Quercetin Improves Baroreflex Sensitivity in Spontaneously Hypertensive Rats

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
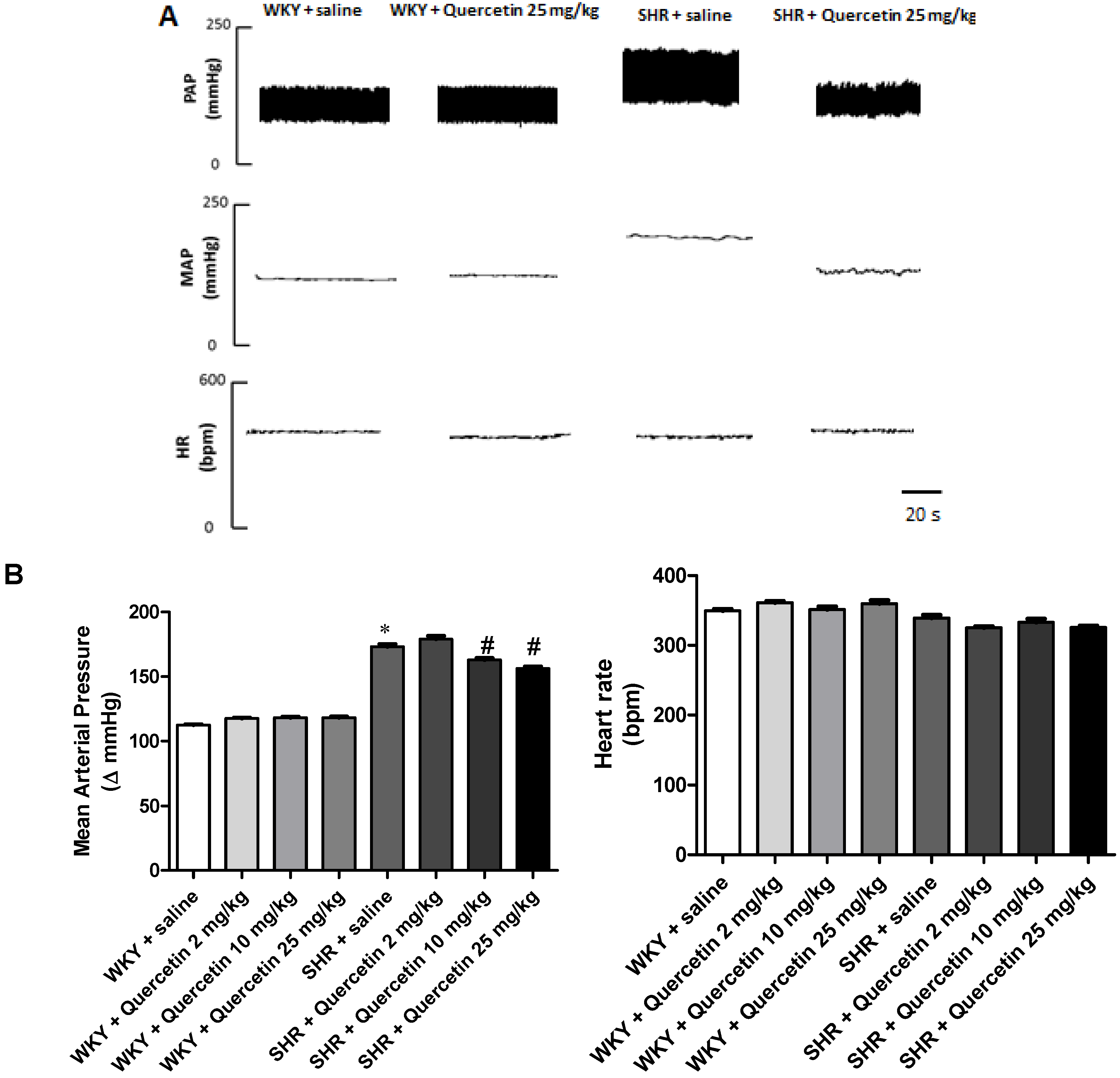
2.1. Administration of Quercetin Reduces Blood Pressure in Hypertensive Rats
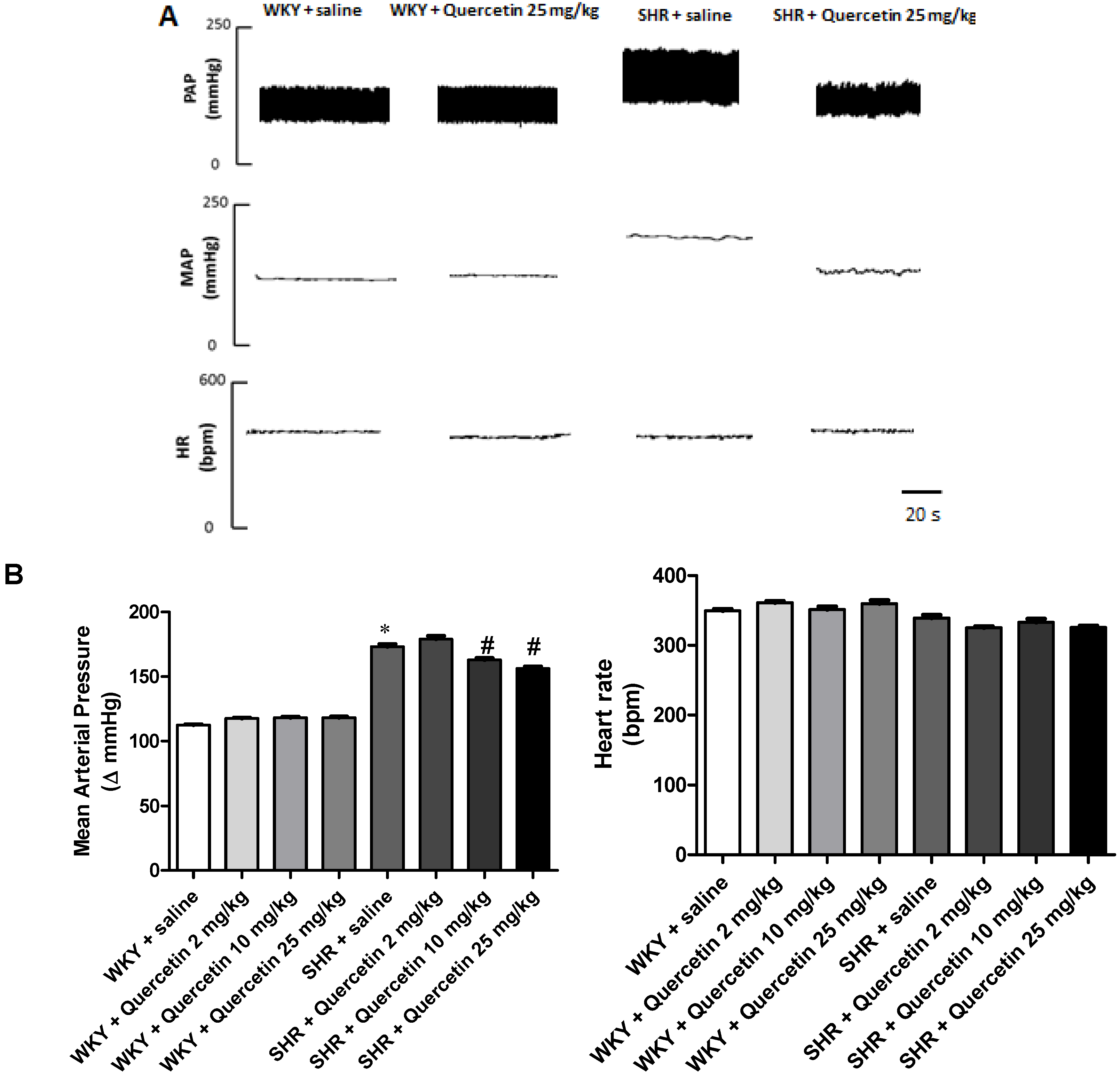
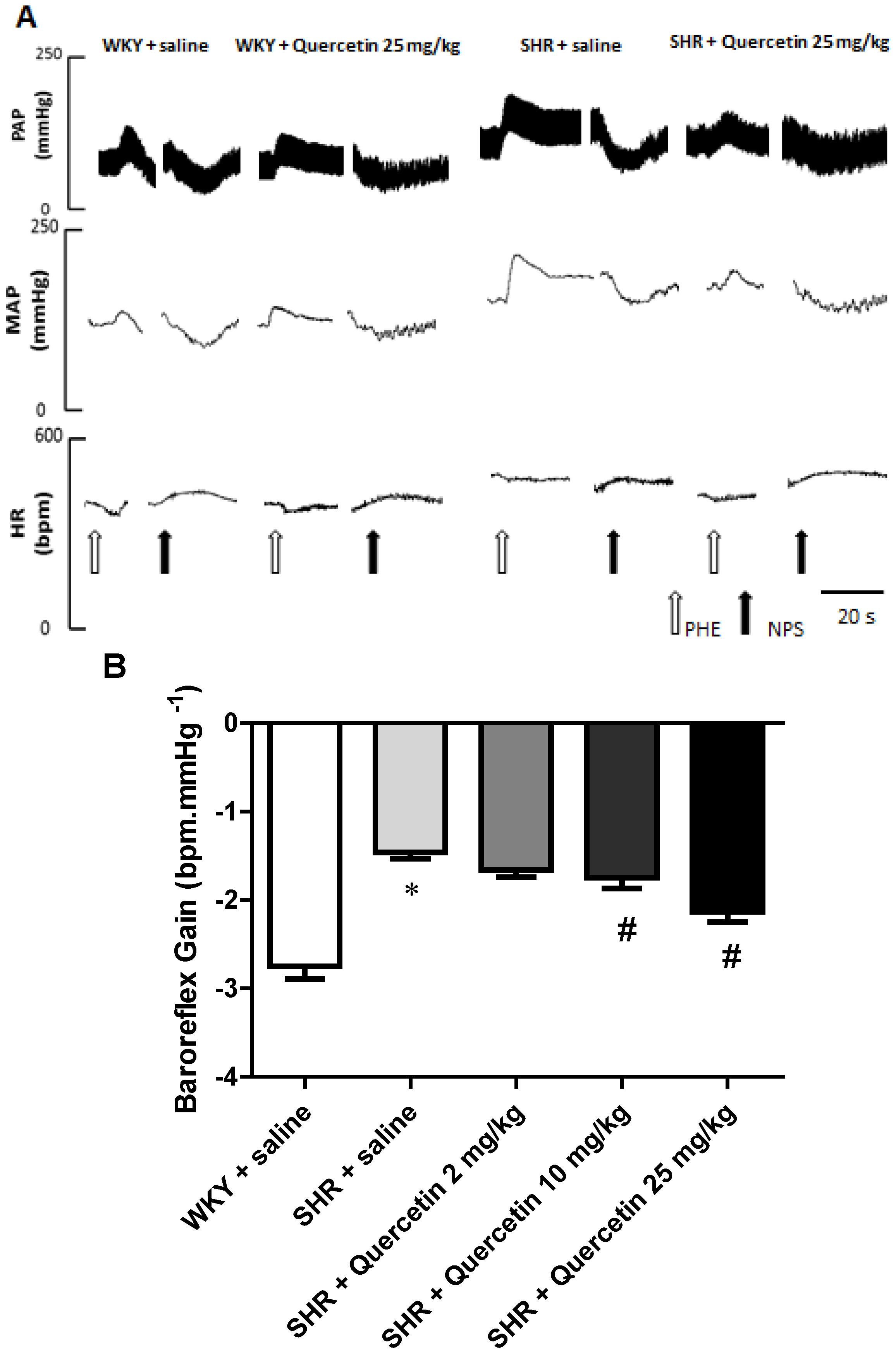
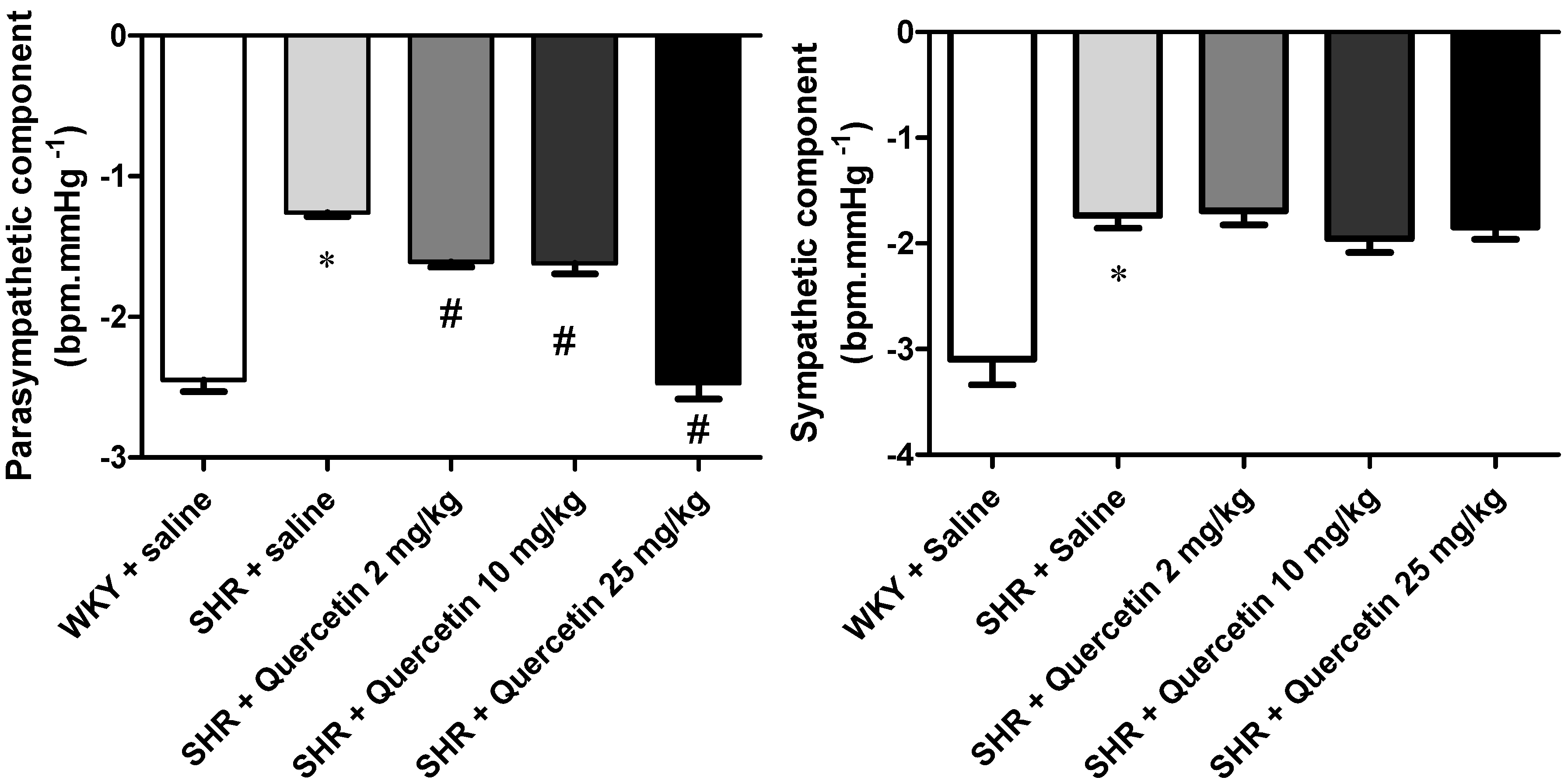
2.2. Administration of Quercetin Improves Baroreflex Sensitivity in Spontaneously Hypertensive Rats
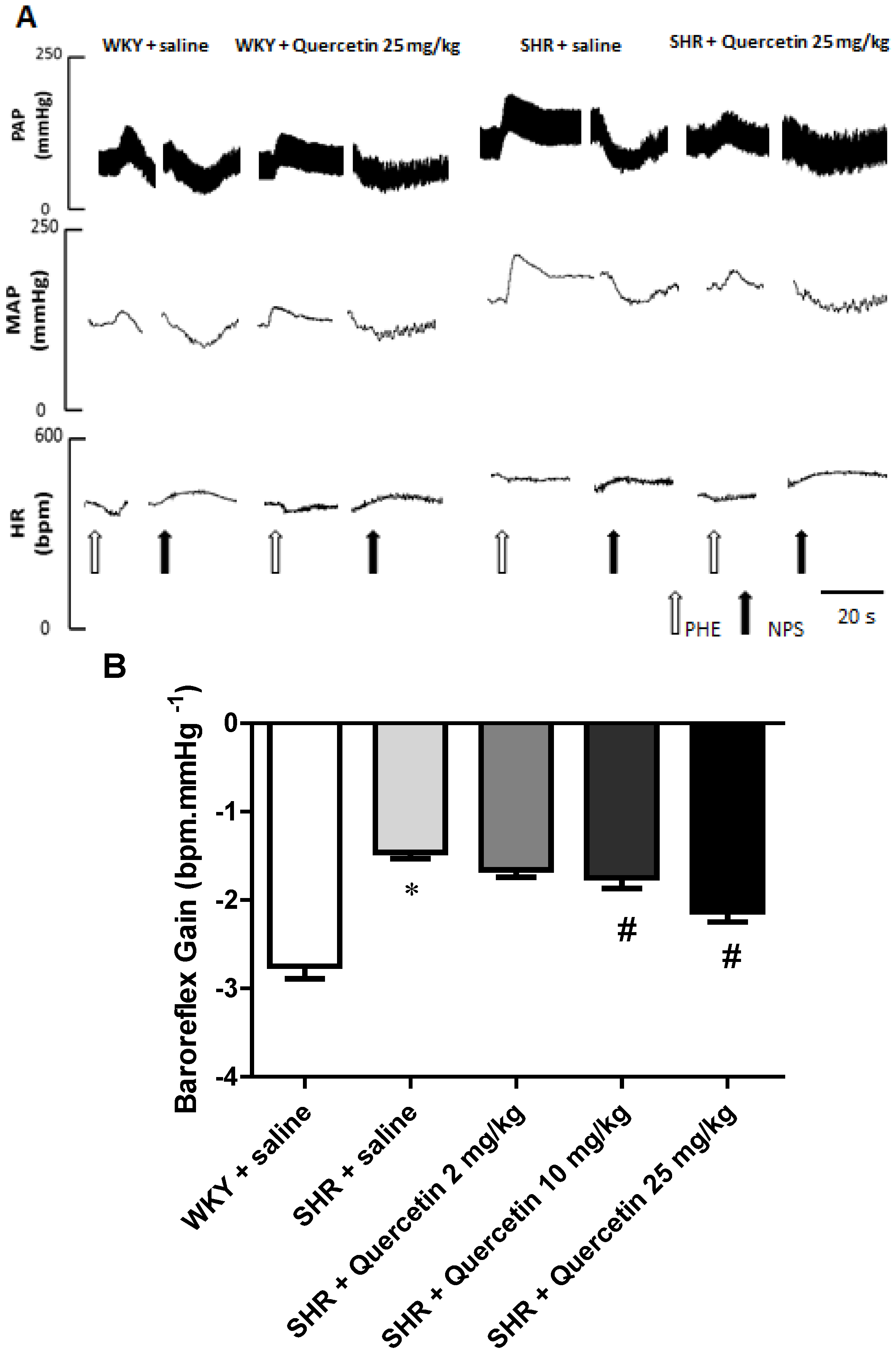

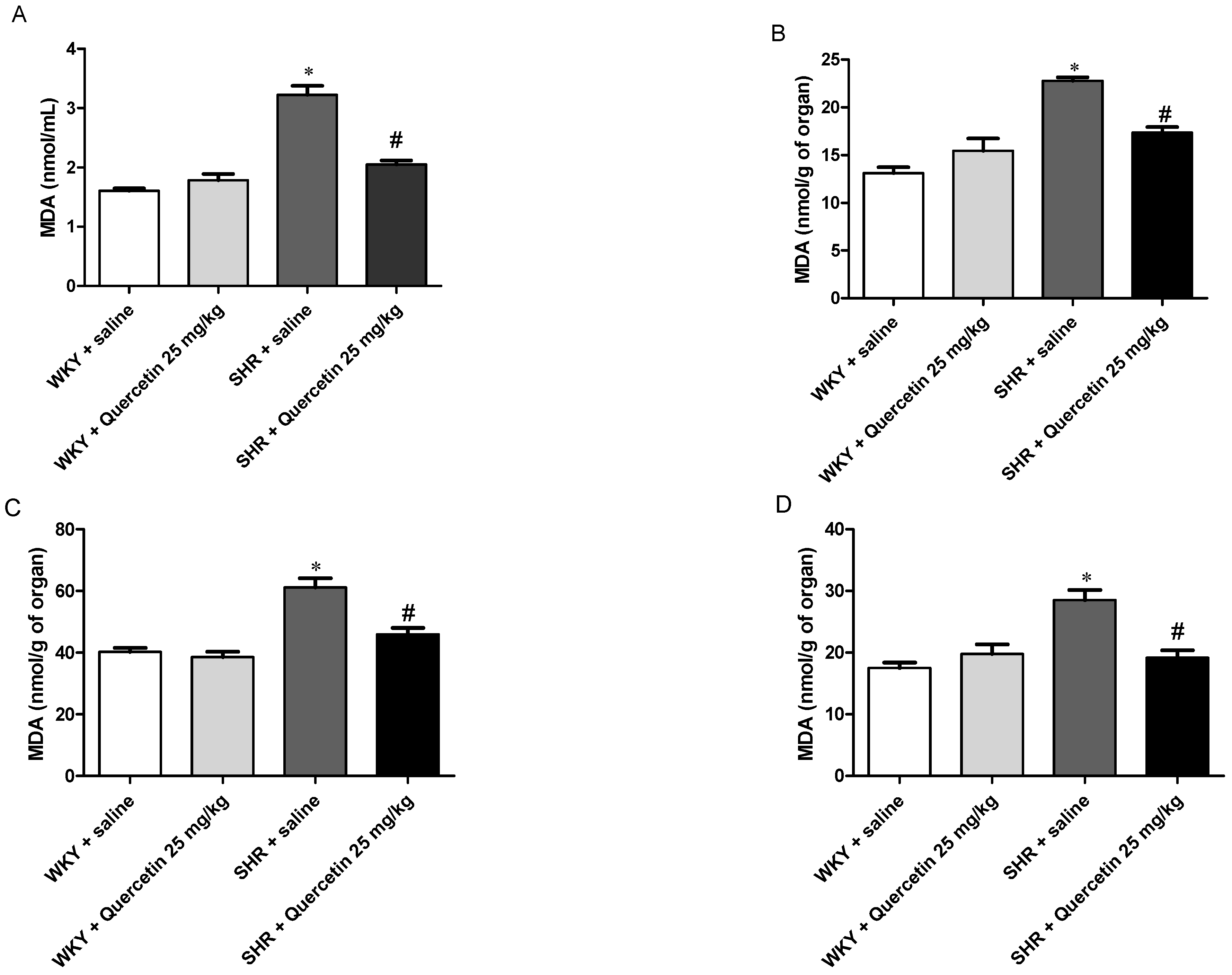
2.3. Quercetin Reduces Oxidative Stress in Hypertensive Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum (nmol/mL) | n | Heart (nmol/g) | n | Liver (nmol/g) | n | Kidney (nmol/g) | n | |
|---|---|---|---|---|---|---|---|---|
| WKY + Saline | 1.6 ± 1.1 | 9 | 13.13 ± 1.5 | 7 | 17.5 ± 2.1 | 6 | 40.2 ± 3.0 | 5 |
| WKY + Quercetin (25 mg/kg) | 1.78 ± 0.2 | 4 | 15.45 ± 2.5 | 4 | 19.79 ± 3.3 | 5 | 38.55 ± 4.7 | 7 |
| SHR + Saline | 3.2 ± 0.3 * | 6 | 22.78 ± 0.9 * | 6 | 28.51 ± 3.9 * | 6 | 61.13 ± 7.3 * | 6 |
| SHR + Quercetin (25 mg/kg) | 2.04 ± 0.2 # | 6 | 17.33 ± 1.6 # | 7 | 19.19 ± 2.6 # | 5 | 45.98 ± 4.1 # | 4 |

3. Experimental
3.1. Animals and Treatment
3.2. Blood Pressure and Heart Rate Recordings
3.3. Baroreflex Sensitivity Test
3.4. Tiobarbituric Acid Reactive Species (TBARS) Assay
