Detection of Cytotoxic Activity of Lectin on Human Colon Adenocarcinoma (Sw480) and Epithelial Cervical Carcinoma (C33-A)


 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Tepary bean lectin purification

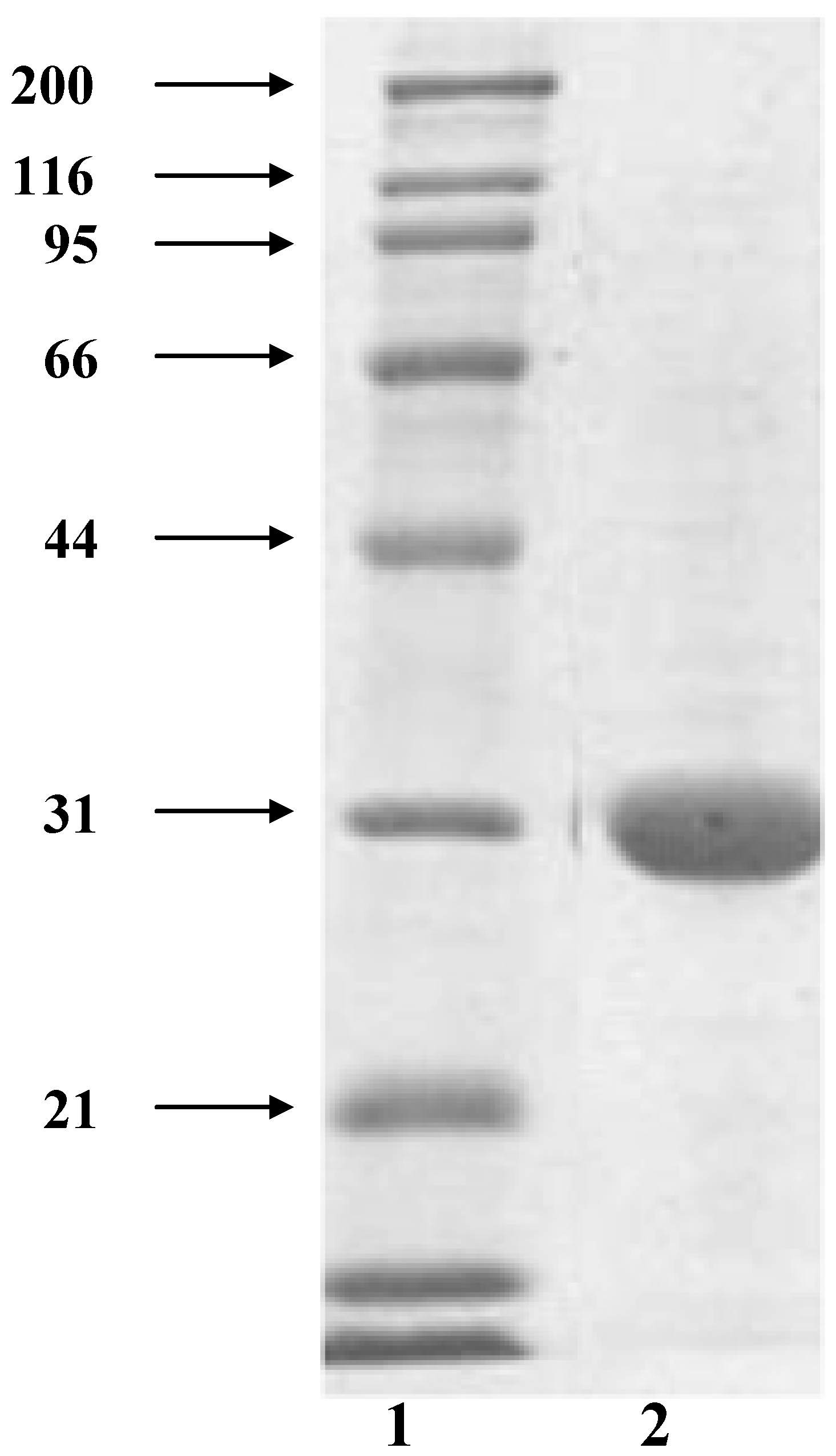{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin | Lectin Concentration (mg/mL) | Human erythrocytes | |
|---|---|---|---|
| Type A | Type O | ||
| Tepary | 3.5 | 292.6 ± 32.9 | 36.6 ± 4.3 |
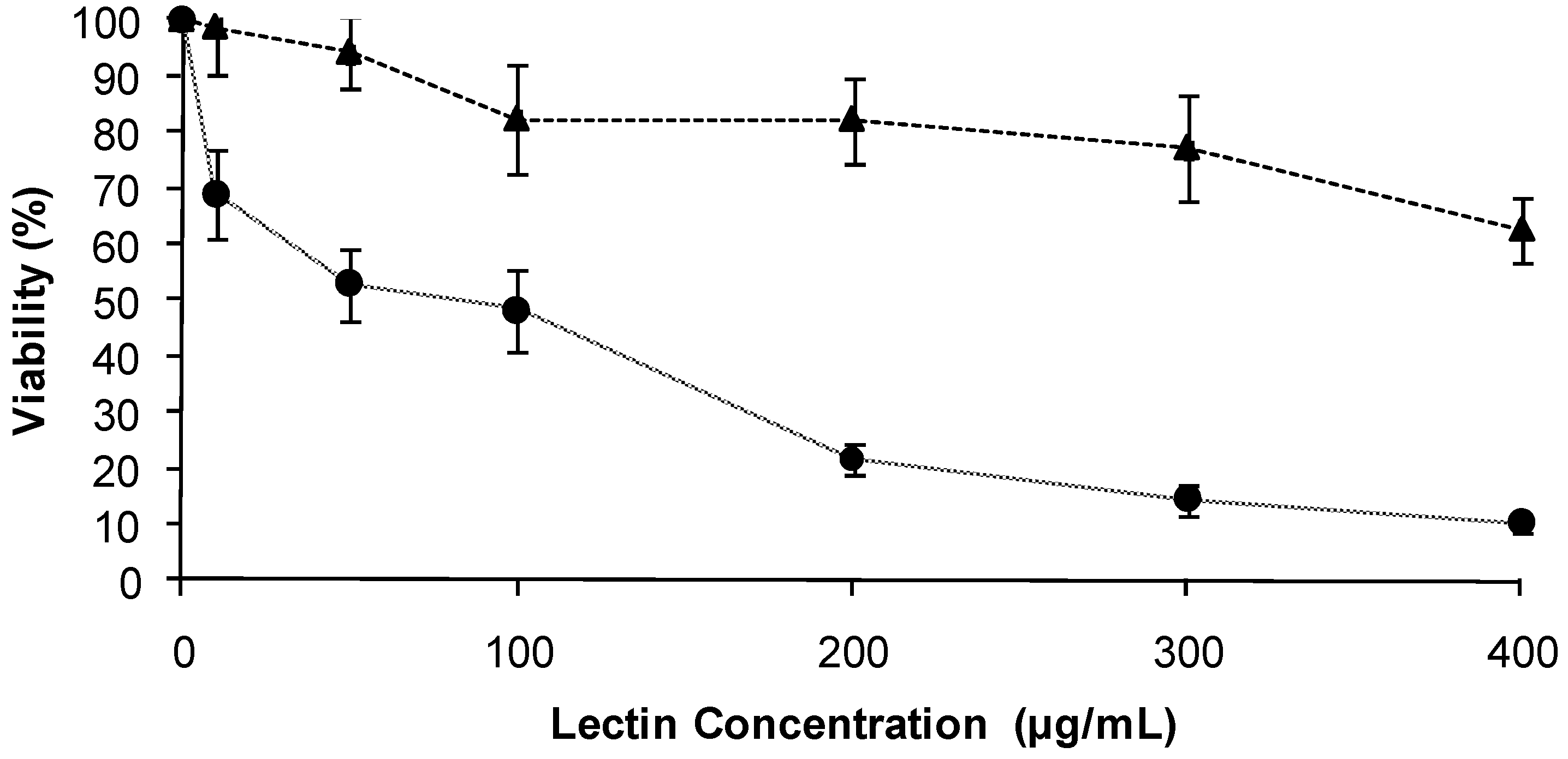
2.2. Effect of the TBL on the viability of human malignant cells

 SW480
SW480  .
