Citrus Taste Modification Potentials by Genetic Engineering
by
 and
and
Li-Jun Li
1,2,3, 
Wan-Seng Tan
1,
Wen-Jing Li
1,
Yan-Bing Zhu
1,2,3,
Yi-Sheng Cheng
4,5 and
Hui Ni
1,2,3,* 1
College of Food and Biological Engineering, Jimei University, Xiamen 361021, China
2
Fujian Provincial Key Laboratory of Food Microbiology and Enzyme Engineering, Xiamen 361021, China
3
Research Center of Food Biotechnology of Xiamen City, Xiamen 361021, China
4
Department of Life Science, National Taiwan University, Taipei 10617, Taiwan
5
Institute of Plant Biology, National Taiwan University, Taipei 10617, Taiwan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(24), 6194; https://doi.org/10.3390/ijms20246194
Submission received: 31 October 2019
/
Revised: 25 November 2019
/
Accepted: 4 December 2019
/
Published: 8 December 2019
(This article belongs to the Collection Genetics and Molecular Breeding in Plants)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Citrus fruits are mainly consumed as fresh fruit and processed juice products. They serve as nutritional and a tasty diet in our daily life. However, the formidable bitterness and delayed bitterness significantly impact the citrus industry attributable to the two major bitter compounds naringin and limonin. The extremely sour and acidic also negatively affects the sensory quality of citrus products. Citrus breeding programs have developed different strategies to improve citrus quality and a wealth of studies have aimed to uncover the genetic and biochemical basis of citrus flavor. In this minireview, we outline the major genes characterized to be involved in pathways shaping the sweet, bitter, or sour taste in citrus, and discuss briefly about the possible approaches to modify citrus taste by genetic engineering.
1. Introduction
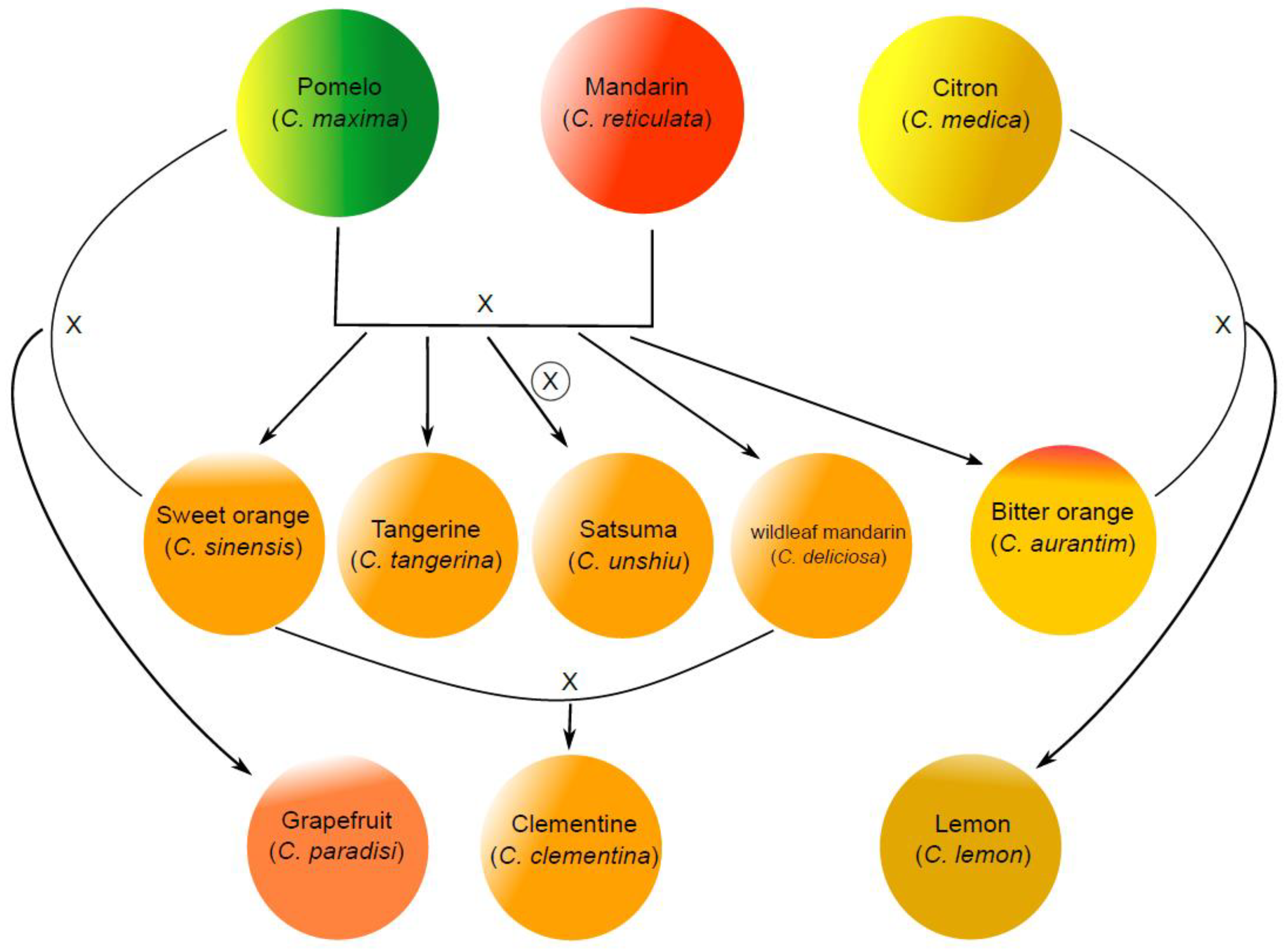
It was suggested that the Citrus genus originated from the Malay Archipelago and Southeast Asia, and the varieties of edible citrus fruits on the market are generated by hybridizations of ancestral species, natural or artificial mutations, and human selection during domestication [1,2]. The genealogy and genetic origin of the major citrus species are roughly summarized in Figure 1. The major ancestral species largely include mandarin (Citrus reticulata), pomelo (Citrus maxima), and citron (Citrus medica). The classification of commercial mandarins is very complicated, which largely include tangerine, satsuma, and clementine. Citrus hybrids are more diversified, and their taxonomy is somewhat inconsistent. The major hybrids as our common consumption include sweet orange (Citrus sinensis), bitter orange (Citrus aurantim), grapefruit (Citrus paradisi), lemon (Citrus limon), and limes [3]. Orange is a hybrid between pomelo and mandarin, and grapefruit is the product of a natural backcross of a sweet orange with a pomelo [4]. The original lemon is a hybrid of bitter orange and citron, while lime is highly diverse including several species. According to the Food and Agriculture Organization of the United Nations (FAO) citrus fruits statistics 2017 (FAO statistics, 2017), citrus fruit accounts for about 17% of total fruit production, in which about 50% and 23% of citrus fruits are orange and mandarins, respectively.
Citrus fruits have great nutritional and pharmacological values. Citrus is rich in vitamins, especially in vitamin C and is a good source of minerals and dietary fibers [5]. The phytochemicals such as monoterpenes, limonoids (triterpenes), flavonoids, carotenoids, and hydroxycinnamic acid have great potentials for the protection of humans against chronic diseases and cancer [6]. The citrus products consumed consist mostly of fresh fruits and fruit juice. The orange is more favored as processed orange juice, such as single-strength orange juice and frozen juice concentrate, which can be processed through up to 95% of orange fruit produced in Florida, US and 70% in Brazil, according the United States Department of Agriculture (USDA) report about citrus world markets released in July 2019. Generally, flavor is sensed by a combination of the taste and olfactory systems. Five primary taste sensations in our mouth are salty, sour, sweet, bitter, and umami [7]. As for citrus fruits and juice, they normally impress us with their sweet and sour. Although flavor is favored differently for different individuals, there is a generally accepted notion of good taste which is attributed to an appropriate balance of sweetness and sourness in citrus [7]. Excessive bitterness and the delayed bitterness in citrus pose great challenge for the citrus industry to provide consumers readily acceptable fresh fruit and citrus juice, thus causing great lose in citrus market value [8]. Extremely sour and acidic can destroy the sweet taste of citrus and may be physiologically harmful to humans. To improve citrus quality, traditional breeding techniques including cross-pollination, self-pollination, selection of natural mutations or induced mutations, and rootstock hybrid. Extensive studies have been performed to understand the citrus flavor-contributing factors through biochemical and genetic approaches [7,9]. In this review, we mainly focus on the understanding of molecular basis governing citrus sweetness, bitterness, and sourness, providing an overview of the potentials for genetic modification or molecular-assisted breeding to improve citrus overall taste.