3.5. Statistical Analyses
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Thomas, G.D. Neural control of the circulation. Adv. Physiol. Educ. 2011, 35, 28–32. [Google Scholar] [CrossRef]
- Chobanian, A.V.; Bakris, G.L.; Black, H.R.; Cushman, W.C.; Green, L.A.; Izzo, J.L., Jr.; Jones, D.W.; Materson, B.J.; Oparil, S.; Wright, J.T., Jr.; et al. Seventh report of the Joint National Committee on Prevention, Detection, Evaluation, and Treatment of High Blood Pressure. Hypertension 2003, 42, 1206–1252. [Google Scholar]
- Botelho-Ono, M.S.; Pina, H.V.; Sousa, K.H.F.; Nunes, F.C.; Medeiros, I.A.; Braga, V.A. Acute superoxide scavenging restores depressed baroreflex sensitivity in renovascular hypertensive rats. Auton. Neurosci. 2011, 159, 38–44. [Google Scholar] [CrossRef]
- Guimarães, D.D.; Carvalho, C.C.; Braga, V.A. Scavenging of NADPH oxidase-derived superoxide anions improves depressed baroreflex sensitivity in spontaneously hypertensive rats. Clin. Exp. Pharmacol. Physiol. 2012, 39, 373–378. [Google Scholar] [CrossRef]
- Braga, V.A. Depressed Baroreflex Sensitivity in Hypertensive Rats: A Role for Reactive Oxygen Species. J. Hypertens. 2012, 1, e103. [Google Scholar]
