Anti-Lung Cancer Activity through Enhancement of Immunomodulation and Induction of Cell Apoptosis of Total Triterpenes Extracted from Ganoderma luncidum (Leyss. ex Fr.) Karst.

Abstract
:1. Introduction
2. Results
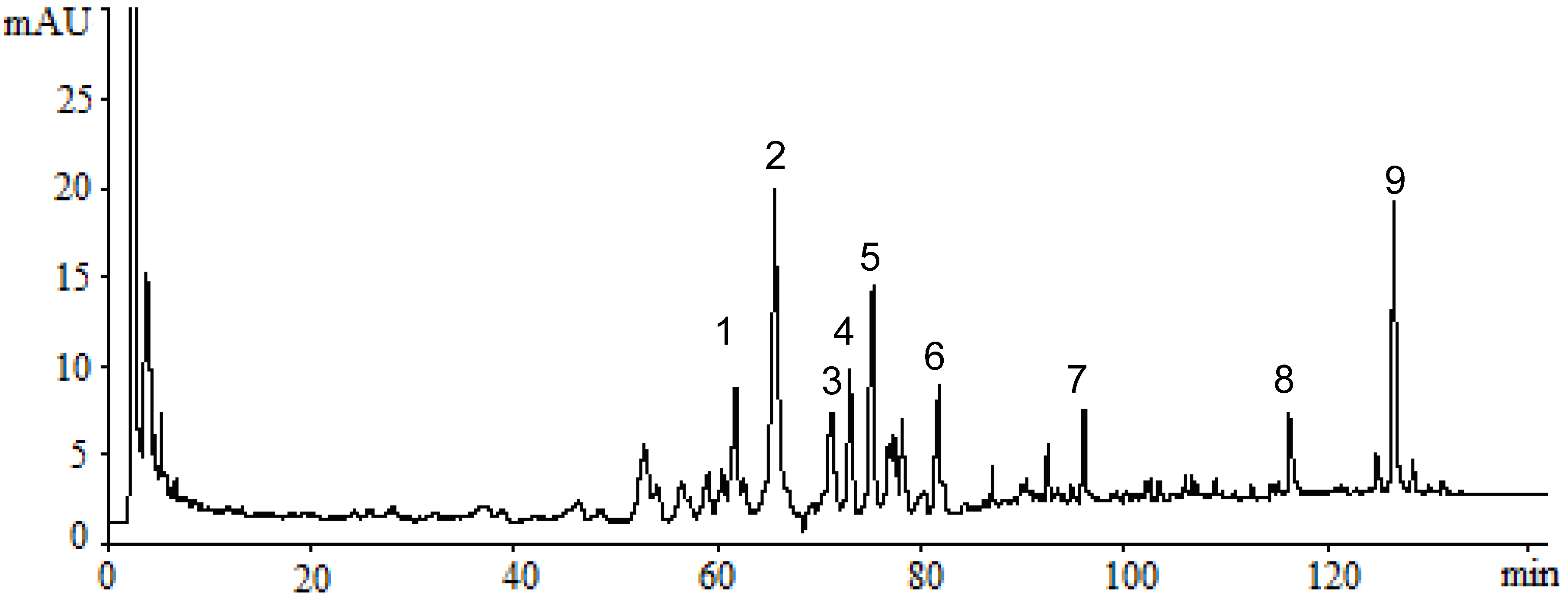
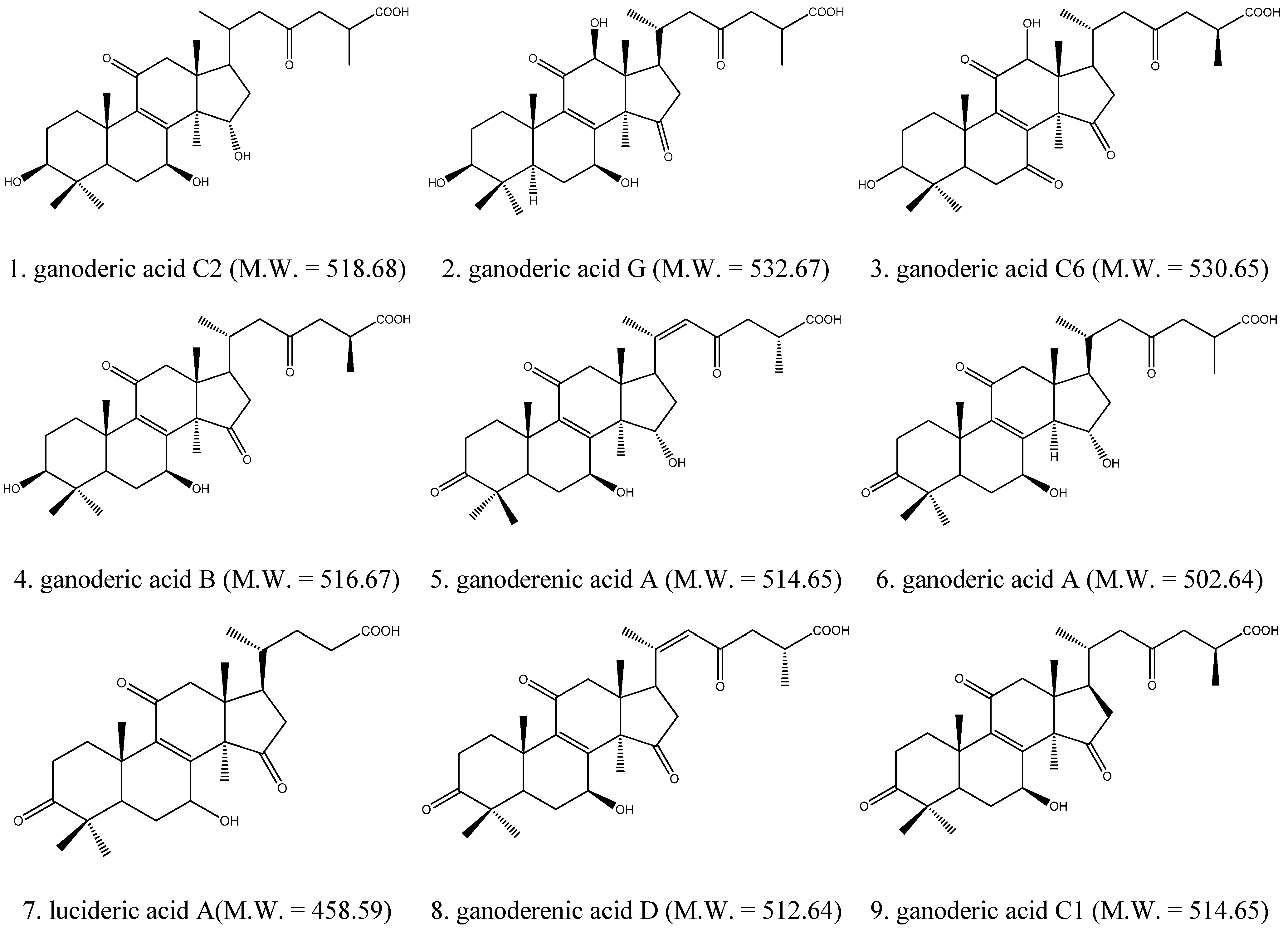
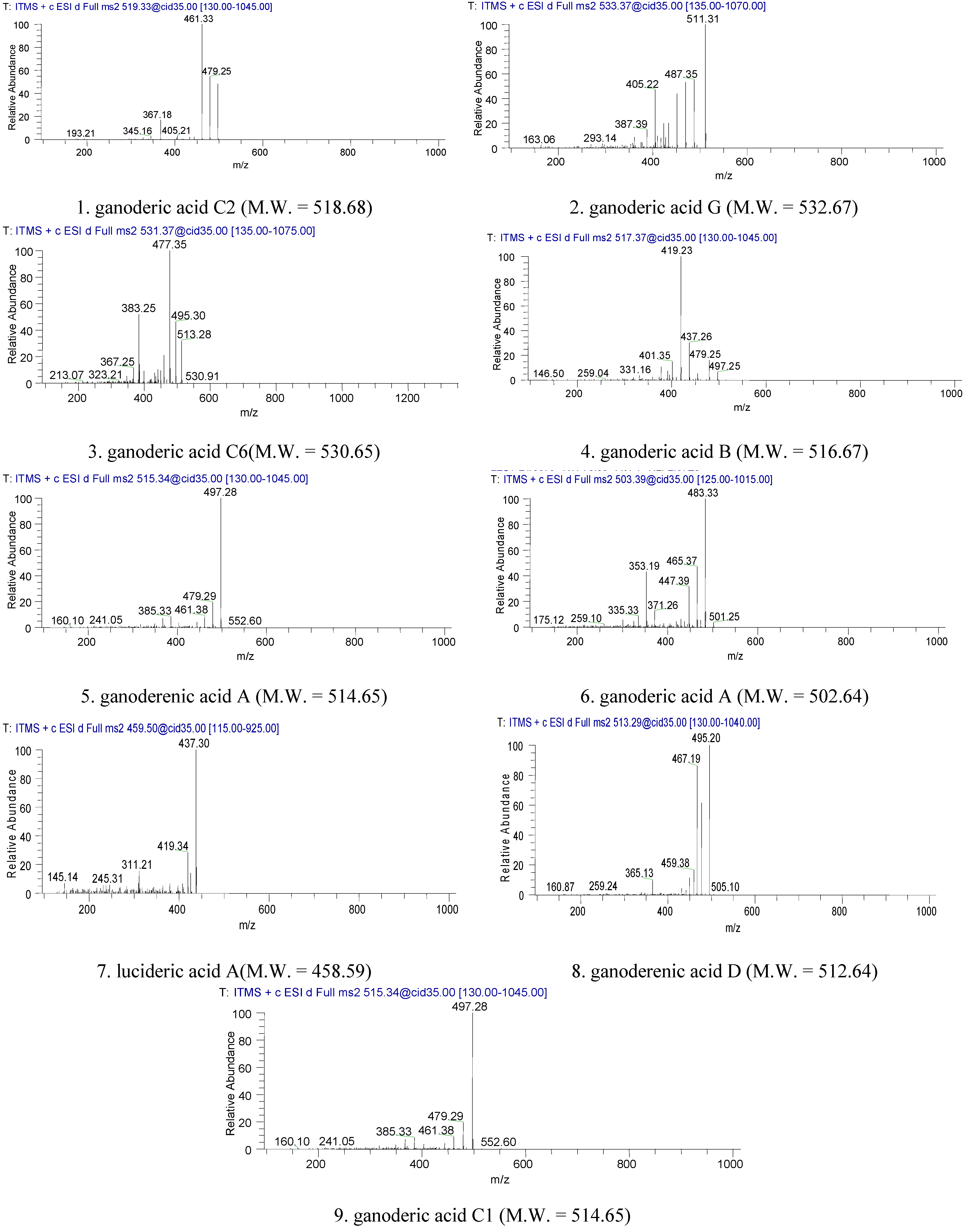
2.1. Identification of Triterpenes by HPLC-DAD and LC/MS Analysis
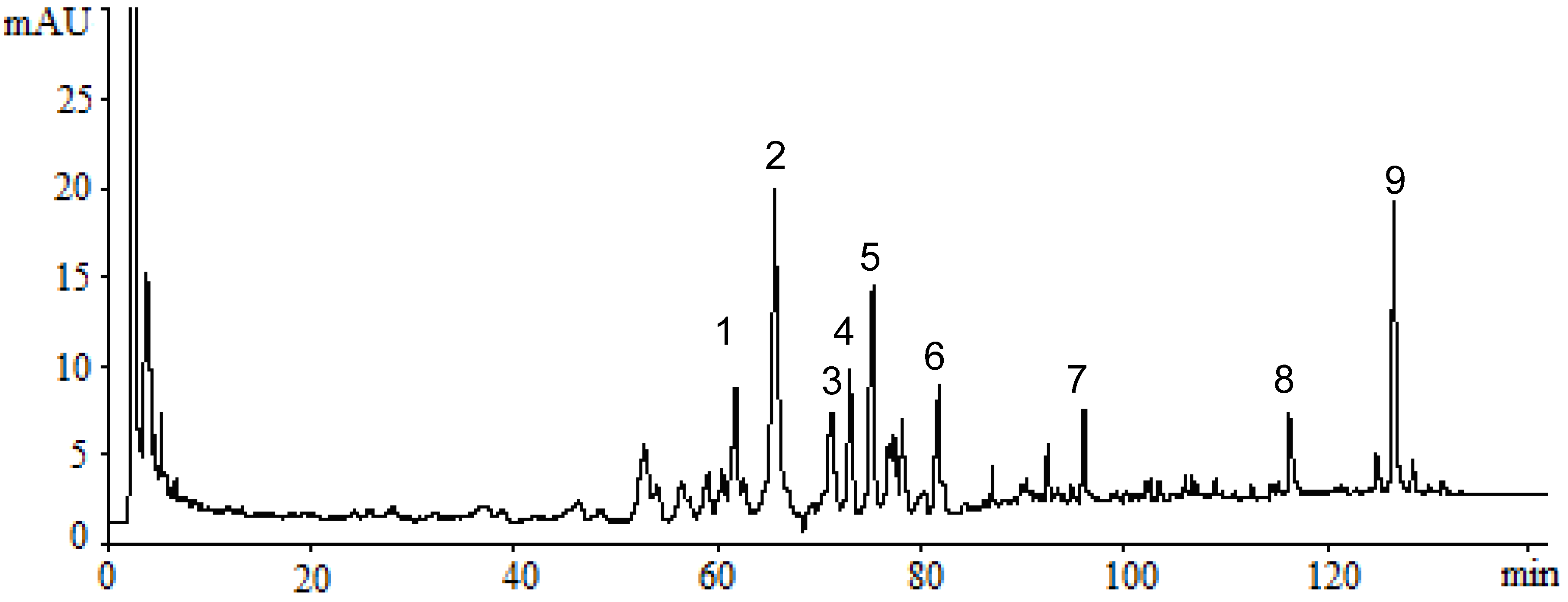
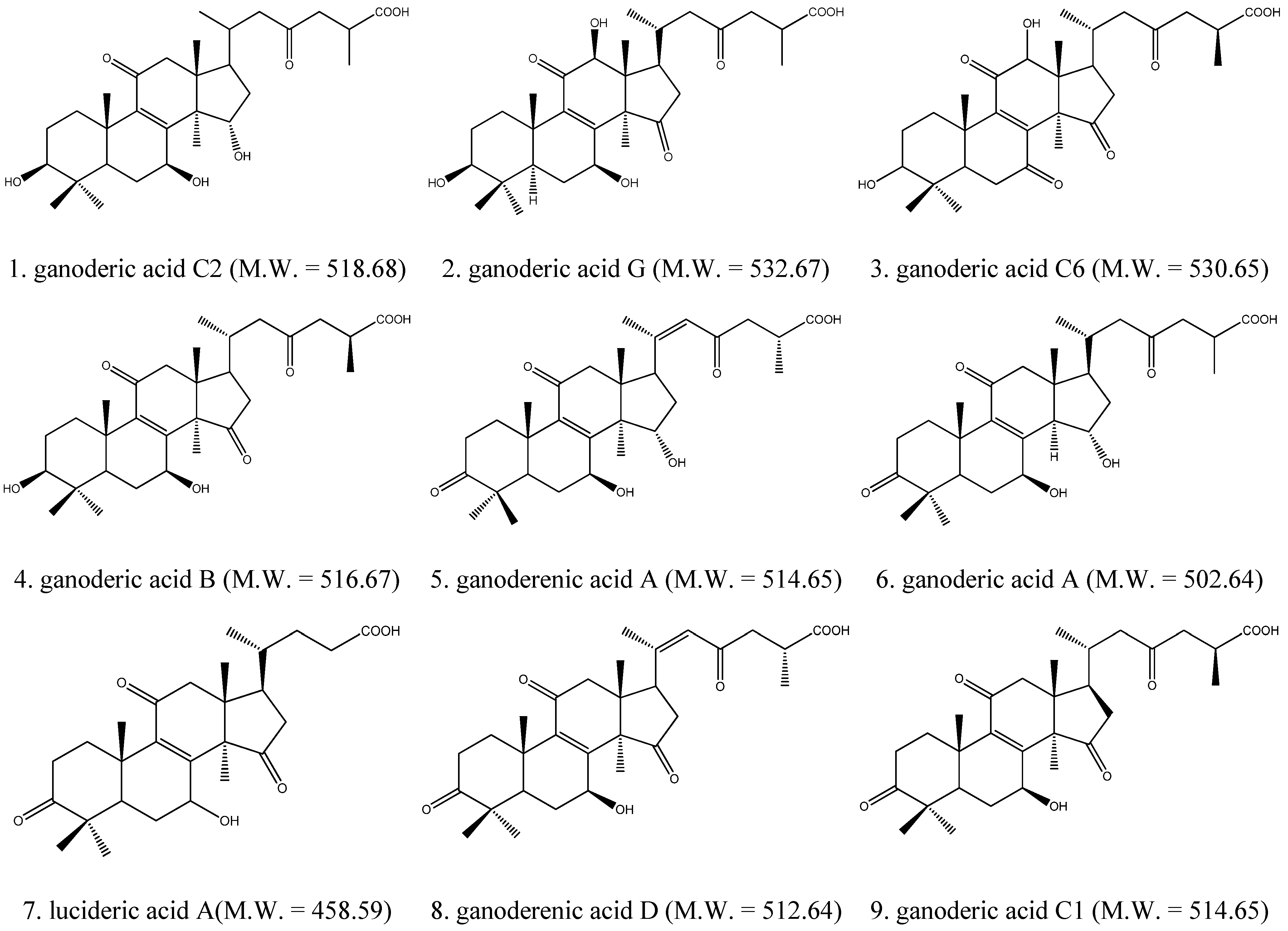
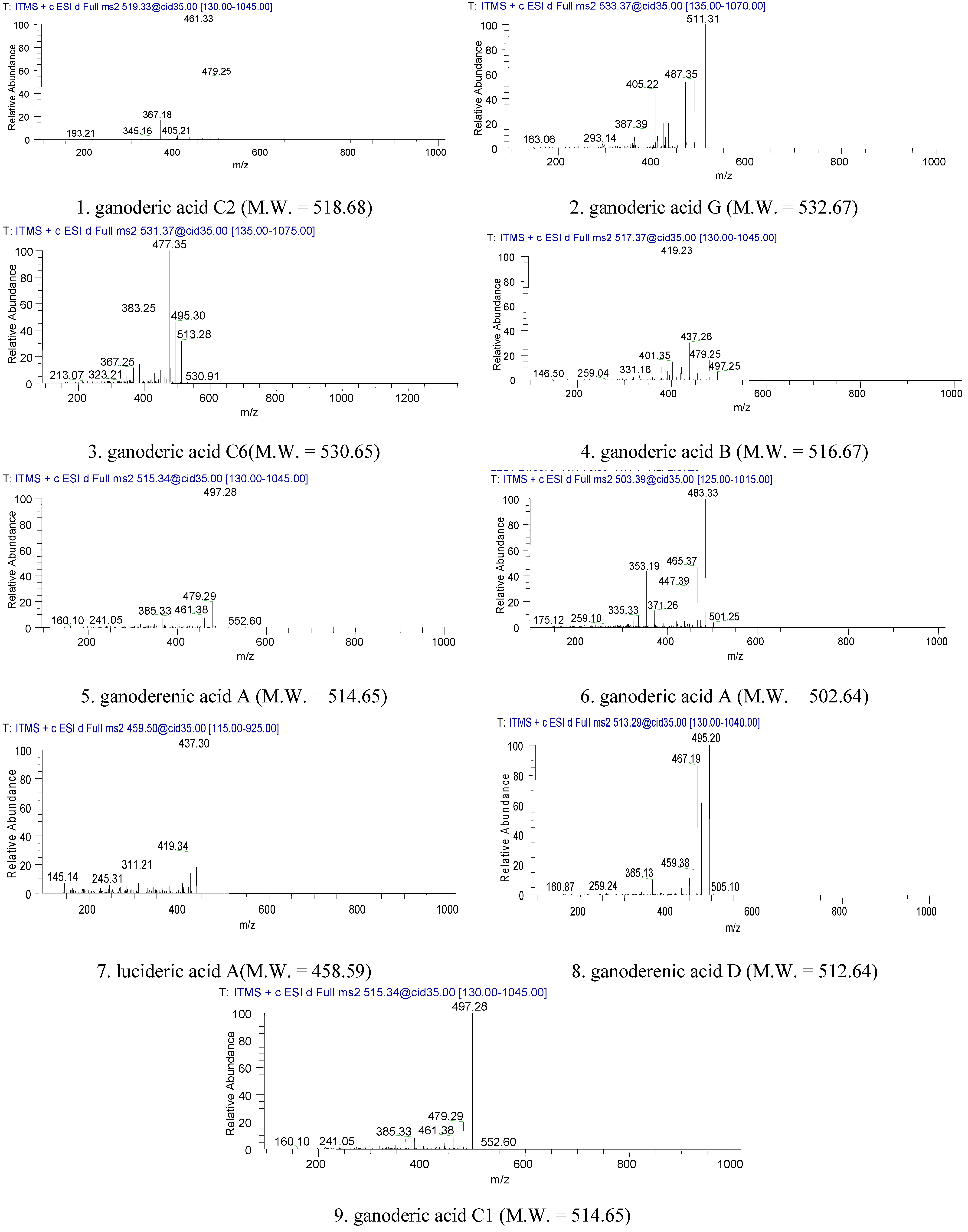
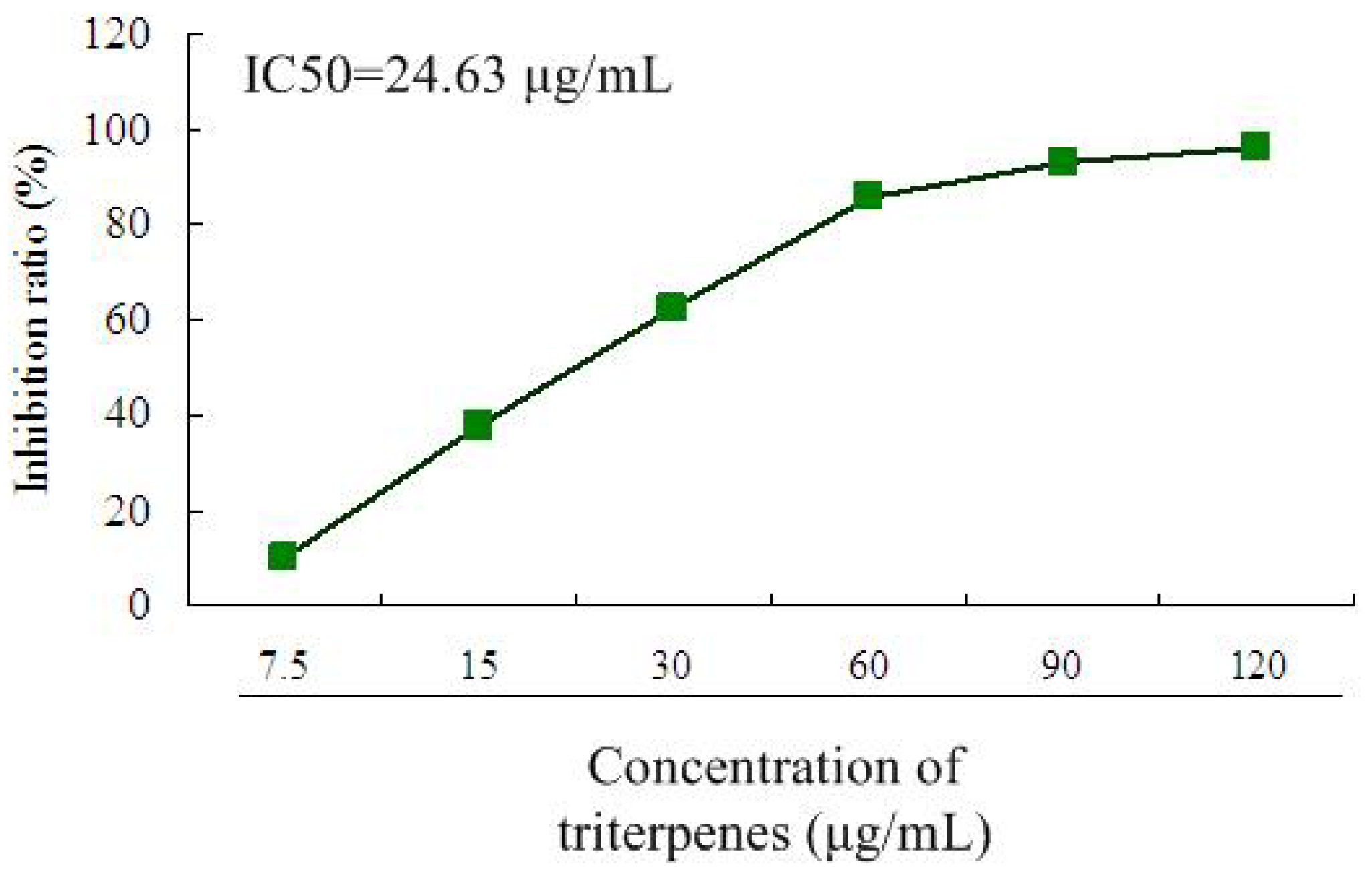
2.2. Inhibitory Effect of Triterpenes on A549 Cell Proliferation
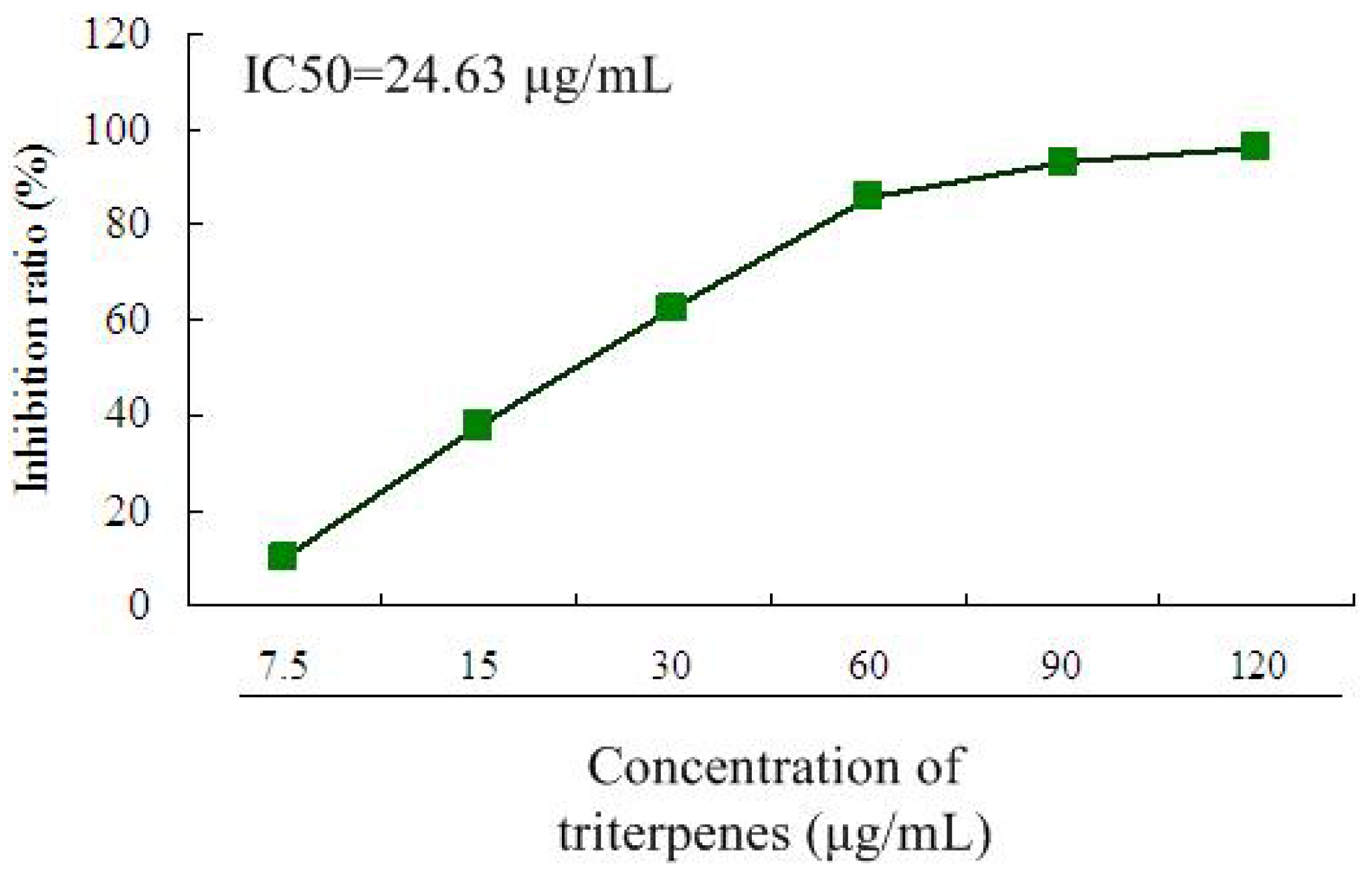
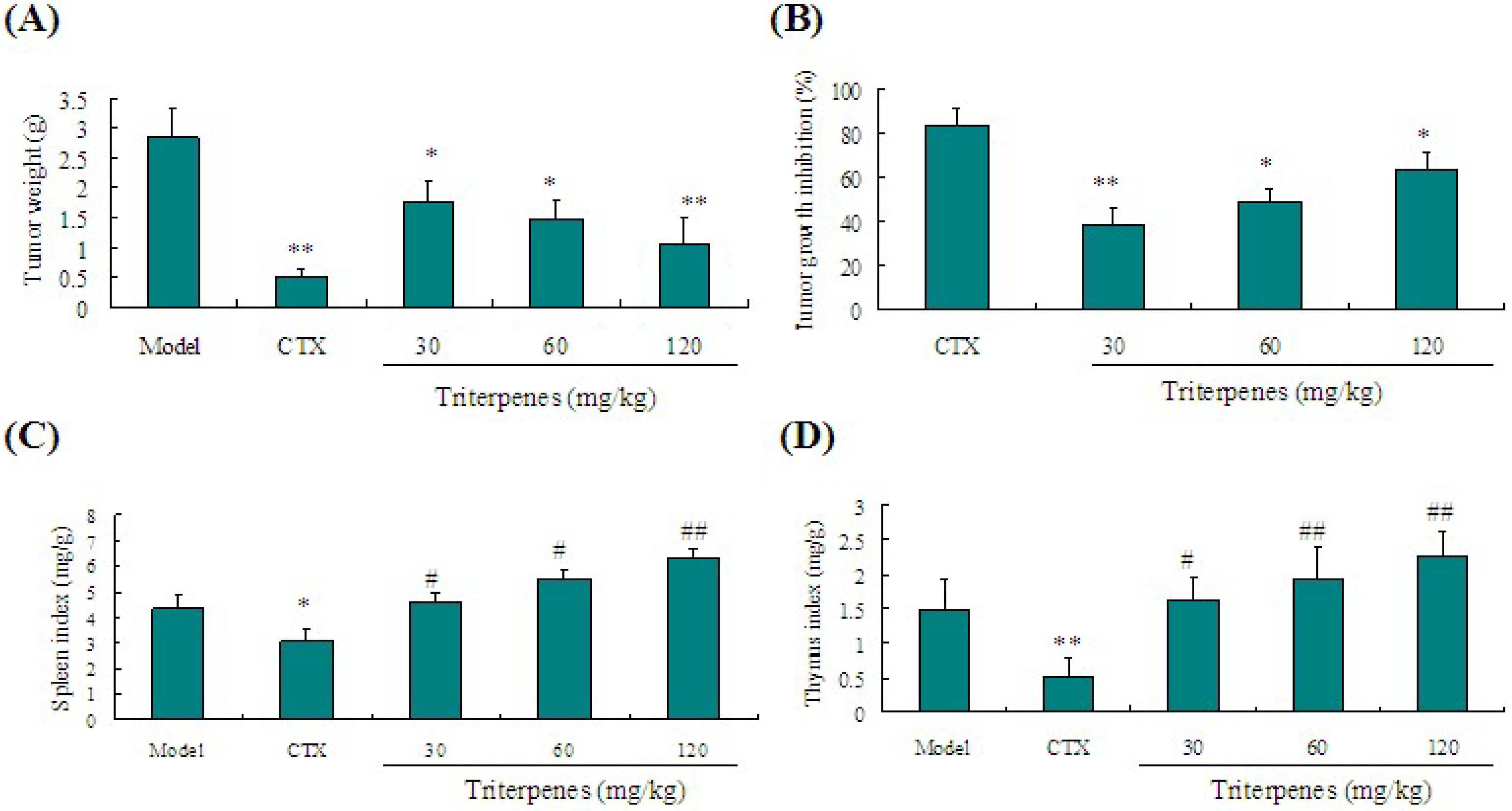
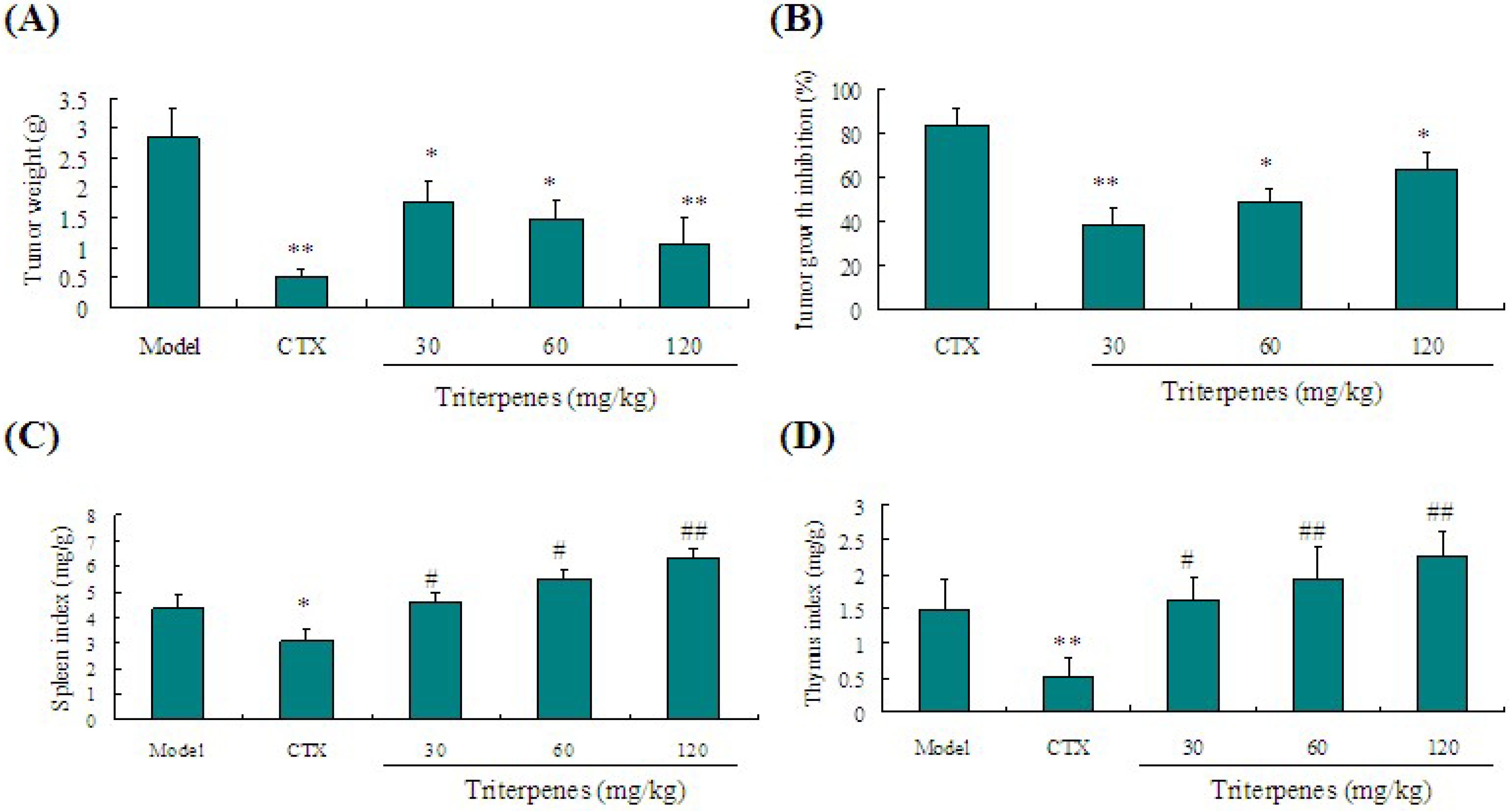
2.3. Anti-Tumor Activity of Triterpenes in Lewis Tumor-Bearing Mice
2.4. Immunostimulation Activity of Triterpenes on Spleen and Thymus Indexes in Lewis Tumor-Bearing Mice
2.5. Regulation of Triterpenes on IL-6 and TNF-α Levels in Lewis Tumor-Bearing Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Concentration of TNF-α (pg/mL) | Concentration of IL-6 (pg/mL) |
|---|---|---|
| Model | 42.07 ± 0.86 # | 34.53 ± 7.89 # |
| CTX (20 mg/kg) | 31.72 ± 7.87 * | 20.60 ± 1.60 * |
| Triterpenes (30 mg/kg) | 46.84 ± 4.30 # | 38.50 ± 7.86 # |
