Chemokine CXCL13 as a New Systemic Biomarker for B-Cell Involvement in Acute T Cell-Mediated Kidney Allograft Rejection

 and
and
Abstract
1. Introduction
2. Results
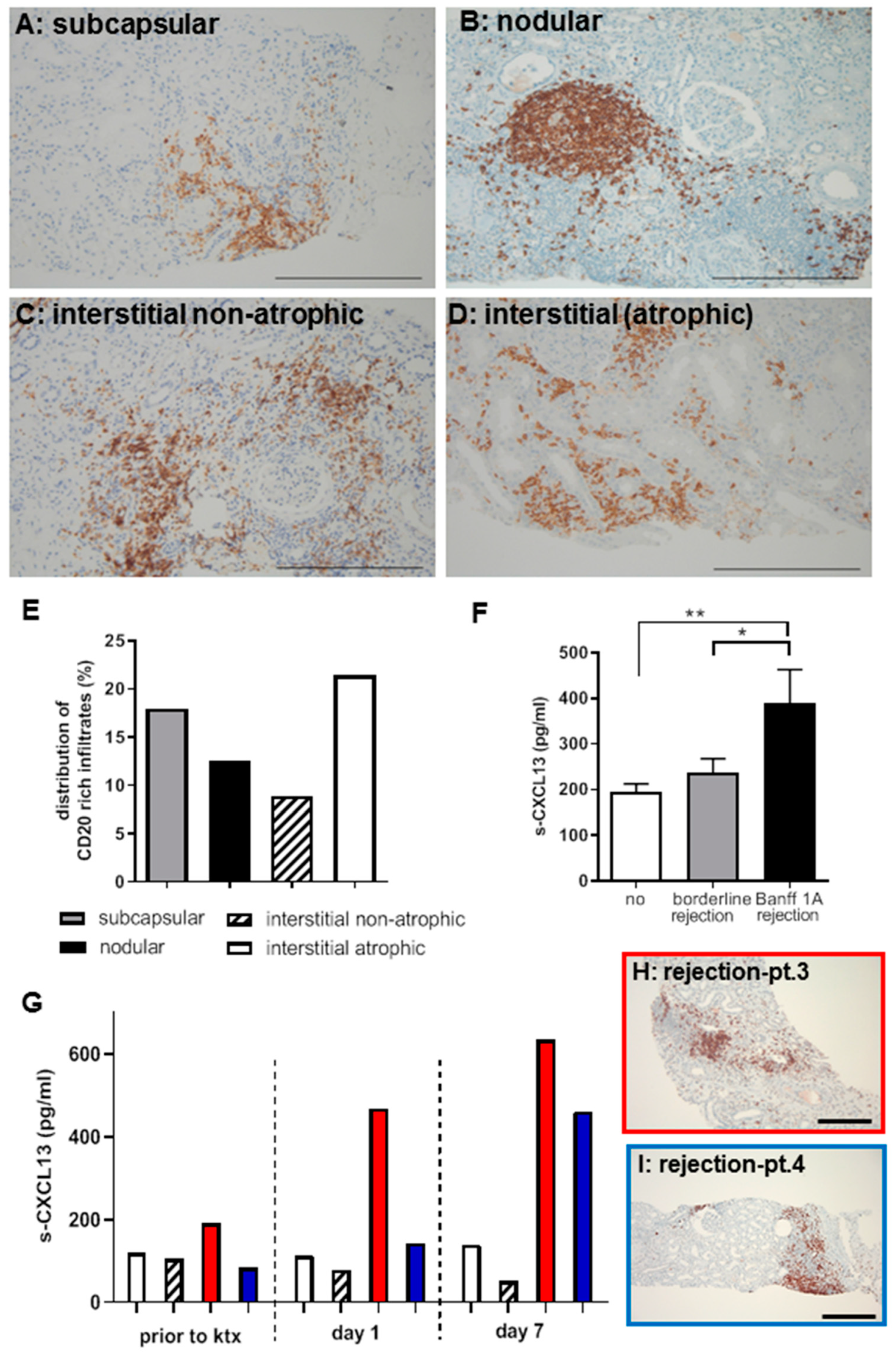
2.1. B-cell Involvement in Kidney Allograft Rejection in Patients
2.2. CXCL13 as a Systemic Biomarker of TCMR Rejection in Patients
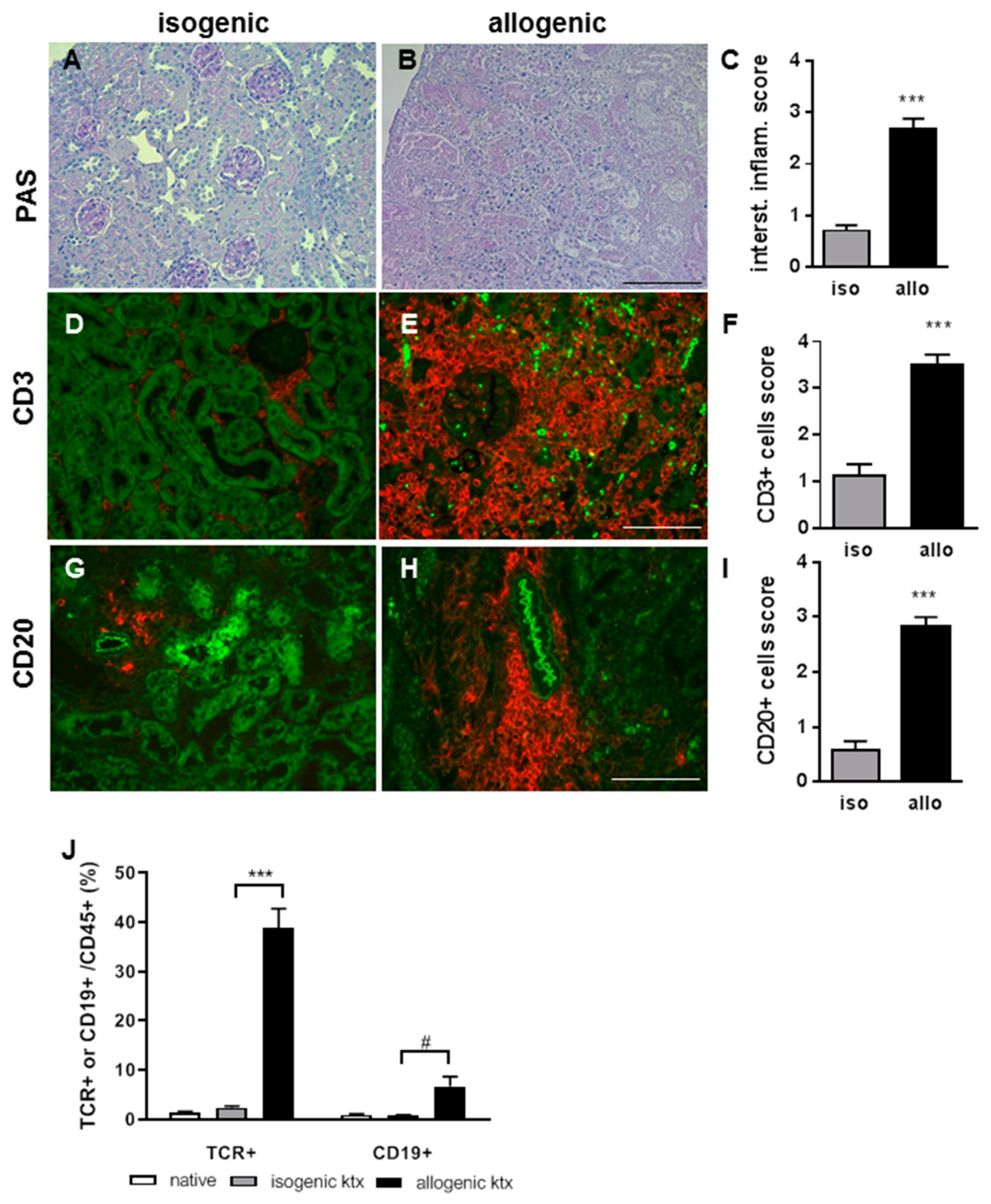
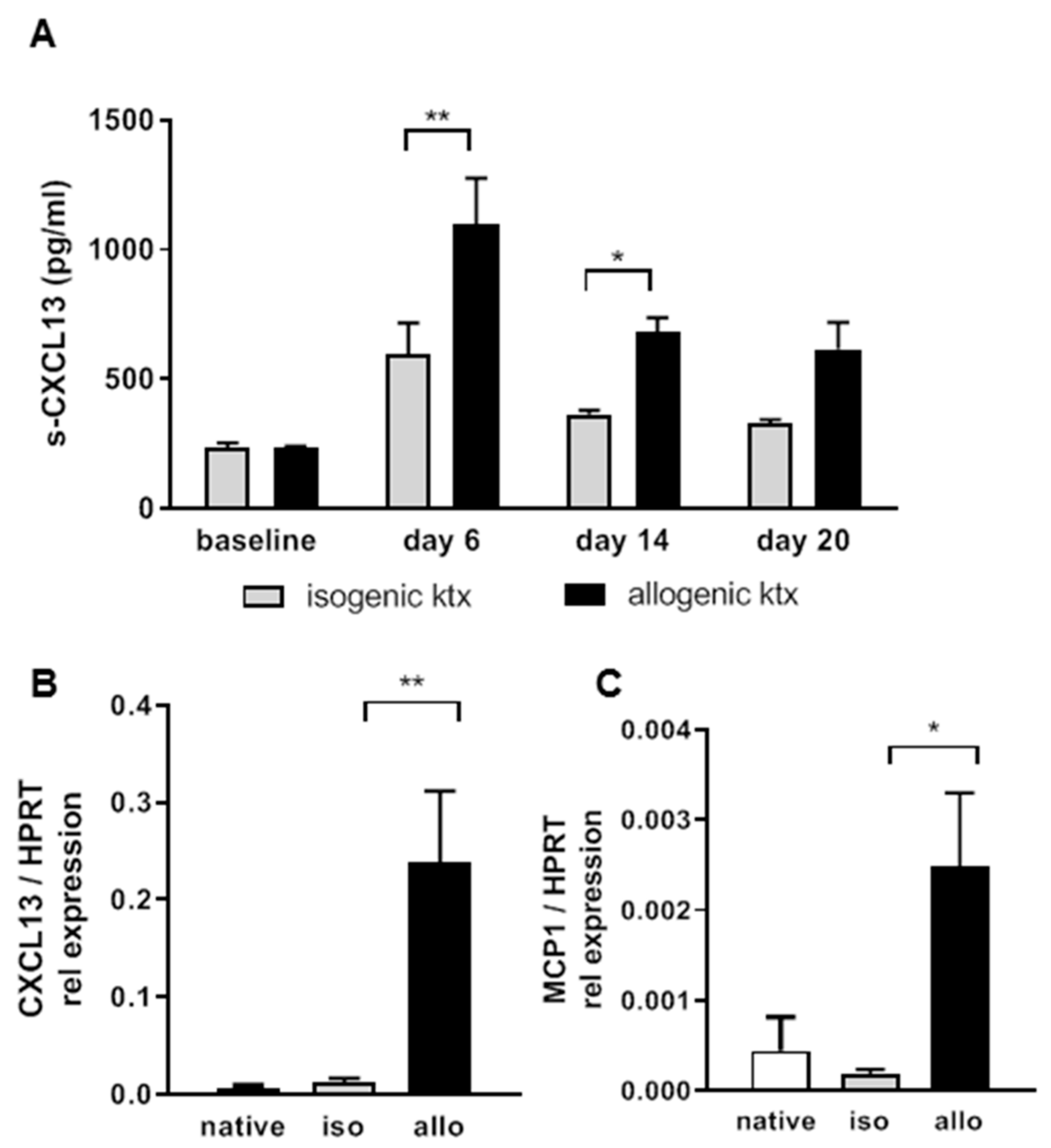
2.3. Allograft Rejection in a Translational Mouse Model for TCMR
3. Discussion
4. Materials and Methods
4.1. Patient Samples
4.2. CXCL13 ELISA
4.3. Renal Morphology and Immunohistochemistry of Patient Allograft Biopsies
4.4. Mice
4.5. Kidney Transplantation
4.6. Renal Morphology and Immunohistochemistry of Mouse Kidney Grafts
4.7. Flow Cytometry
4.8. CXCL13 Expression in Kidney Graft Tissue
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BCA | B-cell attracting chemokine |
| CXCL13 | CXC ligand 13 protein |
| ktx | Kidney transplantation |
| allograft rejection | T cell-mediated rejection |
References
- Cardinal, H.; Dieude, M.; Hebert, M.J. The Emerging Importance of Non-HLA Autoantibodies in Kidney Transplant Complications. J. Am. Soc. Nephrol. 2017, 28, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Lefaucheur, C.; Viglietti, D.; Bouatou, Y.; Philippe, A.; Pievani, D.; Aubert, O.; Duong Van Huyen, J.P.; Taupin, J.L.; Glotz, D.; Legendre, C.; et al. Non-HLA agonistic anti-angiotensin II type 1 receptor antibodies induce a distinctive phenotype of antibody-mediated rejection in kidney transplant recipients. Kidney Int. 2019. (Epub ahead of print). [Google Scholar] [CrossRef] [PubMed]
