Protective Effect of Total Flavonoids of Seabuckthorn (Hippophae rhamnoides) in Simulated High-Altitude Polycythemia in Rats

Abstract
:1. Introduction
2. Results
2.1. Effect of Total Flavonoids on Routine Blood Parameters

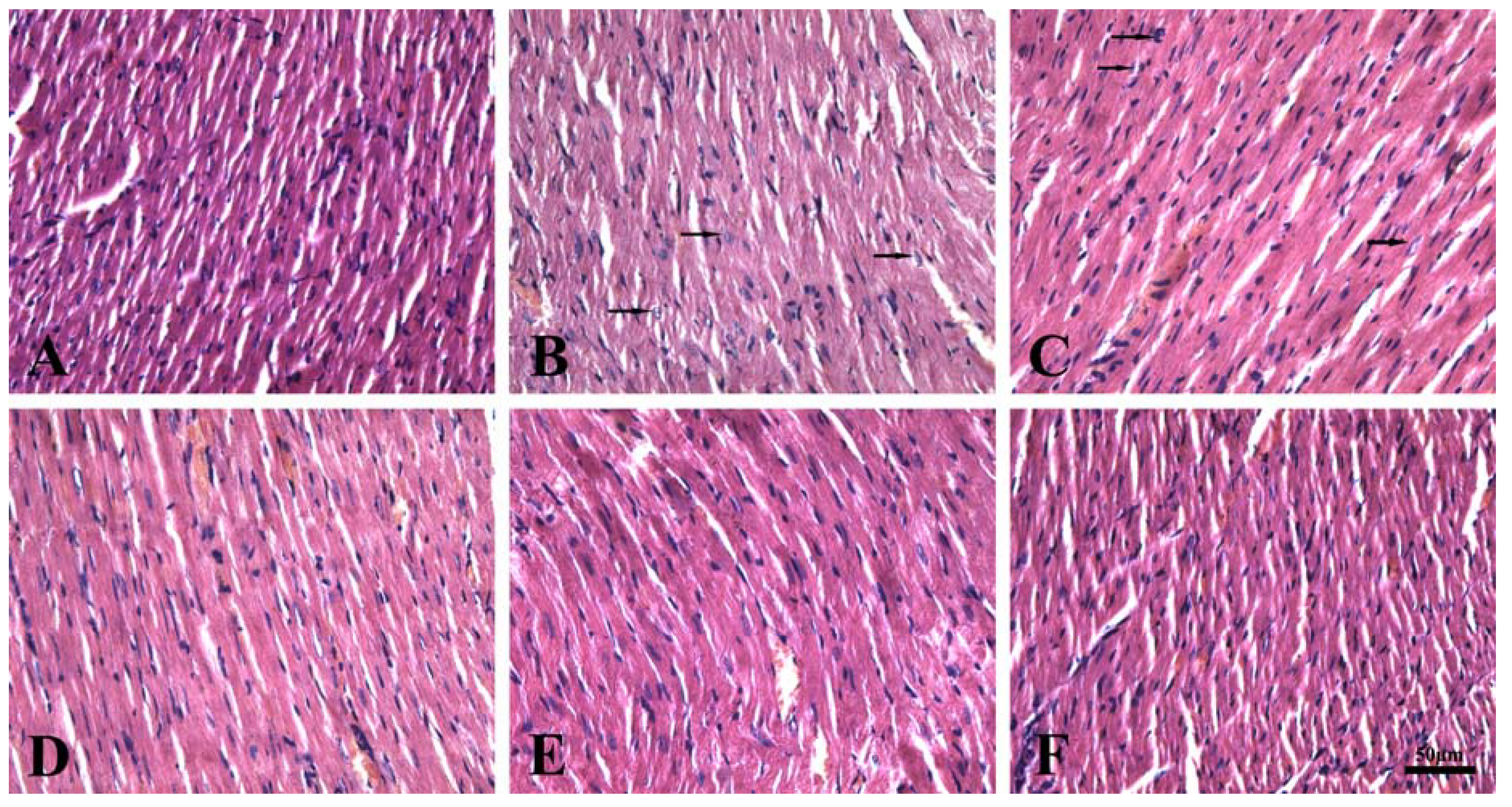{kind=link}
| Groups | RBC (×1012/L) | Hemoglobin (g/L) | Hematocrit (L/L) | MCH (pg) | Erythropoietin (mIU) |
|---|---|---|---|---|---|
| Normoxic control | 7.14 ± 0.62 | 134.49 ± 13.33 | 0.392 ± 0.010 | 18.83 ± 0.94 | 7.05 ± 0.43 |
| Hypoxic control | 9.86 ± 0.54 * | 236.70 ± 13.09 * | 0.665 ± 0.013 * | 24.06 ± 1.50 * | 8.54 ± 0.25 * |
| Hypoxic + 35 mg/kg TF | 9.68 ± 0.83 | 231.49 ± 10.25 | 0.657 ± 0.014 | 24.02 ± 1.53 | 8.45 ± 0.31 |
| Hypoxic + 70 mg/kg TF | 8.28 ± 0.58 †† | 184.52 ± 8.48 †† | 0.518 ± 0.010 †† | 22.36 ± 1.35 †† | 7.51 ± 0.27 †† |
| Hypoxic + 140 mg/kg TF | 8.13 ± 0.46 †† | 174.45 ± 9.50 †† | 0.479 ± 0.018 †† | 21.50 ± 1.32 †† | 7.42 ± 0.39 †† |
| Hypoxic + Tianji capsule | 8.43 ± 0.55 †† | 191.83 ± 10.49 †† | 0.538 ± 0.010 †† | 22.55 ± 1.59 † | 7.70 ± 0.15 †† |
2.2. Effect of Total Flavonoids on Serum Erythropoietin Content
2.3. Effect of Total Flavonoids on Hematologic Rheology Parameters
| Groups | Whole blood viscosity at | Plasma viscosity | RBC aggre-gation index | RBC deforma-tion index | RBC electro-phoretic time | ||
|---|---|---|---|---|---|---|---|
| High shear rate (200/s) | Med. shear rate (30/s) | Low shear rate (3/s) | |||||
| Normoxic control | 4.87 ± 0.90 | 6.09 ± 0.60 | 12.38 ± 1.20 | 1.44 ± 0.15 | 8.39 ± 0.85 | 0.70 ± 0.08 | 16.74 ± 0.95 |
| Hypoxic control | 15.80 ± 1.03 * | 16.27 ± 1.45 * | 29.06 ± 2.61 * | 2.23 ± 0.30 * | 16.17 ± 1.97 * | 0.48 ± 0.06 * | 28.28 ± 1.98 * |