- Lazartigues, E. Inflammation and neurogenic hypertension: A new role for the circumventricular organs? Circ. Res. 2010, 107, 166–167. [Google Scholar]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Zimmerman, M.C.; Lazartigues, E.; Sharma, R.V.; Davisson, R.L. Hypertension Caused by Angiotensin II Infusion Involves Increased Superoxide Production in the Central Nervous System. Circ. Res. 2004, 95, 210–216. [Google Scholar] [CrossRef]
- Pedro-Botet, J.; Covas, M.I.; Martín, S.; Rubiés-Prat, J. Decreased endogenous antioxidant enzymatic status in essential hypertension. J. Hum. Hypertens 2000, 14, 343–345. [Google Scholar]
- Romero, J.C.; Reckelhoff, J.F. Role of Angiotensin and Oxidative Stress in Essential Hypertension. Hypertension 1999, 34, 943–949. [Google Scholar] [CrossRef]
- Nalivaiko, E. Animal models of psychogenic cardiovascular disorders: What we can learn from them and what we cannot. Clin. Exp. Pharmacol. Physiol. 2011, 38, 115–125. [Google Scholar] [CrossRef]
- Braga, V.A.; Medeiros, I.A.; Ribeiro, T.P.; França-Silva, M.S.; Botelho-Ono, M.S.; Guimarães, D.D. Angiotensin-II-induced reactive oxygen species along the SFO-PVN-RVLM pathway: Implications in neurogenic hypertension. Braz. J. Med. Biol. Res. 2011, 44, 871–876. [Google Scholar] [CrossRef]
- Braga, V.A. Dietary salt enhances angiotensin-II-induced superoxide formation in the rostral ventrolateral medulla. Auton. Neurosci. 2010, 155, 14–18. [Google Scholar] [CrossRef]
