Zoledronate Enhances Osteocyte-Mediated Osteoclast Differentiation by IL-6/RANKL Axis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Zoledronate, a Nitrogen-Containing Bisphosphonate, Significantly Increased RANKL and Sclerostin Expression from Osteocyte-Like MLO-Y4 Cells
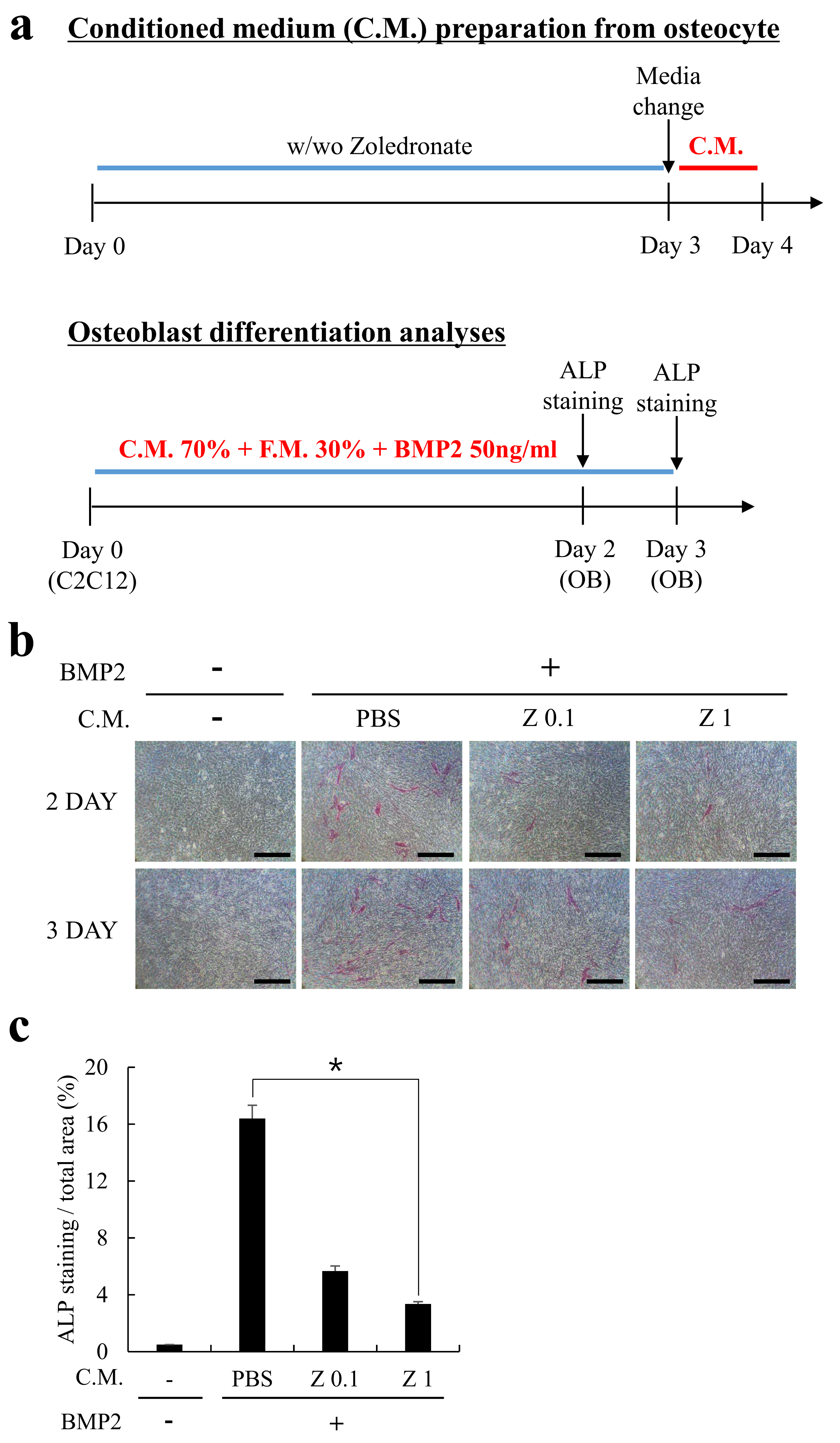
2.2. Conditioned Medium (C.M.) Obtained from Zoledronate-Treated MLO-Y4 Cells Reduced by Osteoblast Differentiation
2.3. Zoledronate Treatment Increased the Expression of Osteoclastogenesis Supporting Factor from MLO-Y4 Cells
2.4. The Autocrine Expression of IL-6 Upon Zoledronate Treatment Resulted in RANKL Expression in MLO-Y4 Cells through JAK2/STAT3 Pathway
2.5. AG490 and IL-6 Neutralizing Antibody Inhibited the Expression of RANKL and Reduced the Differentiation of Osteoclasts co-Cultured with MLO-Y4 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Cultures
4.3. Cytotoxicity Assay
4.4. Conditioned Medium (C.M.) Preparation from MLO-Y4 Cells
4.5. Co-Culture of MLO-Y4 Cells and BMMs
4.6. Quantitative Real-Time Polymerase-Chain Reaction (PCR) Analyses
4.7. Western Blotting
4.8. Statistics
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| RANKL | Receptor Activator of Nuclear Factor kB Ligand |
| BRONJ | Bisphosphonate-Related Osteonecrosis of the Jaw |
| BMM | Bone Marrow-derived Macrophage |
| M-CSF | Macrophage-Colony Stimulating Factor |
References
- Fleisch, H. Diphosphonates: History and mechanisms of action. Metab. Bone Dis. Relat. Res. 1981, 3, 279–287. [Google Scholar] [CrossRef]
- Endo, Y.; Kumamoto, H.; Nakamura, M.; Sugawara, S.; Takano-Yamamoto, T.; Sasaki, K.; Takahashi, T. Underlying Mechanisms and Therapeutic Strategies for Bisphosphonate-Related Osteonecrosis of the Jaw (BRONJ). Biol. Pharm. Bull. 2017, 40, 739–750. [Google Scholar] [CrossRef]
- Rodan, G.A.; Martin, T.J. Therapeutic approaches to bone diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef]
- Rogers, M.J. New insights into the molecular mechanisms of action of bisphosphonates. Curr. Pharm. Des. 2003, 9, 2643–2658. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.G. Bisphosphonates: The first 40 years. Bone 2011, 49, 2–19. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, D.B. Mechanism of action, pharmacokinetic and pharmacodynamic profile, and clinical applications of nitrogen-containing bisphosphonates. J. Dent. Res. 2007, 86, 1022–1033. [Google Scholar] [CrossRef]
- Reszka, A.A.; Rodan, G.A. Nitrogen-containing bisphosphonate mechanism of action. Mini Rev. Med. Chem. 2004, 4, 711–719. [Google Scholar]