- Sellares, J.; de Freitas, D.G.; Mengel, M.; Reeve, J.; Einecke, G.; Sis, B.; Hidalgo, L.G.; Famulski, K.; Matas, A.; Halloran, P.F. Understanding the causes of kidney transplant failure: The dominant role of antibody-mediated rejection and nonadherence. Am. J. Transpl. 2012, 12, 388–399. [Google Scholar] [CrossRef]
- Mengel, M.; Gwinner, W.; Schwarz, A.; Bajeski, R.; Franz, I.; Brocker, V.; Becker, T.; Neipp, M.; Klempnauer, J.; Haller, H.; et al. Infiltrates in protocol biopsies from renal allografts. Am. J. Transpl. 2007, 7, 356–365. [Google Scholar] [CrossRef]
- Hasegawa, J.; Honda, K.; Omoto, K.; Wakai, S.; Shirakawa, H.; Okumi, M.; Ishida, H.; Fuchinoue, S.; Hattori, M.; Tanabe, K. Clinical and Pathological Features of Plasma Cell-Rich Acute Rejection After Kidney Transplantation. Transplantation 2018, 102, 853–859. [Google Scholar] [CrossRef]
- Kwun, J.; Manook, M.; Page, E.; Burghuber, C.; Hong, J.; Knechtle, S.J. Crosstalk Between T and B Cells in the Germinal Center After Transplantation. Transplantation 2017, 101, 704–712. [Google Scholar] [CrossRef]
- Ansel, K.M.; Ngo, V.N.; Hyman, P.L.; Luther, S.A.; Forster, R.; Sedgwick, J.D.; Browning, J.L.; Lipp, M.; Cyster, J.G. A chemokine-driven positive feedback loop organizes lymphoid follicles. Nature 2000, 406, 309–314. [Google Scholar] [CrossRef]
- Luther, S.A.; Ansel, K.M.; Cyster, J.G. Overlapping roles of CXCL13, interleukin 7 receptor alpha, and CCR7 ligands in lymph node development. J. Exp. Med. 2003, 197, 1191–1198. [Google Scholar] [CrossRef]
- Gunn, M.D.; Ngo, V.N.; Ansel, K.M.; Ekland, E.H.; Cyster, J.G.; Williams, L.T. A B-cell-homing chemokine made in lymphoid follicles activates Burkitt’s lymphoma receptor-1. Nature 1998, 391, 799–803. [Google Scholar] [CrossRef] [PubMed]
