (Hydrangea macrophylla) |
| 37 | Yarrow (Achillea millefolium) |
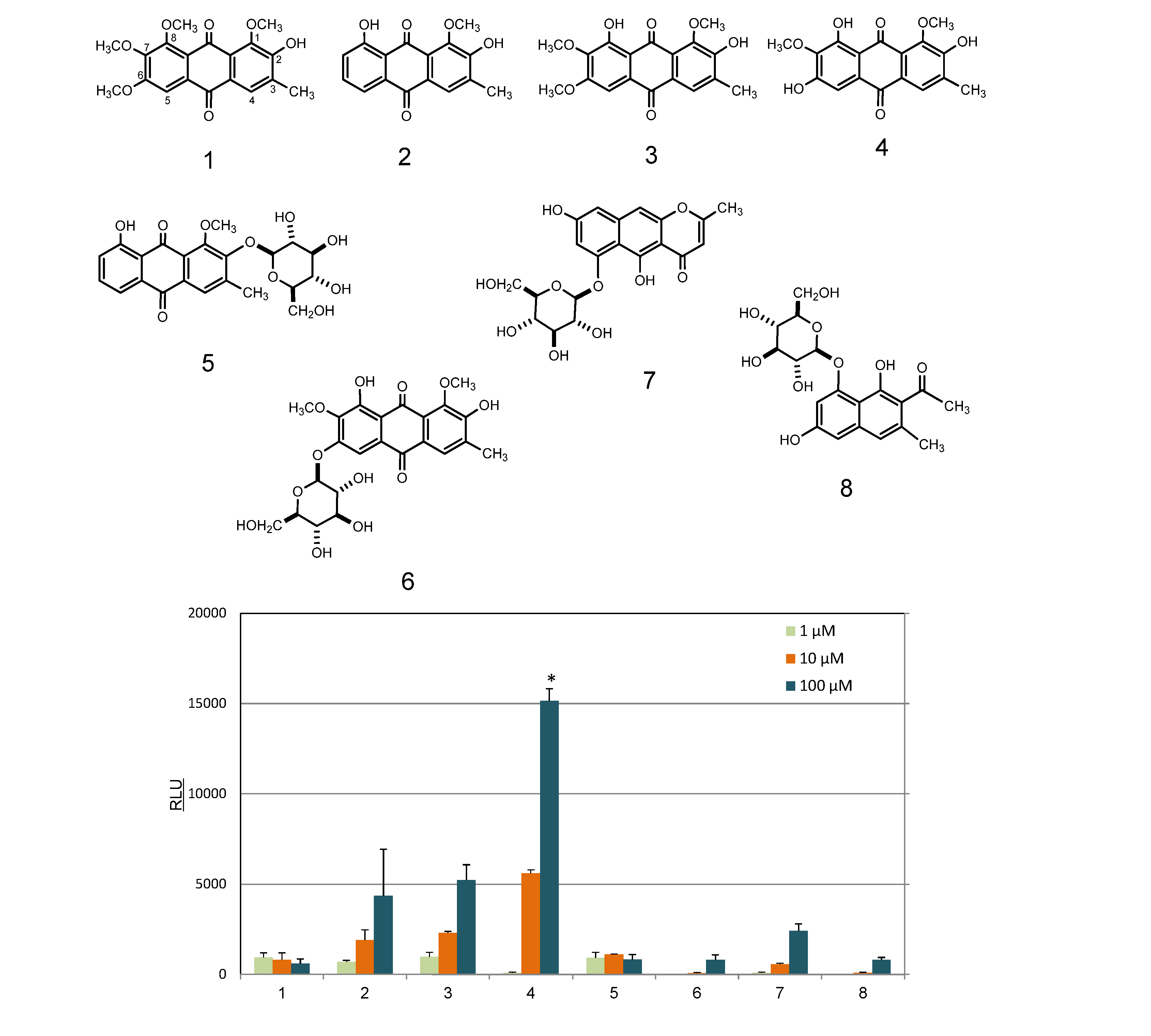
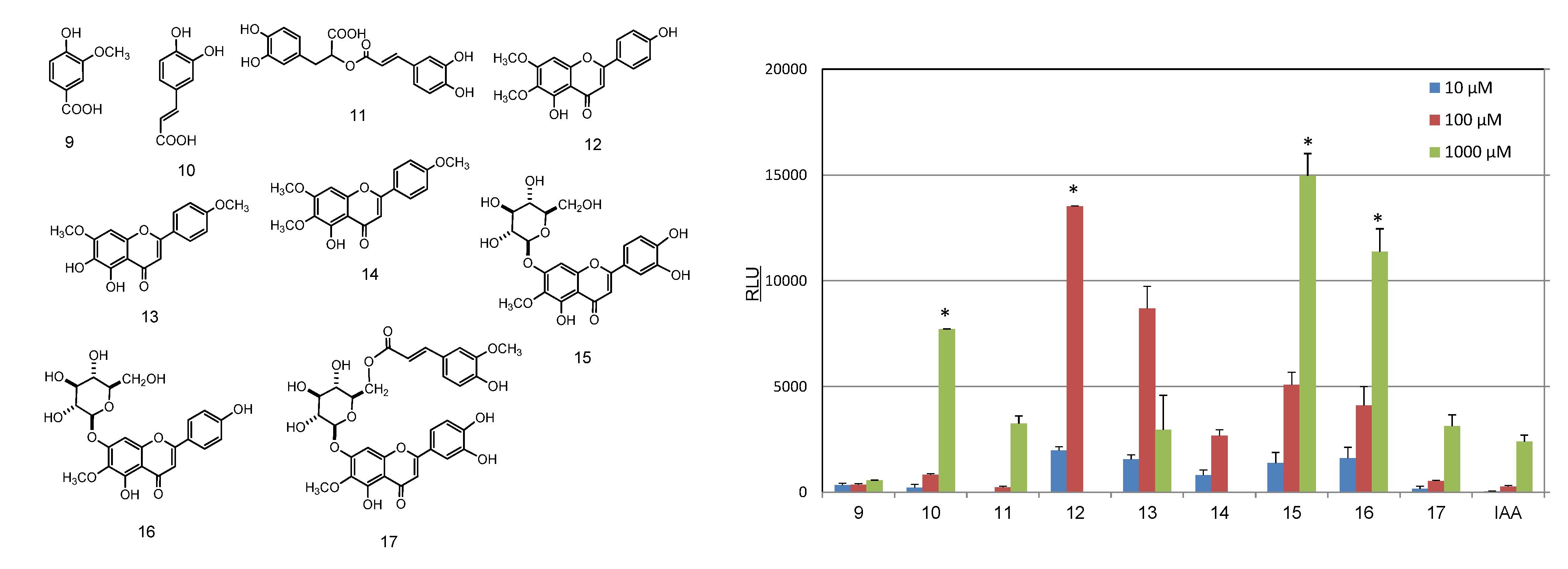
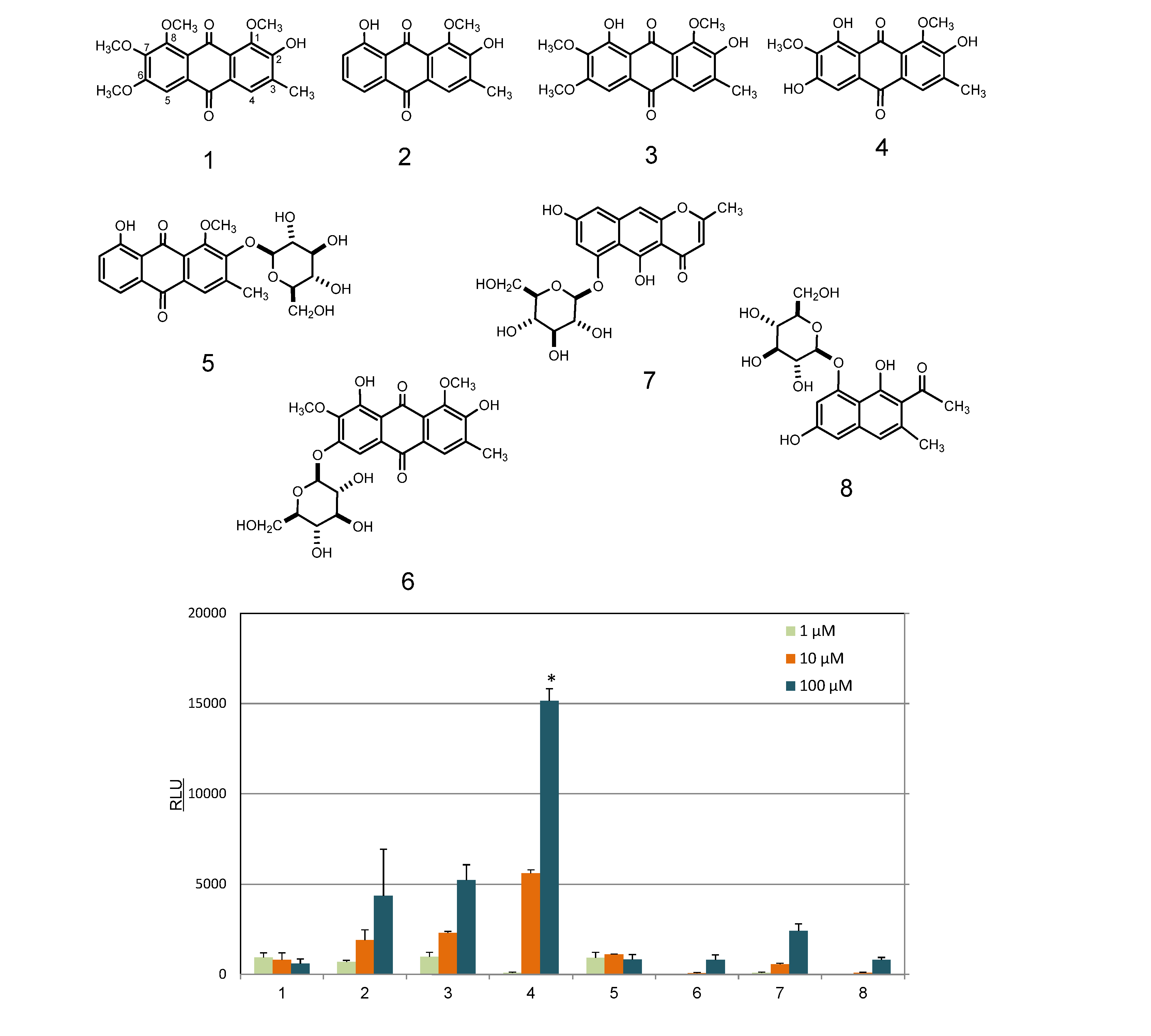
2.2. Identification and AhR Activity of Constituents

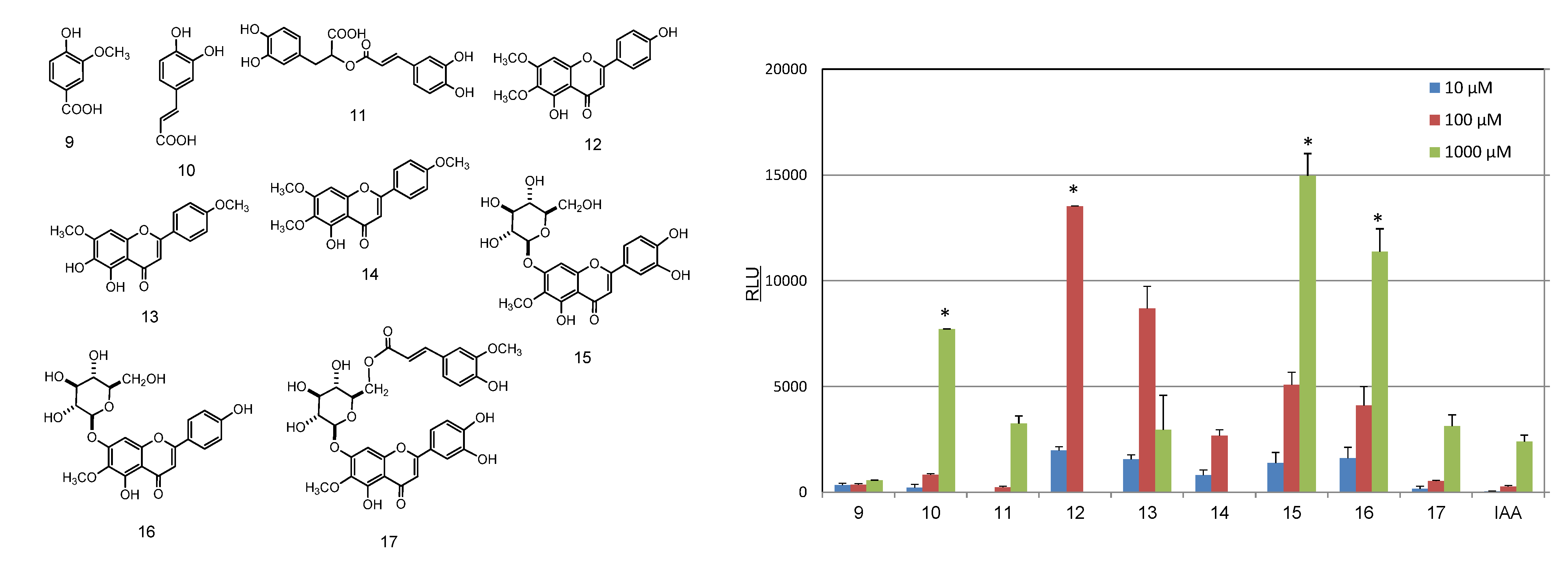
3. Experimental
3.1. General
3.2. Samples and Reagents
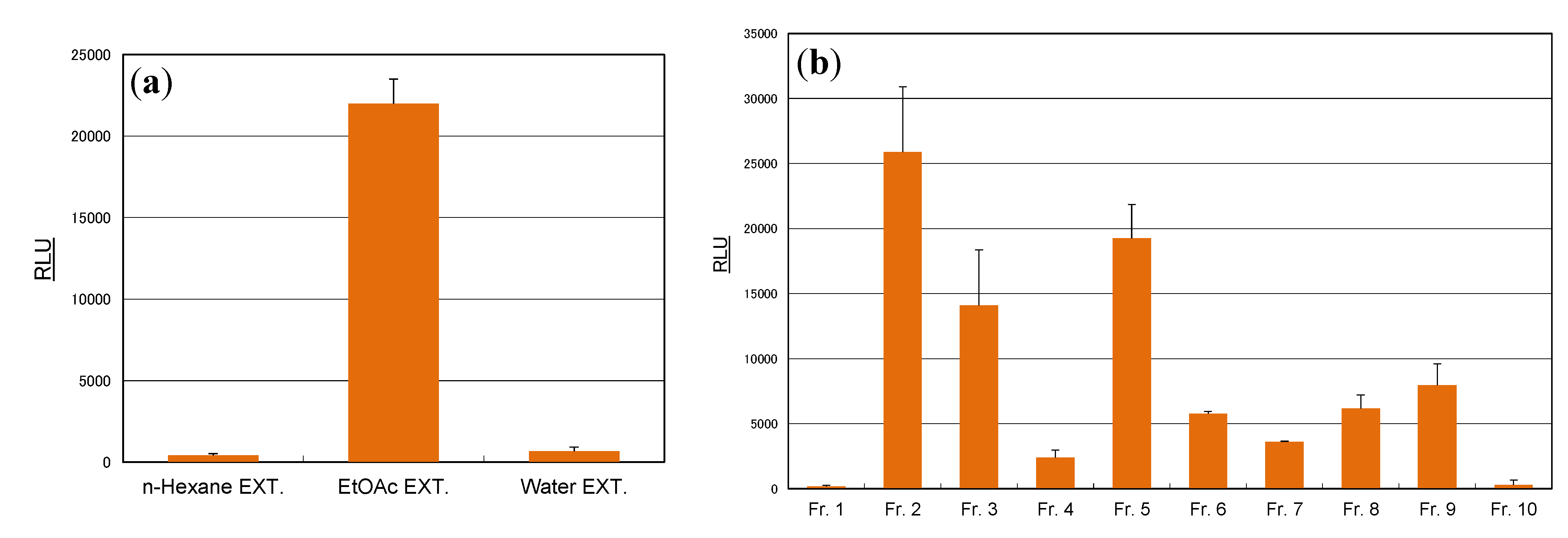
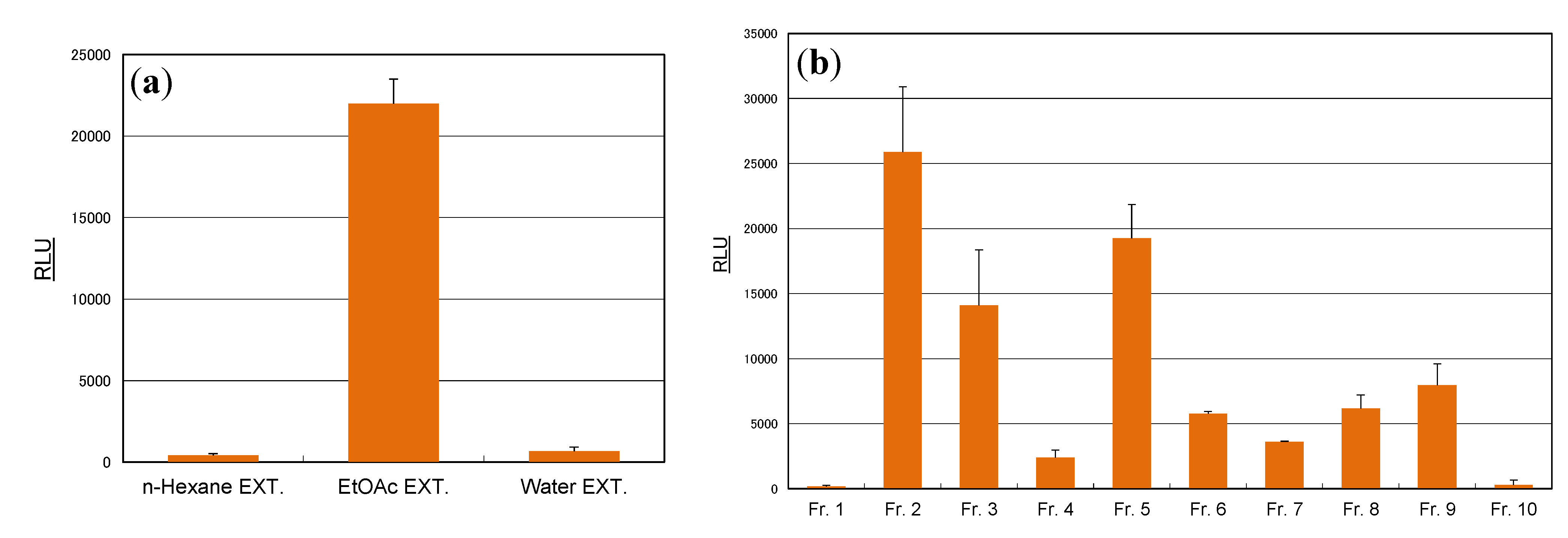
3.3. Extraction
3.4. Isolation of Compounds from Cassia Seeds
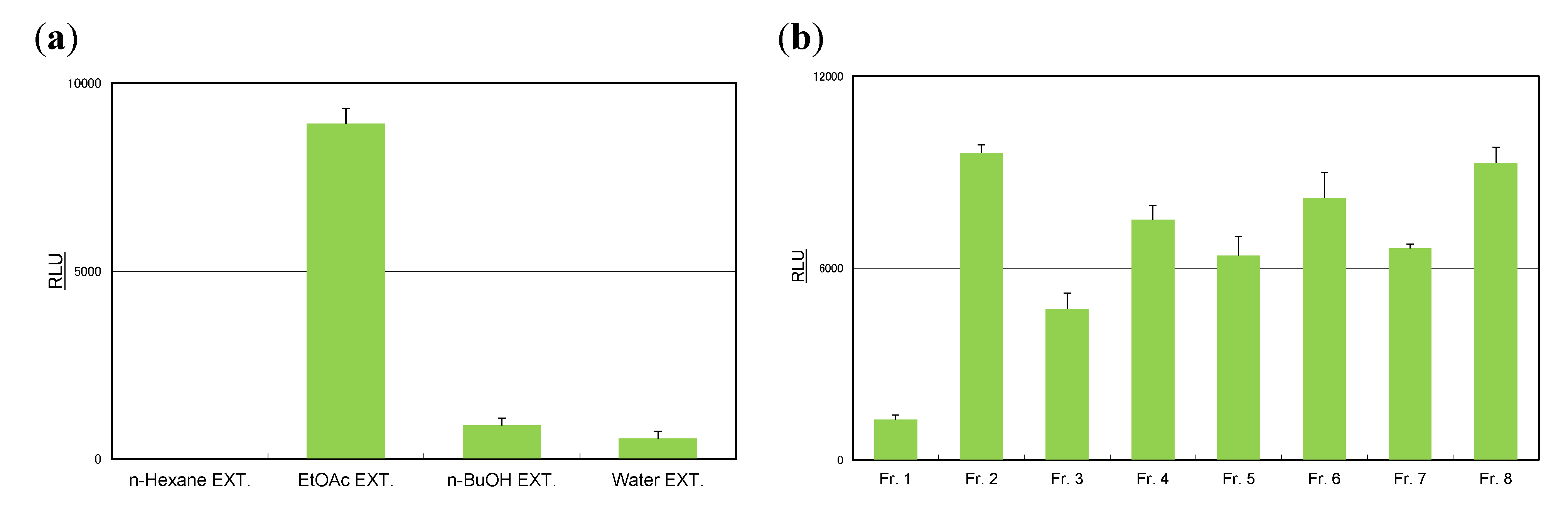
3.5. Isolation of Compounds from Rosemary
3.6. Estimation of AhR Ligand Activity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Machala, M.; Vondráček, J.; Bláha, L.; Ciganek, M.; Neča, J. Aryl hydrocarbon receptor-mediated activity of mutagenic polycyclic aromatic hydrocarbons determined using in vitro reporter gene assay. Mutat. Res. 2001, 31, 347–364. [Google Scholar]