| Triterpenes (60 mg/kg) | 50.44 ± 3.2 *,# | 42.78 ± 5.64 *,# |
| Triterpenes (120 mg/kg) | 56.77 ± 6.32 *,# | 48.59 ± 3.45 *,# |
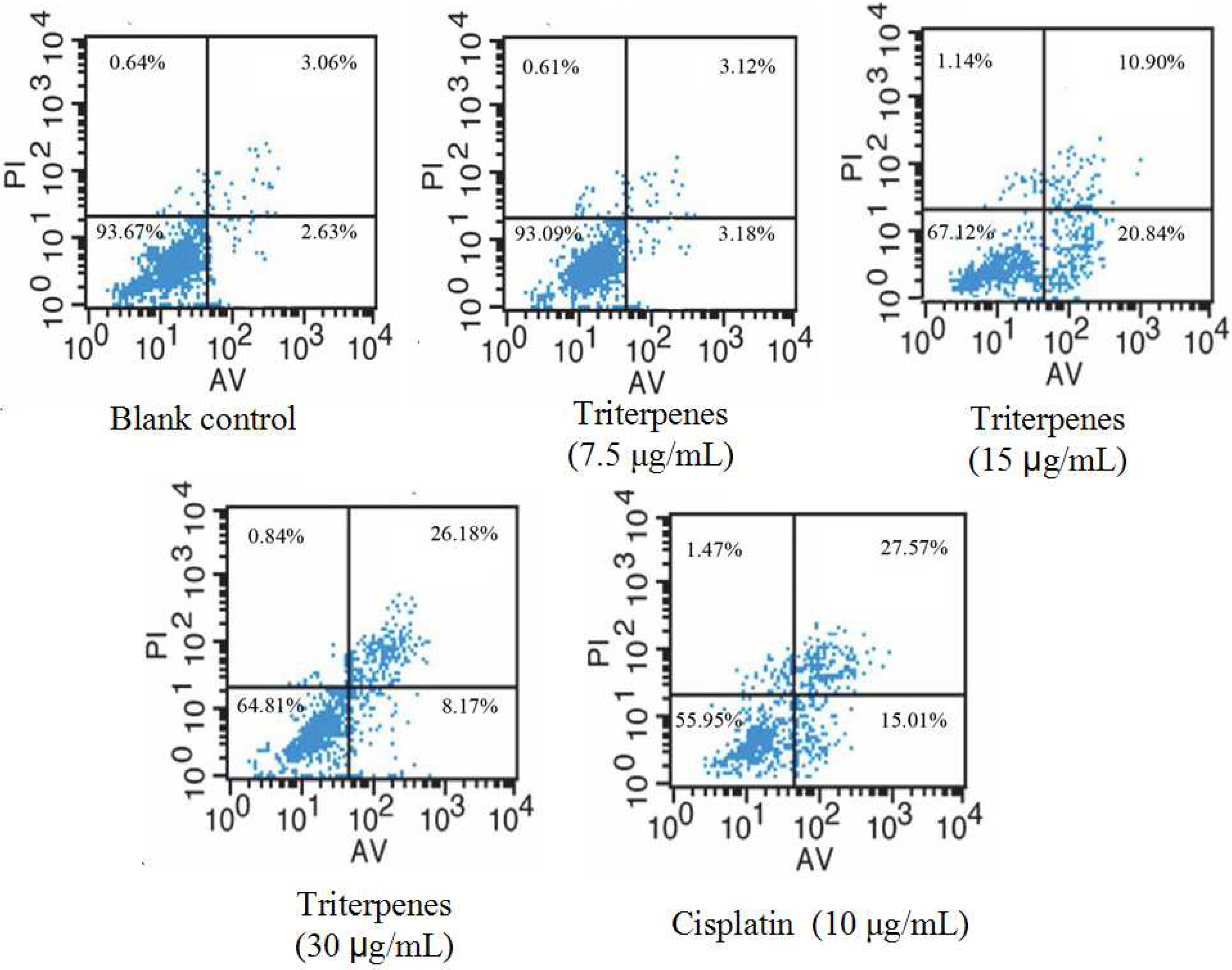
2.6. Induction of Triterpenes on Apoptosis of A549 Cells
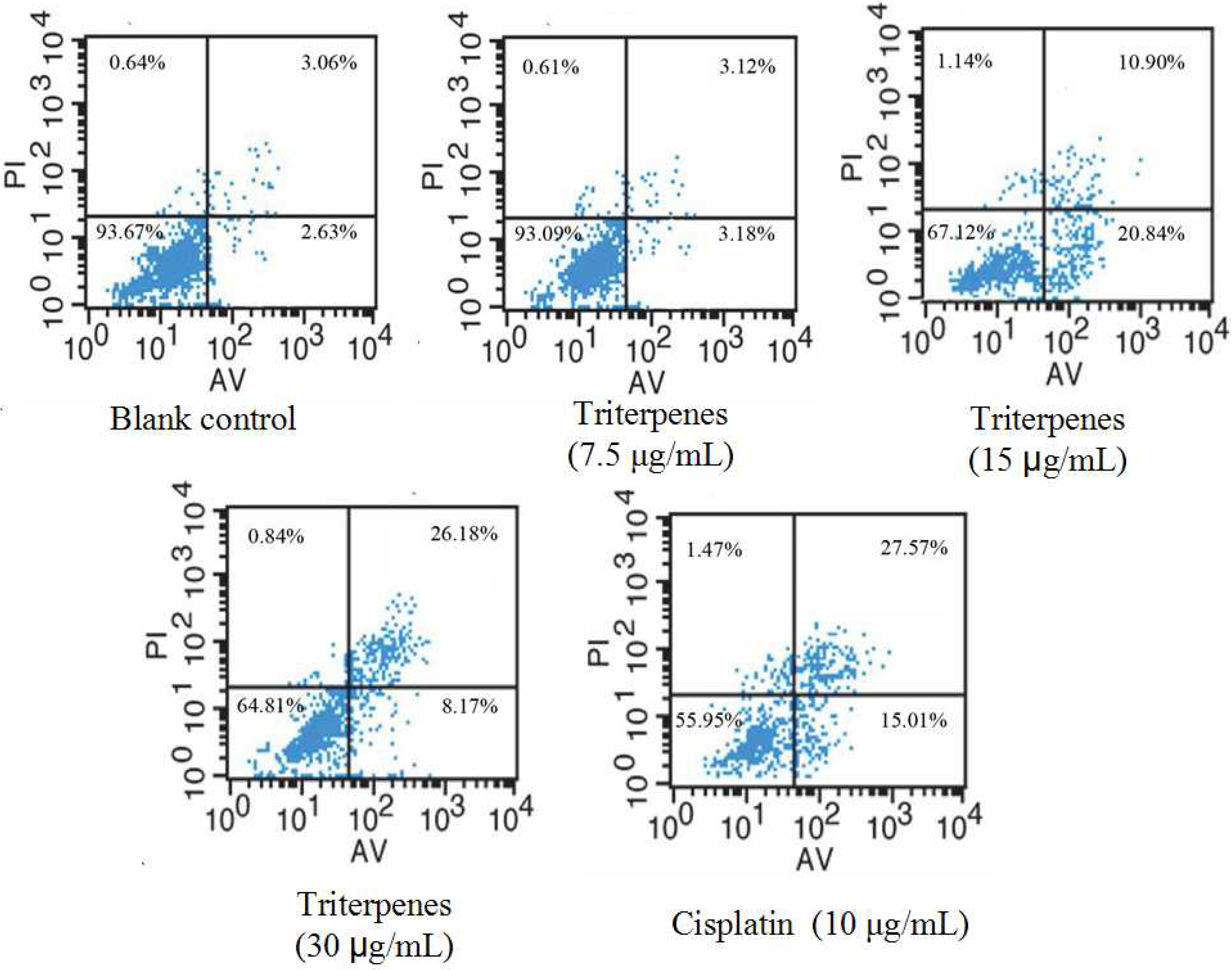
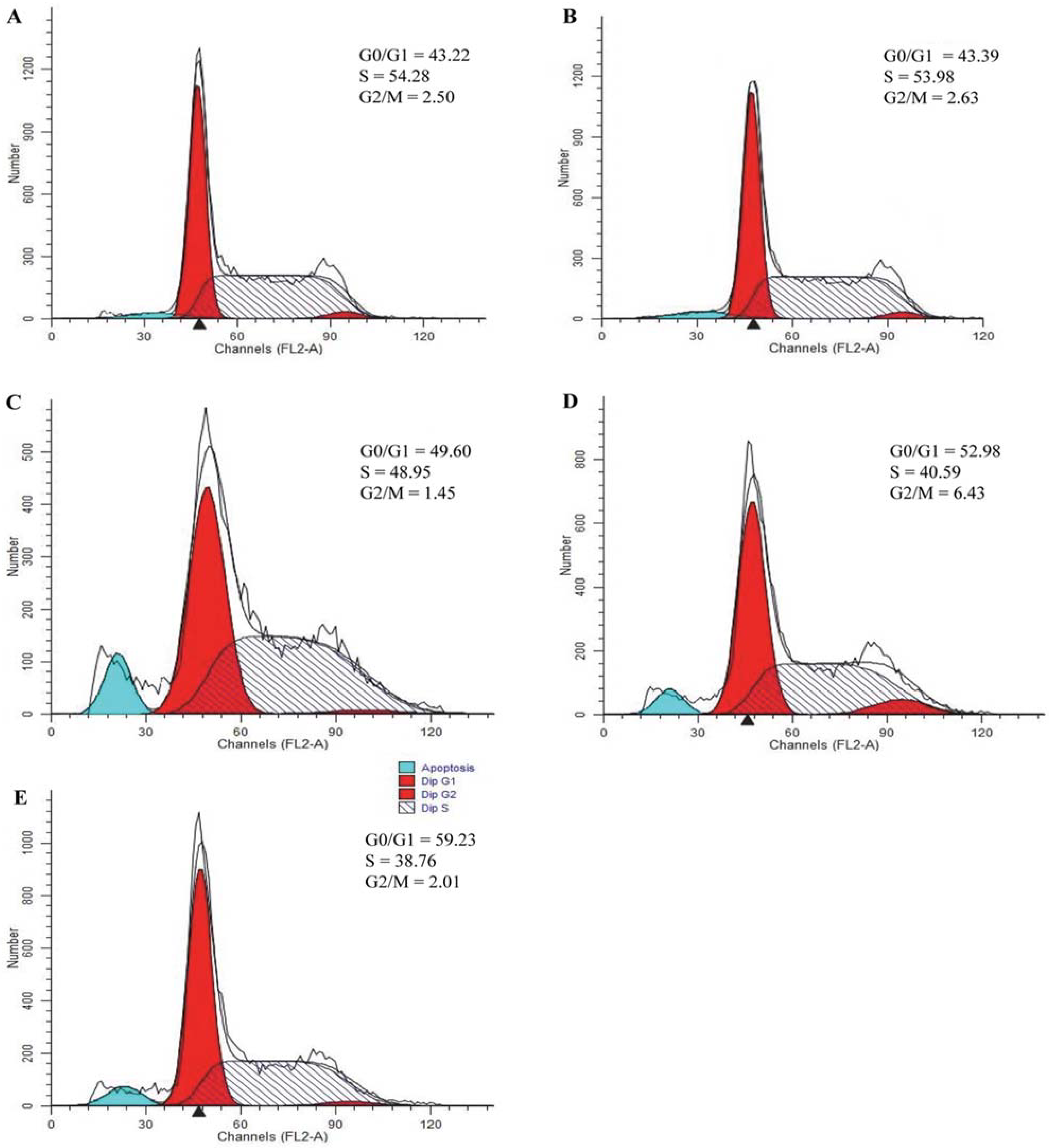
2.7. Effect of Triterpenes on Cell Cycle Arrest of A549 Cells

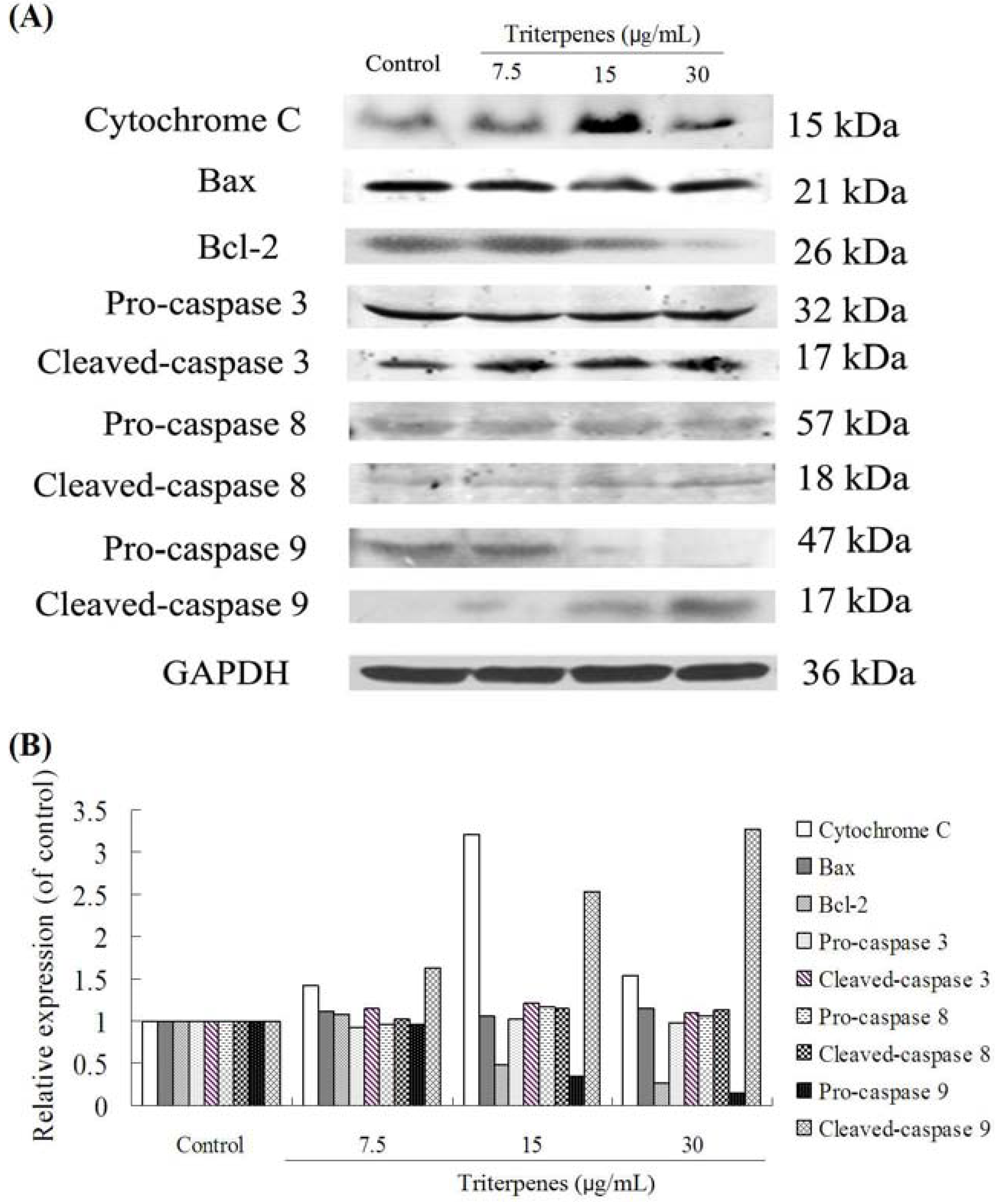
2.8. Apoptosis-Related Protein Expressions
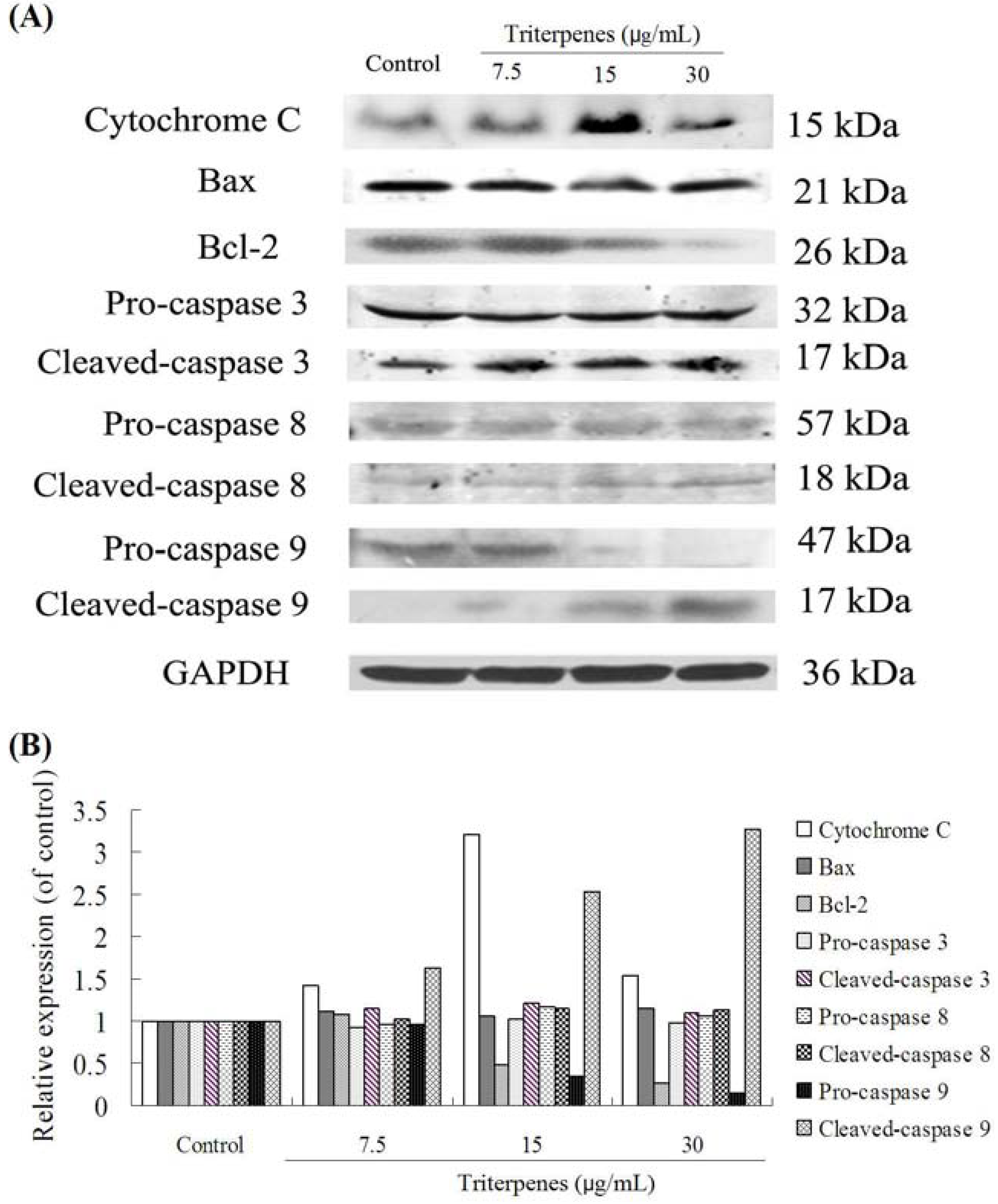
3. Discussion
4. Experimental
4.1. Chemicals and Reagents
4.2. Plant Materials
4.3. Preparation of Plant Extracts
4.4. Chromatographic Analysis of Triterpenes
4.5. Cell Culture
4.6. Cell Proliferation Assay
4.7. Anti-Tumor Activity on Lewis Tumor-Bearing C57BL/6
4.8. Flow Cytometric Analysis on Cell Cycle
4.9. Measurement of Apoptotic Ratio of A549 Cells
4.10. Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Jemal, A.; Grey, N.; Ferlay, J.; Forman, D. Global cancer transitions according to the Human Development Index (2008-2030): A population-based study. Lancet Oncol. 2012, 13, 790–801. [Google Scholar] [CrossRef]
- Kalaivani, T.; Rajasekaran, C.; Suthindhiran, K.; Mathew, L. Free radical scavenging, cytotoxic and hemolytic activities from leaves of acacia nilotica (L.) Wild. ex. Delile subsp. indica (Benth.) brenan. Evid. Based Complement. Alternat. Med. 2011, 2011, 274741. [Google Scholar]