SW480 .
.
SW480 .
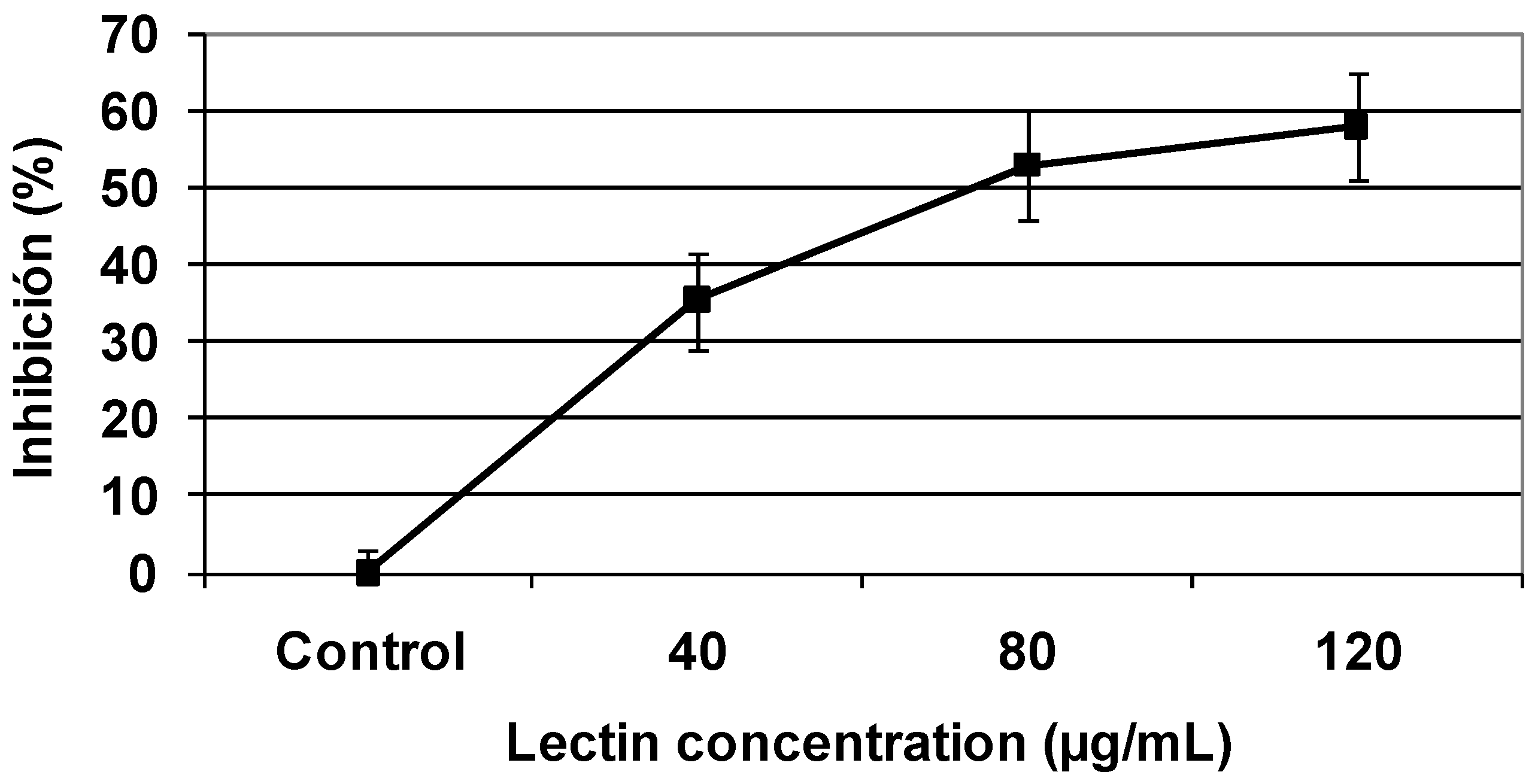
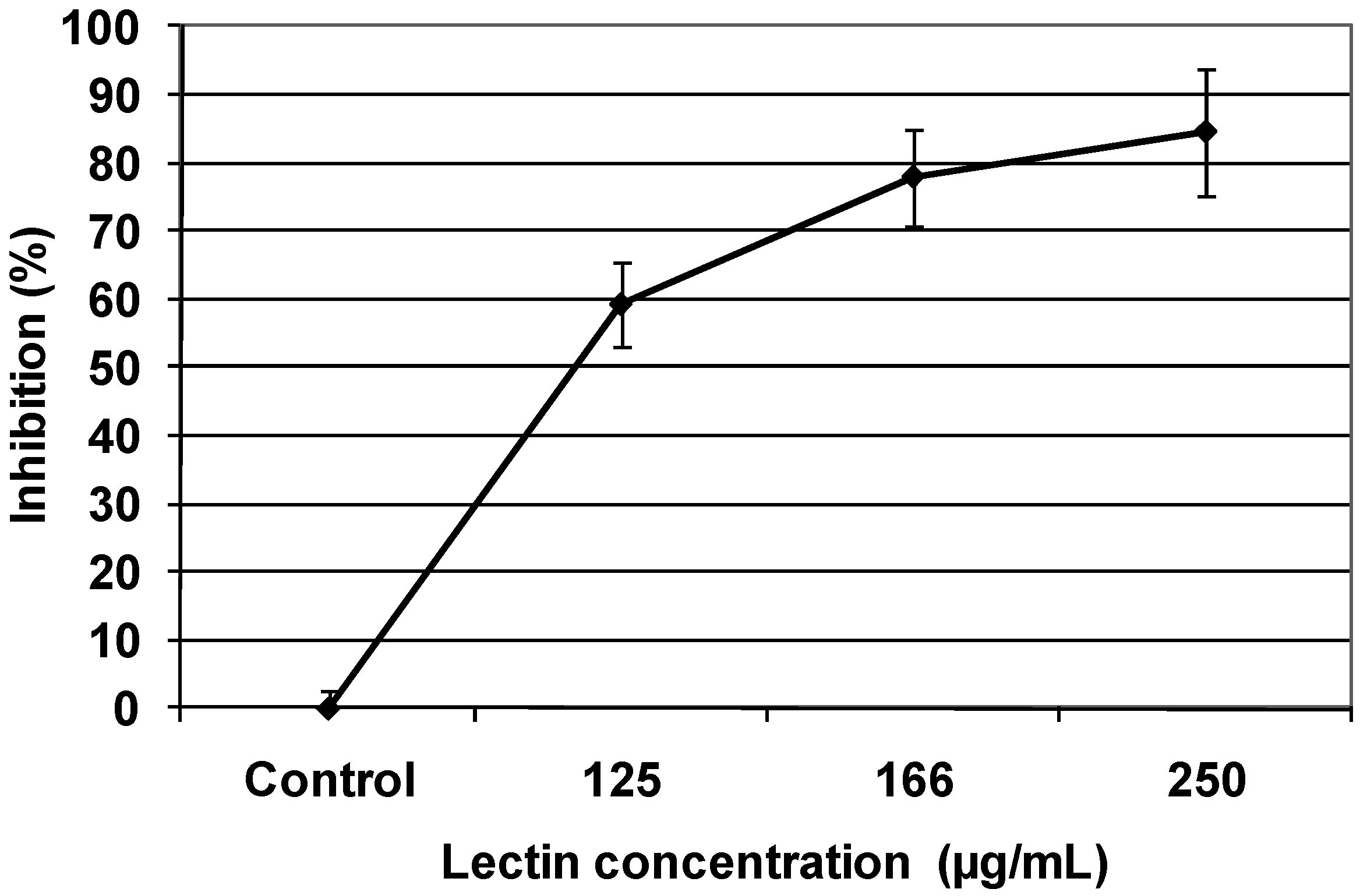
2.3. Cytotoxic effect of TBL in human malignant cells
| Treatment | DNA (% incorporation 3[H] thymidine) | |
|---|---|---|
| SW480 | C33-A | |
| Control | 100 ± 10.56 | 100 ± 11.06 |
| 10 µg/mL | 71.3 ± 5.93 | 42.6 ± 11.35 |
| 25 µg/mL | 66.7 ± 6.19 | 23.6 ± 12.08 |
| 50 µg/mL | 59.7 ± 5.43 | 21.2 ± 13.47 |
| 100 µg/mL | 33.4 ± 18.77 | 43.1 ± 4.96 |
2.4. Colony formation


2.5. Discussion
3. Experimental
3.1. Extraction and purification of lectins
3.2. Quantification of protein
3.3. Hemagglutination assays
3.4. Polyacrilamide gel electrophoresis (PAGE)
3.5. Cell culture
3.6. Cell viability assay
3.7. Cytotoxicity assay
3.8. Colony formation assay
4. Conclusions
References
- Gazdar, A.F.; Girard, L.; Lockwood, W.W.; Lam, W.L.; Minna, J.D. Lung Cancer Cell Lines as Tools for Biomedical Discovery and Research. J. Natl. Cancer Inst. 2010, 102, 1310–1321. [Google Scholar]
- Issah, F.; Maree, J.E.; Mwinituo, P.P. Expressions of cervical cancer-related signs and symptoms. Eur. J. Oncol. Nurs. 2010, in press. [Google Scholar]
- Pramateftakis, M.G.; Kanellos, D.; Vrakas, G.; Tsachalis, T.; Raptis, D.; Makrantonakis, A.; Koukouritaki, Z.; Kanellos, I. Progress in rectal cancer staging and treatment. Tech. Coloproctol. 2010, 14, 529–531. [Google Scholar]
- Bishayee, A.; Háznagy-Radnai, E.; Mbimba, T.; Sipos, P.; Morazzoni, P.; Darvesh, A.S.; Bhatia, D.; Hohmann, J. Anthocyanin-rich black currant extract suppresses the growth of human hepatocellular carcinoma cells. Nat. Prod. Commun. 2010, 5, 1613–1618. [Google Scholar]
- Qiu, W.; Wang, X.; Leibowitz, B.; Liu, H.; Barker, N.; Okada, H.; Oue, N.; Yasui, W.; Clevers, H.; Schoen, R.E.; Yu, J.; Zhang, L. Chemoprevention by nonsteroidal anti-inflammatory drugs eliminates oncogenic intestinal stem cells via SMAC-dependent apoptosis. Proc. Natl. Acad. Sci. USA 2010, 107, 20027–20032. [Google Scholar]