- Briones, A.M.; Touyz, R.M. Oxidative stress and hypertension: Current concepts. Curr. Hypertens. Rep. 2010, 12, 135–142. [Google Scholar] [CrossRef]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- Duarte, J.; Pérez-Palencia, R.; Vargas, F.; Ocete, M.A.; Pérez-Vizcaino, F.; Zarzuelo, A.; Tamargo, J. Antihypertensive effects of the flavonoid quercetin in spontaneously hypertensive rats. Br. J. Pharmacol. 2001, 133, 117–124. [Google Scholar] [CrossRef]
- Pérez-Vizcaíno, F.; Ibarra, M.; Cogolludo, A.L.; Duarte, J.; Zaragozá-Arnáez, F.; Moreno, L.; López-López, G.; Tamargo, J. Endothelium-independent vasodilator effects of the flavonoid quercetin and its methylated metabolites in rat conductance and resistance arteries. J. Pharmacol. Exp. Ther. 2002, 302, 66–72. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Oue, E. Antihypertensive effect of quercetin in rats fed with a high-fat high-sucrose diet. Biosci. Biotechnol. Biochem. 2006, 70, 933–939. [Google Scholar] [CrossRef]
- Romero, M.; Jiménez, R.; Sánchez, M.; López-Sepúlveda, R.; Zarzuelo, M.J.; O’Valle, F.; Zarzuelo, A.; Pérez-Vizcaíno, F.; Duarte, J. Quercetin inhibits vascular superoxide production induced by endothelin-1: Role of NADPH oxidase, uncoupled eNOS and PKC. Atherosclerosis 2009, 202, 58–67. [Google Scholar] [CrossRef]
