A Multivalent ICAM-1 Binding Nanoparticle which Inhibits ICAM-1 and LFA-1 Interaction Represents a New Tool for the Investigation of Autoimmune-Mediated Dry Eye

 ,
,
Abstract
:1. Introduction
2. Results
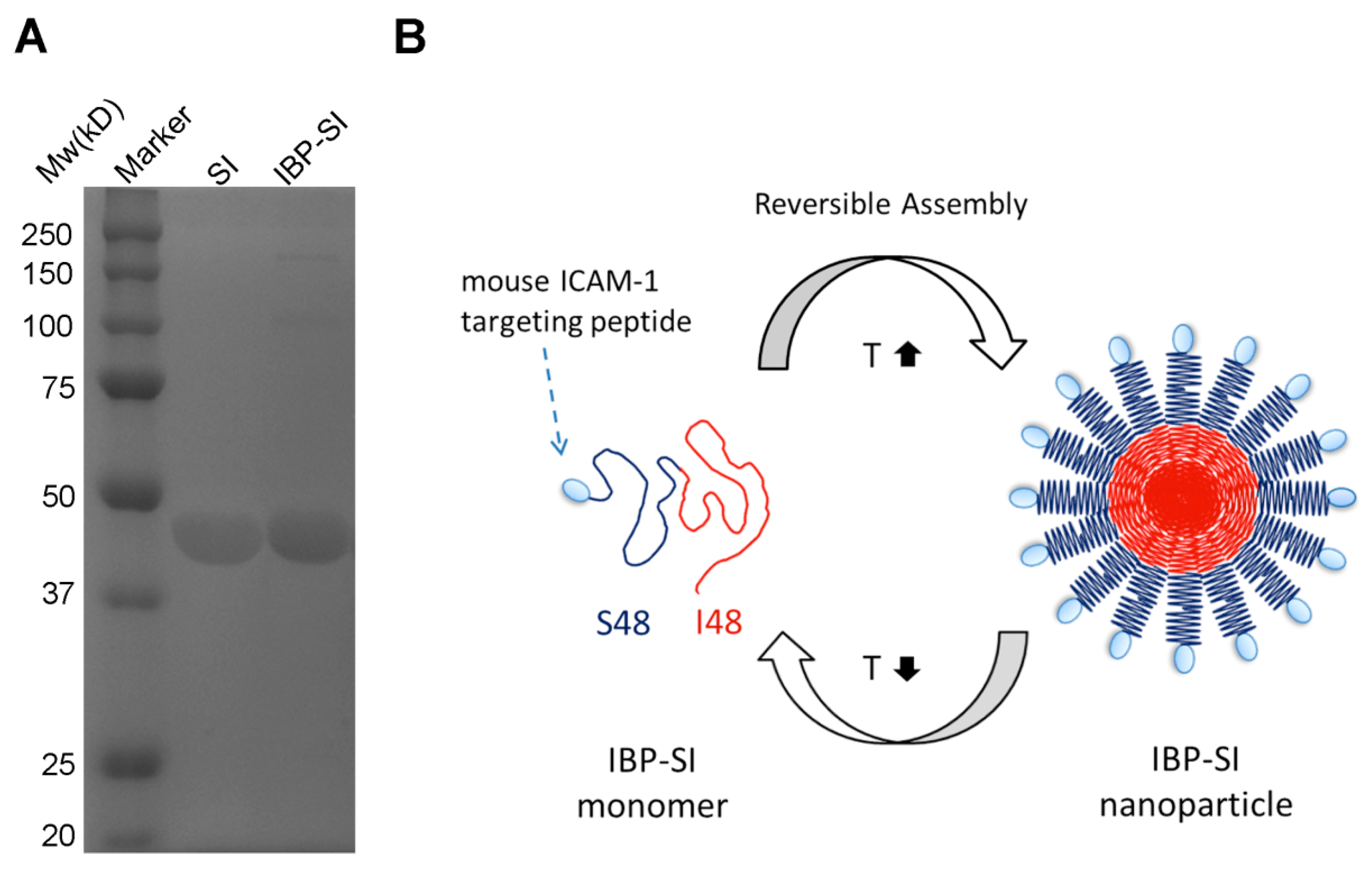
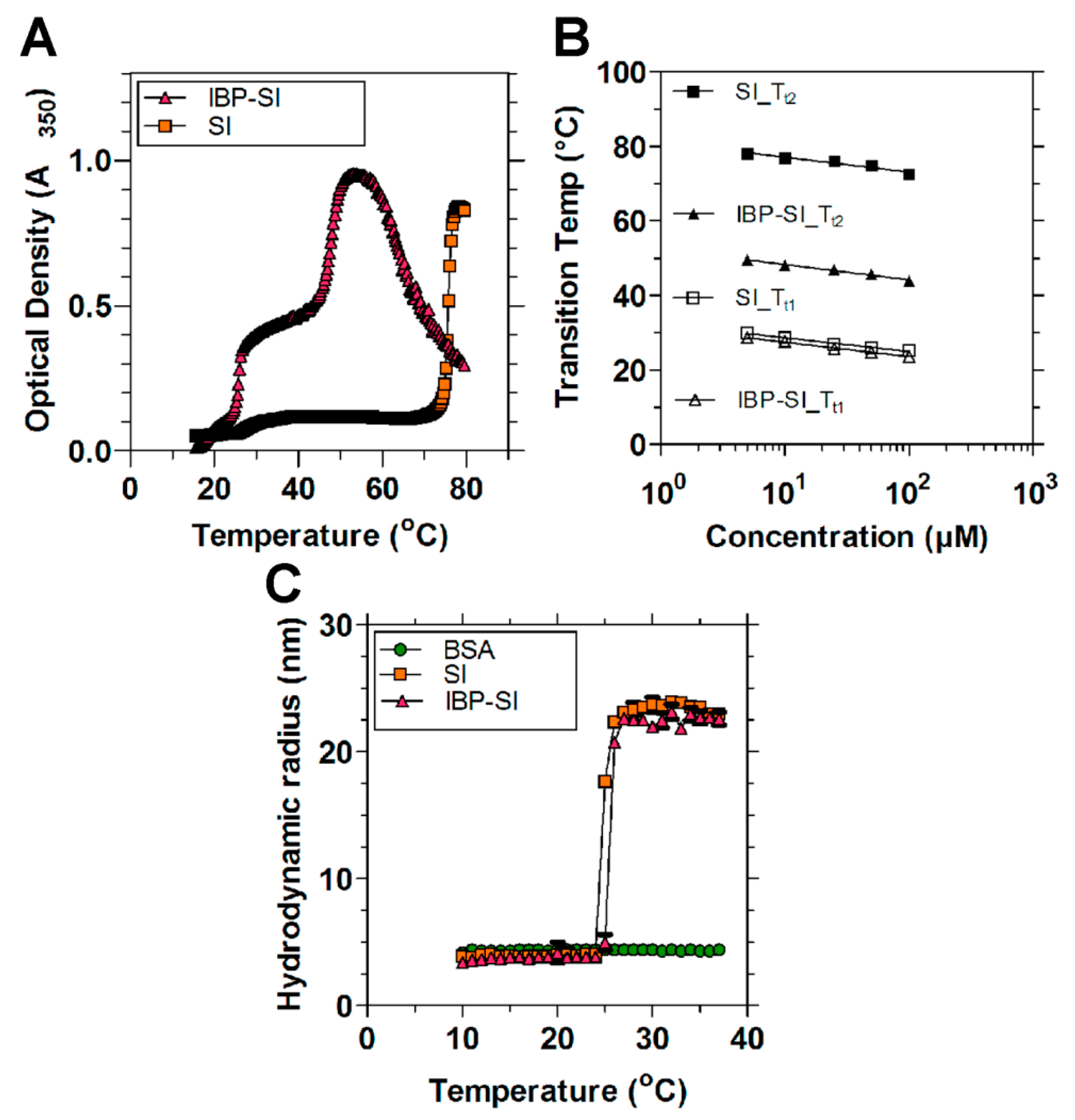
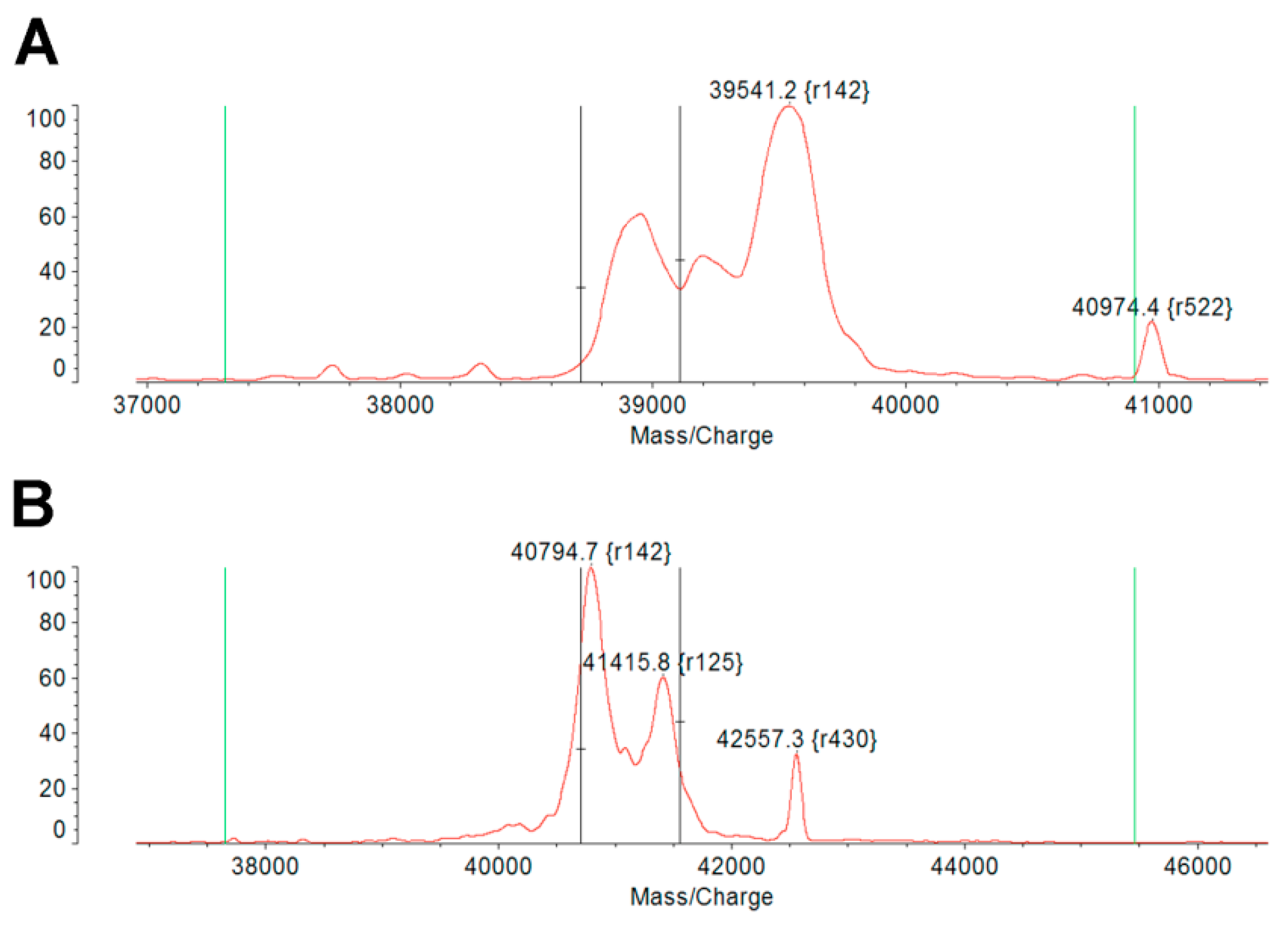
2.1. IBP-SI Forms Nanoparticles at Physiological Temperature
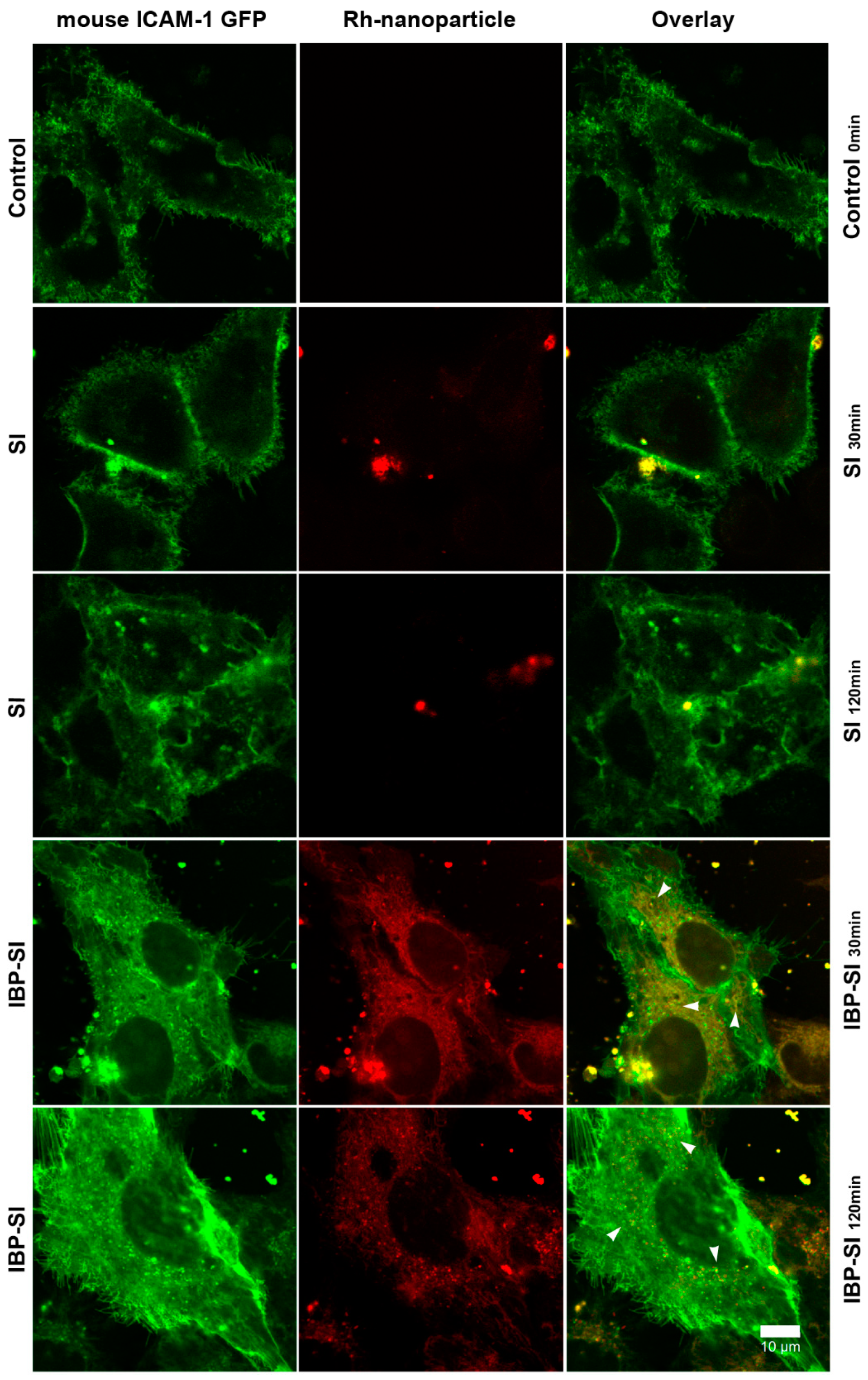
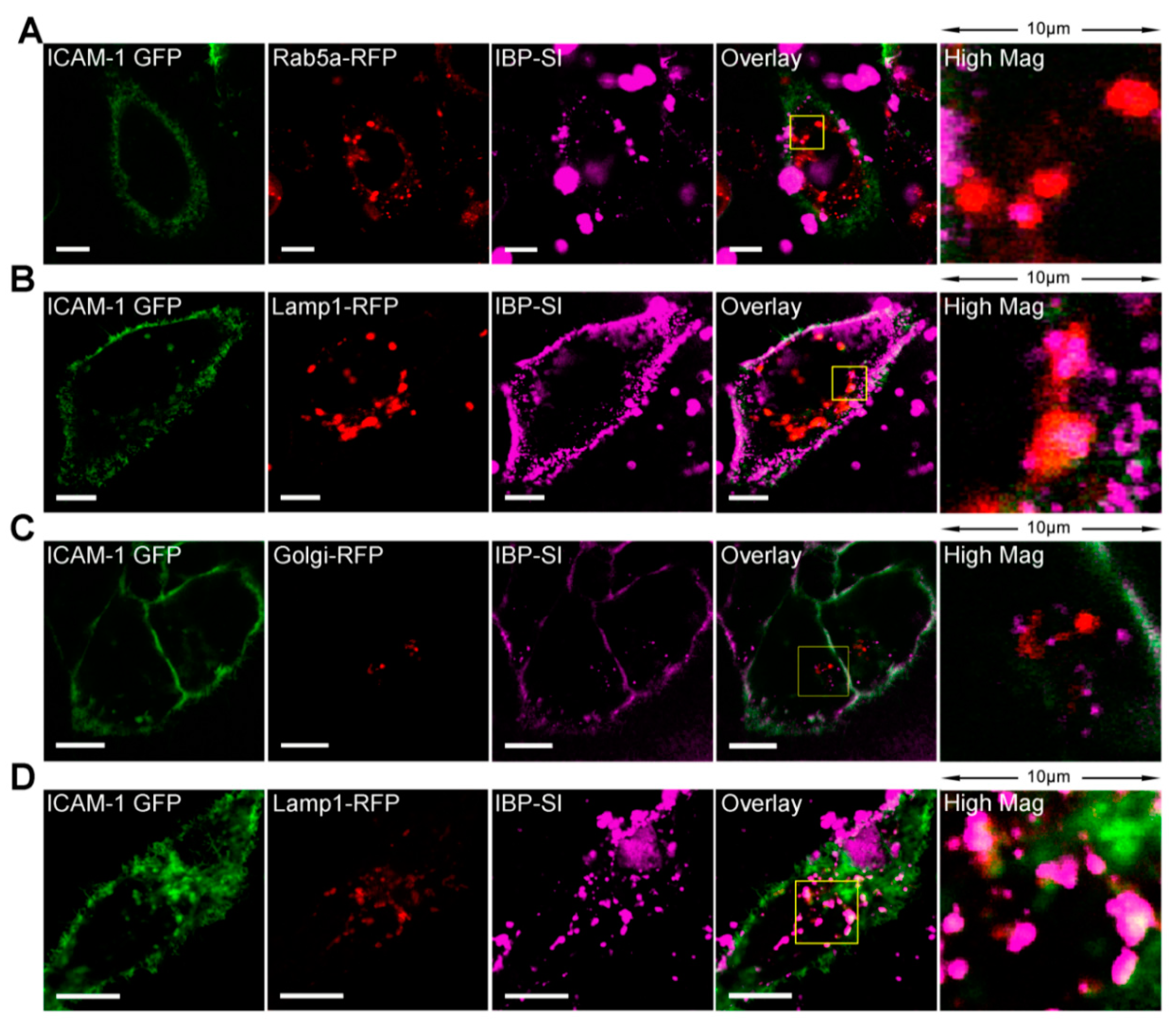
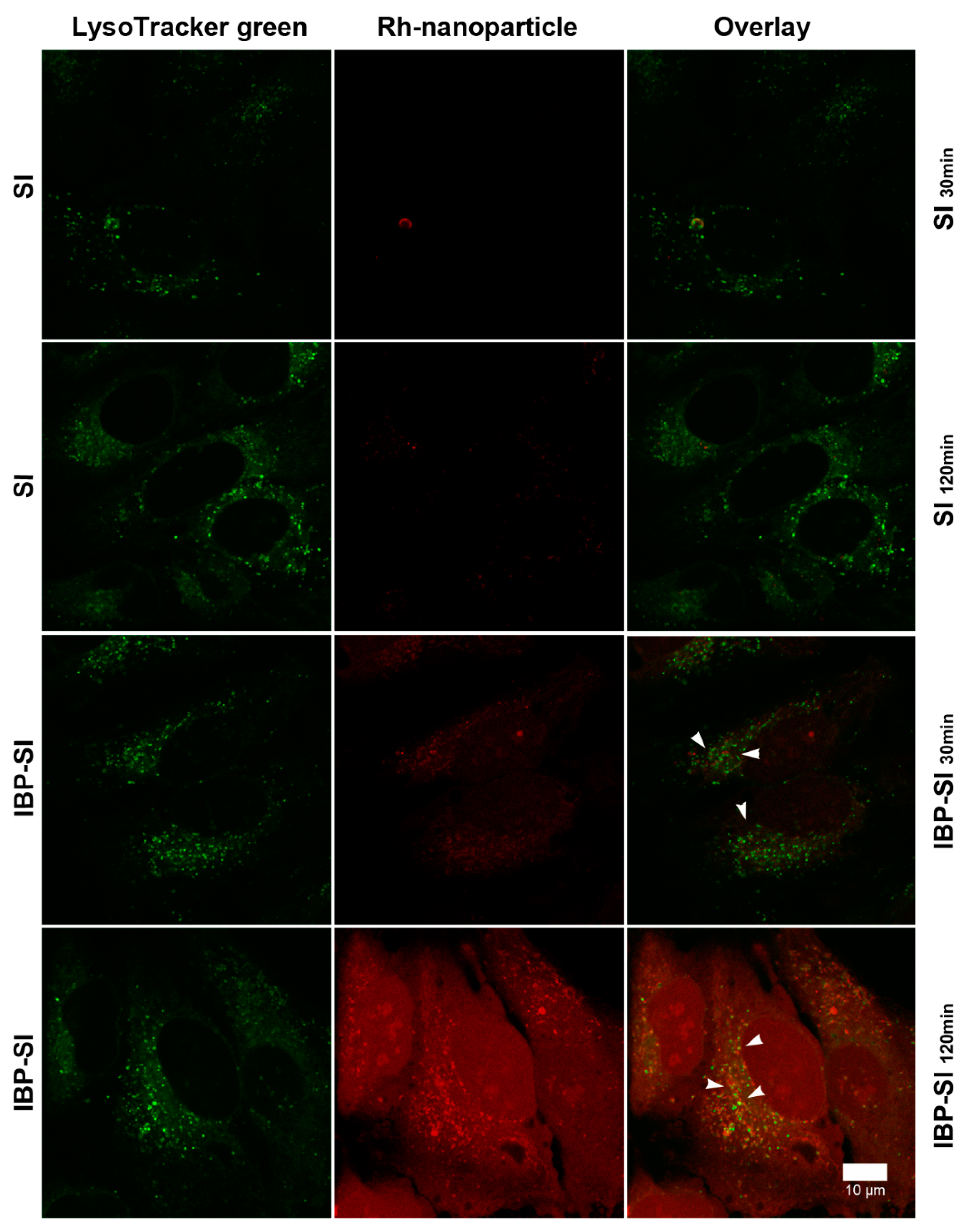
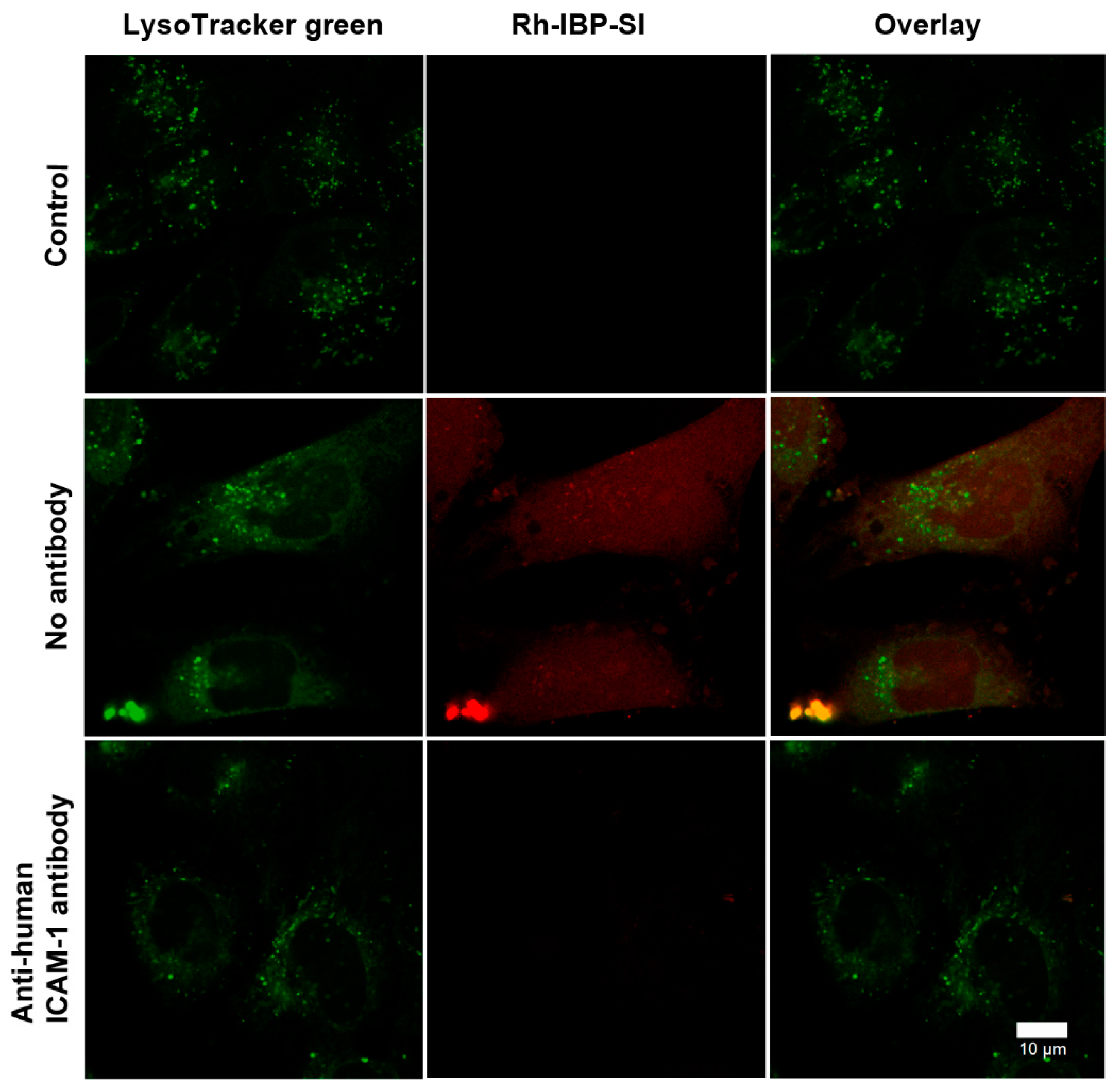
2.2. IBP-SI Nanoparticles Target ICAM-1 In Vitro
2.3. ICAM-1 Is Significantly Overexpressed in the LG of NOD Mice Compared with Healthy Mice
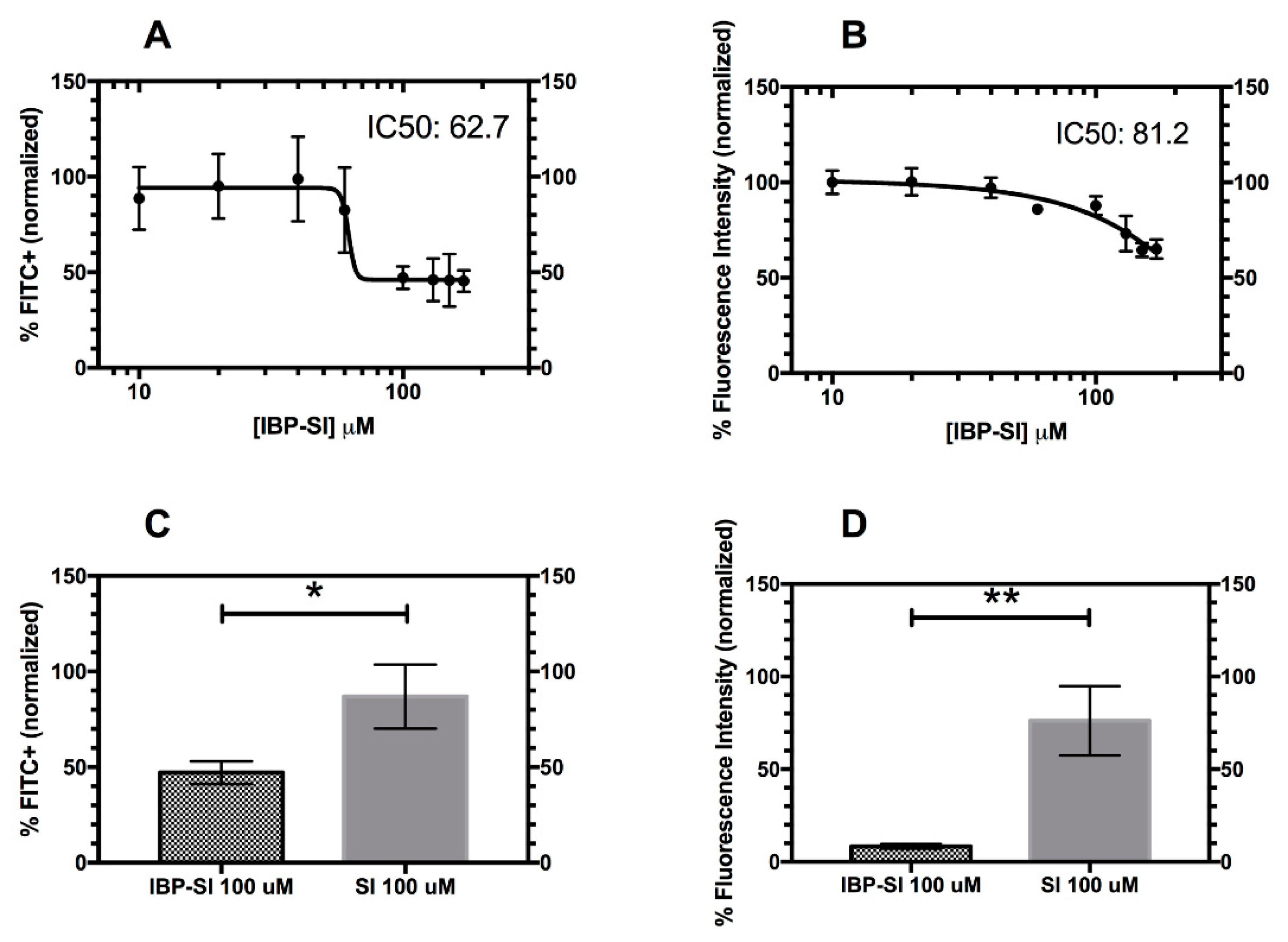
2.4. Inhibition of Lymphocyte Binding to ICAM-1 Enriched bEnd.3 Cells by IBP-SI
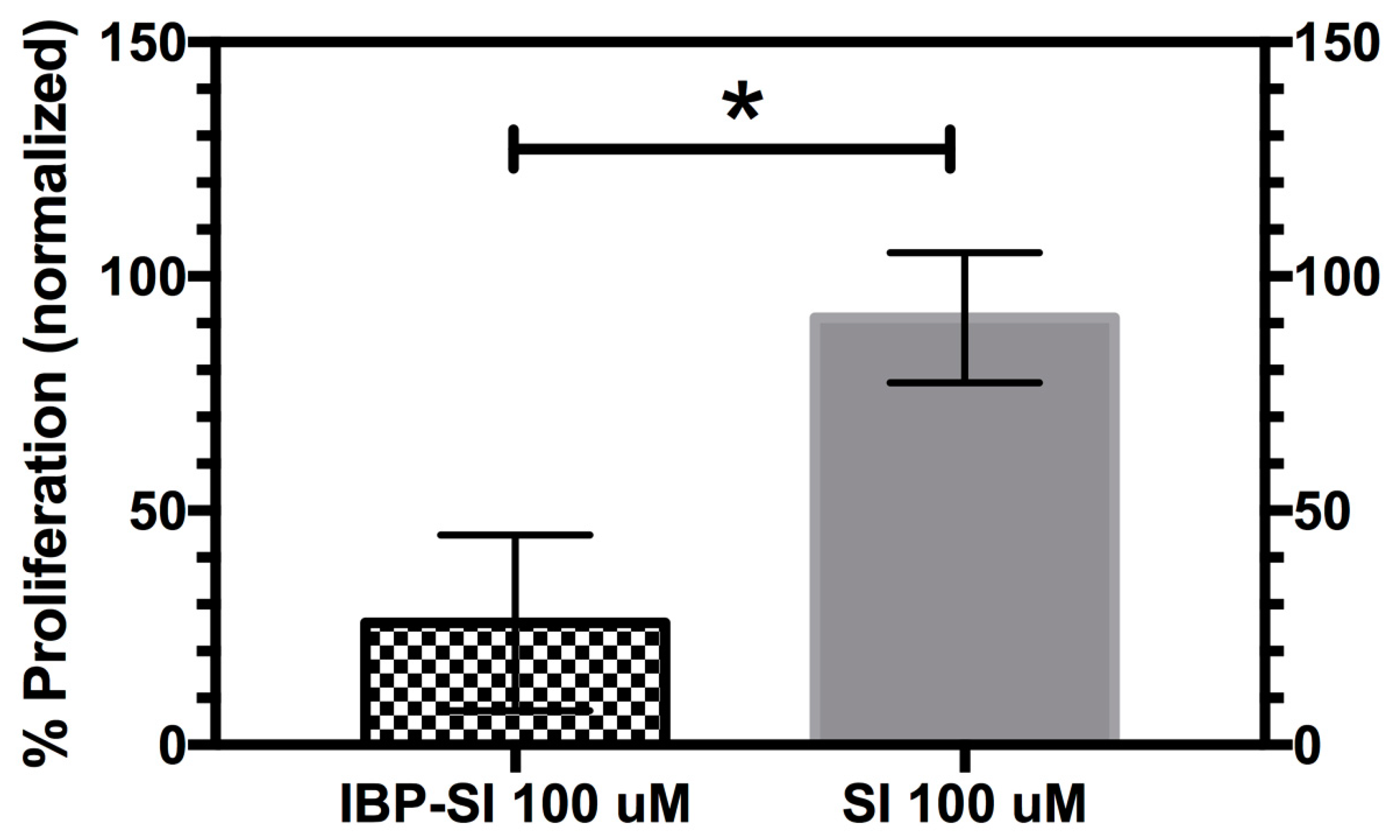
2.5. IBP-SI Inhibits the Activation and Proliferation of T-Cells
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Animals
4.3. Construction of IBP-SI ELP Nanoparticles
4.4. Characterization of IBP-SI Phase Transition Behavior and Nanoparticle Formation
4.5. Fluorescent Labelling of ELP Nanoparticles
4.6. In Vitro Cell Uptake
4.7. Immunofluorescence of Mouse LG
4.8. Splenocyte Isolation, Labelling and Quantitation
4.9. In Vitro Static Adhesion Assay
4.10. Antigen Presentation Assay
4.11. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATCC CFSE DAPI | American Type Culture Collection carboxyfluorescein diacetate succinimidyl ester 4′6-diamindino-2-phenylindole |
| DLS EDTA | dynamic light scattering ethylenediaminetetraacetic acid |
| ELP FACS Fc | elastin-like polypeptide fluorescence-activated cell sorting fragment crystallizable region |
| ICAM-1 | intercellular adhesion molecule-1 |
| IBP-SI IL-1 | ICAM-1 binding peptide S48I48 interleukin-1 |
| LFA-1 | lymphocyte function-associated antigen 1 |
| LG | lacrimal gland |
| LGAC | lacrimal gland acinar cell |
| MCC | moth cytochrome C |
| MEM MHC | Modified Eagle’s Medium major histocompatibility complex |
| mICAM-1 | mouse ICAM-1 |
| NOD PLGA | non-obese diabetic poly lactic-co-glycolic acid |
| PMA RFP | phorbol 12-myristate 13-acetate red fluorescent protein |
| rh RPMI SG | rhodamine roswell park memorial institute medium salivary gland |
| SI | S48I48 |
| SS TCR | Sjögrens’ syndrome T cell receptor |
| TNF-α | tumor necrosis factor-α |
Appendix A





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constructs | Temperature-Concentration Phase | Diagram |
|---|---|---|
