Effect of Leguminous Lectins on the Growth of Rhizobium tropici CIAT899


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion

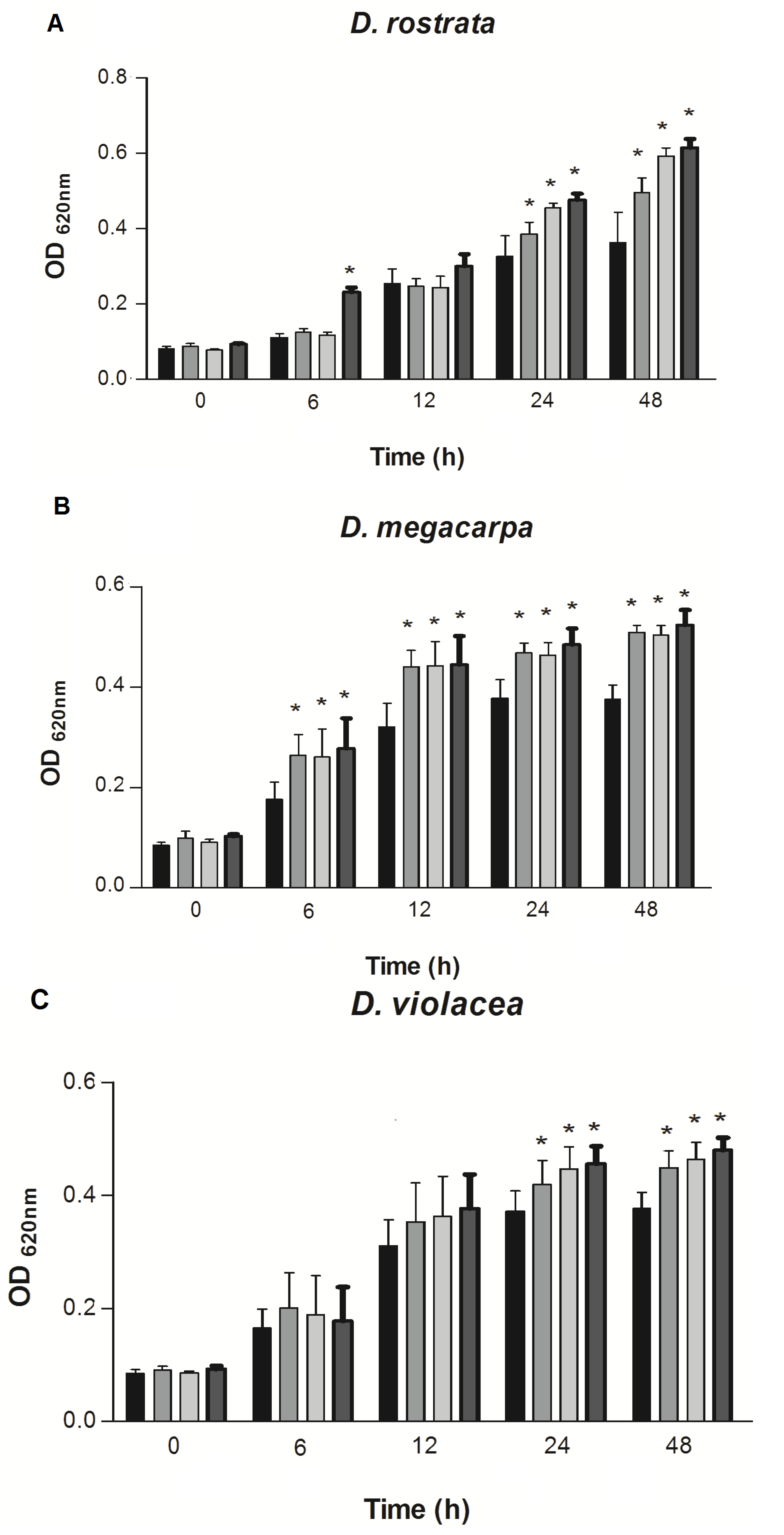
 ) Control, (
) Control, (  ) lectins at 125 µg/mL, (
) lectins at 125 µg/mL, (  ) 250 µg/mL and (
) 250 µg/mL and (  ) 500 µg/mL.
) Control, ( ) lectins at 125 µg/mL, ( ) 250 µg/mL and ( ) 500 µg/mL.
) 500 µg/mL.
) Control, ( ) lectins at 125 µg/mL, ( ) 250 µg/mL and ( ) 500 µg/mL. ) Control, ( ) lectins at 125 µg/mL, ( ) 250 µg/mL and ( ) 500 µg/mL.
) Control, ( ) lectins at 125 µg/mL, ( ) 250 µg/mL and ( ) 500 µg/mL.
) Control, ( ) lectins at 125 µg/mL, ( ) 250 µg/mL and ( ) 500 µg/mL.