- Fujii-Kuriyama, Y.; Mimura, J. Molecular mechanisms of AhR functions in the regulation of cytochrome P450 genes. Biochem. Biophys. Res. Commun. 2005, 338, 311–317. [Google Scholar] [CrossRef]
- Nebert, D.W.; Dalton, T.P. The role of cytochrome P450 enzymes in endogenous signalling pathways and environmental carcinogenesis. Nat. Rev. Cancer. 2006, 6, 947–960. [Google Scholar] [CrossRef]
- Amakura, Y.; Tsutsumi, T.; Sasaki, K.; Nakamura, M.; Yoshida, T.; Maitani, T. Influence of food polyphenols on aryl hydrocarbon receptor-signaling pathway estimated by in vitro bioassay. Phytochemistry 2008, 69, 3117–3130. [Google Scholar]
- Ashida, H.; Nishiumi, S.; Fukuda, I. An update on the dietary ligands of the AhR. Expert Opin. Drug Metab. Toxicol. 2008, 4, 1429–1447. [Google Scholar] [CrossRef]
- Wang, H.K.; Yeh, C.H.; Iwamoto, T.; Satsu, H.; Shimizu, M.; Totsuka, M. Dietary flavonoid naringenin induces regulatory T cells via an aryl hydrocarbon receptor mediated pathway. J. Agric. Food Chem. 2012, 60, 2171–2178. [Google Scholar]