2. Citrus Taste
2.1. Sweet
The content of three major carbohydrates (sucrose, fructose, and glucose) as the main components of total soluble solids (TTS) makes the major contribution to citrus sweetness [10]. Sucrose is exported to the developing citrus fruit from the source leaves with sugar accumulation through photosynthesis and conversion to translocation sucrose [11,12]. The partitioning of sucrose into citrus fruit was believed to be determined by the sink strength which is monitored by the enzyme activities of three sucrose-metabolizing enzymes, sucrose phosphate synthase (SPS), sucrose synthase (SS), and invertase (IVR) [13]. The schematic presentation of sucrose transport is shown in Figure 2. The sucrose accumulated in the sink leaves is loaded into the phloem through the well-recognized action of plasmalemma-localized sucrose/H+ symporter and transported to the filial storage site [14,15]. Sucrose is unloaded to the parenchyma tissue and further imported into citrus flesh through apoplastic and symplastic pathways. It was suggested that there existed P-ATPase involved transport of sucrose from apoplast to cytosol and an endocytic pathway of assimilating sucrose directly from apoplast into vacuole, while the sucrose/H+ antiporters for sucrose import into vacuole from cytosol remain to be identified if any [16]. The translocated sucrose is to be either hydrolyzed to fructose and glucose by IVR or cleaved to fructose and uridine diphosphate glucose (UDP-glucose) by SS, and then resynthesized by the activity of SPS. The two steps of sucrose degradation and resynthesis favor the sucrose import by generating a sucrose concentration gradient in the sink local zone for phloem unloading [17]. A collection of genes encoding the enzymes for sucrose transport and metabolism have been reported, including three sucrose transporters (CitSUT1, CitSUT2, and CitSUT3) for sucrose loading or unloading [18], three sucrose synthase isoforms (CitSUS1/2 and CitSUSA) from Citrus unshiu [17], and six SS homologs from publicly available genome database for sweet orange and clementine (citrus.hzau.edu.cn/orange and www.phytozome.net) [19], and their transcriptional and translational expression patterns were investigated during citrus fruit development. Generally, the sucrose synthase activity and expression levels are relatively low at the citrus fruit immature stage favoring sucrose accumulation and increased during fruit development favoring improving sink strength and sucrose import, while different sucrose synthases showed their unique expression patterns [19,20]. Komatsu et al. (2002) investigated the transcript levels of CitSUS1/2 and CitSUSA during citrus development and suggested that CitSUS1 may help provide the sucrose degradation products for growth and cell wall construction while CitSUSA plays more roles in providing substrates for sucrose resynthesis in concert with the function of SPS [17]. Katz et al. (2012) performed proteomic and metabolic analysis during citrus fruit development and revealed that sucrose invertase expression largely remained unchanged while an invertase inhibitor was upregulated in the later stages of fruit development [21]. This further supported the notion that sucrose synthase is the major player mediating sucrose degradation and provides substrates for sucrose re-synthesis by SPS, as manifested that SPS showed co-upregulation with SS at later stages in the sink tissue.
Taken together, sucrose accumulation in citrus fruit is regulated at multiple levels during fruit development. A set of factors may determine the partitioning of sucrose into the fruits, including photosynthesis and conversion of translocation sucrose in leaves, sucrose loading into and unloading from phloem, and the coordination of the major sucrose metabolism-related enzymes and transporters [11]. It has been shown that drought stress or treatment can improve the sink strength by increasing the sucrose synthase activity thus enhancing sucrose importing into citrus fruit [18]. Given the involvement of a set of genes and knowledge of their spatial and temporal expression patterns, it is still not clear if a single gene modification would improve sucrose accumulation in citrus fruit until a comprehensive analysis of knockout mutants or controlled downregulation or upregulation of individual genes to examine their effects on sucrose partitioning.
2.2. Bitter
There are two types of bitterness, namely immediate bitterness and delayed bitterness, in citrus fruits, imparted by two different types of compounds [22,23]. The immediate bitterness is largely conferred by naringin and neohesperidin [1], and the delayed bitterness is mainly produced by limonin of limonoids [24]. Delayed bitterness is gradually developed upon fruit is mechanically damaged, juiced, or frozen [25,26]. Some enzymes are natural debittering enzymes providing citrus palatable quality. The overall picture of citrus bitter-tasting and non-bitter compounds synthesis pathways is sketched in Figure 3.
2.2.1. Cm1,2RhaT
Flavanones are the major group of flavonoids in citrus and the primary bitterness in citrus fruit is determined by flavanone-neohesperidosides in bitter citrus species such as pummelo/pomelo, grapefruit, and bitter orange [27]. The flavanone naringenin is the dominant flavonoid backbone in some citrus species, while it might undergo a variety of modifications to form other flavanones, for instance, hydroxylation and methylation to generate its derivative hesperetin [1,28]. The bitter-tasting naringin is the major contributor to the bitter taste in grapefruit and neohesperidin in sour orange [27,29]. Naringinase is widely used as a debittering enzyme in the commercial production of citrus juice, and our lab has reported a mutated α-L-rhamnosidase of naringinase complex with enhanced performance on thermostability and citrus juice debittering efficacy [30].
In grapefruit, the bitter-tasting naringin is predominant over the tasteless narirutin. They are derived from the same flavanone skeleton naringenin but differ in the rhamnose-glucose disaccharide connected through an O-linkage at position 7. In naringin, the rhamnose is attached to the glucose moiety at the C-2 position, while it is attached at the C-6 position in narirutin, resulting in the formation of naringenin-7-O-neohesperidoside and naringenin-7-O-rutinoside, respectively. The gene for Cm1,2RhaT (flavanone 7-O-glucoside-1,2-rhamnosyltytransferase) was isolated from pummelo (C. maxima) [27], and its expression product of recombinant Cm1,2RhaT enzyme in tobacco BY2 cells exhibited UDP-glycosyltransferase activity specifically converting naringenin-7-O-glucoside into naringin. In addition, Cm1,2RhaT displayed similar enzymatic function on another flavonoid subgroup, flavone. Spatial and temporal expression patterns suggested that Cm1,2RhaT gene is highly expressed in young leaves and young fruit but is not actively transcribed in mature leaves and mature fruit, which match with the developmental accumulation pattern of flavanone-neohesperidosides [27,31]. Cm1,2RhaT expresses exclusively in naringin-containing citrus species (pummelo, grapefruit, and sour-orange), but not in sweet species like orange, citron, and mandarin [27]. A Cm1,2RhaT ortholog was identified with two frameshift mutations abolishing the functional 1,2RhaT expression in sweet orange (C. sinensis) [1]. These findings provide a possibility to modulate the expression of 1,2RhaT by genetic engineering to modify the bitter-tasting naringin content in those citrus species with readily detecTable 1,2RhaT expression.
2.2.2. Cs1,6RhaT
In citrus, naringenin produced from naringenin chalcone catalyzed by chalcone isomerase can be further processed to form different subgroups of flavonoids featuring different structures of C ring, including flavanone, flavone, flavonol, and anthocyanidin [32]. After cloning and characterization of Cm1,2RhaT [27], Frydman et al. (2013) isolated Cs1,6RhaT (flavanone-7-O-glucoside-1,6- rhamnosyltytransferase) from orange (C. sinensis) which diverts another branch of glycosylation of flavanone-7-O-glucoside [1]. Cs1,6RhaT adds rhamnose to the C-6 position of glucoside leading to the formation of the tasteless flavanone-7-O-rutinoside, such as hesperidin and narirutin. Additionally, it turned out that Cs1,6RhaT is substrate-promiscuous which can catalyze 1,6-rhamnosylation of flavonoid-7-O-glucoside in flavanone, flavone, flavonol, and anthocyanidin as well as flavonoid-3-O-glucoside in flavonol and anthocyanidin. Cs1,6RhaT is highly expressed in the non-bitter citrus species such as orange, mandarin and lemon, but with relatively low expression in citron containing overall low levels of flavanones [33]. Contrary to Cm1,2RhaT, Cs1,6RhaT has low levels in grapefruit and bitter orange, and was not detected in pummelo which is rich in flavanone neohesperidosides but lacks rutinosides [1].