- Ji, H.F.; Li, X.J.; Zhang, H.Y. Natural products and drug discovery. Can thousands of years of ancient medical knowledge lead us to new and powerful drug combinations in the fight against cancer and dementia? EMBO Rep. 2009, 10, 194–200. [Google Scholar] [CrossRef]
- Ma, H.D.; Deng, Y.R.; Tian, Z.; Lian, Z.X. Traditional chinese medicine and immune regulation. Clin. Rev. Allergy Immunol. 2013, 44, 229–241. [Google Scholar] [CrossRef]
- Liu, H.; Guo, Y.Q. Evaluation of immune regulation effects of Chinese medicinal formulae based on immunomics of traditional Chinese medicine. Zhongguo Zhong Yao Za Zhi 2008, 33, 2958–2960. [Google Scholar]
- Sanodiya, B.S.; Thakur, G.S.; Baghel, R.K.; Prasad, G.B.; Bisen, P.S. Ganoderma lucidum: A potent pharmacological macrofungus. Curr. Pharm. Biotechnol. 2009, 10, 717–742. [Google Scholar] [CrossRef]
- Yu, F.; Gao, J.; Zeng, Y.; Liu, C.X. Effects of adlay seed oil on blood lipids and antioxidant capacity in hyperlipidemic rats. J. Sci. Food Agric. 2011, 91, 1843–1848. [Google Scholar] [CrossRef]
- Jin, X.; Ruiz Beguerie, J.; Sze, D.M.; Chan, G.C. Ganoderma lucidum (Reishi mushroom) for cancer treatment. Cochrane Database Syst. Rev. 2012. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, Q.; Zhou, C.; Jia, W.; Da, S.L.; Nguyen, L.D.; Reutter, W.; Fan, H. GLIS, a bioactive proteoglycan fraction from Ganoderma lucidum, displays anti-tumour activity by increasing both humoral and cellular immune response. Life Sci. 2010, 87, 628–637. [Google Scholar] [CrossRef]
- Martinez-Montemayor, M.M.; Acevedo, R.R.; Otero-Franqui, E.; Cubano, L.A.; Dharmawardhane, S.F. Ganoderma lucidum (Reishi) inhibits cancer cell growth and expression of key molecules in inflammatory breast cancer. Nutr. Cancer 2011, 63, 1085–1094. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Wu, J.M. Regulation of cell cycle transition and induction of apoptosis in HL-60 leukemia cells by the combination of Coriolus versicolor and Ganoderma lucidum. Int. J. Mol. Med. 2013, 32, 251–257. [Google Scholar]
- Guo, Y.; Luo, X.; Yu, M.; Zheng, L. Active ingredients and efficacies of Ganoderma lucidum cultivated on non-medicinal parts of Chinese medicinal herbs. Wei Sheng Wu Xue Bao 2011, 51, 764–768. [Google Scholar]
- Dudhgaonkar, S.; Thyagarajan, A.; Sliva, D. Suppression of the inflammatory response by triterpenes isolated from the mushroom Ganoderma lucidum. Int. Immunopharmacol. 2009, 9, 1272–1280. [Google Scholar] [CrossRef]
- Yao, X.; Li, G.; Xu, H.; Lü, C. Inhibition of the JAK-STAT3 signaling pathway by ganoderic acid A enhances chemosensitivity of HepG2 cells to cisplatin. Planta Med. 2012, 78, 1740–1748. [Google Scholar]