- Tan, Z.; Chang, X.; Puga, A.; Xia, Y. Activation of mitogen-activated protein kinase (MAPKs) by aromatic hydrocarbons: role in the regulation of aryl hydrocarbon receptor (AHR) function. Biochem. Pharmacol. 2002, 64, 771–780. [Google Scholar] [CrossRef]
- Kawajiri, K.; Kobayashi, Y.; Ohtake, F.; Ikuta, T.; Matsushima, Y.; Mimura, J.; Pettersson, S.; Pollenz, R.S.; Sakaki, T.; Hirokawa, T.; et al. Aryl hydrocarbon receptor suppresses intestinal carcinogenesis in ApcMin/+ mice with natural ligands. Proc. Natl. Acad. Sci. USA 2009, 106, 13481–13486. [Google Scholar] [CrossRef]
- Kimura, A.; Naka, T.; Nohara, K.; Fujii-Kuriyama, Y.; Kishimoto, T. Aryl hydrocarbon receptor regulates Stat1 activation and participates in the development of Th17 cells. Proc. Natl. Acad. Sci. USA 2008, 105, 9721–9726. [Google Scholar]
- Quintana, F.J.; Basso, A.S.; Iglesias, A.H.; Korn, T.; Farez, M.F.; Bettelli, E.; Caccamo, M.; Oukka, M.; Weiner, H.L. Control of Treg and TH17 cell differentiation by the aryl hydrocarbon receptor. Nature 2008, 453, 65–71. [Google Scholar] [CrossRef]