A more recent study by Ohashi et al. (2016) was performed on the branch-forming RhaT (rhamnose transferase) in the engineered fission yeast expressing Arabidopsis rhamnose synthase making UDP-Rha available in yeast system [34]. The Cm1,6RhaT was newly isolated from pummelo. Substrate preference studies of Cm1,2RhaT, Cs1,6RhaT, and Cm1,6RhaT indicated that Cm1,6RhaT can transfer rhamnose residue of UDP-rhamnose toward naringenin-7-O-gulcoside as Cs1,6RhaT; Cm1,2Rhat has glycosyl transfer activity using UDP-xylose in addition to UDP-rhamnose as donor toward naringenin-7-O-glucoside. Additionally, Cs1,6RhaT can transfer another rhamnose residue toward narirutin to yield naringenin-7-O-rhamnosylrutinoside which seems not to exist naturally in citrus species or is quite low in abundance if any [34]. Given that the amino acid sequences of Cm1,6RhaT and Cs1,6RhaT share about 94% in identity, the failure of the detection of Cm1,6RhaT by western blot using anti-Cs1,6RhaT antibody might be on account of the extremely low expression of Cm1,6RhaT in pummelo [1,34].
2.2.3. CitdGlcT
1,2RhaT and 1,6RhaT confer the branched rhamnosylation of flavanone/flavone-7-O-glucoside in citrus. To further explore other enzymes governing the flavor-related glycosylation in citrus, especially the di-glycosylation, Chen et al. (2019) performed Basic Local Alignment Search Tool (BLAST) against citrus genome database (http://citrus.hzau.edu.cn/orange/ and phytozome.jgi.doe.gov) using Cm1,2RhaT CDS as query and identified another 1,2RhaT and three dGlcT (di-glucosyltransferase) genes from citrus genome which were referred to as Cit1,2RhaT and CitdGlcT-1, CitdGlcT-2, and CitdGlcT-3, respectively [35]. These genes are not distributed evenly among different citrus species. Metabolite profiling by LC-MS in tobacco BY2 cells harboring the construct of Cit1,2RhaT, CitdGlc-1, or CitdGlc-2 provided with different flavonoid substrates revealed that Cit1,2RhaT and CitdGlcT-1/-2 display high preference for flavanone-glucoside as substrate for glycosylation. Notably, RNAseq analysis in C. maxima fruit juice sac demonstrated that CmdGlcT-2 is the most active GlcT and the dynamic pattern of CmdGlcT-2 expression does not match with the availability of flavanone-7-O-glucoside during fruit development, consequently, there is high abundance of bitter naringin but no detectable bitterless flavanone-7-O-di-glucoside in pummelo. One of the possibilities to modify the bitterness of pummelo or citrus in general is through manipulation of the expression of dGlcTs to compete for the flavonoid substrates for better balance between flavor and nutrients [35]. Furthermore, the co-segregation of the presence of bitter-tasting naringin and 1,2RhaT provides a genetic marker to assist citrus breeding for directed selection [35].
2.2.4. CitLGTs
Limonoid is the other major type of bitter compound in addition to naringin in citrus fruit, including nomilin and limonin [8,24]. A study has shown that generally limonoids accumulate relatively high at young fruit or fruit expanding stage and then fall to very low in concentration or under detectable levels in mature fruit [36]. The majority of the bitter-tasting nomilin and limonin exists in fruit tissue as their non-bitter precursors nomilioate A-ring lactone (NARL) and limonoate A-ring lactone (LARL), respectively, before harvest or processing, and LARL is predominant [37]. The limonoid A-ring lactones in citrus are synthesized in leaves and transported into fruit where they are stored in the seeds and cell cytoplasm at a neutral to slightly alkaline pH environment [23,37,38]. During the juicing process, the precursors NARL and LARL are converted to the bitter nomilin and limonin, respectively, under the acidic condition (pH < 6.5) accelerated with the aid of limonoid D-ring lactone hydrolase (LLH) when the membranous sacs are ruptured [24,37]. The conversion of LARL to limonin can also occur naturally under acidic conditions when citrus fruits are young or when the fruits encounter freezing or mechanical damage [39,40]. The gradual conversion of the non-bitter precursors to the bitter limonoids is known as the process of delayed bitterness.
There are two forms of limonoid in citrus fruits and seeds, the bitter aglycones and non-bitter glucosides [40]. Limonoid aglycones are partly converted to the limonoid glucosides during fruit maturation [41], and soluble limonin glucoside is the predominant limonoid glucoside in citrus juice [42]. In citrus seeds, there exists the only natural repository of limonoid aglycones, where aglycone and glucoside of nomilin exceed those of limonin in concentration [43,44]. The conversion of aglycone of limonoids to glucoside of limonoids is catalyzed by limonoid glycosyltransferase (LGT) during citrus fruit maturation as natural debittering; specifically, LGT converts nomilin and LARL to nomilin glucoside and limonin glucoside, respectively [40].
Researchers have made progress on isolation and characterization of LGT genes from citrus species including satsuma mandarin (C. unshiu) and navel orange (C. sinensis) [39,45], Kinnow mandarin (C. reticulata) [40], sweet lime (C. limettioides), grapefruit (C. paradisi), and sour orange (C. aurantium) [26]. Overall these LGTs share high similarity in their coding sequences but they display different expression patterns during citrus development [26]. The satsuma mandarin and navel orange have two different alleles of LGTs where satsuma mandarin is heterozygous for CitLGT-1/CitLGT-2 and navel orange is homozygous for CitLGT-1. The lack of delayed bitterness in satsuma mandarin was thought to be attributed to the presence of CitLGT-2 which is highly expressed throughout the entire citrus fruit development, while the navel orange has the unpleasant delayed bitterness was on account of the much delayed expression (120 DAF) of CitLGT-1 allele in the juice sac/segment epidermis [39]. Similar observations have been obtained from the citrus species with different levels of delayed bitterness demonstrating that the delayed presence of CitLGTs in the albedo tissue of citrus fruit is in agreement with the intensity of delayed bitterness [26]. The differential expression patterns provide a clue for marker-assisted selection during hybrid breeding and a target molecule for genetic engineering for debittering enhancement.
2.3. Sour
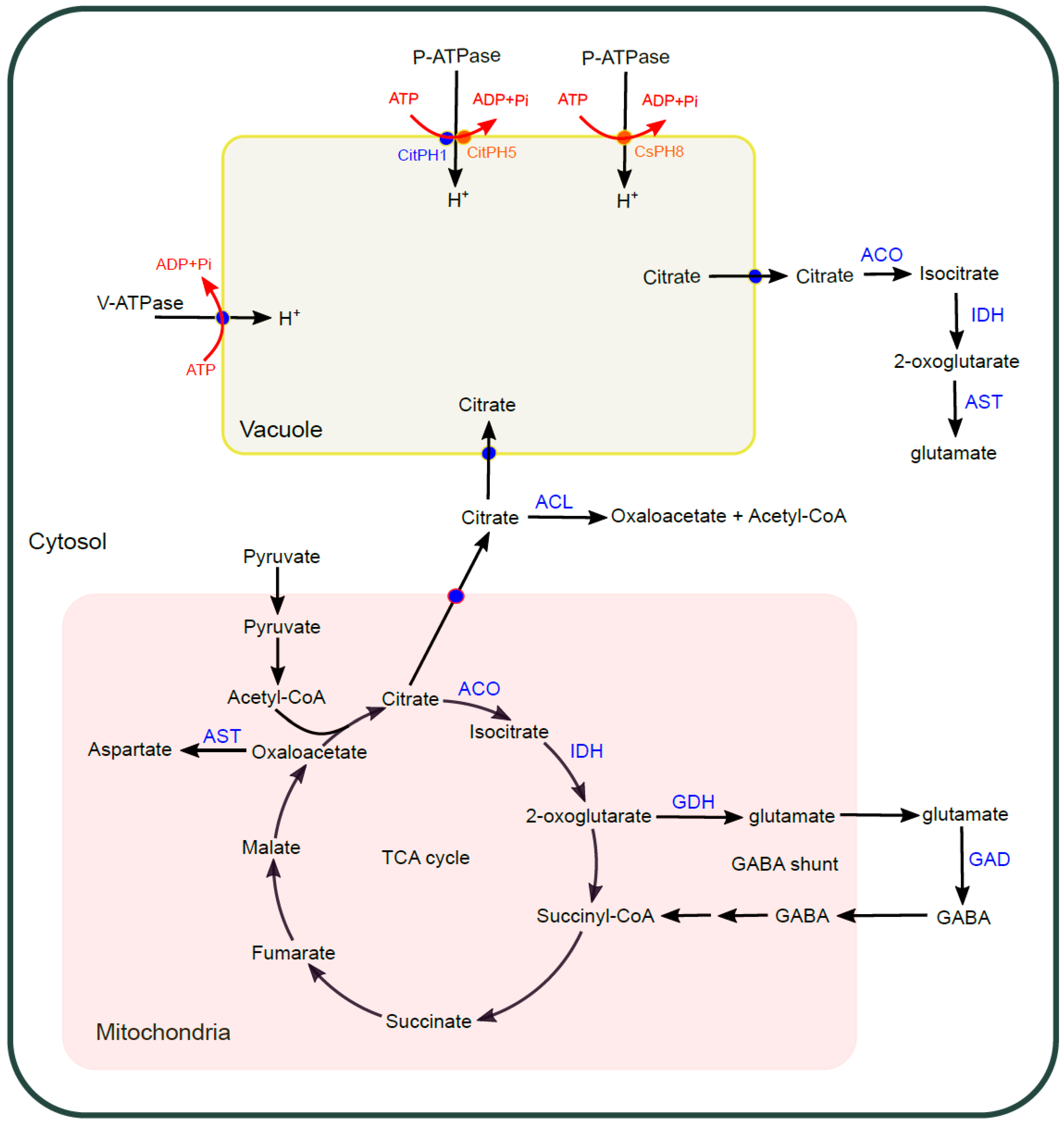