) Control, ( ) lectins at 125 µg/mL, ( ) 250 µg/mL and ( ) 500 µg/mL.
3. Experimental
3.1. Lectin Purification
3.1.1. Dioclea Lectin Purification
3.1.2. VML Purification
3.1.3. BVL Purification
3.1.4. EVL Purification
3.2. SDS-PAGE
3.3. Cultivation of Microorganisms
3.4. Bacterial Growth Assay
3.5. Statistical Analyses
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Denison, R.F.; Kiers, E.T. Life histories of symbiotic rhizobia and mycorrhizal fungi. Curr. Biol. 2011, 21, 775–785. [Google Scholar] [CrossRef]
- Martinez-Romero, E.; Segovia, L.; Mercante, F.M.; Franco, A.A.; Graham, P.; Pardo, M.A. Rhizobium tropici, a novel species nodulating Phaseolusvulgaris L. beans and Leucaena sp. trees. Int. J. Syst. Bacteriol. 1991, 41, 417–426. [Google Scholar] [CrossRef]
- Ribeiro, R.A; Rogel, M.A.; López-López, A.; Ormenõ-Orrillo, E.; Barcellos, F.G.; Martínez, J.; Thompson, F.L.; Martínez-Romero, E.; Hungria, M. Reclassification of Rhizobium tropici type A strains as Rhizobium leucaenae sp. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 1179–1184. [Google Scholar] [CrossRef]
- Hungria, M.; Campo, R.J.; Mendes, I.C. Benefits of inoculation of common bean (Phaseolus. vulgaris) crop with efficient and competitive Rhizobium. tropici strains. Biol. Fertil. Soil 2003, 39, 88–93. [Google Scholar] [CrossRef]
- Gomes, D.F.; Batista, J.S.S.; Schiavon, A.L.; Andrade, D.S.; Hungria, M. Proteomic profiling of Rhizobium. tropici PRF 81: Identification of conserved and specific responses to heat stress. BMC Microbiol. 2012, 12, 1–12. [Google Scholar] [CrossRef]
- Chueire, L.M.O.; Bangel, E.V.; Mostasso, F.L.; Campo, R.J.; Pedrosa, F.O.; Hungria, M. Taxonomic classification of rhizobial strains recommended for soybean and common bean crops in Brazil based on the sequencing of the 16s rRNA gene. Rev. Bras. Ciênc. Solo 2003, 27, 833–840. [Google Scholar] [CrossRef]
- Van Damme, E.J.M.; Peumans, W.J.; Barre, A.; Rougé, P. Plant Lectins: A composite of several distinct families of structurally and evolutionary related proteins with diverse biological roles. CRC Crit. Rev. Plant. Sci. 1998, 17, 575–592. [Google Scholar]
- Sharon, N.; Lis, H. History of lectins from hemagglutinins to biological recognition molecules. Glycobiology 2004, 14, R53–R62. [Google Scholar] [CrossRef]
- Sytnikov, D.M. How to Increase the Productivity of the Soybean-Rhizobial Symbiosis. In A Comprehensive Survey of International Soybean Research—Genetics, Physiology, Agronomy and Nitrogen Relationships, 1st ed.; James, E.B., Ed.; InTech: Rijeka, Croatia, 2013; Volume 1, pp. 51–82. [Google Scholar]
- Wang, D.; Yang, S.; Tang, S.; Zhu, H. Symbiosis specificity in the legume–rhizobial mutualismo. Cell. Microbiol. 2012, 14, 334–342. [Google Scholar] [CrossRef]
- Vandenbosch, K.A.; Rodgers, L.R.; Sherrier, D.J.; Kishinevsky, B.D. A peanut nodule lectin in infected cells and in vacuoles and the extracellular matrix of nodule parenchyma. Plant Physiol. 1994, 104, 327–337. [Google Scholar]
- Geurts, R.; Bisseling, T. Rhizobium. Nod factor perception and signalling. Plant Cell. 2002, 14, 239–249. [Google Scholar]
- Roberts, N.J.; Morieri, G.; Kalsi, G.; Rose, A.; Stiller, J.; Edwards, A.; Xie, F.; Gresshoff, P.M.; Oldroyd, G.E.D.; Downie, J.A. Rhizobial and mycorrhizal symbioses in Lotus japonicus require lectin nucleotide phosphohydrolase, which acts upstream of calcium signaling. Plant Physiol. 2013, 161, 556–567. [Google Scholar] [CrossRef]
- Antonyuk, L.P.; Ignatov, V.V. The role of wheat germ agglutinin in plant–bacteria interactions: A hypothesis and the evidence in its support. Russ. J. Plant. Physiol. 2001, 48, 364–369. [Google Scholar] [CrossRef]