- Veldhoen, M.; Hirota, K.; Westendorf, A.M.; Buer, J.; Dumoutier, L.; Renauld, J.; Stockinger, B. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 2008, 453, 106–110. [Google Scholar] [CrossRef]
- Kiss, E.A.; Vonarbourg, C.; Kopfmann, S.; Hobeika, E.; Finke, D.; Esser, C.; Diefenbach, A. Natural aryl hydrocarbon receptor ligands control organogenesis of intestinal lymphoid follicles. Science 2011, 334, 1561–1565. [Google Scholar] [CrossRef]
- Hu, W.; Zhao, J.; Pei, G. Activation of aryl hydrocarbon receptor by tranilast, an anti-allergy drug, promotes miR-302 expression and reprogramming. J. Biol. Chem. 2013, 288, 22972–22984. [Google Scholar] [CrossRef]
- Tsutsumi, T.; Amakura, Y.; Nakamura, M.; Brown, D.J.; Clark, G.C.; Sasaki, K.; Toyoda, M.; Maitani, T. Validation of the CALUX bioassay for the screening of PCDD/Fs and dioxin-like PCBs in retail fish. Analyst 2003, 128, 486–492. [Google Scholar] [CrossRef]
- Overmeire, I.V.; Clark, G.C.; Brown, D.J.; Chu, M.D.; Cooke, W.M.; Denison, M.S.; Baeyens, W.; Srebrnik, S.; Goeyens, L. Analysis of Ah receptor pathway activation by brominated flame retardants. Environ. Sci. Policy 2001, 4, 345–357. [Google Scholar] [CrossRef]
- Han, D.; Nagy, S.R.; Denison, M.S. Comparison of recombinant cell bioassays for the detection of Ah receptor agonists. BioFactors 2004, 20, 11–22. [Google Scholar] [CrossRef]
- Moghaddam, G.; Ebrahimi, S.A.; Rahbar-Roshandel, N.; Foroumadi, A. Antiproliferative activity of flavonoids: Influence of the sequential methoxylation state of the flavonoid structure. Phytother. Res. 2012, 26, 1023–1028. [Google Scholar] [CrossRef]
- Agarwal, O.P. The anti-inflammatory action of nepitrin, a flavonoid. Agents Actions 1982, 12, 298–302. [Google Scholar] [CrossRef]