- Sánchez, M.; Galisteo, M.; Vera, R.; Villar, I.C.; Zarzuelo, A.; Tamargo, J.; Pérez-Vizcaíno, F.; Duarte, J. Quercetin downregulates NADPH oxidase, increases eNOS activity and prevents endothelial dysfunction in spontaneously hypertensive rats. J. Hypertens. 2006, 24, 75–84. [Google Scholar] [CrossRef]
- García-Saura, M.F.; Galisteo, M.; Villar, I.C.; Bermejo, A.; Zarzuelo, A.; Vargas, F.; Duarte, J. Effects of chronic quercetin treatment in experimental renovascular hypertension. Mol. Cell. Biochem. 2005, 270, 147–155. [Google Scholar] [CrossRef]
- Khoo, N.K.H.; White, C.R.; Pozzo-Miller, L.; Zhou, F.; Constance, C.; Inoue, T.; Patel, R.P.; Parks, D.A. Dietary flavonoid quercetin stimulates vasorelaxation in aortic vessels. Free Radic. Biol. Med. 2010, 49, 339–347. [Google Scholar]
- Larson, A.J.; Symons, J.D.; Jalili, T. Therapeutic potential of quercetin to decrease blood pressure: Review of efficacy and mechanisms. Adv. Nutr. 2012, 3, 39–46. [Google Scholar]
- Carlstrom, J.; Symons, J.D.; Wu, T.C.; Bruno, R.S.; Litwin, S.E.; Jalili, T. A quercetin supplemented diet does not prevent cardiovascular complications in spontaneously hypertensive rats. J. Nutr. 2007, 137, 628–633. [Google Scholar]
- Galindo, P.; González-Manzano, S.; Zarzuelo, M.J.; Gómez-Guzmán, M.; Quintela, A.M.; González-Paramás, A.; Santos-Buelga, C.; Pérez-Vizcaíno, F.; Duarte, J.; Jiménez, R. Different cardiovascular protective effects of quercetin administered orally or intraperitoneally in spontaneously hypertensive rats. Food Funct. 2012, 3, 643–650. [Google Scholar] [CrossRef]