| Slope, m (°CLog(µM)) | Intercept, b (°C) | |
| SI (Tt1) | −3.7 ± 0.3 | 32.4 ± 0.4 |
| SI (Tt2) | −4.0 ± 0.5 | 81.1 ± 0.8 |
| IBP-SI (Tt1) | −3.9 ± 0.1 | 31.4 ± 0.2 |
| IBP-SI (Tt2) | −4.1 ± 0.3 | 52.4 ± 0.4 |
References
- Mavragani, C.P.; Moutsopoulos, H.M. Sjogren’s syndrome. Annu. Rev. Pathol. 2014, 9, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Nocturne, G.; Mariette, X. Sjogren Syndrome-associated lymphomas: An update on pathogenesis and management. Br. J. Haematol. 2015, 168, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Nikolov, N.P.; Illei, G.G. Pathogenesis of Sjögren’s syndrome. Curr. Opin. Rheumatol. 2009, 21, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.E.; Schaumburg, C.S.; Dana, R.; Calonge, M.; Niederkorn, J.Y.; Pflugfelder, S.C. Autoimmunity at the ocular surface: Pathogenesis and regulation. Mucosal Immunol. 2010, 3, 425–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulks, G.N.; Forstot, S.L.; Donshik, P.C.; Forstot, J.Z.; Goldstein, M.H.; Lemp, M.A.; Nelson, J.D.; Nichols, K.K.; Pflugfelder, S.C.; Tanzer, J.M.; et al. Clinical Guidelines for Management of Dry Eye Associated with Sjogren Disease. Ocul. Surf. 2015, 13, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Vivino, F.B.; Carsons, S.E.; Foulks, G.; Daniels, T.E.; Parke, A.; Brennan, M.T.; Forstot, S.L.; Scofield, R.H.; Hammitt, K.M. New Treatment Guidelines for Sjogren’s Disease. Rheum. Dis. Clin. N. Am. 2016, 42, 531–551. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Edman, M.C.; Janga, S.R.; Shi, P.; Dhandhukia, J.; Liu, S.; Louie, S.G.; Rodgers, K.; Mackay, J.A.; Hamm-Alvarez, S.F. A rapamycin-binding protein polymer nanoparticle shows potent therapeutic activity in suppressing autoimmune dacryoadenitis in a mouse model of Sjogren’s syndrome. J. Control. Release Off. J. Control. Release Soc. 2013, 171, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Edman, M.C.; Reddy Janga, S.; Yarber, F.; Meng, Z.; Klinngam, W.; Bushman, J.; Ma, T.; Liu, S.; Louie, S.; et al. Rapamycin Eye Drops Suppress Lacrimal Gland Inflammation In a Murine Model of Sjogren’s Syndrome. Investig. Ophthalmol. Vis. Sci. 2017, 58, 372–385. [Google Scholar] [CrossRef]
- Bella, J.; Kolatkar, P.R.; Marlor, C.W.; Greve, J.M.; Rossmann, M.G. The structure of the two amino-terminal domains of human ICAM-1 suggests how it functions as a rhinovirus receptor and as an LFA-1 integrin ligand. Proc. Natl. Acad. Sci. USA 1998, 95, 4140–4145. [Google Scholar] [CrossRef] [Green Version]