- Nugroho, A.; Kim, M.H.; Choi, J.; Beak, N.I.; Park, H.J. In vivo sedative and gastroprotective activities of Salvia plebeian extract and its composition of polyphenols. Arch. Pharm. Res. 2012, 35, 1403–1411. [Google Scholar]
- Li, X.C.; Dunbar, D.C.; Elsohly, H.N.; Jacob, M.R.; Nimrod, A.C.; Walker, L.A.; Clark, A.M. A new naphthopyrone derivative from Cassia quinquangulata and structural revision of quinquangulin and its glycosides. J. Nat. Prod. 2001, 64, 1153–1156. [Google Scholar] [CrossRef]
- Yun-Choi, H.S.; Kim, J.H. Potential inhibitors of platelet aggregation from plant sources, V. Anthraquinones from seeds of Cassia obtusifolia and related compounds. J. Nat. Prod. 1990, 53, 630–633. [Google Scholar] [CrossRef]
- Begum, S.; Wahab, A.; Siddiqui, B.S.; Qamar, F. Nematicidal constituents of the aerial parts of Lantana camara. J. Nat. Prod. 2000, 63, 765–767. [Google Scholar] [CrossRef]
- Ly, T.N.; Shimoyamada, M.; Yamauchi, R. Isolation and characterization of rosmarinic acid oligomers in Celastrus hindsii Benth leaves and their antioxidative activity. J. Agric. Food Chem. 2006, 54, 3786–3793. [Google Scholar] [CrossRef]
- Wang, M.; Li, J.; Rangarajan, M.; Shao, Y.; LaVoie, E.; Huang, T.C.; Ho, C.T. Antioxidative phenolic compounds from sage (Salvia officinalis). J. Agric. Food Chem. 1998, 46, 4869–4873. [Google Scholar] [CrossRef]
- Pèrez-Fons, L.; Garzón, M.T.; Micol, V. Relationship between the antioxidant capacity and effect of rosemary (Rosmarinus officinalis L.) polyphenols on membrane phospholipid order. J. Agric. Food Chem. 2010, 58, 161–171. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compound 11 are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Amakura, Y.; Yoshimura, M.; Takaoka, M.; Toda, H.; Tsutsumi, T.; Matsuda, R.; Teshima, R.; Nakamura, M.; Handa, H.; Yoshida, T. Characterization of Natural Aryl Hydrocarbon Receptor Agonists from Cassia Seed and Rosemary. Molecules 2014, 19, 4956-4966. https://doi.org/10.3390/molecules19044956