| Hypoxic + 35 mg/kg TF | 15.66 ± 1.26 | 15.59 ± 1.14 | 28.79 ± 1.83 | 2.12 ± 0.31 | 16.09 ± 1.70 | 0.49 ± 0.07 | 28.16 ± 1.84 |
| Hypoxic + 70 mg/kg TF | 11.65 ± 0.91 † | 11.92 ± 1.82 † | 19.69 ± 2.17 † | 1.75 ± 0.14 † | 9.82 ± 0.74 † | 0.63 ± 0.09 † | 21.14 ± 1.41 † |
| Hypoxic + 140 mg/kg TF | 11.02 ± 1.17 † | 10.77 ± 1.43 † | 18.77 ± 2.50 † | 1.65 ± 0.14 † | 9.70 ± 1.17 † | 0.64 ± 0.09 † | 20.95 ± 1.24 † |
| Hypoxic + Tianji capsule | 11.83 ± 1.43 † | 11.07 ± 1.14 † | 20.94 ± 2.13 † | 1.72 ± 0.14 † | 10.35 ± 1.60 † | 0.62 ± 0.06 † | 21.21 ± 1.96 † |
2.4. Effect of Total Flavonoids on Hemodynamic Parameters
| Groups | Left ventricle systolic pressure (mmHg) | Left ventricular end-diastolic pressure (mmHg) | Left ventricular +dp/dt (max) | Left ventricular −dp/dt (max) | Heart rate | Mean arterial pressure (mmHg) | RV/(LV + S)Ratio (%) |
|---|---|---|---|---|---|---|---|
| Normoxic control | 172.387 ± 9.942 | 4.122 ± 0.335 | 18260.974 ± 1653.984 | 10574.945 ± 1086.160 | 388.0 ± 33.9 | 117.11 ± 8.43 | 0.34 ± 0.03 |
| Hypoxic control | 152.864 ± 8.646 * | 2.943 ± 0.319 * | 13502.003 ± 1361.975 * | 6980.370 ± 998.746* | 441.0 ± 18.5* | 97.15 ± 3.79* | 0.48 ± 0.02* |
| Hypoxic + 35 mg/kg TF | 154.924 ± 10.793 | 2.947 ± 0.386 | 13638.763 ± 909.229 | 7108.272 ± 867.963 | 438.0 ± 19.3 | 99.26 ± 4.90 | 0.45 ± 0.03 |
| Hypoxic + 70 mg/kg TF | 164.574 ± 9.250 † | 3.541 ± 0.535 †† | 14663.368 ± 1112.082 † | 7813.614 ± 793.876 † | 391.0 ± 26.9 †† | 105.77 ± 4.56 †† | 0.41 ± 0.03 †† |
| Hypoxic + 140 mg/kg TF | 166.387 ± 12.709 †† | 3.614 ± 0.477 †† | 15090.811 ± 887.559 †† | 7906.700 ± 741.118 † | 387.0 ± 13.4 †† | 108.56 ± 4.89 †† | 0.40 ± 0.03 †† |
| Hypoxic + Tianji capsule | 162.420 ± 8.740 † | 3.643 ± 0.542 †† | 14796.312 ± 1261.311 † | 7927.825 ± 781.023 † | 396.0 ± 20.1 †† | 103.10 ± 6.14 † | 0.41 ± 0.02 †† |