- Giorelli, M.; De Blasi, A.; Defazio, G.; Avolio, C.; Iacovelli, L.; Livrea, P.; Trojano, M. Differential regulation of membrane bound and soluble ICAM 1 in human endothelium and blood mononuclear cells: Effects of interferon beta-1a. Cell Commun. Adhes. 2002, 9, 259–272. [Google Scholar] [CrossRef]
- Min, J.K.; Kim, Y.M.; Kim, S.W.; Kwon, M.C.; Kong, Y.Y.; Hwang, I.K.; Won, M.H.; Rho, J.; Kwon, Y.G. TNF-related activation-induced cytokine enhances leukocyte adhesiveness: Induction of ICAM-1 and VCAM-1 via TNF receptor-associated factor and protein kinase C-dependent NF-kappaB activation in endothelial cells. J. Immunol. (Baltim. Md. 1950) 2005, 175, 531–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, E.O. ICAM-1: Getting a grip on leukocyte adhesion. J. Immunol. (Baltim. Md. 1950) 2011, 186, 5021–5023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, M.D.; Bielinski, S.J.; Suarez-Lopez, J.R.; Reiner, A.P.; Bailey, K.; Thyagarajan, B.; Carr, J.J.; Duprez, D.A.; Jacobs, D.R., Jr. Circulating soluble intercellular adhesion molecule 1 and subclinical atherosclerosis: The Coronary Artery Risk Development in Young Adults Study. Clin. Chem. 2012, 58, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Fotis, L.; Agrogiannis, G.; Vlachos, I.S.; Pantopoulou, A.; Margoni, A.; Kostaki, M.; Verikokos, C.; Tzivras, D.; Mikhailidis, D.P.; Perrea, D. Intercellular adhesion molecule (ICAM)-1 and vascular cell adhesion molecule (VCAM)-1 at the early stages of atherosclerosis in a rat model. In Vivo 2012, 26, 243–250. [Google Scholar] [PubMed]
- Mendivil, C.O.; Robles-Osorio, L.; Horton, E.S.; Hamdy, O.; Caballero, A.E. Young Hispanics at risk of type 2 diabetes display endothelial activation, subclinical inflammation and alterations of coagulation and fibrinolysis. Diabetol. Metab. Syndr. 2013, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Jing, L.; Wang, J.G.; Zhang, J.Z.; Cao, C.X.; Chang, Y.; Dong, J.D.; Guo, F.Y.; Li, P.A. Upregulation of ICAM-1 in diabetic rats after transient forebrain ischemia and reperfusion injury. J. Inflamm. 2014, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Adukpo, S.; Kusi, K.A.; Ofori, M.F.; Tetteh, J.K.; Amoako-Sakyi, D.; Goka, B.Q.; Adjei, G.O.; Edoh, D.A.; Akanmori, B.D.; Gyan, B.A.; et al. High plasma levels of soluble intercellular adhesion molecule (ICAM)-1 are associated with cerebral malaria. PLoS ONE 2013, 8, e84181. [Google Scholar] [CrossRef] [Green Version]
- Stem, M.E.; Gao, J.; Morgan, G.A.; Brees, D.; Schwalb, T.A.; Humphreys-Behr, M.; Smith, J.A. The role of ICAM-1 as a signal protein for predisposition of ocular surface inflammation. Adv. Exp. Med. Biol. 2002, 506, 753–759. [Google Scholar]