- Dragsted, L.; Srube, M.; Larsen, J. Cancer-protective factors in fruits and vegetables: Biochemical and biological background. Pharmacol.Toxicol. 1993, 72, 116–135. [Google Scholar]
- Pezzuto, J. Plant-derived anticancer agents. Biochem. Pharmacol. 1996, 53, 121–133. [Google Scholar]
- Abdullaev, F.I.; Frenkel, G.D. Effect of saffron on cell colony formation and cellular nucleic acid and protein synthesis. BioFactors 1992, 3, 201–204. [Google Scholar]
- Abdullaev, F.I.; MacVicar, C.; Frenkel, G.D. Inhibition by selenium of DNA and RNA synthesis in normal and malignant human cells in vitro. Cancer Lett. 1992, 65, 43–49. [Google Scholar] [CrossRef]
- Abdullaev, F.I.; Rivera-Luna, R.; García-Carranca, A.; Ayala-Fierro, F.; Espinosa-Aguirre, J.J. Cytotoxic effect of three arsenic compounds in HeLa human tumor and bacterial cells. Mutant Res. 2001, 493, 31–38. [Google Scholar]
- Pustai, A.J. Plat Lectins; Cambridge University Press: Cambridge, UK, 1991; p. 253. [Google Scholar]
- Drikarmer, K.; Taylor, M.E. biology of animal lectins. Annu. Rev. Cell. Biol. 1993, 9, 237–264. [Google Scholar]
- Gabius, H.J. Animal lectins. Eur. J. Biochem. 1997, 243, 543–576. [Google Scholar]
- Wong, J.H.; Ng, T.B.; Cheung, R.C.; Ye, X.J.; Wang, H.X.; Lam, S.K.; Lin, P.; Chan, Y.S.; Fang, E.F.; Ngai, P.H.; Xia, L.X.; Ye, X.Y.; Jiang, Y.; Liu, F. Proteins with antifungal properties and other medicinal applications from plants and mushrooms. Appl. Microbiol. Biotechnol. 2010, 87, 1221–1235. [Google Scholar] [CrossRef]
- Sharon, N.; Lis, H. Lectin-proteins with a sweet tooth: Functions in cell recognition. Assays Biochem. 1995, 30, 59–75. [Google Scholar]
- Goldstein, I.J.; Hughes, R.S.; Monsigny, M.; Osawa, T.; Sharon, N. What should be called a lectin? Nature 1980, 285, 665–666. [Google Scholar]
- Sabová, L.; Pilátová, M.; Szilagyi, K.; Sabo, R.; Mojzis, J. Cytotoxic effect of mistletoe (Viscum album L.) extract on Jurkat cells and its interaction with doxorubicin. Phytother. Res. 2010, 24, 365–368. [Google Scholar] [CrossRef]