2.5. Effect of Total Flavonoids on Arterial Blood Gases
| Groups | pH | pO2 (mmHg) | SpO2 (%) | pCO2 (mmHg) |
|---|---|---|---|---|
| Normoxic control | 7.36 ± 0.03 | 121.8 ± 7.5 | 98.5 ± 0.5 | 35.6 ± 3.8 |
| Hypoxic control | 7.27 ± 0.03 * | 71.0 ± 4.0 * | 92.8 ± 2.3 * | 21.2 ± 2.0 * |
| Hypoxic + 35 mg/kg TF | 7.28 ± 0.01 | 74.1 ± 3.6 | 92.1 ± 2.1 | 21.3 ± 1.8 |
| Hypoxic + 70 mg/kg TF | 7.32 ± 0.02 †† | 83.3 ± 4.4 †† | 95.3 ± 0.9 †† | 24.1 ± 1.0 †† |
| Hypoxic + 140 mg/kg TF | 7.33 ± 0.04 †† | 88.0 ± 3.3 †† | 96.7 ± 1.2 †† | 24.9 ± 1.9 †† |
| Hypoxic + Tianji capsule | 7.33 ± 0.04 †† | 82.4 ± 3.4 †† | 94.2 ± 1.3 † | 23.5 ± 1.6 † |
2.6. Effect of Total Flavonoids on Electrolyte Concentration
| Groups | Na+ (mmol/L) | K+ (mmol/L) | Cl− (mmol/L) | HCO3− (mmol/L) |
|---|---|---|---|---|
| Normoxic control | 135.6 ± 3.3 | 4.97 ± 0.41 | 104.2 ± 2.7 | 18.1 ± 1.6 |
| Hypoxic control | 117.3 ± 3.0 * | 6.03 ± 0.25 * | 82.9 ± 2.1 * | 11.4 ± 1.3 * |
| Hypoxic + 35 mg/kg TF | 118.4 ± 2.9 | 5.87 ± 0.30 | 84.4 ± 2.8 | 11.5 ± 1.1 |
| Hypoxic + 70 mg/kg TF | 130.8 ± 1.2 † | 5.23 ± 0.25 † | 102.6 ± 2.1 † | 12.9 ± 1.3 † |
| Hypoxic + 140 mg/kg TF | 131.7 ± 1.8 † | 5.26 ± 0.23 † | 103.8 ± 2.2 † | 13.8 ± 1.2 † |
| Hypoxic + Tianji capsule | 128.6 ± 2.5 † | 5.14 ± 0.18 † | 101.9 ± 1.4 † | 13.2 ± 0.8 † |
2.7. Effect of Total Flavonoids on Right Ventricle Morphology

3. Discussion
4. Experimental
4.1. Reagents
4.2. Experimental Procedure
4.3. Hemodynamic Measurements
4.4. Hematologic Measurements
4.5. Determination of Heart Weight
4.6. Histology
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
- Samples Availability: Samples of the compounds are available from the authors.