- Gao, J.; Morgan, G.; Tieu, D.D.; Schwalb, T.A.; Ngo, M.; Stern, M.E. ICAM-1: Its role in the pathophysiology of immune activation in the MRL/LPR mouse. Adv. Exp. Med. Biol. 2002, 506, 777–781. [Google Scholar]
- Ju, Y.; Guo, H.; Yarber, F.; Edman, M.C.; Peddi, S.; Janga, S.R.; MacKay, J.A.; Hamm-Alvarez, S.F. Molecular Targeting of Immunosuppressants Using a Bifunctional Elastin-Like Polypeptide. Bioconj. Chem. 2019, 30, 2358–2372. [Google Scholar] [CrossRef]
- Zhong, M.; Hanan, E.J.; Shen, W.; Bui, M.; Arkin, M.R.; Barr, K.J.; Evanchik, M.J.; Hoch, U.; Hyde, J.; Martell, J.R.; et al. Structure-activity relationship (SAR) of the alpha-amino acid residue of potent tetrahydroisoquinoline (THIQ)-derived LFA-1/ICAM-1 antagonists. Bioorganic Med. Chem. Lett. 2011, 21, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Yusuf-Makagiansar, H.; Anderson, M.E.; Yakovleva, T.V.; Murray, J.S.; Siahaan, T.J. Inhibition of LFA-1/ICAM-1 and VLA-4/VCAM-1 as a therapeutic approach to inflammation and autoimmune diseases. Med. Res. Rev. 2002, 22, 146–167. [Google Scholar] [CrossRef] [PubMed]
- Nicolls, M.R.; Gill, R.G. LFA-1 (CD11a) as a therapeutic target. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc.Transplant Surg. 2006, 6, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Semba, C.P.; Gadek, T.R. Development of lifitegrast: A novel T-cell inhibitor for the treatment of dry eye disease. Clin. Ophthalmol. 2016, 10, 1083–1094. [Google Scholar] [CrossRef] [Green Version]
- Belizaire, A.K.; Tchistiakova, L.; St-Pierre, Y.; Alakhov, V. Identification of a murine ICAM-1-specific peptide by subtractive phage library selection on cells. Biochem. Biophys. Res. Commun. 2003, 309, 625–630. [Google Scholar] [CrossRef]
- Urry, D. Physical chemistry of biological free energy transduction as demonstrated by elastic protein-based polymers. J. Phys. Chem. B 1997, 101, 11007–11028. [Google Scholar] [CrossRef]
- Janib, S.M.; Gustafson, J.A.; Minea, R.O.; Swenson, S.D.; Liu, S.; Pastuszka, M.K.; Lock, L.L.; Cui, H.; Markland, F.S.; Conti, P.S.; et al. Multimeric disintegrin protein polymer fusions that target tumor vasculature. Biomacromolecules 2014, 15, 2347–2358. [Google Scholar] [CrossRef]
