Light Stress-Induced Increase of Sphingosine 1-Phosphate in Photoreceptors and Its Relevance to Retinal Degeneration

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Light-Emitting Diode Exposure Enhanced S1P in Photoreceptor Cells
2.2. All-Trans Retinal Induces the Expression of Sphingosine Kinase (SphK) in Photoreceptor Cells
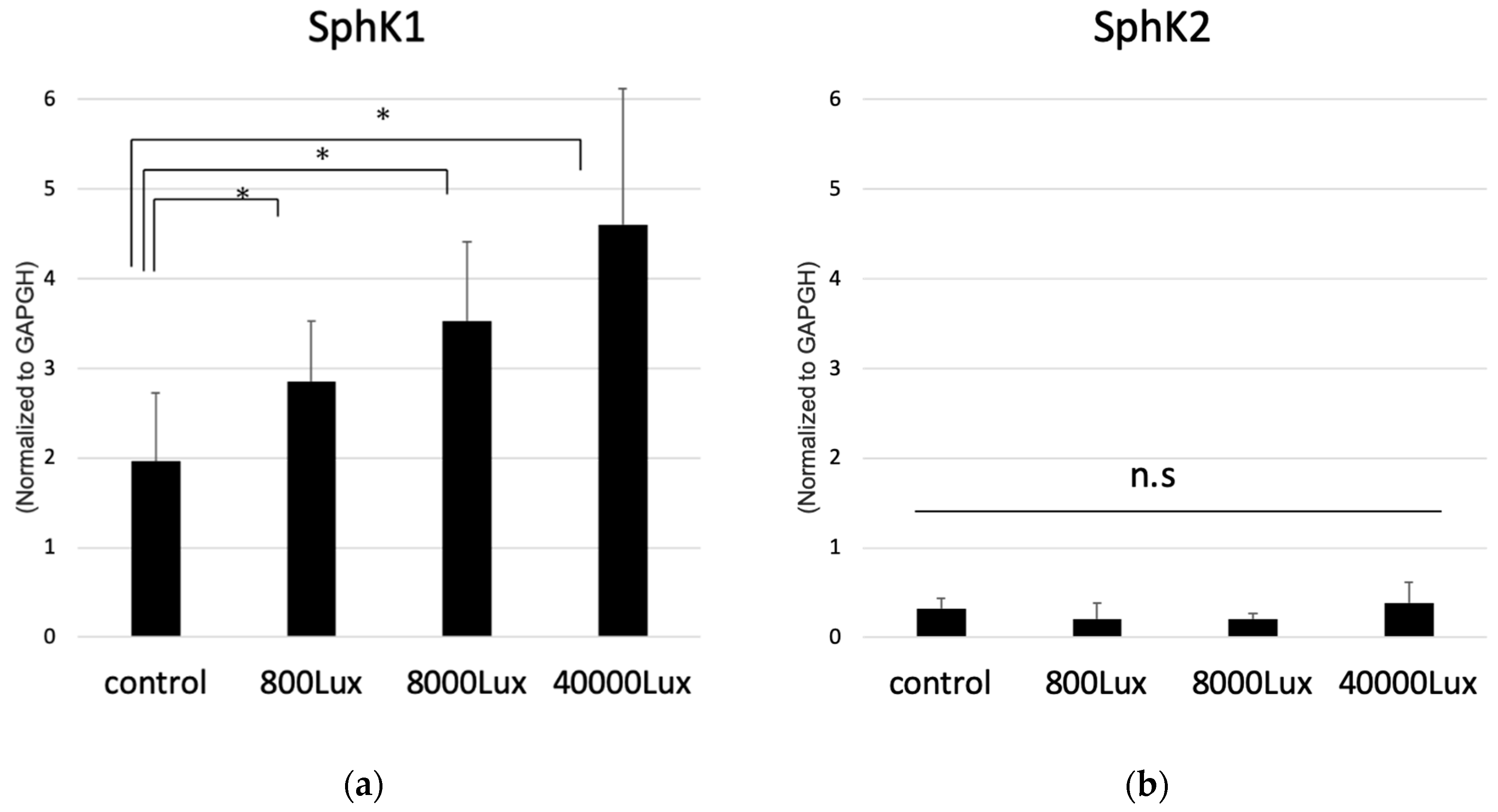
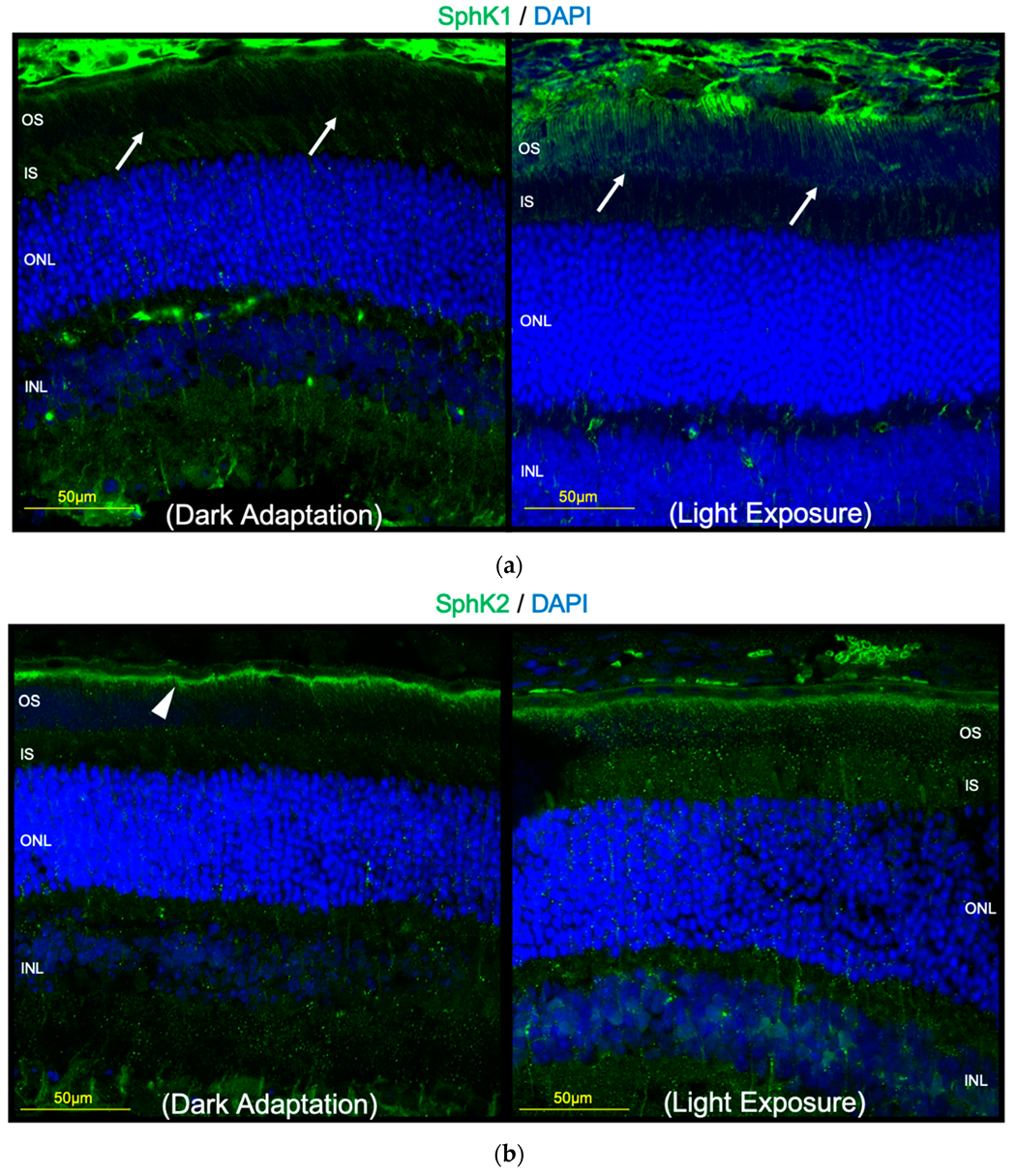
2.3. Light Exposure Increases the Expression of SphK1 in Murine Retina
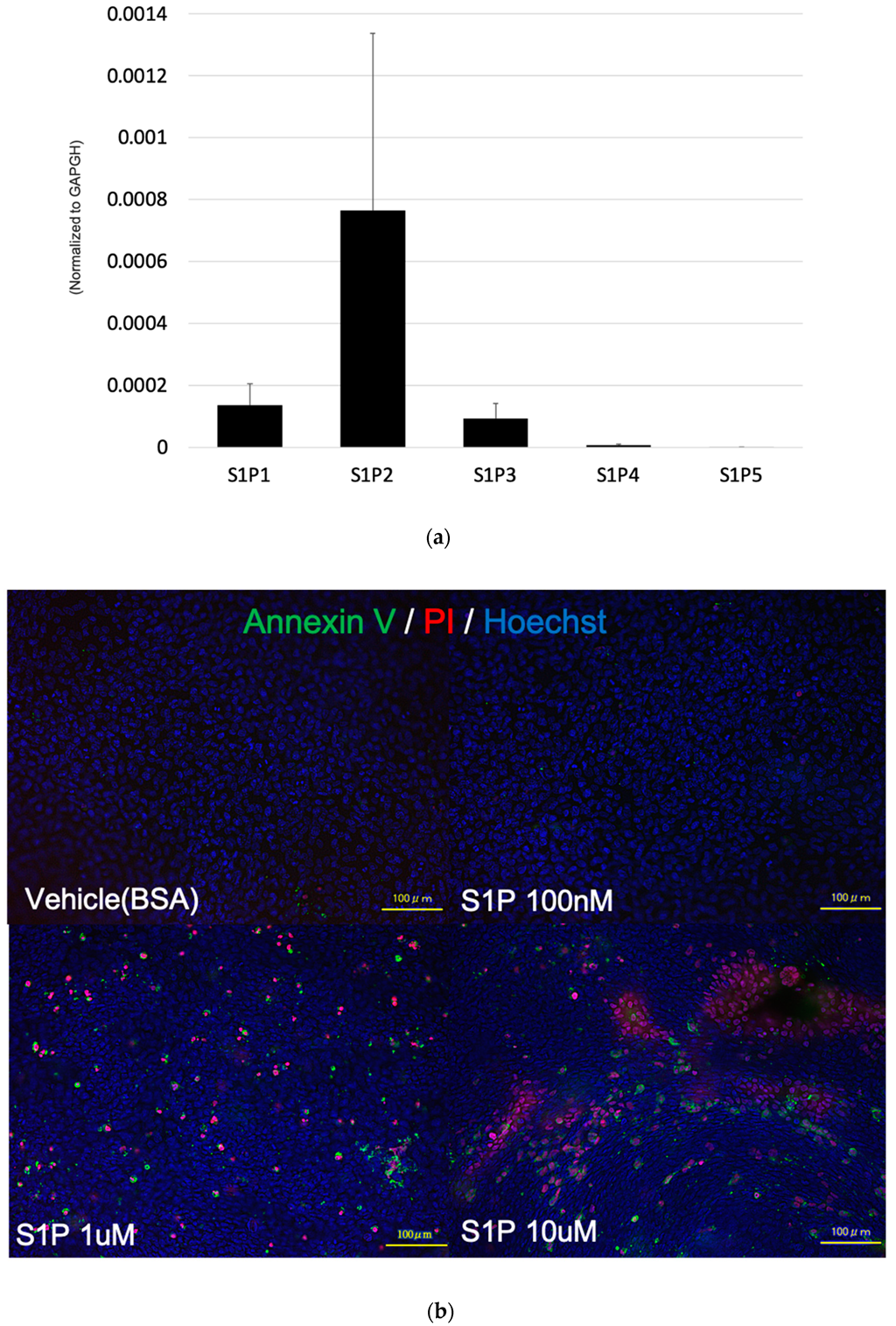
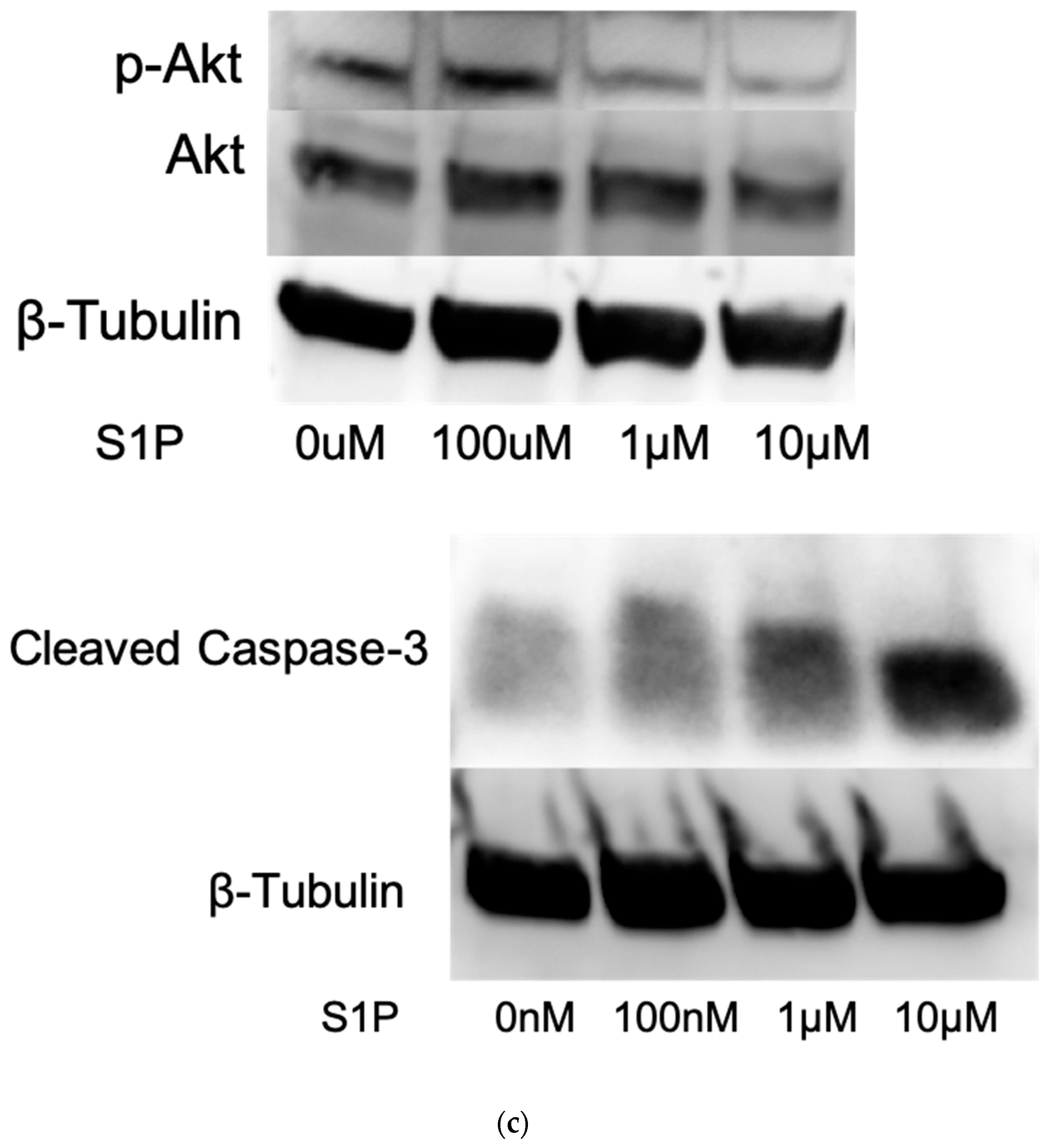
2.4. S1P Induced the Apoptosis of Photoreceptor Cells
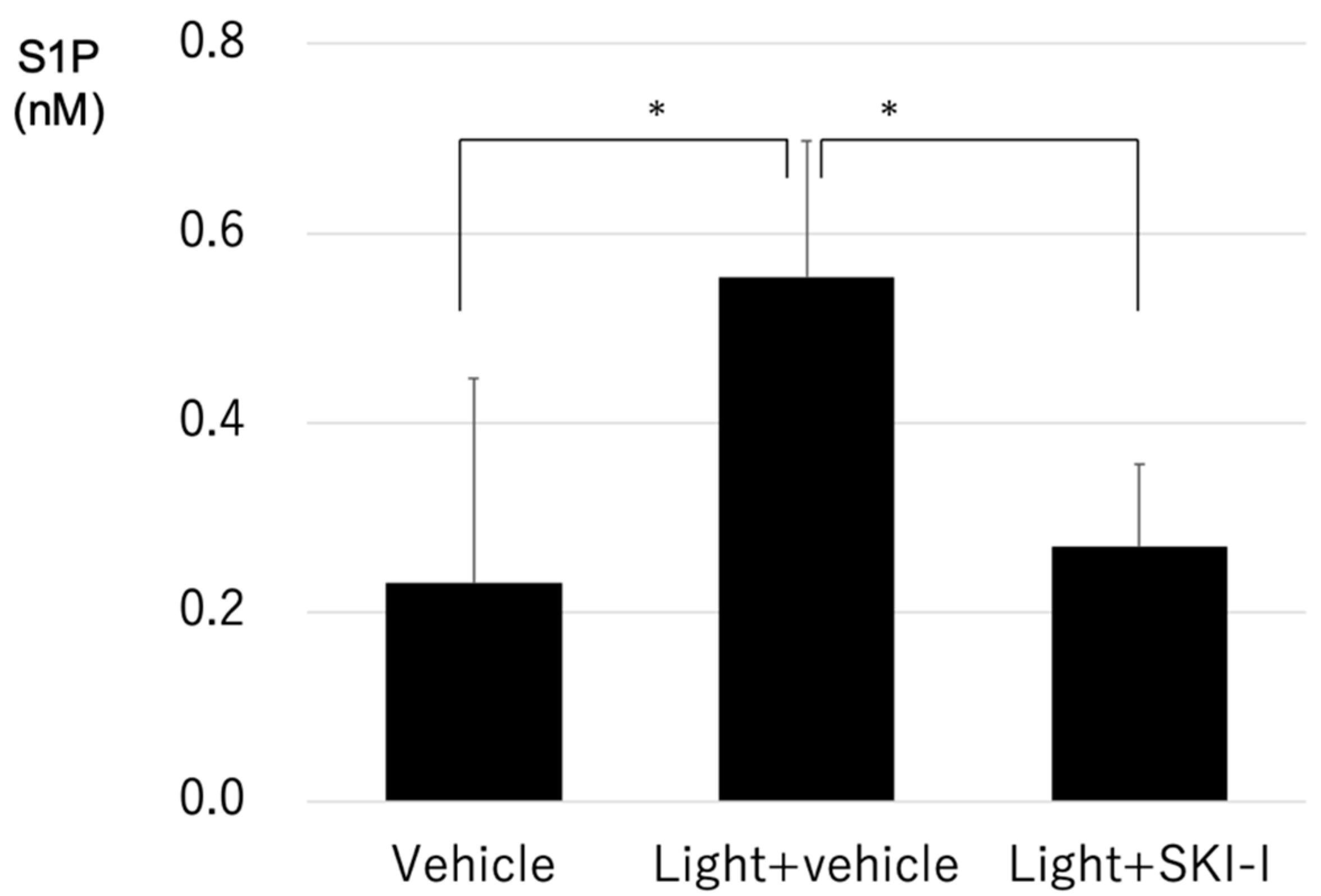
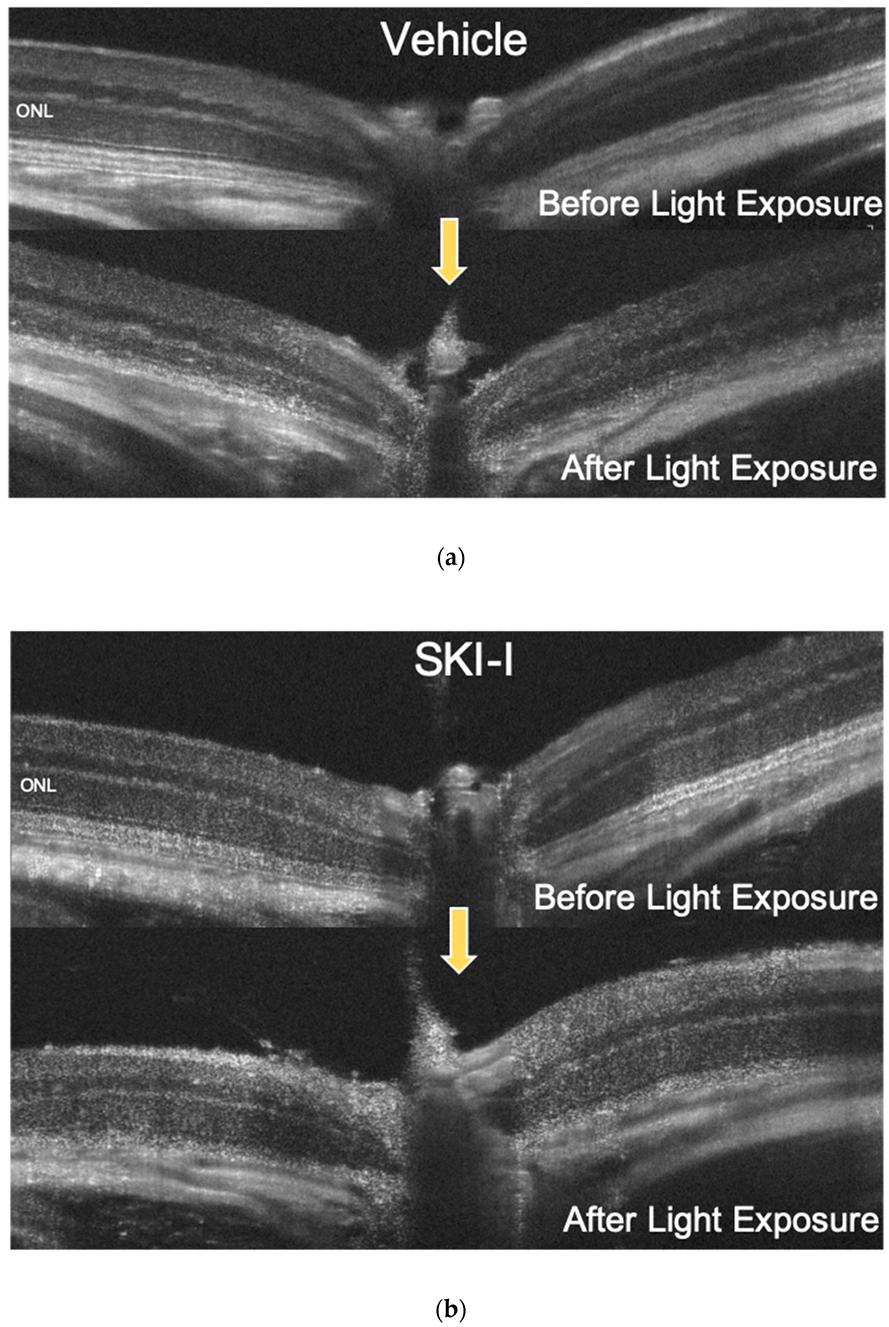