- Li, C.Y.; Xu, H.L.; Liu, B.; Bao, J.K. Concanavalin A, from an old protein to novel candidate anti-neoplastic drug. Curr. Mol. Pharmacol. 2010, 3, 123–128. [Google Scholar]
- Matsuda, A.; Kuno, A.; Kawamoto, T.; Matsuzaki, H.; Irimura, T.; Ikehara, Y.; Zen, Y.; Nakanuma, Y.; Yamamoto, M.; Ohkohchi, N.; Shoda, J.; Hirabayashi, J.; Narimatsu, H. Wisteria floribunda agglutinin-positive mucin 1 is a sensitive biliary marker for human cholangiocarcinoma. Hepatology 2010, 52, 174–182. [Google Scholar] [CrossRef]
- Kaur, M.; Singh, K.; Rup, P.J.; Saxena, A.K.; Khan, R.H.; Ashraf, M.T.; Kamboj, S.S.; Singh, J. A tuber lectin from Arisaemahelleborifolium Schott with anti-insect activity against melon fruit fly, Bactroceracucurbitae (Coquillett) and anti-cancer effect on human cancer cell lines. Arch. Biochem. Biophys. 2006, 445, 156–165. [Google Scholar] [CrossRef]
- Blonski, K.; Schumacher, U.; Burkholder, I.; Edler, L.; Nikbakht, H.; Boeters, I.; Peters, A.; Kugler, C.; Horny, H.P.; Langer, M.; Wilhelm-Ogunbiyi, K.; Witthohn, K.; Laack, E. Binding of recombinant mistletoe lectin (aviscumine) to resected human adenocarcinoma of the lung. Anticancer Res. 2005, 25, 3303–3307. [Google Scholar]
- Nozoe, T.; Korenaga, D.; Ohga, T.; Futatsugi, M.; Machara, Y. Suppression of the phytohemagglutinin response to lymphocytes is an independent prognosticator in patients with squamous cell carcinoma of the esophagus. Ann. Thorac. Surg. 2003, 76, 260–265. [Google Scholar] [CrossRef]
- Valentiner, U.; Ian, S.; Schumacher, U.; Leathem, A.J. the influence of dietary lectins on the cell proliferation of human breast cancer cell lines in vitro. Anticancer Res. 2003, 23, 1197–1206. [Google Scholar]
- Gastman, B.; Wang, K.; Han, J.; Zhu, Z.Y.; Huang, X.; Wang, G.Q.; Rabinowich, H.; Gorelik, E. A novel apoptotic pathway as defined by lectin cellular initiation. Biochem. Biophys. Res. Commun. 2004, 26, 263–271. [Google Scholar]
- Banchonglikitkul, C.; Smart, J.D.; Gibbs, R.V.; Donovan, S.J.; Cook, D.J. An in vitro evaluation of lectincytotoxicity using cell lines derived from the ocular surface. J. Drug. Target. 2002, 10, 601–606. [Google Scholar] [CrossRef]