- Chen, N.H.; Liu, J.W.; Zhong, J.J. Ganoderic acid T inhibits tumor invasion in vitro and in vivo through inhibition of MMP expression. Pharmacol. Rep. 2010, 62, 150–163. [Google Scholar] [CrossRef]
- Chang, U.M.; Li, C.H.; Lin, L.I.; Huang, C.P.; Kan, L.S.; Lin, S.B. Ganoderiol F, a ganoderma triterpene, induces senescence in hepatoma HepG2 cells. Life Sci. 2006, 79, 1129–1139. [Google Scholar] [CrossRef]
- Lin, S.B.; Li, C.H.; Lee, S.S.; Kan, L.S. Triterpene-enriched extracts from Ganoderma lucidum inhibit growth of hepatoma cells via suppressing protein kinase C, activating mitogen-activated protein kinases and G2-phase cell cycle arrest. Life Sci. 2003, 72, 2381–2390. [Google Scholar] [CrossRef]
- Yue, Q.X.; Song, X.Y.; Ma, C.; Feng, L.X.; Guan, S.H.; Wu, W.Y.; Yang, M.; Jiang, B.H.; Liu, X.; Cui, Y.J.; et al. Effects of triterpenes from Ganoderma lucidum on protein expression profile of HeLa cells. Phytomedicine 2010, 17, 606–613. [Google Scholar] [CrossRef]
- Thyagarajan, A.; Jedinak, A.; Nguyen, H.; Terry, C.; Baldridge, L.A.; Jiang, J.; Sliva, D. Triterpenes from Ganoderma Lucidum induce autophagy in colon cancer through the inhibition of p38 mitogen-activated kinase (p38 MAPK). Nutr. Cancer 2010, 62, 630–640. [Google Scholar] [CrossRef]