- El-Haibi, C.P.; Singh, R.; Gupta, P.; Sharma, P.K.; Greenleaf, K.N.; Singh, S.; Lillard, J.W., Jr. Antibody Microarray Analysis of Signaling Networks Regulated by Cxcl13 and Cxcr5 in Prostate Cancer. J. Proteom. Bioinform. 2012, 5, 177–184. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, J.; Peng, W.; Weng, C.; Chen, J. Urinary CXC motif chemokine 13 is a noninvasive biomarker of antibodymediated renal allograft rejection. Mol. Med. Rep. 2018, 18, 2399–2406. [Google Scholar] [CrossRef]
- Halloran, P.F.; Famulski, K.; Reeve, J. The molecular phenotypes of rejection in kidney transplant biopsies. Curr. Opin. Organ Transpl. 2015, 20, 359–367. [Google Scholar] [CrossRef]
- Einecke, G.; Reeve, J.; Mengel, M.; Sis, B.; Bunnag, S.; Mueller, T.F.; Halloran, P.F. Expression of B cell and immunoglobulin transcripts is a feature of inflammation in late allografts. Am. J. Transpl. 2008, 8, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Carpio, V.N.; Noronha Ide, L.; Martins, H.L.; Jobim, L.F.; Gil, B.C.; Kulzer, A.S.; Loreto Mda, S.; Goncalves, L.F.; Manfro, R.C.; Veronese, F.V. Expression patterns of B cells in acute kidney transplant rejection. Exp. Clin. Transpl. 2014, 12, 405–414. [Google Scholar]
- Firl, D.J.; Benichou, G.; Kim, J.I.; Yeh, H. A Paradigm Shift on the Question of B Cells in Transplantation? Recent Insights on Regulating the Alloresponse. Front Immunol. 2017, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Hippen, B.E.; DeMattos, A.; Cook, W.J.; Kew, C.E., 2nd; Gaston, R.S. Association of CD20+ infiltrates with poorer clinical outcomes in acute cellular rejection of renal allografts. Am. J. Transpl. 2005, 5, 2248–2252. [Google Scholar] [CrossRef]
- Tsai, E.W.; Rianthavorn, P.; Gjertson, D.W.; Wallace, W.D.; Reed, E.F.; Ettenger, R.B. CD20+ lymphocytes in renal allografts are associated with poor graft survival in pediatric patients. Transplantation 2006, 82, 1769–1773. [Google Scholar] [CrossRef]