- Janssen, O.; Scheffler, A.; Kabelitz, D. In vitro effects of mistletoe extracts and mistletoe lectins. Cytotoxicity towards tumor cells due to the induction of programmed cell death (apoptosis). Arzneimittelforschung 1993, 43, 1221–1227. [Google Scholar]
- Schwarz, R.E.; Wojciechowicz, D.C.; Picon, A.I.; Schwarz, M.Z.; Paty, P.B. Wheatgerm agglutinin-mediated toxicity in pancreatic cancer cells. J. Cancer 1999, 80, 1754–1762. [Google Scholar] [CrossRef]
- Kiss, R.; Camby, I.; Duckworth, C.; De Decker, R.; Salmon, I.; Pasteel, J.L.; Danguy, A.; Yeaton, P. In vitro influence of Phaseolusvulgaris, Griffoniasimpliciflia, oncanavalina A, Wheat germ, and peanut agglutinin on HCT-15, Lovo, d SW 837 human colorectal cancer cell growth. Gut 1997, 40, 253–261. [Google Scholar]
- Hammar, L.; Hirsch, I.; Machado, A.A.; De Mareuil, J.; Baillon, J.G.; Bolmont, C.; Chermann, J.C. Lectin-mediated effects on HIV type I infection in vitro. AIDS Res. Hum. Retrovir. 1995, 11, 87–95. [Google Scholar] [CrossRef]
- Ito, T.; Suzuki, Y.; Mitnaul, L.; Vines, A.; Kida, H.; Kawaoka, Y. Receptor specificity on influenza A viruses correlates with the agglutination of erythrocytes from different animal species. Virology 1997, 227, 493–499. [Google Scholar] [CrossRef]
- Efferth, T. Cancer therapy with natural products and medicinal plants. Planta Med. 2010, 76, 1035–1036. [Google Scholar]
- Gullett, N.P.; Ruhul Amin, A.R.; Bayraktar, S.; Pezzuto, J.M.; Shin, D.M.; Khuri, F.R.; Aggarwal, B.B.; Surh, Y.J.; Kucuk, O. Cancer Prevention with Natural Compounds. Semin. Oncol. 2010, 37, 258–281. [Google Scholar] [CrossRef]
- Ojeswi, B.K.; Khoobchandani, M.; Hazra, D.K.; Srivastava, M.M. Protective effect of Thujaoccidentalis against DMBA-induced breast cancer with reference to oxidative stress. Hum. Exp. Toxicol. 2010, 29, 369–375. [Google Scholar] [CrossRef]
- Tayarani-Najaran, Z.; Mousavi, S.H.; Asili, J.; Emami, S.A. Growth-inhibitory effect of Scutellarialindbergii in human cancer cell lines. Food. Chem. Toxicol. 2010, 48, 599–604. [Google Scholar]