The sour taste of citrus fruits is mainly due to the presence of citric acid and malic acid, with citric acid as the dominant organic acid [46]. More specifically, the ratio of total soluble solid to titratable acid (TSS/TA) governs the quality and maturity of citrus fruits, which mainly reflects the ratio of sugar to citric acid content [47,48]. In the US, a minimum TSS/TA ratio of 7–9:1 is favored for oranges and mandarins, and 5–7:1 for grapefruit (http://www.yara.us/agriculture/). During citrus fruit ripening process, normally the sucrose content increases and the citric acid decreases, leading to the declined acidity and sourness; however, acidity increases with maturity in lemon causing the ratio of TSS/TA close to 1–2:1 [3,49]. High content of citric acid may mask the perception of sweet taste, influencing the overall taste of citrus fruit [50]. On the other hand, some natural citrus variants are acidless and thus taste sweet [51]. Citric acid is produced through tricarboxylic acid cycle (TCA cycles) in mitochondria and accumulated in vacuole of juice sacs during early-mid stage of fruit development and gradually degraded at the later stages in cytosol after released from vacuole [52,53]. Metabolic pathways were proposed to degrade citric acid to control its accumulation; its breakdown can be through the TCA cycle or γ-aminobutyric acid shunt (GABA shunt) in mitochondria, or through the acetyl-CoA pathway catalyzed by ATP-citrate lyase (ACL) in cytosol, characterized by the upregulated expression of related genes during fruit maturation [13,46]. It is also hypothesized that the citric acid after its export from vacuole to cytosol can be converted to 2-oxoglutarate catalyzed by the sequential action of cytosolic aconitase (Aco) and isocitrate dehydrogenase (IDH) for glutamate synthesis and further metabolism. The activity of aconitase was proposed to paly important role in the metabolism of citric acid during citrus fruit development [52,54], while a more thorough investigation is still needed. The metabolism network of citrate in citrus is summarized in Figure 4. Nevertheless, the vacuolar storage of citrate after export and degradation has a determinative effect on citrus fruit sour taste. Here we will give more details about understanding the underlying molecular mechanism regulating hyperacidity in vacuole and citrate flux into/out of vacuole.
2.3.1. V-ATPase
It has been suggested that vacuolar H+-ATPase (V-ATPase) acts as the proton pump driving the H+ import into vacuole at the expense of ATP hydrolysis, and the resulting ΔpH and H+ electrochemical gradient facilitate the transport of citrate into vacuole [55,56]. Li et al. (2016) suggested that CitVHA-c4, one of the V-ATPases in C. reticulata, is involved in the citrate accumulation in vacuole considering its substantially high expression at fruit late development stage [57]. Biochemical assays indicated that the V-ATPase CitVHA-c4 can physically interact with the transcription factor CitERF13, and they can act alone or cooperatively in promoting citrus acid accumulation in transfected tobacco leaves [57]. An earlier research by Shimada et al. (2006) reported the isolation of a vacuolar H+-ATPase from C. sinesis, namely CsCit1, which functions as an active co-transporter of citrate and H+ in an electroneutral manner (citrateH2−/2H+) for citrate export from vacuole. The action manner of citrateH2−/2H+ possibly enhances the buffer capacity by selectively exporting the conjugate base of citrateH2− thus favoring the maintenance of high vacuolar acidity during citrus fruit maturation [58]. These findings suggest the regulatory role of V-ATPase in vacuolar citrate buffer content and acidity.
2.3.2. P-ATPase
The pH values in citrus such as sour lemon and lime can be as low as 2, which require ATPase with low H+/ATP ratio (<2) for hyper-acidification except for the regular V-ATPase for vacuolar lumens acidification [59,60]. In petunia, the interacting PH1 of P3B-ATPase (Mg2+ pump) and PH5 of P3A-ATPase (H+ pump) were reported to be the indispensable players for proton translocation across tonoplast against a larger electrochemical gradient for vacuolar hyperacidification in epidermal petal cells [61]. Strazzer et al. (2019) provided a wealth of evidence demonstrating that the two citrus homologs, CitPH1 and CitPH5, act as a heterodimer holding major responsibility for the hyperacidity in juice vesicles [62]. CitPH1 and CitPH5 are highly expressed in acidic (low pH) citrus fruit while strongly suppressed in non-acidic (high pH) one. Their downregulated expressions in high-pH citrus were attributed to the mutations in the transcription regulator CitAN1 (in sweet lemon and sweet oranges) or the reduced expression of the transcription factors CitAN1, CitPH3, and/or CitPH4 as a result of altered functions of their upstream trans-acting factors [62]. An in-depth investigation of those acidless variants in citrus species including citron, lemon, sweet lime, and sweet orange uncovered that a bHLH-type transcription factor, namely Noemi, is the potent contributor to citrus acidity except for its role in anthocyanin production [51]. The Noemi protein is a homolog of AN1 in petunia. The abolished or reduced expression of Noemi leading to acidless or reduced acidity was attributed to the mutation in Noemi alleles with deletion or transposon insertion in the coding region or small lesion adjacent to TATA-box in the promoter region [51]. The regulatory role of AN1 in CitPH1 and CitPH5 expression can account for the conserved regulatory role of Noemi in citrus acidity. Moreover, the CsPH4 in C. sinensis was characterized to be a physical interactor with Noemi and positively regulate the expression of Noemi as CsPH4-Noemi complex. Their expression levels positively correlated with the expression of CsPH5 and the citrus acidity [63], providing another line of evidence in support of the citrus acidity regulatory complex involving CitPH1, CitPH5, and their transcription regulators. Additionally, Shi et al. (2019) further characterized CsPH8, another citrus homolog of petunia PH5, as a positive regulator of citrate content in citrus fruits based on previous profiling of CsPH8 expression in citrus cultivars with different acidity [64,65]. These findings substantiate the notion that the steep pH gradient across the tonoplast is the powerful driving force for titratable citrate uptake into vacuole providing the sensation of sour taste [62,64]. CitPH1, CitPH5, CsPH8, and their transcription regulators provide promising genetic loci for molecular marker selection and genetic breeding in citrus for desired taste.
3. Citrus Genetic Modification Approach
Conventional citrus breeding is time-consuming and difficult as citrus has an extended juvenile period and possesses several special reproductive characteristics such as high heterozygosity, polyembryony, and self-incompatibility [66]. The parent lines with desirable traits are to be identified and genetically crossed, and it will take about another 20 years before the hybrid sets fruits for taste or flavor evaluation [9]. Ectopic expression of the citrus homolog of the Flowering locus T (FT) has been reported to be able to induce early flowering in trifoliate orange [67]. Dutt et al. (2005) also successfully generated early flowering Carrizo rootstock by constitutively expressing Clementine FT gene in the transgenic plant [68], and the induction of early flowering by FT was also documented in citrus plants of different genotypes by graft inoculation with the Citrus leaf blotch virus-based vector harboring the AtFT (Arabidopsis FT gene) or CiFT (Valencia late sweet orange FT gene) [69]. As documented, the transgenic plantlets can start to flower when they were 3–6 months old under the induction of the FT signal [68,69]. These findings provide a feasible approach to speeding up the breeding process. Given the mobile property of the FT signal, the exotic gene contamination can be avoided when the transgenic citrus is used as rootstock. The virus-based expression vector has this intrinsic advantage for transient expression of an exogenous gene of interest efficiently and systemically. It is encouraging to observe the precocious flowering phenotype induced by viral vector-medicated FT expression without compromising plant growth and citrus fruit development [69].