- Steinmetz, O.M.; Panzer, U.; Kneissler, U.; Harendza, S.; Lipp, M.; Helmchen, U.; Stahl, R.A. BCA-1/CXCL13 expression is associated with CXCR5-positive B-cell cluster formation in acute renal transplant rejection. Kidney Int. 2005, 67, 1616–1621. [Google Scholar] [CrossRef] [PubMed]
- Loupy, A.; Haas, M.; Solez, K.; Racusen, L.; Glotz, D.; Seron, D.; Nankivell, B.J.; Colvin, R.B.; Afrouzian, M.; Akalin, E.; et al. The Banff 2015 Kidney Meeting Report: Current Challenges in Rejection Classification and Prospects for Adopting Molecular Pathology. Am. J. Transpl. 2017, 17, 28–41. [Google Scholar] [CrossRef]
- Schiffer, L.; Henke-Gendo, C.; Wilsdorf, N.; Hussein, K.; Pape, L.; Schmitt, C.; Haller, H.; Schiffer, M.; Klein, C.; Kreipe, H.; et al. CXCL13 as a novel marker for diagnosis and disease monitoring in pediatric PTLD. Am. J. Transpl. 2012, 12, 1610–1617. [Google Scholar] [CrossRef]
- Worthmann, K.; Gueler, F.; von Vietinghoff, S.; Davalos-Misslitz, A.; Wiehler, F.; Davidson, A.; Witte, T.; Haller, H.; Schiffer, M.; Falk, C.S.; et al. Pathogenetic role of glomerular CXCL13 expression in lupus nephritis. Clin. Exp. Immunol. 2014, 178, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Gueler, F.; Rong, S.; Gwinner, W.; Mengel, M.; Brocker, V.; Schon, S.; Greten, T.F.; Hawlisch, H.; Polakowski, T.; Schnatbaum, K.; et al. Complement 5a receptor inhibition improves renal allograft survival. J. Am. Soc. Nephrol. 2008, 19, 2302–2312. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Nagai, S.; Sato, T.; Akadegawa, K.; Yoneyama, H.; Zhang, Y.Y.; Onai, N.; Matsushima, K. Increased circulating CD11b+CD11c+ dendritic cells (DC) in aged BWF1 mice which can be matured by TNF-alpha into BLC/CXCL13-producing DC. Eur. J. Immunol. 2002, 32, 1881–1887. [Google Scholar] [CrossRef]