- Frohlich, E.D.; Pfeffer, M.A. Adrenergic mechanisms in human hypertension and in spontaneously hypertensive rats. Clin. Sci. Mol. Med. Suppl. 1975, 2, 225–238. [Google Scholar]
- Folkow, B. Physiological aspects of primary hypertension. Physiol. Rev. 1982, 62, 347–504. [Google Scholar]
- Kezeli, T.; Gongadze, N.; Chapichadze, Z.; Bakuridze, K.; Chirakadze, K. Effect of combination of zafirlukast and quercetin on baroreflex sensitivity and endothelin production in rats with myocardial infarction. Int. J. Clin. Pharmacol. Ther. 2010, 48, 335–341. [Google Scholar]
- Chapleau, M.W.; Cunningham, J.T.; Sullivan, M.J.; Wachtel, R.E.; Abboud, F.M. Structural versus functional modulation of the arterial baroreflex. Hypertension 1995, 26, 341–347. [Google Scholar] [CrossRef]
- Li, Z.; Mao, H.Z.; Abboud, F.M.; Chapleau, M.W. Oxygen-derived free radicals contribute to baroreceptor dysfunction in atherosclerotic rabbits. Circ. Res. 1996, 79, 802–811. [Google Scholar] [CrossRef]
- Girouard, H.; Denault, C.; Chulak, C.; de Champlain, J. Treatment by n-acetylcysteine and melatonin increases cardiac baroreflex and improves antioxidant reserve. Am. J. Hypertens. 2004, 17, 947–954. [Google Scholar] [CrossRef]