- Shi, P.; Aluri, S.; Lin, Y.A.; Shah, M.; Edman, M.; Dhandhukia, J.; Cui, H.; MacKay, J.A. Elastin-based protein polymer nanoparticles carrying drug at both corona and core suppress tumor growth in vivo. J. Control. Release Off. J.Control. Release Soc. 2013, 171, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, P.; Edman, M.C.; Sun, G.; Shi, P.; Xu, S.; Lin, Y.; Cui, H.; Hamm-Alvarez, S.F.; Andrew MacKay, J. Tear-mediated delivery of nanoparticles through transcytosis of the lacrimal gland. J. Control. Release Off. J. Control. Release Soc. 2015, 208, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Despanie, J.; Shi, P.; Edman-Woolcott, M.C.; Lin, Y.; Cui, H.; Heur, J.M.; Fini, M.E.; Hamm-Alvarez, S.F.; MacKay, J.A. Lacritin-mediated regeneration of the corneal epithelia by protein polymer nanoparticles. J. Mater. Chem. B Mater. Biol. Med. 2014, 2, 8131–8141. [Google Scholar] [CrossRef] [Green Version]
- Muro, S.; Wiewrodt, R.; Thomas, A.; Koniaris, L.; Albelda, S.M.; Muzykantov, V.R.; Koval, M. A novel endocytic pathway induced by clustering endothelial ICAM-1 or PECAM-1. J. Cell Sci. 2003, 116, 1599–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muro, S.; Gajewski, C.; Koval, M.; Muzykantov, V.R. ICAM-1 recycling in endothelial cells: A novel pathway for sustained intracellular delivery and prolonged effects of drugs. Blood 2005, 105, 650–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.H.; Tudares, M.A.; Nguyen, C.Q. Sjogren’s syndrome: An old tale with a new twist. Arch. Immunol. Et Ther. Exp. 2009, 57, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, K.S.; Klickstein, L.B.; Weber, C. Specific activation of leukocyte beta2 integrins lymphocyte function-associated antigen-1 and Mac-1 by chemokines mediated by distinct pathways via the alpha subunit cytoplasmic domains. Mol. Biol. Cell 1999, 10, 861–873. [Google Scholar] [CrossRef]
- Rey-Ladino, J.A.; Huber, M.; Liu, L.; Damen, J.E.; Krystal, G.; Takei, F. The SH2-containing inositol-5′-phosphatase enhances LFA-1-mediated cell adhesion and defines two signaling pathways for LFA-1 activation. J. Immunol. (Baltim. Md. 1950) 1999, 162, 5792–5799. [Google Scholar]
- Kandula, S.; Abraham, C. LFA-1 on CD4+ T Cells Is Required for Optimal Antigen-Dependent Activation In Vivo. J. Immunol. 2004, 173, 4443. [Google Scholar] [CrossRef] [Green Version]
- Mesri, M.; Liversidge, J.; Forrester, J.V. ICAM-1/LFA-1 interactions in T-lymphocyte activation and adhesion to cells of the blood-retina barrier in the rat. Immunology 1994, 83, 52–57. [Google Scholar]
- Varga, G.; Nippe, N.; Balkow, S.; Peters, T.; Wild, M.K.; Seeliger, S.; Beissert, S.; Krummen, M.; Roth, J.; Sunderkötter, C.; et al. LFA-1 Contributes to Signal I of T-Cell Activation and to the Production of Th1 Cytokines. J. Investig. Dermatol. 2010, 130, 1005–1012. [Google Scholar] [CrossRef]