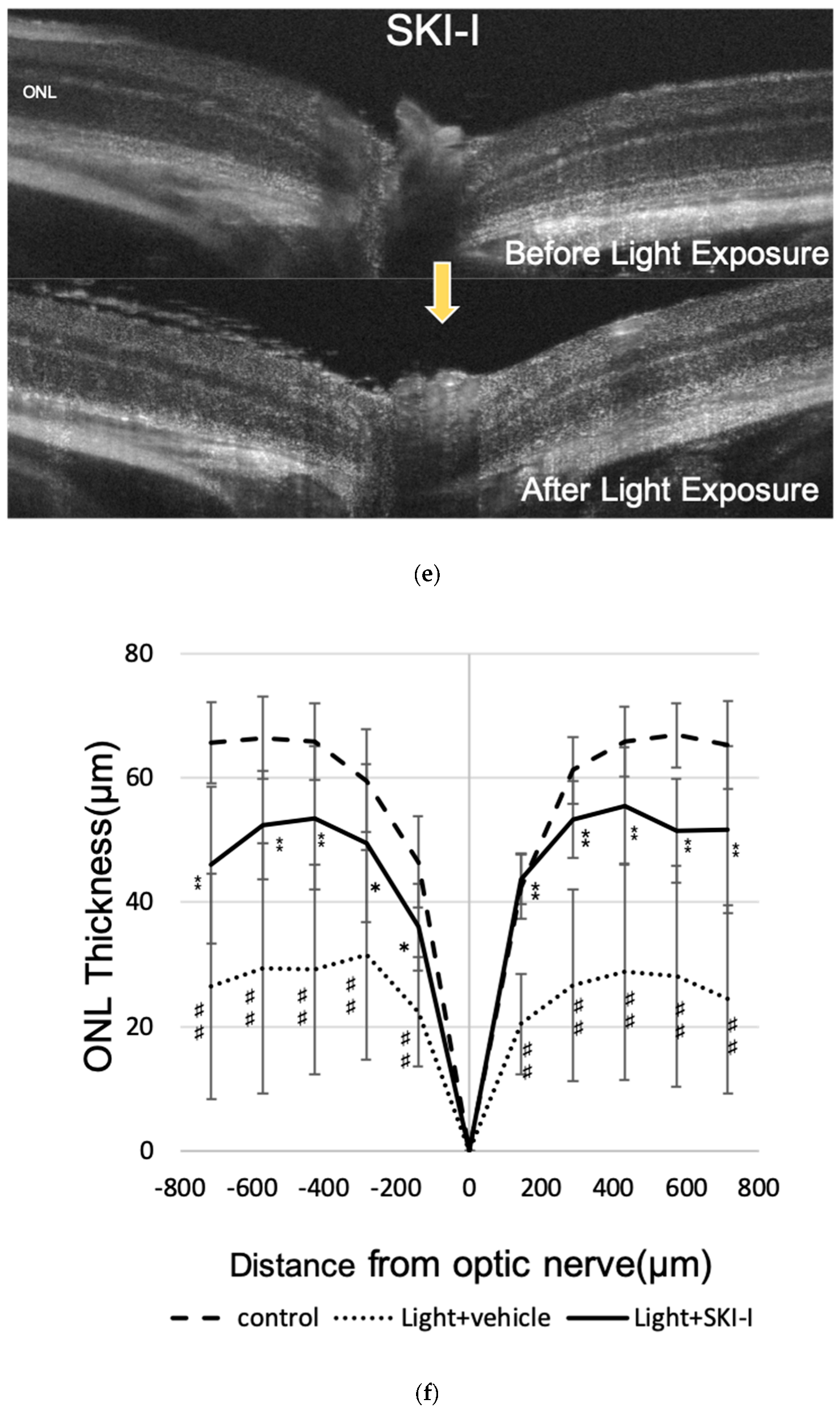
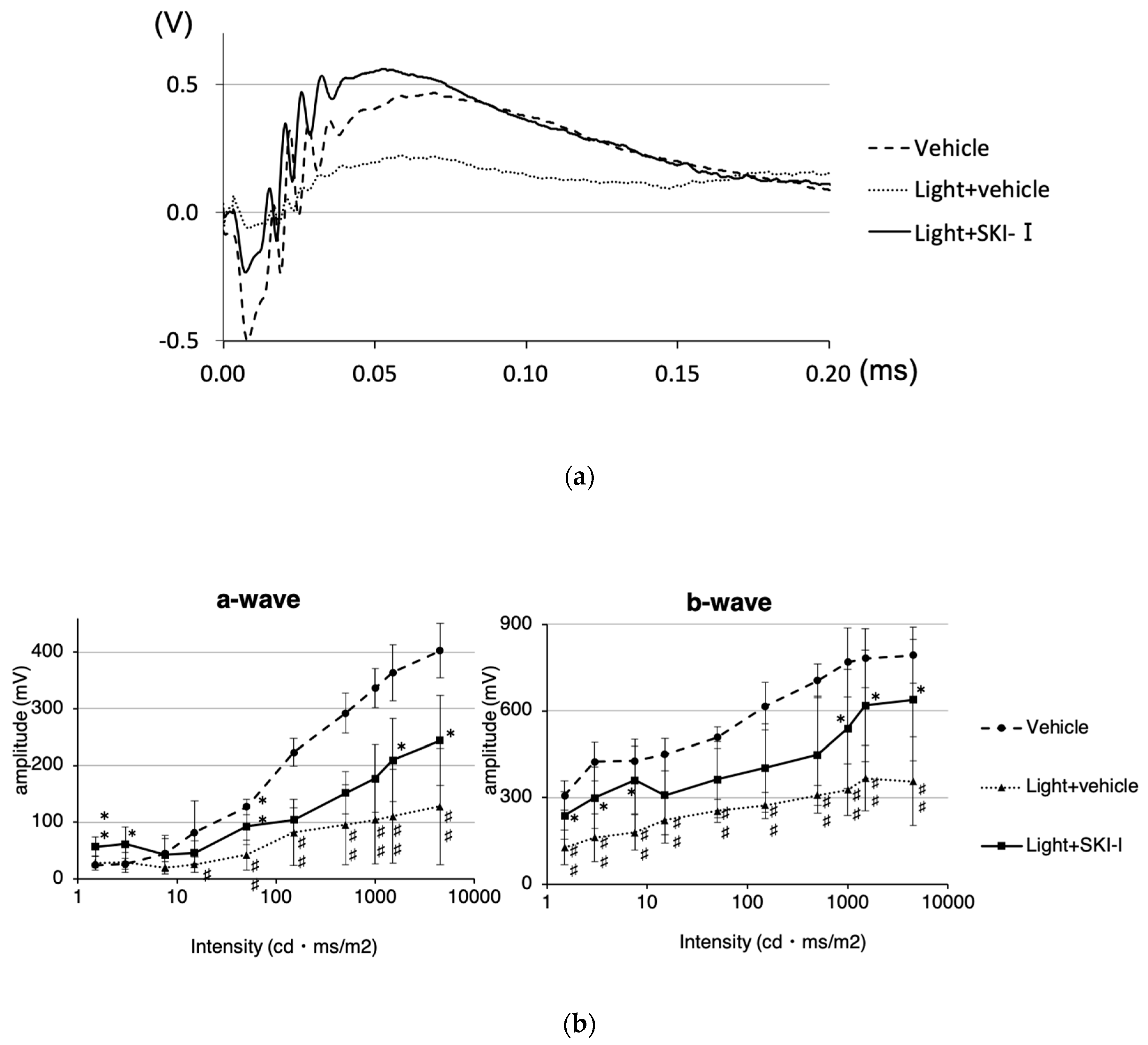
2.5. A SphK Inhibitor Conferred a Protective Effect against LED Light-Induced Retinal Degeneration
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals
4.3. Cell Culture
4.4. Light Exposure
4.5. Measurement of SIP by LC-MS
4.6. qPCR
4.7. Sphingosine Kinase Assay
4.8. Immunohistochemistry
4.9. Apoptosis Assay
4.10. Western Blotting
4.11. OCT
4.12. ERG
4.13. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Ambati, J.; Fowler, B.J. Mechanisms of age-related macular degeneration. Neuron 2012, 75, 26–39. [Google Scholar] [CrossRef]