- Xu, X.; Han, Y.; Wang, Q.; Cai, M.; Qian, Y.; Wang, X.; Huang, H.; Xu, L.; Xiao, L.; Shi, B. Characterisation of Tertiary Lymphoid Organs in Explanted Rejected Donor Kidneys. Immunol. Invest. 2016, 45, 38–51. [Google Scholar] [CrossRef]
- Rong, S.; Lewis, A.G.; Kunter, U.; Haller, H.; Gueler, F. A knotless technique for kidney transplantation in the mouse. J. Transpl. 2012, 2012, 127215. [Google Scholar] [CrossRef]
- Gueler, F.; Shushakova, N.; Mengel, M.; Hueper, K.; Chen, R.; Liu, X.; Park, J.K.; Haller, H.; Wensvoort, G.; Rong, S. A novel therapy to attenuate acute kidney injury and ischemic allograft damage after allogenic kidney transplantation in mice. PLoS ONE 2015, 10, e0115709. [Google Scholar] [CrossRef]
- Hueper, K.; Gutberlet, M.; Brasen, J.H.; Jang, M.S.; Thorenz, A.; Chen, R.; Hertel, B.; Barrmeyer, A.; Schmidbauer, M.; Meier, M.; et al. Multiparametric Functional MRI: Non-Invasive Imaging of Inflammation and Edema Formation after Kidney Transplantation in Mice. PLoS ONE 2016, 11, e0162705. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.; Sis, B.; Racusen, L.C.; Solez, K.; Glotz, D.; Colvin, R.B.; Castro, M.C.; David, D.S.; David-Neto, E.; Bagnasco, S.M.; et al. Banff 2013 meeting report: Inclusion of c4d-negative antibody-mediated rejection and antibody-associated arterial lesions. Am. J. Transpl. 2014, 14, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org (accessed on 1 May 2018).
- Touloumis, A. R Package multgee: A Generalized Estimating Equations Solver for Multinomial Responses. J. Stat. Softw. 2015, 64, 1–14. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type of Rejection | Banff 1A or Higher | Borderline | No Rejection |