- Abraham, C.; Griffith, J.; Miller, J. The Dependence for Leukocyte Function-Associated Antigen-1/ICAM-1 Interactions in T Cell Activation Cannot Be Overcome by Expression of High Density TCR Ligand. J. Immunol. 1999, 162, 4399–4405. [Google Scholar]
- Pflugfelder, S.C.; Stern, M.; Zhang, S.; Shojaei, A. LFA-1/ICAM-1 Interaction as a Therapeutic Target in Dry Eye Disease. J. Ocul. Pharmacol. Ther. Off. J. Assoc. Ocul. Pharmacol. Ther. 2017, 33, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Whitcup, S.M.; Chan, C.-C.; Kozhich, A.T.; Magone, M.T. Blocking ICAM-1 (CD54) and LFA-1 (CD11a) Inhibits Experimental Allergic Conjunctivitis. Clin. Immunol. 1999, 93, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Roybal, K.T.; Mace, E.M.; Mantell, J.M.; Verkade, P.; Orange, J.S.; Wülfing, C. Early Signaling in Primary T Cells Activated by Antigen Presenting Cells Is Associated with a Deep and Transient Lamellal Actin Network. PLoS ONE 2015, 10, e0133299. [Google Scholar] [CrossRef] [PubMed]
- Wülfing, C.; Sjaastad, M.D.; Davis, M.M. Visualizing the dynamics of T cell activation: Intracellular adhesion molecule 1 migrates rapidly to the T cell/B cell interface and acts to sustain calcium levels. Proc. Natl. Acad. Sci. USA 1998, 95, 6302–6307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnacho, C.; Serrano, D.; Muro, S. A fibrinogen-derived peptide provides intercellular adhesion molecule-1-specific targeting and intraendothelial transport of polymer nanocarriers in human cell cultures and mice. J. Pharmacol. Exp. Ther. 2012, 340, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Chittasupho, C.; Duangrat, C.; Siahaan, T.J.; Berkland, C. PLGA nanoparticle--peptide conjugate effectively targets intercellular cell-adhesion molecule-1. Bioconj. Chem. 2008, 19, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Muro, S.; Cui, X.; Gajewski, C.; Murciano, J.C.; Muzykantov, V.R.; Koval, M. Slow intracellular trafficking of catalase nanoparticles targeted to ICAM-1 protects endothelial cells from oxidative stress. Am. J. Physiol. Cell Physiol. 2003, 285, C1339–C1347. [Google Scholar] [CrossRef] [Green Version]
- Muro, S.; Schuchman, E.H.; Muzykantov, V.R. Lysosomal enzyme delivery by ICAM-1-targeted nanocarriers bypassing glycosylation- and clathrin-dependent endocytosis. Mol. Ther. J. Am. Soc. Gene Ther. 2006, 13, 135–141. [Google Scholar] [CrossRef]