- Martínez, C.R.; Albertini, A.V.P.; Figueiredo, M.V.B.; Silva, V.L.; Sampaio, A.H.; Cavada, B.S.; Lima-Filho, J.L. Respiratory stimulus in Rhizobium. sp. by leguminous lectins. World J. Microbiol. Biotechnol. 2004, 20, 77–83. [Google Scholar] [CrossRef]
- Martínez, C.R.; Chanway, C.P.; Albertini, A.V.P.; Figueiredo, M.V.B.; Sampaio, A.H.; Castellon, R.R.; Cavada, B.S.; Lima-Filho, J.L. The interaction of Vatairea. macrocarca and Rhizobium. tropici: net H+ efflux stimulus and alteration of extracellular Na+ concentration. FEMS Microbiol. Lett. 2004, 238, 17–22. [Google Scholar]
- Bajaj, M.; Soni, G.; Singh, C.K. Interaction of pea (Pisum. sativum L.) lectins with rhizobial strains. Microbiol. Res. 2011, 156, 71–74. [Google Scholar] [CrossRef]
- Vasconcelos, M.A.; Cunha, C.O.; Arruda, F.V.S.; Carneiro, V.A.; Mercante, F.M.; Neto, L.G.N.; Sousa, G.S.; Rocha, B.A.M.; Teixeira, E.H.; Cavada, B.S. Lectin from Canavalia. brasiliensis Seeds (ConBr) is a valuable biotechnological tool to stimulate the growth of Rhizobium. tropici in vitro. Molecules 2002, 17, 5244–5254. [Google Scholar]
- Sytnikov, D.M.; Kirizii, D.A.; Malichenko, S.M.; Kots, S.Y. Productivity of soybean-Rhizobium symbiosis after modification of root nodule bacteria activity with exogenous proteins. Russ. J. Plant Physiol. 2007, 54, 366–372. [Google Scholar] [CrossRef]
- Sytnikov, D.M.; Kots, S.Y.; Datsenko, V.K. Efficacy of biological preparations of soybean root nodule bacteria modified with a homologous lectin. Appl. Biochem. Microbiol. 2007, 43, 274–279. [Google Scholar] [CrossRef]
- Cavada, B.S.; Barbosa, T.; Arruda, S.; Grangeiro, T.B.; Baral-Neto, M. Revisiting proteus: do minor changes in lectin structure matter in biological activity? Lessons from and potential biotechnological uses of the Diocleinae subtribe lectins. Curr. Protein Pept. Sci. 2001, 2, 1–13. [Google Scholar]
- Andrade, J.L.; Arruda, S.; Barbosa, T.; Paim, L.; Ramos, M.V.; Cavada, B.S.; Barral-Netto, M. Lectin-induced nitric oxide production. Cell Immunol. 1999, 194, 98–112. [Google Scholar] [CrossRef]
- Assreuy, A.M.S.; Shibuya, M.D.; Martins, G.J.; Souza, M.L.P.; Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A.; Ribeiro, R.A.; Flores, C.A. Anti-inflammatory effect of glucose-mannose binding lectins isolated from brazilian beans. Mediators Inflamm. 1997, 6, 201–210. [Google Scholar] [CrossRef]
- Cavalcante, T.T.; Rocha, B.A.M.; Carneiro, V.A.; Arruda, F.V.S.; Nascimento, A.S.F.; Sá, N.C.; Nascimento, K.S.; Cavada, B.S.; Teixeira, E.H. Effect of lectins from Diocleinae subtribe against oral Streptococci. Molecules 2011, 16, 3530–3543. [Google Scholar] [CrossRef]
- Pinto, F.G.S.; Chueire, L.M.O.; Vasconcelos, A.T.R.; Nicolás, M.F.; Almeida, L.G.P.; Souza, R.C.; Menna, P.; Barcellos, F.G.; Megías, M.; Hungria, M. Novel genes related to nodulation, secretion systems, and surface structures revealed by a genome draft of Rhizobiu. tropici strain PRF 81. Funct. Integr. Genomics 2009, 9, 263–270. [Google Scholar] [CrossRef]
- Laus, M.C.; Logman, T.J.; Lamers, G.E.; Van Brussel, A.A.N.; Carlson, R.W.; Kijne, J.W. A novel polar surface polysaccharide from Rhizobium. leguminosarum binds host plant lectin. Mol. Microbiol. 2006, 59, 1704–1713. [Google Scholar] [CrossRef]
- Fraysse, N.; Couderc, F.; Poinsot, V. Surface polysaccharide involvement in establishing the rhizobium–legume symbiosis. Eur. J. Biochem. 2003, 270, 1365–1380. [Google Scholar] [CrossRef]