|---|---|---|---|
| Number of patients | 9 | 19 | 45 |
| Number of serum samples | 10 | 24 | 65 |
| Time between transplantation and biopsy (days; ±SD) | 108.9 (±56.2) | 110.4 (±59.0) | 117.0 (±53.6) |
| Age at transplantation (years; ±SD) | 54.1 (±19.6) | 48.3 (±10.2) | 54.5 (±12.6) |
| HLA-Mismatch (mean ± SD) | 1.5 (±2.3) | 1.9 (±1.6) | 2.1 (±1.7) |
| Data available for the following number of patients | 8 (9) | 15 (19) | 41 (45) |
| Creatinine level at time of biopsy (μmol/l; ±SD) | 160.1 (±72.9) | 194.0 (±91.8) | 154.1 (±72.4) |
| Creatinine level after 1 year (μmol/l; ±SD) | 156.4 (±41.1) | 188.1 (±73.7) | 142.2 (±63.9) |
| Creatinine level after 5 year (μmol/l; ±SD) | 146.2 (±41.9) | 196.9 (±91.5) | 162.9 (±99.9) |
| Immunosuppression at time of biopsy | |||
| Number of immunosuppressants (±SD) | 2.2 (±0.6) | 2.2 (±0.4) | 2.6 (±0.6) |
| Data available for the following number of samples | 10 (10) | 23 (24) | 62(65) |
| Prednisolon dose (mg; ±SD) | 10.5 (±5.7) | 12.2 (±5.3) | 9.6 (±5.4) |
| Cyclosporine A | 50% | 91.7% | 88.7% |
| Mycophenolat mofetil | 20% | 25% | 59.7% |
| Sirolimus | 0% | 8.3% | 16.1% |
| Tacrolimus | 30% | 4.2% | 3.2% |
| Belatacept | 20% | 0% | 4.8% |
| Number of rejections (mean ± SD) | 1.2 (±0.4) | 0.2 (±0.4) | 0 (±0) |
| DSA-Status | n.d. | n.d. | n.d. |
| Patient Characteristics | Patient 1 | Patient 2 | Patient 3 | Patient 4 |
|---|---|---|---|---|
| Recipient age (years) | 22 | 48 | 51 | 50 |
| Hemodialysis (years) | None | 6 | 12 | 12 |
| Type of ktx | living donation | postmortal, AM-Program | postmortal | postmortal |
| Plasmapheresis | no | 2× day 0 + 1 | day 15, 16, 17 | no |
| Delayed graft function | no | no | no | yes |
| Allograft biopsy (day) | n/a | n/a | 10 | 14 |