The molecular marker-assisted breeding can identify the citrus candidates with potential desired flavor at early stage [70]. Quantitative trait locus (QTL) mapping and other molecular marker genetic linkage analysis have been widely applied to explore certain associated target traits in marker-assisted selection program. A collection of single nucleotide polymorphisms based QTLs (SNP-based QTLs) was mapped to be associated with mandarin aroma volatiles production, which could facilitate the mandarin fruit flavor improvement [71]. The above-mentioned key genes controlling citrus sour and bitter taste qualities can serve as excellent DNA markers to evaluate the sensory traits of citrus fruit when the plants are still at juvenile stages. In addition, agrobacterium-mediated genetic transformation is a well-developed approach for direct introduction of gene of interest into citrus genome, and some advances have been made in using genetic element of plant origin as driving promoter or selection marker to avoid foreign gene introduction [72]. Other transformation methods such as poly ethylene glycol mediated (PEG-mediated) plasmid DNA uptake by protoplasts and biolistic DNA delivery also made success in citrus genetic modification [73,74]. Generally, antibiotic or herbicide resistance is used as selection marker for transgenic plantlets production. To overcome the risk of introduction of exotic DNA fragments into donor plants, it is worthwhile to mention that Ruby gene and citrus-derived embryo-specific promoter have been employed as visual reporter gene to monitor the success of transgene in citrus, as Ruby can act as a transcriptional activator of anthocyanin production [68,73]. The best-case scenarios would be upregulated or ectopic expression of the genes conferring “good” taste or repression of the genes controlling “bad” taste leads to enhancement of the overall taste in citrus. Moreover, the development of CRISPR/Cas system has facilitated the precision genome editing in plant, making it possible to generate transgene-free edited crops [75]. Several studies have reported using CRISPR/Cas9 system [76,77,78,79,80,81] or CRISPR/Cas12 system [82] as an efficient tool for genome editing in citrus. Great efforts have been focused on the improvement of citrus resistance to bacterial canker and HuangLongbing (HLB) through CRISPR/Cas system and conventional transformation strategies [83]. Interestingly, the isolation of monoembryonic early-flowering “Mini-Citrus”, its sequence homology to cultivated citrus species, and its amenability to CRISPR/Cas9 precise editing render it a potent model system to study gene functions in citrus [81]. By projected editing of the DNA sequence of the promoter or coding region of citrus taste-contributing genes to tune their expression, it would be achievable to improve the citrus fruit quality using CRISPR/Cas strategy in the long run.
4. Conclusions and Perspectives
This review outlines the major genes governing citrus taste properties including sweetness, bitterness, and sourness. The sweet sensation is largely determined by the sucrose content in citrus fruit, and the ratio of TSS/TA confers the dominance of sweet or sour taste of citrus. Sucrose is synthesized in leaves through photosynthesis and transported to citrus fruit, via a multiplayer-involved regulatory pathway. The content of titratable acid, mainly citric acid in the vacuole, delivers the sour taste and its import is greatly facilitated through the H+ gradient across the tonoplast. The characterized V-ATPase and P-ATPase and their regulators play essential roles in citrus acidification. The bitterness is influenced by the branch-forming glycosylation on flavanone-7-glucoside. The activity and availability of 1,2RhaT, 1,6RhaT, and dGlcT determine the bitter or non-bitter taste of citrus. The delayed bitterness is generated mostly by the conversion of non-bitter precursor to bitter limonin, and the glycosylation of limonoid catalyzed by limonoid glycosyltransferase (LGT) can act to debitter. The molecular evidence of these core taste-shaping genes by detecting their existence in citrus varieties would help evaluate the citrus fruit quality at juvenile stages. In addition, these genes can serve as a guide for genetic engineering for citrus taste modification.
The flavor of citrus fruit and juice is perceived and evaluated by our sense organs through their appearance (color and texture), aroma, and the juice and solid contents [84]. Other physical properties such as fruit size and shape, peel strength and thickness, and seedless also contribute to the customer acceptance of citrus fresh fruits [85]. The genetic makeup of flavor-forming pathways provides the basis for citrus flavor, while other favorable conditions such as temperature, fertilization and irrigation schedule, and harvest timing should be met as well for optimal citrus flavor and production. With the ever-growing knowledge about the underlying molecular mechanism shaping the citrus flavor and the advances in molecular genetic tools and techniques, the citrus breeding would be projected to grow faster than ever to provide consumers a variety of healthy and tasty citrus products.
Author Contributions
Funding
This work was funded by the Natural Science Foundation of China (No. U1805235) and the Science and Technology Project of Xiamen City (No. 3502Z20183029).
Acknowledgments
Thank Wen-Guang Zheng very much for helping improve English writing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Frydman, A.; Liberman, R.; Huhman, D.V.; Carmeli-Weissberg, M.; Sapir-Mir, M.; Ophir, R.; L, W.S.; Eyal, Y. The molecular and enzymatic basis of bitter/non-bitter flavor of citrus fruit: Evolution of branch-forming rhamnosyltransferases under domestication. Plant J. 2013, 73, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.A.; Terol, J.; Ibanez, V.; Lopez-Garcia, A.; Perez-Roman, E.; Borreda, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Heying, E.; Tanumihardjo, S.A. History, global distribution, and nutritional importance of citrus fruits. Compr. Rev. Food Sci. Food Saf. 2012, 11, 530–545. [Google Scholar] [CrossRef]
- Shimizu, T.; Kitajima, A.; Nonaka, K.; Yoshioka, T.; Ohta, S.; Goto, S.; Toyoda, A.; Fujiyama, A.; Mochizuki, T.; Nagasaki, H.; et al. Hybrid origins of citrus varieties inferred from dna marker analysis of nuclear and organelle genomes. PLoS ONE 2016, 11, e0166969. [Google Scholar] [CrossRef]
- Bharate, S.S.; Bharate, S.B. Non-enzymatic browning in citrus juice: Chemical markers, their detection and ways to improve product quality. J. Food Sci. Technol. 2014, 51, 2271–2288. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, R.; Kim, K.S. Anticancer phytochemicals of citrus fruits—A review. J. Anim. Res. 2014, 4, 85–95. [Google Scholar] [CrossRef]
- Klee, H.J. Improving the flavor of fresh fruits: Genomics, biochemistry, and biotechnology. New Phytol. 2010, 187, 44–56. [Google Scholar] [CrossRef]
- Kore, V.T.; Chakraborty, I. Efficacy of various techniques on biochemical characteristics and bitterness of pummelo juice. J. Food Sci. Technol. 2015, 52, 6073–6077. [Google Scholar] [CrossRef] [Green Version]
- Talon, M.; Gmitter, F.G., Jr. Citrus genomics. Int. J. Plant Genom. 2008, 2008, 17. [Google Scholar] [CrossRef]
- Echeverria, E.; Ismail, M. Sugars unrelated to brix changes in stored citrus fruits. HortScience 1990, 25, 710. [Google Scholar] [CrossRef]
- Yamaki, S. Metabolism and accumulation of sugars translocated to fruit and their regulation. J. Japan. Soc. Hort. Sci. 2010, 79, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Stein, O.; Granot, D. An overview of sucrose synthases in plants. Front. Plant Sci. 2019, 10, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; Wang, C.; Dong, W.; Jiang, Q.; Wang, D.; Li, S.; Chen, M.; Liu, C.; Sun, C.; Chen, K. Transcriptome and metabolome analyses of sugar and organic acid metabolism in Ponkan (Citrus reticulata) fruit during fruit maturation. Gene 2015, 554, 64–74. [Google Scholar] [CrossRef]