- Nishihara, M.; Hirooka, Y.; Kishi, T.; Sunagawa, K. Different role of oxidative stress in paraventricular nucleus and rostral ventrolateral medulla in cardiovascular regulation in awake spontaneously hypertensive rats. J. Hypertens. 2012, 30, 1758–1765. [Google Scholar] [CrossRef]
- Kwon, T.W.; van der Veen, J. Ultraviolet and infrared absorption spectra of malonaldehyde in organic solvents. J. Agric. Food Chem. 1968, 16, 639–642. [Google Scholar] [CrossRef]
- Gutteridge, J.M. Thiobarbituric acid-reactivity following iron-dependent free-radical damage to amino acids and carbohydrates. FEBS Lett. 1981, 128, 343–346. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar]
- Liu, C.M.; Sun, Y.Z.; Sun, J.M.; Ma, J.Q.; Cheng, C. Protective role of quercetin against lead-induced inflammatory response in rat kidney through the ROS-mediated MAPKs and NF-κB pathway. Biochim. Biophys. Acta 1820, 1693–1703. [Google Scholar]
- Zhang, Y.; Gao, Z.; Liu, J.; Xu, Z. Protective effects of baicalin and quercetin on an iron-overloaded mouse: Comparison of liver, kidney and heart tissues. Nat. Prod. Res. 2011, 25, 1150–1160. [Google Scholar] [CrossRef]