- Zhou, X.X.; Ji, F.; Zhao, J.L.; Cheng, L.F.; Xu, C.F. Anti-cancer activity of anti-p185HER-2 ricin A chain immunotoxin on gastric cancer cells. J. Gastroenterol. Hepatol. 2010, 25, 1266–1275. [Google Scholar] [CrossRef]
- Behnckern, A.; Saeger, W. LectinBinding in pituitary Adenoma and Normal Pituitaries. Phat. Res. Pract. 1991, 187, 629–631. [Google Scholar] [CrossRef]
- Damjanou, I.; Sobrinho-Simoes, M. LectinHistochemistry of Human Thyroid Tumors. In Lectinsand Cancer; Gabius, S., Gabius, H.J., Eds.; Springer-Verlag: Berlin, Germany, 1991; pp. 41–52. [Google Scholar]
- Bardocz, S.; Grant, G.; Duguid, T.J.; Brown, D.S.; Pusztai, A.; Pryme, I.F. Effect of phytohaemoagglutinin on the growth of Krebs II tumour cells, body metabolism and internal organs of mice. Int. J. Oncol. 1994, 5, 1369–1374. [Google Scholar]
- Pryme, I.F.; Bardocz, S. Anti-cancer therapy: Diversion of polyamines in the gut. Eur. J. Gastroen. Hepatol. 2001, 13, 1041–1046. [Google Scholar] [CrossRef]
- Miller, J.B.; Noyes, C.; Heinrikson, R.; Kingdon, H.S.; Yachnin, S. Phytohemagglutinin mitogenic proteins. Structural evidence for a family of isomitogenic proteins. J. Exp. Med. 1973, 138, 939–951. [Google Scholar] [CrossRef]
- González de Mejía, E.; Grajeda Cota, P.; Celada, E.; Valencia, M.E. Characterization of the nutritional potential of tepari bean (Phaseolus acutifolius) grown in Mexico. Arch. Latinoam. Nutr. 1988, 38, 907–924. [Google Scholar]
- Lowry, O.H.; Rosebrough, J.N.; Fan, A.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Jaffe, W.G.; Brucher, O.; Palozzo, A. Detection of tour types of specific phytohemagglutinins in different lines of beans (Phaseolus vulgaris) Z. Immun. Forsh. Bd. 1972, 142, 439–447. [Google Scholar]
- Laemmli, V.K. Cleavage of structural protein during the assembly of the head of bacteriophage t4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Hansen, M.B.; Nielsen, S.E.; Berg, K. Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol. Methods 1989, 119, 203–210. [Google Scholar] [CrossRef]