- De Jong, P.T. Age-related macular degeneration. N. Engl. J. Med. 2006, 355, 1474–1485. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.Z. Oxidative stress, polyunsaturated fatty acids-derived oxidation products and bisretinoids as potential inducers of CNS diseases: Focus on age-related macular degeneration. Pharmacol. Rep. 2013, 65, 288–304. [Google Scholar] [CrossRef]
- Age-Related Eye Disease Study Research Group. A randomized, placebo-controlled, clinical trial of high-dose supplementation with vitamins C and E, beta carotene, and zinc for age-related macular degeneration and vision loss: AREDS report no. 8. Arch. Ophthalmol. 2001, 119, 1417–1436. [Google Scholar] [CrossRef] [PubMed]
- Cruickshanks, K.J.; Klein, R.; Klein, B.E. Sunlight and age-related macular degeneration. The Beaver Dam Eye Study. Arch. Ophthalmol. 1993, 111, 514–518. [Google Scholar] [CrossRef]
- Organisciak, D.T.; Darrow, R.M.; Jiang, Y.I.; Marak, G.E.; Blanks, J.C. Protection by dimethylthiourea against retinal light damage in rats. Investig. Ophthalmol. Vis. Sci. 1992, 33, 1599–1609. [Google Scholar]
- Izawa, H.; Inoue, Y.; Ohno, Y.; Ojino, K.; Tsuruma, K.; Shimazawa, M.; Hara, H. Protective Effects of Antiplacental Growth Factor Antibody Against Light-Induced Retinal Damage in Mice. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6914–6924. [Google Scholar] [CrossRef] [Green Version]