- Lowell, C.A.; Tomlinson, P.T.; Koch, K.E. Sucrose-metabolizing enzymes in transport tissues and adjacent sink structures in developing citrus fruit. Plant Physiol. 1989, 90, 1394–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, N. Molecular physiology of higher plant sucrose transporters. FEBS Lett. 2007, 581, 2309–2317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etxeberria, E.; Gonzalez, P. Sucrose transport into citrus juice cells: Evidence for an endocytic transport system. J. Am. Soc. Hort. Sci. 2005, 130, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, A.; Moriguchi, T.; Koyama, K.; Omura, M.; Akihama, T. Analysis of sucrose synthase genes in citrus suggests different roles and phylogenetic relationships. J. Exp. Bot. 2002, 53, 61–71. [Google Scholar] [CrossRef]
- Islam, M.Z.; Jin, L.-F.; Shi, C.-Y.; Liu, Y.-Z.; Peng, S.-A. Citrus sucrose transporter genes: Genome-wide identification and transcript analysis in ripening and ABA-injected fruits. Tree Genet. Genomes 2015, 11, 97. [Google Scholar] [CrossRef]
- Islam, M.Z.; Hu, X.M.; Jin, L.F.; Liu, Y.Z.; Peng, S.A. Genome-wide identification and expression profile analysis of citrus sucrose synthase genes: Investigation of possible roles in the regulation of sugar accumulation. PLoS ONE 2014, 9, e113623. [Google Scholar] [CrossRef] [Green Version]
- Omura, M.; Shimada, T. Citrus breeding, genetics and genomics in Japan. Breed. Sci. 2016, 66, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Katz, E.; Boo, K.H.; Kim, H.Y.; Eigenheer, R.A.; Phinney, B.S.; Shulaev, V.; Negre-Zakharov, F.; Sadka, A.; Blumwald, E. Label-free shotgun proteomics and metabolite analysis reveal a significant metabolic shift during citrus fruit development. J. Exp. Bot. 2011, 62, 5367–5384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kore, V.T.; Tawade, S.S.; Devi, L.H.; Chakraborty, I. Use of pre and post harvest low cost techniques to control/minimize citrus juice bitterness. Am. Res. J. Food Nutr. 2018, 1, 11–21. [Google Scholar]
- Puri, M.; MarwahaJ, S.S.; KothariJ, R.M.; Kennedy, J.F. Biochemical basis of bitterness in citrus fruit juices and biotech approaches for debittering. Crit. Rev. Biotechnol. 1996, 16, 145–155. [Google Scholar] [CrossRef]
- Raithore, S.; Dea, S.; McCollum, G.; Manthey, J.A.; Bai, J.; Leclair, C.; Hijaz, F.; Narciso, J.A.; Baldwin, E.A.; Plotto, A. Development of delayed bitterness and effect of harvest date in stored juice from two complex citrus hybrids. J. Sci. Food Agric. 2016, 96, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Li, H.; Wang, H. Delayed bitterness of citrus wine is removed through the selection of fining agents and fining optimization. Front. Chem. 2019, 7. [Google Scholar] [CrossRef]
- Zaare-Nahandi, F.; Hosseinkhani, S.; Zamani, Z.; Asadi-Abkenar, A.; Omidbaigi, R. Delay expression of limonoid UDP-glucosyltransferase makes delayed bitterness in citrus. Biochem. Biophys. Res. Commun. 2008, 371, 59–62. [Google Scholar] [CrossRef]
- Frydman, A.; Weisshaus, O.; Bar-Peled, M.; Huhman, D.V.; Sumner, L.W.; Marin, F.R.; Lewinsohn, E.; Fluhr, R.; Gressel, J.; Eyal, Y. Citrus fruit bitter flavors: Isolation and functional characterization of the gene Cm1,2RhaT encoding a 1,2 rhamnosyltransferase, a key enzyme in the biosynthesis of the bitter flavonoids of citrus. Plant J. 2004, 40, 88–100. [Google Scholar] [CrossRef]
- Lewinsohn, E.; Britsch, L.; Mazur, Y.; Gressel, J. Flavanone glycoside biosynthesis in citrus: Chalcone synthase, UDP-glucose: flavanone-7-o-glucosyl-transferase and -rhamnosyl-transferase activities in cell-free extracts. Plant Physiol. 1989, 91, 1323–1328. [Google Scholar] [CrossRef] [Green Version]
- Stevens, Y.; Van Rymenant, E.; Grootaert, C.; Van Camp, J.; Possemiers, S.; Masclee, A.; Jonkers, D. The intestinal fate of citrus flavanones and their effects on gastrointestinal health. Nutrients 2019, 11, 1464. [Google Scholar] [CrossRef] [Green Version]
- Li, L.J.; Wu, Z.Y.; Yu, Y.; Zhang, L.J.; Zhu, Y.B.; Ni, H.; Chen, F. Development and characterization of an alpha-l-rhamnosidase mutant with improved thermostability and a higher efficiency for debittering orange juice. Food Chem. 2018, 245, 1070–1078. [Google Scholar] [CrossRef]
- Bar-Peled, M.; Fluhr, R.; Gressel, J. Juvenile-specific localization and accumulation of a rhamnosyltransferase and its bitter flavonoid in foliage, flowers, and young citrus fruits. Plant Physiol. 1993, 103, 1377–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berhow, M.A.; Tisserat, B.; Kanes, K.; Vandercook, C. Survey of phenolic compounds produced in citrus. In US Department of Agriculture/Agricultural Research Service; Bulletin, T., Ed.; US Government Printing Office: Washington, DC, USA, 1998. [Google Scholar]
- Ohashi, T.; Hasegawa, Y.; Misaki, R.; Fujiyama, K. Substrate preference of citrus naringenin rhamnosyltransferases and their application to flavonoid glycoside production in fission yeast. Appl. Microbiol. Biotechnol. 2016, 100, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yuan, Z.; Zhang, H.; Li, W.; Shi, M.; Peng, Z.; Li, M.; Tian, J.; Deng, X.; Cheng, Y.; et al. Cit1,2RhaT and two novel CitdGlcTs participate in flavor-related flavonoid metabolism during citrus fruit development. J. Exp. Bot. 2019, 70, 2759–2771. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Liu, X.; Xiong, B.; Qiu, X.; Sun, G.; Wang, X.; Zhang, X.; Dong, Z.; Wang, Z. Variation in limonin and nomilin content in citrus fruits of eight varieties determined by modified HPLC. Food Sci. Biotechnol. 2019, 28, 641–647. [Google Scholar] [CrossRef]
- Breksa, A.P.; Zukas, A.A.; Manners, G.D. Determination of limonoate and nomilinoate A-ring lactones in citrus juices by liquid chromatography-electrospray ionization mass spectrometry. J. Chromatogr. A 2005, 1064, 187–191. [Google Scholar] [CrossRef]
- Maier, V.P.; Hasegawa, S.; Bennett, R.D.; Echols, L.C. Limonin and limonoids chemistry, biochemistry, and juice bitterness. Citrus Nutr. Qual. 1980, 143, 63–82, Chapter 4. [Google Scholar]
- Kita, M.; Endo, T.; Shimada, T.; Moriguchi, T.; Hirata, Y.; Hasegawa, S.; Omura, M. Allelic structures of UDP-glucose: Limonoid glucosyltransferase affect limonoid bitterness in Citrus unshiu and C. sinensis. Euphytica 2003, 132, 87–94. [Google Scholar] [CrossRef]
- Arora, S.; Mohanpuria, P.; Sidhu, G.S.; Yadav, I.S.; Kumari, V. Cloning and characterization of limonoid glucosyltransferase from Kinnow Mandarin (Citrus reticulata Blanco). Food Technol. Biotechnol. 2018, 56, 228–237. [Google Scholar] [CrossRef]
- Endo, T.; Kita, M.; Shimada, T.; Moriguchi, T.; Hidaka, T.; Matsumoto, R.; Hasegawa, S.; Omura, M. Modification of limonoid metabolism in suspension cell culture of citrus. Plant Biotechnol. 2002, 19, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Herman, Z.; Fong, C.H.; Ou, P.; Hasegawa, S. Limonoid glucosides in orange juices by HPLC. J. Agric. Food Chem. 1990, 38, 1860–1861. [Google Scholar] [CrossRef]
- Vikram, A.; Jayaprakasha, G.K.; Patil, B.S. Simultaneous determination of citrus limonoid aglycones and glucosides by high performance liquid chromatography. Anal. Chim. Acta 2007, 590, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Ohta, H.; Nogata, Y.; Yano, M.; Hasegawa, S. Limonoids in seeds of iyo tangor (Citrus iyo hort. ex Tanaka). Food Sci. Technol. Res. 2003, 9, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Kita, M.; Hirata, Y.; Moriguchi, T.; Endo-Inagaki, T.; Matsumoto, R.; Hasegawa, S.; Suhayda, C.G.; Omura, M. Molecular cloning and characterization of a novel gene encoding limonoid UDP-glucosyltransferase in Citrus. FEBS Lett. 2000, 469, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Etienne, A.; Genard, M.; Lobit, P.; Mbeguie, A.M.D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.H.; Liu, J.J.; Chen, K.L.; Li, H.W.; He, J.; Guan, B.; He, L. Comparative transcriptome and proteome profiling of two Citrus sinensis cultivars during fruit development and ripening. BMC Genom. 2017, 18. [Google Scholar] [CrossRef] [Green Version]
- Goldenberg, L.; Yaniv, Y.; Porat, R.; Carmi, N. Mandarin fruit quality: A review. J. Sci. Food Agric. 2018, 98, 18–26. [Google Scholar] [CrossRef]
- Al-Mouei, R.; Choumane, W. Physiochemical juice characteristics of various citrus species in Syria. Int. J. Plant Soil Sci. 2014, 3, 1083–1095. [Google Scholar] [CrossRef]
- Güler, Z.; Gül, E. Volatile organic compounds in the aril juices and seeds from selected five pomegranate (Punica granatum L.) cultivars. Int. J. Food Prop. 2017, 20, 281–293. [Google Scholar] [CrossRef]