- Braga, V.A.; Burmeister, M.A.; Sharma, R.V.; Davisson, R.L. Cardiovascular responses to peripheral chemoreflex activation and comparison of different methods to evaluate baroreflex gain in conscious mice using telemetry. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R1168–R1174. [Google Scholar] [CrossRef]
- Rios, E.R.V.; Rocha, N.F.M.; Venâncio, E.T.; Moura, B.A.; Feitosa, M.L.; Cerqueira, G.S.; Soares, P.M.G.; Woods, D.J.; de Sousa, F.C.F.; Leal, L.K.A.M.; et al. Mechanisms involved in the gastroprotective activity of esculin on acute gastric lesions in mice. Chem. Biol. Interact. 2010, 188, 246–254. [Google Scholar] [CrossRef]
- Sample Availability: Samples of compounds are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Monteiro, M.M.O.; França-Silva, M.S.; Alves, N.F.B.; Porpino, S.K.P.; Braga, V.A. Quercetin Improves Baroreflex Sensitivity in Spontaneously Hypertensive Rats. Molecules 2012, 17, 12997-13008. https://doi.org/10.3390/molecules171112997
Monteiro MMO, França-Silva MS, Alves NFB, Porpino SKP, Braga VA. Quercetin Improves Baroreflex Sensitivity in Spontaneously Hypertensive Rats. Molecules. 2012; 17(11):12997-13008. https://doi.org/10.3390/molecules171112997
Chicago/Turabian StyleMonteiro, Matheus M. O., Maria S. França-Silva, Naiane F. B. Alves, Suênia K. P. Porpino, and Valdir A. Braga. 2012. "Quercetin Improves Baroreflex Sensitivity in Spontaneously Hypertensive Rats" Molecules 17, no. 11: 12997-13008. https://doi.org/10.3390/molecules171112997