- Fang, S.; Lin, C.; Zhang, Q.; Wang, L.; Lin, P.; Zhang, J.; Wang, X. Anticancer potential of aqueous extract of alocasia macrorrhiza against hepatic cancer in vitro and in vivo. J. Ethnopharmacol. 2012, 141, 947–956. [Google Scholar] [CrossRef]
- Guckenberger, M.; Allgäuer, M.; Appold, S.; Dieckmann, K.; Ernst, I.; Ganswindt, U.; Holy, R.; Nestle, U.; Nevinny-Stickel, M.; Semrau, S.; et al. Safety and efficacy of stereotactic body radiotherapy for stage I non-small-cell lung cancer in routine clinical practice: A patterns-of-care and outcome analysis. J. Thorac. Oncol. 2013, 8, 1050–1058. [Google Scholar] [CrossRef]
- Olishevsky, S.; Shlyakhovenko, V.; Kozak, V.; Yanish, Y. Response of different organs of immune system of mice upon administration bacterial CpG DNA. Exp. Oncol. 2005, 27, 290–297. [Google Scholar]
- Nishihara, K.; Barth, R.F.; Wilkie, N.; Lang, J.C.; Oda, Y.; Kikuchi, H.; Everson, M.P.; Lotze, M.T. Increased in vitro and in vivo tumoricidal activity of a macrophage cell line genetically engineered to express IFN-gamma, IL-4, IL-6, or TNF-alpha. Cancer Gene Ther. 1995, 2, 113–124. [Google Scholar]