- Rossin, R.; Muro, S.; Welch, M.J.; Muzykantov, V.R.; Schuster, D.P. In vivo imaging of 64Cu-labeled polymer nanoparticles targeted to the lung endothelium. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2008, 49, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Chittasupho, C.; Siahaan, T.J.; Vines, C.M.; Berkland, C. Autoimmune therapies targeting costimulation and emerging trends in multivalent therapeutics. Ther. Deliv. 2011, 2, 873–889. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Matthews, K.; Siahaan, T.J.; Kevil, C.G. Alpha L-integrin I domain cyclic peptide antagonist selectively inhibits T cell adhesion to pancreatic islet microvascular endothelium. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G67–G73. [Google Scholar] [CrossRef] [Green Version]
- Tibbetts, S.A.; Seetharama Jois, D.; Siahaan, T.J.; Benedict, S.H.; Chan, M.A. Linear and cyclic LFA-1 and ICAM-1 peptides inhibit T cell adhesion and function. Peptides 2000, 21, 1161–1167. [Google Scholar] [CrossRef]
- Yusuf-Makagiansar, H.; Yakovleva, T.V.; Tejo, B.A.; Jones, K.; Hu, Y.; Verkhivker, G.M.; Audus, K.L.; Siahaan, T.J. Sequence recognition of alpha-LFA-1-derived peptides by ICAM-1 cell receptors: Inhibitors of T-cell adhesion. Chem. Biol. Drug Des. 2007, 70, 237–246. [Google Scholar] [CrossRef]
- Shannon, J.P.; Silva, M.V.; Brown, D.C.; Larson, R.S. Novel cyclic peptide inhibits intercellular adhesion molecule-1-mediated cell aggregation. J. Pept. Res. Off. J. Am. Pept. Soc. 2001, 58, 140–150. [Google Scholar] [CrossRef]
- Semba, C.P.; Torkildsen, G.L.; Lonsdale, J.D.; McLaurin, E.B.; Geffin, J.A.; Mundorf, T.K.; Kennedy, K.S.; Ousler, G.W. A phase 2 randomized, double-masked, placebo-controlled study of a novel integrin antagonist (SAR 1118) for the treatment of dry eye. Am. J. Ophthalmol. 2012, 153, 1050–1060.e1. [Google Scholar]
- Sheppard, J.D.; Torkildsen, G.L.; Lonsdale, J.D.; D’Ambrosio, F.A., Jr.; McLaurin, E.B.; Eiferman, R.A.; Kennedy, K.S.; Semba, C.P. Lifitegrast ophthalmic solution 5.0% for treatment of dry eye disease: Results of the OPUS-1 phase 3 study. Ophthalmology 2014, 121, 475–483. [Google Scholar] [CrossRef]
- Muro, S. A DNA-Device that Mediates Selective Endosomal Escape and Intracellular Delivery of Drugs and Biologicals. Adv. Funct. Mater. 2014, 24, 2899–2906. [Google Scholar] [CrossRef] [Green Version]
- MacEwan, S.R.; Chilkoti, A. Applications of elastin-like polypeptides in drug delivery. J. Control. Release Off. J. Control. Release Soc. 2014, 190, 314–330. [Google Scholar] [CrossRef] [Green Version]