- Gu, R.; Tang, W.; Lei, B.; Ding, X.; Jiang, C.; Xu, G. Glucocorticoid-Induced Leucine Zipper Protects the Retina from Light-Induced Retinal Degeneration by Inducing Bcl-xL in Rats. Investig. Ophthalmol. Vis. Sci. 2017, 58, 3656–3668. [Google Scholar] [CrossRef]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef]
- Sanchez, T.; Hla, T. Structural and functional characteristics of S1P receptors. J. Cell. Biochem. 2004, 92, 913–922. [Google Scholar] [CrossRef]
- Lee, J.F.; Zeng, Q.; Ozaki, H.; Wang, L.; Hand, A.R.; Hla, T.; Wang, E.; Lee, M.J. Dual roles of tight junction-associated protein, zonula occludens-1, in sphingosine 1-phosphate-mediated endothelial chemotaxis and barrier integrity. J. Biol. Chem. 2006, 281, 29190–29200. [Google Scholar] [CrossRef] [PubMed]
- Takabe, K.; Paugh, S.W.; Milstien, S.; Spiegel, S. “Inside-out” signaling of sphingosine-1-phosphate: Therapeutic targets. Pharmacol. Rev. 2008, 60, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Terao, R.; Honjo, M.; Aihara, M. Apolipoprotein M Inhibits Angiogenic and Inflammatory Response by Sphingosine 1-Phosphate on Retinal Pigment Epithelium Cells. Int. J. Mol. Sci. 2017, 19, 112. [Google Scholar] [CrossRef] [PubMed]
- Terao, R.; Honjo, M.; Totsuka, K.; Miwa, Y.; Kurihara, T.; Aihara, M. The role of sphingosine 1-phosphate receptors on retinal pigment epithelial cells and murine choroidal neovascularization. Prostaglandins Lipid Mediat. under reviews.