Amakura Y, Yoshimura M, Takaoka M, Toda H, Tsutsumi T, Matsuda R, Teshima R, Nakamura M, Handa H, Yoshida T. Characterization of Natural Aryl Hydrocarbon Receptor Agonists from Cassia Seed and Rosemary. Molecules. 2014; 19(4):4956-4966. https://doi.org/10.3390/molecules19044956
Chicago/Turabian StyleAmakura, Yoshiaki, Morio Yoshimura, Masashi Takaoka, Haruka Toda, Tomoaki Tsutsumi, Rieko Matsuda, Reiko Teshima, Masafumi Nakamura, Hiroshi Handa, and Takashi Yoshida. 2014. "Characterization of Natural Aryl Hydrocarbon Receptor Agonists from Cassia Seed and Rosemary" Molecules 19, no. 4: 4956-4966. https://doi.org/10.3390/molecules19044956
APA StyleAmakura, Y., Yoshimura, M., Takaoka, M., Toda, H., Tsutsumi, T., Matsuda, R., Teshima, R., Nakamura, M., Handa, H., & Yoshida, T. (2014). Characterization of Natural Aryl Hydrocarbon Receptor Agonists from Cassia Seed and Rosemary. Molecules, 19(4), 4956-4966. https://doi.org/10.3390/molecules19044956