- Baiu, D.C.; Prechl, J.; Tchorbanov, A.; Molina, H.D.; Erdei, A.; Sulica, A.; Capel, P.J.; Hazenbos, W.L. Modulation of the humoral immune response by antibody-mediated antigen targeting to complement receptors and Fc receptors. J. Immunol. (Baltim., Md. 1950) 1999, 162, 3125–3130. [Google Scholar]
- Aina, O.H.; Sroka, T.C.; Chen, M.L.; Lam, K.S. Therapeutic cancer targeting peptides. Biopolymers 2002, 66, 184–199. [Google Scholar] [CrossRef]
- Camacho, C.J.; Katsumata, Y.; Ascherman, D.P. Structural and thermodynamic approach to peptide immunogenicity. PLoS Comput. Biol. 2008, 4, e1000231. [Google Scholar] [CrossRef] [Green Version]
- National Research Council (US) Committee. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press (US): Washington, DC, USA, 2011. [Google Scholar]
- Sun, G.; Hsueh, P.Y.; Janib, S.M.; Hamm-Alvarez, S.; MacKay, J.A. Design and cellular internalization of genetically engineered polypeptide nanoparticles displaying adenovirus knob domain. J. Control. Release Off. J. Control. Release Soc. 2011, 155, 218–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]







| Label | Amino Acid Sequence a | Tt1b (°C) | Tt2c (°C) | Expected MW d (kDa) | Measured MW e (kDa) | Hydrodynamic Radius f (nm) |
|---|---|---|---|---|---|---|
| SI | MG(VPGSG)48(VPGIG)48Y | 25.5 | 73.8 | 39.6 | 39.5 | 23.6 ± 0.4 |
| IBP-SI | MGFEGFSFLAFEDFVSSIG (VPGSG)48(VPGIG)48Y | 25.7 | 46.9 | 41.5 | 41.4 | 21.9 ± 0.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsueh, P.-Y.; Ju, Y.; Vega, A.; Edman, M.C.; MacKay, J.A.; Hamm-Alvarez, S.F. A Multivalent ICAM-1 Binding Nanoparticle which Inhibits ICAM-1 and LFA-1 Interaction Represents a New Tool for the Investigation of Autoimmune-Mediated Dry Eye. Int. J. Mol. Sci. 2020, 21, 2758. https://doi.org/10.3390/ijms21082758
Hsueh P-Y, Ju Y, Vega A, Edman MC, MacKay JA, Hamm-Alvarez SF. A Multivalent ICAM-1 Binding Nanoparticle which Inhibits ICAM-1 and LFA-1 Interaction Represents a New Tool for the Investigation of Autoimmune-Mediated Dry Eye. International Journal of Molecular Sciences. 2020; 21(8):2758. https://doi.org/10.3390/ijms21082758
Chicago/Turabian StyleHsueh, Pang-Yu, Yaping Ju, Adrianna Vega, Maria C. Edman, J. Andrew MacKay, and Sarah F. Hamm-Alvarez. 2020. "A Multivalent ICAM-1 Binding Nanoparticle which Inhibits ICAM-1 and LFA-1 Interaction Represents a New Tool for the Investigation of Autoimmune-Mediated Dry Eye" International Journal of Molecular Sciences 21, no. 8: 2758. https://doi.org/10.3390/ijms21082758