References
- Moore, L.G. Human genetic adaptation to high altitude. High Alt. Med. Biol. 2001, 2, 257–279. [Google Scholar] [CrossRef]
- Monge, C.C.; Whittembury, J. Chronic mountain sickness. Johns Hopkins Med. J. 1976, 139, 87–89. [Google Scholar]
- Windsor, J.S.; Rodway, G.W. Heights and haematology: The story of haemoglobin at altitude. Postgrad. Med. J. 2007, 83, 148–151. [Google Scholar] [CrossRef]
- Jefferson, J.A.; Escudero, E.; Hurtado, M.E.; Kelly, J.P.; Swenson, E.R.; Wener, M.H.; Burnier, M.; Maillard, M.; Schreiner, G.F.; Schoene, R.B.; et al. Hyperuricemia, hypertension, and proteinuria associated with high-altitude polycythemia. Am. J. Kidney Dis. 2002, 39, 1135–1142. [Google Scholar] [CrossRef]
- Koury, M.J. Erythropoietin: The story of hypoxia and a finely regulated hematopoietic hormone. Exp. Hematol. 2005, 33, 1263–1270. [Google Scholar] [CrossRef]
- Fang, J.; Menon, M.; Kapelle, W.; Bogacheva, O.; Bogachev, O.; Houde, E.; Browne, S.; Sathyanarayana, P.; Wojchowski, D.M. EPO modulation of cell-cycle regulatory genes, and cell division, in primary bone marrow erythroblasts. Blood 2007, 110, 2361–2370. [Google Scholar] [CrossRef]
- Gore, C.J.; Rodriguez, F.A.; Truijens, M.J.; Townsend, N.E.; Stray-Gundersen, J.; Levine, B.D. Increased serum erythropoietin but not red cell production after 4 wk of intermittent hypobaric hypoxia (4,000–5,500 m). J. Appl. Physiol. 2006, 101, 1386–1393. [Google Scholar] [CrossRef]
- Simonson, T.S.; Yang, Y.; Huff, C.D.; Yun, H.; Qin, G.; Witherspoon, D.J.; Bai, Z.; Lorenzo, F.R.; Xing, J.; Jorde, L.B.; et al. Genetic evidence for high-altitude adaptation in Tibet. Science 2010, 329, 72–75. [Google Scholar]
- Yi, X.; Liang, Y.; Huerta-Sanchez, E.; Jin, X.; Cuo, Z.X.; Pool, J.E.; Xu, X.; Jiang, H.; Vinckenbosch, N.; Korneliussen, T.S.; et al. Sequencing of 50 human exomes reveals adaptation to high altitude. Science 2010, 329, 75–78. [Google Scholar]
- Rivera-Ch, M.; Leon-Velarde, F.; Huicho, L. Treatment of chronic mountain sickness: Critical reappraisal of an old problem. Respir. Physiol. Neurobiol. 2007, 158, 251–265. [Google Scholar] [CrossRef]
- Richalet, J.P.; Rivera-Ch, M.; Maignan, M.; Privat, C.; Pham, I.; Macarlupu, J.L.; Petitjean, O.; Leon-Velarde, F. Acetazolamide for Monge’s disease: Efficiency and tolerance of 6-month treatment. Am. J. Respir. Crit. Care Med. 2008, 177, 1370–1376. [Google Scholar] [CrossRef]
- Purushothaman, J.; Suryakumar, G.; Shukla, D.; Malhotra, A.S.; Kasiganesan, H.; Kumar, R.; Sawhney, R.C.; Chami, A. Modulatory effects of seabuckthorn (Hippophae rhamnoides L.) in hypobaric hypoxia induced cerebral vascular injury. Brain Res. Bull. 2008, 77, 246–252. [Google Scholar]
- Purushothaman, J.; Suryakumar, G.; Shukla, D.; Jayamurthy, H.; Kasiganesan, H.; Kumar, R.; Sawhney, R.C. Modulation of hypoxia-induced pulmonary vascular leakage in rats by seabuckthorn (Hippophae rhamnoides L.). Evid. Based Complement. Alternat. Med. 2009. [Google Scholar] [CrossRef]