- Porter, H.; Qi, H.; Prabhu, N.; Grambergs, R.; McRae, J.; Hopiavuori, B.; Mandal, N. Characterizing Sphingosine Kinases and Sphingosine 1-Phosphate Receptors in the Mammalian Eye and Retina. Int. J. Mol. Sci. 2018, 19, 3885. [Google Scholar] [CrossRef] [PubMed]
- Rozanowska, M.; Handzel, K.; Boulton, M.E.; Rozanowski, B. Cytotoxicity of all-trans-retinal increases upon photodegradation. Photochem. Photobiol. 2012, 88, 1362–1372. [Google Scholar] [CrossRef]
- Masutomi, K.; Chen, C.; Nakatani, K.; Koutalos, Y. All-trans retinal mediates light-induced oxidation in single living rod photoreceptors. Photochem. Photobiol. 2012, 88, 1356–1361. [Google Scholar] [CrossRef]
- Goldberg, A.F.; Moritz, O.L.; Williams, D.S. Molecular basis for photoreceptor outer segment architecture. Prog. Retin. Eye Res. 2016, 55, 52–81. [Google Scholar] [CrossRef] [Green Version]
- McBee, J.K.; Palczewski, K.; Baehr, W.; Pepperberg, D.R. Confronting complexity: The interlink of phototransduction and retinoid metabolism in the vertebrate retina. Prog. Retin. Eye Res. 2001, 20, 469–529. [Google Scholar] [CrossRef]
- Chen, Y.; Okano, K.; Maeda, T.; Chauhan, V.; Golczak, M.; Maeda, A.; Palczewski, K. Mechanism of all-trans-retinal toxicity with implications for stargardt disease and age-related macular degeneration. J. Biol. Chem. 2012, 287, 5059–5069. [Google Scholar] [CrossRef]
- Allikmets, R.; Singh, N.; Sun, H.; Shroyer, N.F.; Hutchinson, A.; Chidambaram, A.; Gerrard, B.; Baird, L.; Stauffer, D.; Peiffer, A.; et al. A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat. Genet. 1997, 15, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zulfiqar, F.; Xiao, X.; Riazuddin, S.A.; Ayyagari, R.; Sabar, F.; Caruso, R.; Sieving, P.A.; Riazuddin, S.; Hejtmancik, J.F. Severe autosomal recessive retinitis pigmentosa maps to chromosome 1p13.3-p21.2 between D1S2896 and D1S457 but outside ABCA4. Hum. Genet. 2005, 118, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Maeda, T.; Golczak, M.; Chou, S.; Desai, A.; Hoppel, C.L.; Matsuyama, S.; Palczewski, K. Involvement of all-trans-retinal in acute light-induced retinopathy of mice. J. Biol. Chem. 2009, 284, 15173–15183. [Google Scholar] [CrossRef] [PubMed]
- Guzel, Y.; Bildik, G.; Oktem, O. Sphingosine-1-phosphate protects human ovarian follicles from apoptosis in vitro. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 222, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Liu, X.; Yan, Z.; Xie, L. Sphingosine 1-phosphate regulates proliferation, cell cycle and apoptosis of hepatocellular carcinoma cells via syndecan-1. Prog. Biophys. Mol. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Miranda, G.E.; Abrahan, C.E.; Politi, L.E.; Rotstein, N.P. Sphingosine-1-phosphate is a key regulator of proliferation and differentiation in retina photoreceptors. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4416–4428. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Toman, R.E.; Goparaju, S.K.; Maceyka, M.; Nava, V.E.; Sankala, H.; Payne, S.G.; Bektas, M.; Ishii, I.; Chun, J.; et al. Sphingosine kinase type 2 is a putative BH3-only protein that induces apoptosis. J. Biol. Chem. 2003, 278, 40330–40336. [Google Scholar] [CrossRef]
- Wilkerson, J.L.; Stiles, M.A.; Gurley, J.M.; Grambergs, R.C.; Gu, X.; Elliott, M.H.; Proia, R.L.; Mandal, N.A. Sphingosine Kinase-1 Is Essential for Maintaining External/Outer Limiting Membrane and Associated Adherens Junctions in the Aging Retina. Mol. Neurobiol. 2019. [Google Scholar] [CrossRef]
- Fabiani, C.; Zulueta, A.; Bonezzi, F.; Casas, J.; Ghidoni, R.; Signorelli, P.; Caretti, A. 2-Acetyl-5-tetrahydroxybutyl imidazole (THI) protects 661W cells against oxidative stress. Naunyn Schmiedebergs Arch. Pharmacol. 2017, 390, 741–751. [Google Scholar] [CrossRef] [Green Version]