- Butelli, E.; Licciardello, C.; Ramadugu, C.; Durand-Hulak, M.; Celant, A.; Reforgiato Recupero, G.; Froelicher, Y.; Martin, C. Noemi controls production of flavonoid pigments and fruit acidity and illustrates the domestication routes of modern citrus varieties. Curr. Biol. 2019, 29, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Terol, J.; Soler, G.; Talon, M.; Cercos, M. The aconitate hydratase family from Citrus. BMC Plant Biol. 2010, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cercos, M.; Soler, G.; Iglesias, D.J.; Gadea, J.; Forment, J.; Talon, M. Global analysis of gene expression during development and ripening of citrus fruit flesh. A proposed mechanism for citric acid utilization. Plant Mol. Biol. 2006, 62, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Degu, A.; Hatew, B.; Nunes-Nesi, A.; Shlizerman, L.; Zur, N.; Katz, E.; Fernie, A.R.; Blumwald, E.; Sadka, A. Inhibition of aconitase in citrus fruit callus results in a metabolic shift towards amino acid biosynthesis. Planta 2011, 234, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Brune, A.; Gonzalez, P.; Goren, R.; Zehavi, U.; Echeverria, E. Citrate uptake into tonoplast vesicles from acid lime (Citrus aurantifolia) juice cells. J. Membr. Biol. 1998, 166, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, R. Structure, function and regulation of the plant vacuolar H+-translocating ATPase. Biochim. Biophys. Acta 2000, 1465, 17–36. [Google Scholar] [CrossRef] [Green Version]
- Li, S.J.; Yin, X.R.; Xie, X.L.; Allan, A.C.; Ge, H.; Shen, S.L.; Chen, K.S. The Citrus transcription factor, CitERF13, regulates citric acid accumulation via a protein-protein interaction with the vacuolar proton pump, CitVHA-c4. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, T.; Nakano, R.; Shulaev, V.; Sadka, A.; Blumwald, E. Vacuolar citrate/H+ symporter of citrus juice cells. Planta 2006, 224, 472–480. [Google Scholar] [CrossRef]
- Brune, A. Vacuolar acidification in citrus fruit: Comparison between acid lime (Citrus aurantifolia) and sweet lime (Citrus limmetioides) juice cells. J. Am. Soc. Hort. Sci. 2002, 127, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.; Irkens-Kiesecker, U.; Rubinstein, B.; Taiz, L. On the mechanism of hyperacidification in lemon. Comparison of the vacuolar H+-ATPase activities of fruits and epicotyls. J. Biol. Chem. 1996, 271, 1916–1924. [Google Scholar] [CrossRef] [Green Version]
- Faraco, M.; Spelt, C.; Bliek, M.; Verweij, W.; Hoshino, A.; Espen, L.; Prinsi, B.; Jaarsma, R.; Tarhan, E.; de Boer, A.H.; et al. Hyperacidification of vacuoles by the combined action of two different P-ATPases in the tonoplast determines flower color. Cell Rep. 2014, 6, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Strazzer, P.; Spelt, C.E.; Li, S.; Bliek, M.; Federici, C.T.; Roose, M.L.; Koes, R.; Quattrocchio, F.M. Hyperacidification of Citrus fruits by a vacuolar proton-pumping P-ATPase complex. Nat. Commun. 2019, 10, 744. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ye, J.; Liu, C.; Xu, Q.; Long, L.; Deng, X. The citrus CsPH4-Noemi regulatory complex is involved in proanthocyanidin biosynthesis via a positive feedback loop. J. Exp. Bot. 2019. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.Y.; Hussain, S.B.; Yang, H.; Bai, Y.X.; Khan, M.A.; Liu, Y.Z. CsPH8, a P-type proton pump gene, plays a key role in the diversity of citric acid accumulation in Citrus fruits. Plant Sci. 2019, 289, 110288. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.Y.; Song, R.Q.; Hu, X.M.; Liu, X.; Jin, L.F.; Liu, Y.Z. Citrus PH5-like H+-ATPase genes: Identification and transcript analysis to investigate their possible relationship with citrate accumulation in fruits. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liang, M.; Wang, N.; Xu, Q.; Deng, X.; Chai, L. Reproduction in woody perennial Citrus: An update on nucellar embryony and self-incompatibility. Plant Reprod. 2018, 31, 43–57. [Google Scholar] [CrossRef]
- Endo, T.; Shimada, T.; Fujii, H.; Kobayashi, Y.; Araki, T.; Omura, M. Ectopic expression of an FT homolog from citrus confers an early flowering phenotype on trifoliate orange (Poncirus trifoliata L. Raf.). Transgenic Res. 2005, 14, 703–712. [Google Scholar] [CrossRef]
- Dutt, M.; Grosser, J. Using genetically modified biotechnology to improve citrus. Citrus Ind. 2005, 10–13. Available online: https://crec.ifas.ufl.edu/extension/trade_journals/2015/2015_September_biotech.pdf (accessed on 31 October 2019).
- Velazquez, K.; Aguero, J.; Vives, M.C.; Aleza, P.; Pina, J.A.; Moreno, P.; Navarro, L.; Guerri, J. Precocious flowering of juvenile citrus induced by a viral vector based on Citrus leaf blotch virus: A new tool for genetics and breeding. Plant Biotechnol. J. 2016, 14, 1976–1985. [Google Scholar] [CrossRef] [Green Version]
- Iwata, H.; Minamikawa, M.F.; Kajiya-Kanegae, H.; Ishimori, M.; Hayashi, T. Genomics-assisted breeding in fruit trees. Breed. Sci. 2016, 66, 100–115. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Bai, J.; Chen, C.; Plotto, A.; Yu, Q.; Baldwin, E.A.; Gmitter, F.G., Jr. Identification of QTLs controlling aroma volatiles using a ‘Fortune’ × ‘Murcott’ (Citrus reticulata) population. BMC Genom. 2017, 18. [Google Scholar] [CrossRef]
- Rommens, C.M. All-native DNA transformation: A new approach to plant genetic engineering. Trends Plant Sci. 2004, 9, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons control fruit-specific, cold-dependent accumulation of anthocyanins in blood oranges. Plant Cell 2012, 24, 1242–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Acanda, Y.; Canton, M.; Zale, J. Efficient biolistic transformation of immature citrus rootstocks using phosphomannose-isomerase selection. Plants 2019, 8, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeffer, S.M.; Nakata, P.A. The expanding footprint of CRISPR/Cas9 in the plant sciences. Plant Cell Rep. 2016, 35, 1451–1468. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Wang, N. Targeted genome editing of sweet orange using Cas9/sgRNA. PLoS ONE 2014, 9, e93806. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Zou, X.; Orbovic, V.; Wang, N. Genome editing in citrus tree with CRISPR/Cas9. Methods Mol. Biol. 2019, 1917, 235–241. [Google Scholar]
- Jia, H.; Xu, J.; Orbovic, V.; Zhang, Y.; Wang, N. Editing citrus genome via SaCas9/sgRNA system. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Zhang, Y.; Orbovic, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Zheng, X.; Huang, Y.; Ye, J.; Chen, P.; Zhang, C.; Zhao, F.; Xie, Z.; Zhang, S.; Wang, N.; et al. Genome sequencing and CRISPR/Cas9 gene editing of an early flowering Mini-Citrus (Fortunella hindsii). Plant Biotechnol. J. 2019, 17, 2199–2210. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Orbovic, V.; Wang, N. CRISPR-LbCas12a-mediated modification of citrus. Plant Biotechnol. J. 2019, 17, 1928–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Ke, F.; Nie, Z.; Wang, P.; Xu, J. Citrus genetic engineering for disease resistance: Past, present and future. Int. J. Mol. Sci. 2019, 20, 5256. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Bai, J.; Chen, C.; Plotto, A.; Baldwin, E.A.; Gmitter, F.G. Comparative analysis of juice volatiles in selected mandarins, mandarin relatives and other citrus genotypes. J. Sci. Food Agric. 2018, 98, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Abouzari, A.; Mahdi Nezhad, N. The investigation of citrus fruit quality. Popular characteristic and breeding. Acta Univ. Agric. Silvic. Mendel. Brun. 2016, 64, 725–740. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Proposed family tree of citrus species. The three ancestral citrus species of the main citrus species on the market include mandarin, pumelo, and citron. The commercial mandarins consist of tangerine, satsuma, and clementine, and they are hybrids between mandarin and pomelo. Satsuma is a highly inbred line from the hybrid of mandarin and pomelo. Clementine is the hybrid between sweet orange and willowleaf mandarin which are also the hybrid between mandarin and pomelo. Bitter orange is also a hybrid between mandarin and pomelo. Grapefruit is a hybrid between sweet orange and pomelo. Lemon is a hybrid between bitter orange and citron.