- Abdullaev, F.I.; Gonzalez de Mejia, E. Inhibition of colony formation of Hela cells by naturally occurring and synthetic agents. Biofactors 1996, 5, 133–138. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Valadez-Vega, C.; Alvarez-Manilla, G.; Riverón-Negrete, L.; García-Carrancá, A.; Morales-González, J.A.; Zuñiga-Pérez, C.; Madrigal-Santillán, E.; Esquivel-Soto, J.; Esquivel-Chirino, C.; Villagómez-Ibarra, R.; et al. Detection of Cytotoxic Activity of Lectin on Human Colon Adenocarcinoma (Sw480) and Epithelial Cervical Carcinoma (C33-A). Molecules 2011, 16, 2107-2118. https://doi.org/10.3390/molecules16032107
Valadez-Vega C, Alvarez-Manilla G, Riverón-Negrete L, García-Carrancá A, Morales-González JA, Zuñiga-Pérez C, Madrigal-Santillán E, Esquivel-Soto J, Esquivel-Chirino C, Villagómez-Ibarra R, et al. Detection of Cytotoxic Activity of Lectin on Human Colon Adenocarcinoma (Sw480) and Epithelial Cervical Carcinoma (C33-A). Molecules. 2011; 16(3):2107-2118. https://doi.org/10.3390/molecules16032107
Chicago/Turabian StyleValadez-Vega, Carmen, Gerardo Alvarez-Manilla, Leticia Riverón-Negrete, Alejandro García-Carrancá, José A. Morales-González, Clara Zuñiga-Pérez, Eduardo Madrigal-Santillán, Jaime Esquivel-Soto, Cesar Esquivel-Chirino, Roberto Villagómez-Ibarra, and et al. 2011. "Detection of Cytotoxic Activity of Lectin on Human Colon Adenocarcinoma (Sw480) and Epithelial Cervical Carcinoma (C33-A)" Molecules 16, no. 3: 2107-2118. https://doi.org/10.3390/molecules16032107
APA StyleValadez-Vega, C., Alvarez-Manilla, G., Riverón-Negrete, L., García-Carrancá, A., Morales-González, J. A., Zuñiga-Pérez, C., Madrigal-Santillán, E., Esquivel-Soto, J., Esquivel-Chirino, C., Villagómez-Ibarra, R., Bautista, M., & Morales-González, Á. (2011). Detection of Cytotoxic Activity of Lectin on Human Colon Adenocarcinoma (Sw480) and Epithelial Cervical Carcinoma (C33-A). Molecules, 16(3), 2107-2118. https://doi.org/10.3390/molecules16032107