- Gu, J.J.; Hernandez-Ilizaliturri, F.J.; Kaufman, G.P.; Czuczman, N.M.; Mavis, C.; Skitzki, J.J.; Czuczman, M.S. The novel proteasome inhibitor carfilzomib induces cell cycle arrest, apoptosis and potentiates the anti-tumour activity of chemotherapy in rituximab-resistant lymphoma. Br. J. Haematol. 2013. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Thejass, P.; Kutan, G. Diallyl disulfide induces caspase-dependent apoptosis via mitochondria-mediated intrinsic pathway in B16F-10 melanoma cells by up-regulating p53, caspase-3 and down-regulating pro-inflammatory cytokines and nuclear factor-κβ-mediated Bcl-2 activation. J. Environ. Pathol. Toxicol. Oncol. 2010, 29, 113–125. [Google Scholar] [CrossRef]
- Wu, G.S.; Lu, J.J.; Guo, J.J.; Li, Y.B.; Tan, W.; Dang, Y.Y.; Zhong, Z.F.; Xu, Z.T.; Chen, X.P.; Wang, Y.T. Ganoderic acid DM, a natural triterpenoid, induces DNA damage, G1 cell cycle arrest and apoptosis in human breast cancer cells. Fitoterapia 2012, 83, 408–414. [Google Scholar] [CrossRef]
- Li, F.; Wang, Y.; Wang, X.; Li, J.; Cui, H.; Niu, M. Ganoderic acids suppress growth and angiogenesis by modulating the NF-κB signaling pathway in breast cancer cells. Int. J. Clin. Pharmacol. Ther. 2012, 50, 712–721. [Google Scholar]
- Liu, R.M.; Zhong, J.J. Ganoderic acid Mf and S induce mitochondria mediated apoptosis in human cervical carcinoma HeLa cells. Phytomedicine 2011, 18, 349–355. [Google Scholar] [CrossRef]