- Taysi, S.; Gumustekin, K.; Demircan, B.; Aktas, O.; Oztasan, N.; Akcay, F.; Suleyman, H.; Akar, S.; Dane, S.; Gul, M. Hippophae rhamnoides attenuates nicotine-induced oxidative stress in rat liver. Pharm. Biol. 2010, 48, 488–493. [Google Scholar] [CrossRef]
- Upadhyay, N.K.; Kumar, R.; Siddiqui, M.S.; Gupta, A. Mechanism of wound-healing activity of Hippophae rhamnoides L. leaf extract in experimental burns. Evid. Based Complement. Alternat. Med. 2009. [Google Scholar] [CrossRef]
- Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cooper, E.L.; Cossarizza, A. Quercetin and cancer chemoprevention. Evid. Based Complement. Alternat. Med. 2011. [Google Scholar] [CrossRef]
- Zhuang, X.; Zhang, W.; Pang, X.; Wu, W.; Qu, W. Combined effect of total flavonoids from seed residues of Hippophae rhamnoides L. and zinc on advanced glycation end products-induced endothelial cell dysfunction. Food Chem. 2012, 133, 905–911. [Google Scholar]
- Guliyev, V.B.; Gul, M.; Yildirim, A. Hippophae rhamnoides L.: Chromatographic methods to determine chemical composition, use in traditional medicine and pharmacological effects. J. Chromatogr. B 2004, 812, 291–307. [Google Scholar]
- Chen, C.; Zhang, H.; Xiao, W.; Yong, Z.P.; Bai, N. High-performance liquid chromatographic fingerprint analysis for different origins of sea buckthorn berries. J. Chromatogr. A 1154, 250–259. [Google Scholar]
- Zhang, Q.; Cui, H. Simultaneous determination of quercetin, kaempferol, and isorhamnetin in phytopharmaceuticals of Hippophae rhamnoides L. by high-performance liquid chromatography with chemiluminescence detection. J. Sep. Sci. 2005, 28, 1171–1178. [Google Scholar]
- Zu, Y.; Li, C.; Fu, Y.; Zhao, C. Simultaneous determination of catechin, rutin, quercetin kaempferol and isorhamnetin in the extract of sea buckthorn (Hippophae rhamnoides L.) leaves by RP-HPLC with DAD. J. Pharm. Biomed. Anal. 2006, 41, 714–719. [Google Scholar]
- Zhou, J.; Zhou, S.; Gao, Y.; Zeng, S. Modulatory effects of quercetin on hypobaric hypoxic rats. Eur. J. Pharmacol. 2012, 674, 450–454. [Google Scholar] [CrossRef]
- Kelly, G.S. Quercetin. Monograph. Altern. Med. Rev. 2011, 16, 172–194. [Google Scholar]
- Wang, J.; Zhang, W.; Zhu, D.; Zhu, X.; Pang, X.; Qu, W. Hypolipidaemic and hypoglycaemic effects of total flavonoids from seed residues of Hippophae rhamnoides L. in mice fed a high-fat diet. J. Sci. Food Agric. 2011, 91, 1446–1451. [Google Scholar]
- Pang, X.; Zhao, J.; Zhang, W.; Zhuang, X.; Wang, J.; Xu, R.; Xu, Z.; Qu, W. Antihypertensive effect of total flavones extracted from seed residues of Hippophae rhamnoides L. in sucrose-fed rats. J. Ethnopharmacol. 2008, 117, 325–331. [Google Scholar]
- Wang, G.L.; Semenza, G.L. Characterization of hypoxia-inducible factor 1 and regulation of DNA binding activity by hypoxia. J. Biol. Chem. 1993, 268, 21513–21518. [Google Scholar]
- Ou, L.C.; Cai, Y.N.; Tenney, S.M. Responses of blood volume and red cell mass in two strains of rats acclimatized to high altitude. Respir. Physiol. 1985, 62, 85–94. [Google Scholar] [CrossRef]
- West, J.B. Pulmonary Pathophysiology: The Essentials; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2008. [Google Scholar]