Figure 1.
Proposed family tree of citrus species. The three ancestral citrus species of the main citrus species on the market include mandarin, pumelo, and citron. The commercial mandarins consist of tangerine, satsuma, and clementine, and they are hybrids between mandarin and pomelo. Satsuma is a highly inbred line from the hybrid of mandarin and pomelo. Clementine is the hybrid between sweet orange and willowleaf mandarin which are also the hybrid between mandarin and pomelo. Bitter orange is also a hybrid between mandarin and pomelo. Grapefruit is a hybrid between sweet orange and pomelo. Lemon is a hybrid between bitter orange and citron.

Figure 2.
Schematic illustration of sucrose transport from source to sink in citrus. Sucrose is biosynthesized in leaf mesophyll cells through photosynthesis. The translocate sucrose is loaded to the sieve cells of phloem using H+ electrochemical potential gradient as driving force with the help of H+/sucrose symporter. Sucrose is transported in phloem following the turgor pressure in sieve elements towards sink tissue (citrus fruit) and unloaded by the symplastic or apoplastic pathway. Sucrose can be converted to fructose and glucose by IVR or fructose and UDP-glucose by SS, and it can be resynthesized through fructose and UDP-glucose by SPS in the cytosol. Sucrose uptake from apoplastic into cytosol is driven by the H+/sucrose symporter. The apoplastic sucrose can be directly incorporated into vacuole through endocytosis system, while the existence of an active transporter or H+/sucrose antiporter from cytosol to vacuole is still questioning. IVR—invertase; SS—sucrose synthase; SPS—sucrose-phosphate synthase.
Figure 2.
Schematic illustration of sucrose transport from source to sink in citrus. Sucrose is biosynthesized in leaf mesophyll cells through photosynthesis. The translocate sucrose is loaded to the sieve cells of phloem using H+ electrochemical potential gradient as driving force with the help of H+/sucrose symporter. Sucrose is transported in phloem following the turgor pressure in sieve elements towards sink tissue (citrus fruit) and unloaded by the symplastic or apoplastic pathway. Sucrose can be converted to fructose and glucose by IVR or fructose and UDP-glucose by SS, and it can be resynthesized through fructose and UDP-glucose by SPS in the cytosol. Sucrose uptake from apoplastic into cytosol is driven by the H+/sucrose symporter. The apoplastic sucrose can be directly incorporated into vacuole through endocytosis system, while the existence of an active transporter or H+/sucrose antiporter from cytosol to vacuole is still questioning. IVR—invertase; SS—sucrose synthase; SPS—sucrose-phosphate synthase.

Figure 3.
Flavanone glycosylation pathways, and limonoid aglycon or glycon formation pathways. (a) Two major flavanone compounds, naringenin and hesperitin, in citrus are exemplified. They can be converted to their glucoside derivatives at their C7 site by 7GlcT (7-glucose transferase), and further rahmnosylated on glucoside through C1,2 or C1,6 bond formation between rhamnose and glucose catalyzed by 1,2RhaT or 1,6RhaT, respectively. Consequently, the glycosylation of flavanone by neohesperidose (rhamylose-1,2-glucose) leading to the formation of naringin and neohesperidin confers bitterness and the glycosylation by rutinose (rhamylose-1,6-glucose) leading to the formation of narirutin and hesperidin confers non-bitterness. Flavanone-7-O-gluc can also be further glucosylated to form flavanone-7-O-di-glucocide (glucose-1,2-glucose as proposed) catalyzed by dGlcT; (b) Two major limonoid compounds, nomilin and limonin, in citrus are exemplified. Non-bitter NARL and LARL precursors can be converted to bitter-tasting nomilin and limonin, respectively, by LLH under acidic condition. Nomilin can be converted to LARL through multiple steps. Nomilin and LARL can also be converted to the non-bitter derivatives of nomilin-17-O-glucoside and limonin-17-O-glucoside, respectively, catalyzed by LGT. NARL—nomilioate A-ring lactone; LARL—limonoate A-ring lactone; LLH—limonoid D-ring lactone hydrolase; LGT—limonoid glycosyltransferase.
Figure 3.
Flavanone glycosylation pathways, and limonoid aglycon or glycon formation pathways. (a) Two major flavanone compounds, naringenin and hesperitin, in citrus are exemplified. They can be converted to their glucoside derivatives at their C7 site by 7GlcT (7-glucose transferase), and further rahmnosylated on glucoside through C1,2 or C1,6 bond formation between rhamnose and glucose catalyzed by 1,2RhaT or 1,6RhaT, respectively. Consequently, the glycosylation of flavanone by neohesperidose (rhamylose-1,2-glucose) leading to the formation of naringin and neohesperidin confers bitterness and the glycosylation by rutinose (rhamylose-1,6-glucose) leading to the formation of narirutin and hesperidin confers non-bitterness. Flavanone-7-O-gluc can also be further glucosylated to form flavanone-7-O-di-glucocide (glucose-1,2-glucose as proposed) catalyzed by dGlcT; (b) Two major limonoid compounds, nomilin and limonin, in citrus are exemplified. Non-bitter NARL and LARL precursors can be converted to bitter-tasting nomilin and limonin, respectively, by LLH under acidic condition. Nomilin can be converted to LARL through multiple steps. Nomilin and LARL can also be converted to the non-bitter derivatives of nomilin-17-O-glucoside and limonin-17-O-glucoside, respectively, catalyzed by LGT. NARL—nomilioate A-ring lactone; LARL—limonoate A-ring lactone; LLH—limonoid D-ring lactone hydrolase; LGT—limonoid glycosyltransferase.

Figure 4.
Citrate biosynthesis, transport, and accumulation in citrus. Citrate is synthesized in mitochondria through TCA cycle. In mitochondria, citrate can be degraded through TCA cycle initiated by the enzymatic actions of mitochondrial ACO and IDH or through GABA shunt characterized by the functioning of enzymes of GDH and GAD with the shuttle of glutamate and GABA between mitochondria and cytosol. Citrate can be exported from mitochondria to cytosol and further degraded by the action of ACL. Citrate exported from mitochondria to cytosol can be imported to vacuole with the help of H+ electrochemical gradient and pH gradient conferred by the actions of V-ATPase and P-ATPase. Vacuolar citrate can be exported to cytosol by transporters and be degraded by the action of cytosolic ACO and IDH. The integral metabolic pathways affect the citrate content in vacuole and sour taste of citrus fruit. ACO—aconitase; IDH—isocitrate dehydrogenase; GDH—glutamate dehydrogenase; ACL—ATP citrate lyase; GABA—gamma-aminobutyric acid; AST—aspartate aminotransferase.
Figure 4.
Citrate biosynthesis, transport, and accumulation in citrus. Citrate is synthesized in mitochondria through TCA cycle. In mitochondria, citrate can be degraded through TCA cycle initiated by the enzymatic actions of mitochondrial ACO and IDH or through GABA shunt characterized by the functioning of enzymes of GDH and GAD with the shuttle of glutamate and GABA between mitochondria and cytosol. Citrate can be exported from mitochondria to cytosol and further degraded by the action of ACL. Citrate exported from mitochondria to cytosol can be imported to vacuole with the help of H+ electrochemical gradient and pH gradient conferred by the actions of V-ATPase and P-ATPase. Vacuolar citrate can be exported to cytosol by transporters and be degraded by the action of cytosolic ACO and IDH. The integral metabolic pathways affect the citrate content in vacuole and sour taste of citrus fruit. ACO—aconitase; IDH—isocitrate dehydrogenase; GDH—glutamate dehydrogenase; ACL—ATP citrate lyase; GABA—gamma-aminobutyric acid; AST—aspartate aminotransferase.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, L.-J.; Tan, W.-S.; Li, W.-J.; Zhu, Y.-B.; Cheng, Y.-S.; Ni, H. Citrus Taste Modification Potentials by Genetic Engineering. Int. J. Mol. Sci. 2019, 20, 6194. https://doi.org/10.3390/ijms20246194
AMA Style
Li L-J, Tan W-S, Li W-J, Zhu Y-B, Cheng Y-S, Ni H. Citrus Taste Modification Potentials by Genetic Engineering. International Journal of Molecular Sciences. 2019; 20(24):6194. https://doi.org/10.3390/ijms20246194
Chicago/Turabian StyleLi, Li-Jun, Wan-Seng Tan, Wen-Jing Li, Yan-Bing Zhu, Yi-Sheng Cheng, and Hui Ni. 2019. "Citrus Taste Modification Potentials by Genetic Engineering" International Journal of Molecular Sciences 20, no. 24: 6194. https://doi.org/10.3390/ijms20246194
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.