- Stiles, M.; Qi, H.; Sun, E.; Tan, J.; Porter, H.; Allegood, J.; Chalfant, C.E.; Yasumura, D.; Matthes, M.T.; LaVail, M.M.; et al. Sphingolipid profile alters in retinal dystrophic P23H-1 rats and systemic FTY720 can delay retinal degeneration. J. Lipid Res. 2016, 57, 818–831. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, R.; Kurano, M.; Mishima, Y.; Nojiri, T.; Tokuhara, Y.; Kishimoto, T.; Nakamura, K.; Okubo, S.; Hosogaya, S.; Ozaki, Y.; et al. Possible involvement of sphingomyelin in the regulation of the plasma sphingosine 1-phosphate level in human subjects. Clin. Biochem. 2015, 48, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, N.; Okada, T.; Hayashi, S.; Fujita, T.; Jahangeer, S.; Nakamura, S. Sphingosine kinase 2 is a nuclear protein and inhibits DNA synthesis. J. Biol. Chem. 2003, 278, 46832–46839. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tran, J.T.; Eckerd, A.; Huynh, T.P.; Elliott, M.H.; Brush, R.S.; Mandal, N.A. Inhibition of de novo ceramide biosynthesis by FTY720 protects rat retina from light-induced degeneration. J. Lipid Res. 2013, 54, 1616–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.S.; Kim, S.Y.; Kleuser, B.; Schafer-Korting, M.; Kim, K.H.; Park, K.C. Sphingosine-1-phosphate inhibits human keratinocyte proliferation via Akt/protein kinase B inactivation. Cell. Signal. 2004, 16, 89–95. [Google Scholar] [CrossRef]
- Schuppel, M.; Kurschner, U.; Kleuser, U.; Schafer-Korting, M.; Kleuser, B. Sphingosine 1-phosphate restrains insulin-mediated keratinocyte proliferation via inhibition of Akt through the S1P2 receptor subtype. J. Investig. Dermatol. 2008, 128, 1747–1756. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.Q.; Long, L.; Yan, J.Q.; Wei, L.; Pan, M.Q.; Gao, H.M.; Zhou, P.; Liu, M.; Zhu, C.S.; Tang, B.S.; et al. Simvastatin induces neuroprotection in 6-OHDA-lesioned PC12 via the PI3K/AKT/caspase 3 pathway and anti-inflammatory responses. CNS Neurosci. Ther. 2013, 19, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Saigusa, D.; Okudaira, M.; Wang, J.; Kano, K.; Kurano, M.; Uranbileg, B.; Ikeda, H.; Yatomi, Y.; Motohashi, H.; Aoki, J. Simultaneous Quantification of Sphingolipids in Small Quantities of Liver by LC-MS/MS. Mass Spectrom. (Tokyo) 2014, 3, S0046. [Google Scholar] [CrossRef]
- Frej, C.; Andersson, A.; Larsson, B.; Guo, L.J.; Norström, E.; Happonen, K.E.; Dahlbäck, B. Quantification of sphingosine 1-phosphate by validated LC-MS/MS method revealing strong correlation with apolipoprotein M in plasma but not in serum due to platelet activation during blood coagulation. Anal. Bioanal. Chem. 2015, 407, 8533–8542. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligos | Forward (5′–3′) | Reverse (3′–5′) |
|---|---|---|
| SphK1 | ACAGTGGGCACCTTCTTTC | CTTCTGCACCAGTGTAGAGGC |
| SphK2 | ACCACTTATGAGGAGAATCG | CACCACGTGGTCCATACAGC |
| S1P1 | TCATCTGCTGCTTCATCA | CTGCTAATAGGTCCGAGAG |
| S1P2 | CTACAATTACACCAAGGAGAC | CAGCACAAGATGATGATGAA |
| S1P3 | CACCACCATCCTCTTCTT | ATTGACCTTGTATGCTATGC |
| S1P4 | CTCCTGGCTGACATCTTT | TTAATGGCTGAGTTGAACAC |
| S1P5 | TCTCTTGCTATTACTGGATGT | TTGGTGAAGGTGTAGATGA |
| GAPDH | AACTTTGGCATTGTGGAAGG | ACACATTGGGGGTAGGAACA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terao, R.; Honjo, M.; Ueta, T.; Obinata, H.; Izumi, T.; Kurano, M.; Yatomi, Y.; Koso, H.; Watanabe, S.; Aihara, M. Light Stress-Induced Increase of Sphingosine 1-Phosphate in Photoreceptors and Its Relevance to Retinal Degeneration. Int. J. Mol. Sci. 2019, 20, 3670. https://doi.org/10.3390/ijms20153670
Terao R, Honjo M, Ueta T, Obinata H, Izumi T, Kurano M, Yatomi Y, Koso H, Watanabe S, Aihara M. Light Stress-Induced Increase of Sphingosine 1-Phosphate in Photoreceptors and Its Relevance to Retinal Degeneration. International Journal of Molecular Sciences. 2019; 20(15):3670. https://doi.org/10.3390/ijms20153670
Chicago/Turabian StyleTerao, Ryo, Megumi Honjo, Takashi Ueta, Hideru Obinata, Takashi Izumi, Makoto Kurano, Yutaka Yatomi, Hideto Koso, Sumiko Watanabe, and Makoto Aihara. 2019. "Light Stress-Induced Increase of Sphingosine 1-Phosphate in Photoreceptors and Its Relevance to Retinal Degeneration" International Journal of Molecular Sciences 20, no. 15: 3670. https://doi.org/10.3390/ijms20153670