- Tang, W.; Liu, J.W.; Zhao, W.M.; Wei, D.Z.; Zhong, J.J. Ganoderic acid T from Ganoderma lucidum mycelia induces mitochondria mediated apoptosis in lung cancer cells. Life Sci. 2006, 80, 205–211. [Google Scholar] [CrossRef]
- Sample Availability: Contact the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Feng, L.; Yuan, L.; Du, M.; Chen, Y.; Zhang, M.-H.; Gu, J.-F.; He, J.-J.; Wang, Y.; Cao, W. Anti-Lung Cancer Activity through Enhancement of Immunomodulation and Induction of Cell Apoptosis of Total Triterpenes Extracted from Ganoderma luncidum (Leyss. ex Fr.) Karst. Molecules 2013, 18, 9966-9981. https://doi.org/10.3390/molecules18089966
Feng L, Yuan L, Du M, Chen Y, Zhang M-H, Gu J-F, He J-J, Wang Y, Cao W. Anti-Lung Cancer Activity through Enhancement of Immunomodulation and Induction of Cell Apoptosis of Total Triterpenes Extracted from Ganoderma luncidum (Leyss. ex Fr.) Karst. Molecules. 2013; 18(8):9966-9981. https://doi.org/10.3390/molecules18089966
Chicago/Turabian StyleFeng, Liang, Ling Yuan, Meng Du, Yan Chen, Ming-Hua Zhang, Jun-Fei Gu, Jun-Jie He, Ying Wang, and Wei Cao. 2013. "Anti-Lung Cancer Activity through Enhancement of Immunomodulation and Induction of Cell Apoptosis of Total Triterpenes Extracted from Ganoderma luncidum (Leyss. ex Fr.) Karst." Molecules 18, no. 8: 9966-9981. https://doi.org/10.3390/molecules18089966