- D’Haeze, W.; Leoff, C.; Freshour, G.; Noel, K.D.; Carlson, R.W. Rhizobium. etli CE3 bacteroid lipopolysaccharides are structurally similar but not identical to those produced by cultured CE3 bacteria. J. Biol. Chem. 2007, 282, 17101–17113. [Google Scholar]
- Castro, C.; Molinaro, A.; Lanzetta, R.; Silipo, A.; Parrilli, M. Lipopolysaccharide structures from Agrobacterium and Rhizobiaceae species. Carbohydr. Res. 2008, 343, 1924–1933. [Google Scholar] [CrossRef]
- Muszyński, A.; Laus, M.; Kijne, J.W.; Carlson, R.W. Structures of the lipopolysaccharides from Rhizobium. leguminosarum RBL5523 and its UDP-glucose dehydrogenase mutant (exo5). Glycobiology 2011, 21, 55–68. [Google Scholar] [CrossRef]
- Moreira, R.A.; Cordeiro, E.F.; Grangeiro, T.B.; Martins, J.L.; Ramos, M.V.; Oliveira, J.T.A.; Cavada, B.S. Isolation and partial characterization of a lectin from Dioclea. violacea Benth seeds. Rev. Bras. Fisiol. Veg. 1996, 8, 23–29. [Google Scholar]
- Moreira, R.A.; Monteiro, A.C.O.; Horta, A.C.G.; Oliveira, J.T.A.; Cavada, B.S. Isolation and characterization of Dioclea. altissima var. megacarpa seed lectin. Phytochemistry 1997, 46, 139–144. [Google Scholar]
- Cavada, B.S.; Grangeiro, T.B.; Ramos, M.V.; Cordeiro, E.F.; Oliveira, J.T.A.; Moreira, R.A. Isolation and partial characterization of a lectin from Dioclea. rostrata Benth seeds. Rev. Bras. Fisiol. Veg. 1996, 8, 31–36. [Google Scholar]
- Cavada, B.S.; Santos, C.F.; Grangeiro, T.B.; Nunes, E.P.; Sales, P.V.P.; Ramos, R.L.; De Sousa, F.A.M.; Crisostomo, C.V.; Calvete, J.J. Purification and characterization of a lectin from seeds of Vatairea macrocarpa Duke. Phytochemistry 1998, 49, 675–680. [Google Scholar] [CrossRef]
- Moraes, S.M.D.; Cavada, B.S.; Moreira, R.A.; Roquebarreira, M.C.; Santos-De-Oliveira, R.; Pinto, V.P.T.; Oliveira, J.T.A. Purification, Physicochemical characterisation and biological properties of a lectin from Erythrina velutina forma aurantiaca seeds. Braz. J. Med. Biol. Res. 1996, 29, 977–985. [Google Scholar]
- Pinto, L.S.; Nagano, C.S.; Oliveira, T.M.; Moura, T.R.; Sampaio, A.H.; Debray, H.; Pinto, V.P.T.; Dellagostin, O.A.; Cavada, B.S. Purification and molecular cloning of a new galactose-specific lectin from Bauhinia variegata seeds. J. Biosci. 2008, 33, 355–363. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Vincent, J.M. A Manual for the Practical Study of Root Nodule Bacteria, International Biological Programme Handbook No. 15; Blackwell Science Publications: Oxford, UK, 1970; p. 164. [Google Scholar]
- Sample Availability: Contact the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
De Vasconcelos, M.A.; Cunha, C.O.; Arruda, F.V.S.; Carneiro, V.A.; Bastos, R.M.; Mercante, F.M.; Nascimento, K.S.d.; Cavada, B.S.; Santos, R.P.d.; Teixeira, E.H. Effect of Leguminous Lectins on the Growth of Rhizobium tropici CIAT899. Molecules 2013, 18, 5792-5803. https://doi.org/10.3390/molecules18055792
De Vasconcelos MA, Cunha CO, Arruda FVS, Carneiro VA, Bastos RM, Mercante FM, Nascimento KSd, Cavada BS, Santos RPd, Teixeira EH. Effect of Leguminous Lectins on the Growth of Rhizobium tropici CIAT899. Molecules. 2013; 18(5):5792-5803. https://doi.org/10.3390/molecules18055792
Chicago/Turabian StyleDe Vasconcelos, Mayron Alves, Cláudio Oliveira Cunha, Francisco Vassiliepe Sousa Arruda, Victor Alves Carneiro, Rafaela Mesquita Bastos, Fábio Martins Mercante, Kyria Santiago do Nascimento, Benildo Sousa Cavada, Ricardo Pires dos Santos, and Edson Holanda Teixeira. 2013. "Effect of Leguminous Lectins on the Growth of Rhizobium tropici CIAT899" Molecules 18, no. 5: 5792-5803. https://doi.org/10.3390/molecules18055792