- Ou, L.C.; Salceda, S.; Schuster, S.J.; Dunnack, L.M.; Brink-Johnsen, T.; Chen, J.; Leiter, J.C. Polycythemic responses to hypoxia: Molecular and genetic mechanisms of chronic mountain sickness. J. Appl. Physiol. 1998, 84, 1242–1251. [Google Scholar]
- Ou, L.C.; Smith, R.P. Probable strain differences of rats in susceptibilities and cardiopulmonary responses to chronic hypoxia. Respir. Physiol. 1983, 53, 367–377. [Google Scholar] [CrossRef]
- Ou, L.C.; Chen, J.; Fiore, E.; Leiter, J.C.; Brinck-Johnsen, T.; Birchard, G.F.; Clemons, G.; Smith, R.P. Ventilatory and hematopoietic responses to chronic hypoxia in two rat strains. J. Appl. Physiol. 1992, 72, 2354–2363. [Google Scholar]
- Hannon, J.P.; Klain, G.J.; Sudman, D.M.; Sullivan, F.J. Nutritional aspects of high-altitude exposure in women. Am. J. Clin. Nutr. 1976, 29, 604–613. [Google Scholar]
- Hoyt, R.W.; Honig, A. Body fluid and energy metabolism at high altitude. Compr. Physiol. 2011. [CrossRef]
- Erslev, A.J.; Caro, J.; Miller, O.; Silver, R. Plasma erythropoietin in health and disease. Ann. Clin. Lab. Sci. 1980, 10, 250–257. [Google Scholar]
- Kurtz, A.; Eckardt, K.E.; Tannahill, L.; Bauer, C. Regulation of erythropoietin production. Contrib. Nephrol. 1988, 66, 1–16. [Google Scholar]
- Eckardt, K.U.; Dittmer, J.; Neumann, R.; Bauer, C.; Kurtz, A. Decline of erythropoietin formation at continuous hypoxia is not due to feedback inhibition. Am. J. Physiol. 1990, 258, 1432–1437. [Google Scholar]
- Smith, R.P.; Kruszyna, R.; Ou, L.C. Hemoglobinemia in rats exposed to high altitude is not due to an overload of catabolic mechanisms. Aviat. Space Environ. Med. 1979, 50, 9–13. [Google Scholar]
- Mulder, P.; Boujedaini, H.; Richard, V.; Henry, J.P.; Renet, S.; Munter, K.; Thuillez, C. Long-term survival and hemodynamics after endothelin-a receptor antagonism and angiotensin-converting enzyme inhibition in rats with chronic heart failure: Monotherapy versus combination therapy. Circulation 2002, 106, 1159–1164. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, Y.; Luo, Y.; Niu, W.; Li, Z.C. Alteration of serotonin 2C receptor expression in the aorta and the pulmonary artery in rats exposed to hypoxia. Chin. J. Physiol. 2008, 51, 338–347. [Google Scholar]
- Predictive analytics software. version 13.0, 2006.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhou, J.-Y.; Zhou, S.-W.; Du, X.-H.; Zeng, S.-Y. Protective Effect of Total Flavonoids of Seabuckthorn (Hippophae rhamnoides) in Simulated High-Altitude Polycythemia in Rats. Molecules 2012, 17, 11585-11597. https://doi.org/10.3390/molecules171011585
Zhou J-Y, Zhou S-W, Du X-H, Zeng S-Y. Protective Effect of Total Flavonoids of Seabuckthorn (Hippophae rhamnoides) in Simulated High-Altitude Polycythemia in Rats. Molecules. 2012; 17(10):11585-11597. https://doi.org/10.3390/molecules171011585
Chicago/Turabian StyleZhou, Ji-Yin, Shi-Wen Zhou, Xiao-Huang Du, and Sheng-Ya Zeng. 2012. "Protective Effect of Total Flavonoids of Seabuckthorn (Hippophae rhamnoides) in Simulated High-Altitude Polycythemia in Rats" Molecules 17, no. 10: 11585-11597. https://doi.org/10.3390/molecules171011585