| In hospital stay (days) | 8 | 8 | 21 | 14 |
| Steroid boli for rejection treatment | n.a. | n.a. | 3× 500 mg prednisolone | 3× 500 mg prednisolone |
| Creatinine at 4 weeks after ktx (μmol/L) | 145 | 178 | 162 | 169 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiffer, L.; Wiehler, F.; Bräsen, J.H.; Gwinner, W.; Greite, R.; Kreimann, K.; Thorenz, A.; Derlin, K.; Teng, B.; Rong, S.; et al. Chemokine CXCL13 as a New Systemic Biomarker for B-Cell Involvement in Acute T Cell-Mediated Kidney Allograft Rejection. Int. J. Mol. Sci. 2019, 20, 2552. https://doi.org/10.3390/ijms20102552
Schiffer L, Wiehler F, Bräsen JH, Gwinner W, Greite R, Kreimann K, Thorenz A, Derlin K, Teng B, Rong S, et al. Chemokine CXCL13 as a New Systemic Biomarker for B-Cell Involvement in Acute T Cell-Mediated Kidney Allograft Rejection. International Journal of Molecular Sciences. 2019; 20(10):2552. https://doi.org/10.3390/ijms20102552
Chicago/Turabian StyleSchiffer, Lena, Flavia Wiehler, Jan Hinrich Bräsen, Wilfried Gwinner, Robert Greite, Kirill Kreimann, Anja Thorenz, Katja Derlin, Beina Teng, Song Rong, and et al. 2019. "Chemokine CXCL13 as a New Systemic Biomarker for B-Cell Involvement in Acute T Cell-Mediated Kidney Allograft Rejection" International Journal of Molecular Sciences 20, no. 10: 2552. https://doi.org/10.3390/ijms20102552
APA StyleSchiffer, L., Wiehler, F., Bräsen, J. H., Gwinner, W., Greite, R., Kreimann, K., Thorenz, A., Derlin, K., Teng, B., Rong, S., von Vietinghoff, S., Haller, H., Mengel, M., Pape, L., Lerch, C., Schiffer, M., & Gueler, F. (2019). Chemokine CXCL13 as a New Systemic Biomarker for B-Cell Involvement in Acute T Cell-Mediated Kidney Allograft Rejection. International Journal of Molecular Sciences, 20(10), 2552. https://doi.org/10.3390/ijms20102552