- Frith, J.C.; Mönkkönen, J.; Auriola, S.; Mönkkönen, H.; Rogers, M.J. The molecular mechanism of action of the antiresorptive and antiinflammatory drug clodronate: Evidence for the formation in vivo of a metabolite that inhibits bone resorption and causes osteoclast and macrophage apoptosis. Arthritis Rheum. 2001, 44, 2201–2210. [Google Scholar] [CrossRef]
- Lehenkari, P.P.; Kellinsalmi, M.; Näpänkangas, J.P.; Ylitalo, K.V.; Mönkkönen, J.; Rogers, M.J.; Azhayev, A.; Väänänen, H.K.; Hassinen, I.E. Further insight into mechanism of action of clodronate: Inhibition of mitochondrial ADP/ATP translocase by a nonhydrolyzable, adenine-containing metabolite. Mol. Pharmacol. 2002, 61, 1255–1262. [Google Scholar] [CrossRef]
- Marx, R.E. Pamidronate (Aredia) and zoledronate (Zometa) induced avascular necrosis of the jaws: A growing epidemic. J. Oral Maxillofac. Surg. 2003, 61, 1115–1117. [Google Scholar] [CrossRef]
- Sarasquete, M.E.; González, M.; San Miguel, J.F.; García-Sanz, R. Bisphosphonate-related osteonecrosis: Genetic and acquired risk factors. Oral Dis. 2009, 15, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Holtmann, H.; Lommen, J.; Kübler, N.R.; Sproll, C.; Rana, M.; Karschuck, P.; Depprich, R. Pathogenesis of medication-related osteonecrosis of the jaw: A comparative study of in vivo and in vitro trials. J. Int. Med. Res. 2018, 46, 4277–4296. [Google Scholar] [CrossRef] [PubMed]
- Aljohani, S.; Fliefel, R.; Ihbe, J.; Kühnisch, J.; Ehrenfeld, M.; Otto, S. What is the effect of anti-resorptive drugs (ARDs) on the development of medication-related osteonecrosis of the jaw (MRONJ) in osteoporosis patients: A systematic review. J. Craniomaxillofac. Surg. 2017, 45, 1493–1502. [Google Scholar] [CrossRef]
- Basso, F.G.; Silveira Turrioni, A.P.; Hebling, J.; de Souza Costa, C.A. Zoledronic acid inhibits human osteoblast activities. Gerontology 2013, 59, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.H.; Lee, R.S.; Williams, D.; Bae, S.; Woo, J.; Lieberman, M.; Oh, J.E.; Dong, Q.; Shin, K.H.; Kang, M.K.; et al. Bisphosphonates induce senescence in normal human oral keratinocytes. J. Dent. Res. 2011, 90, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Prideaux, M.; Findlay, D.M.; Atkins, G.J. Osteocytes: The master cells in bone remodelling. Curr. Opin. Pharmacol. 2016, 28, 24–30. [Google Scholar] [CrossRef]
- Bellido, T. Osteocyte-driven bone remodeling. Calcif. Tissue Int. 2014, 94, 25–34. [Google Scholar] [CrossRef]
- Ye, T.; Cao, P.; Qi, J.; Zhou, Q.; Rao, DS.; Qiu, S. Protective effect of low-dose risedronate against osteocyte apoptosis and bone loss in ovariectomized rats. PLoS ONE 2017, 12, e0186012. [Google Scholar] [CrossRef]
- Pietrzyk, B.; Smertka, M.; Chudek, J. Sclerostin: Intracellular mechanisms of action and its role in the pathogenesis of skeletal and vascular disorders. Adv. Clin. Exp. Med. 2017, 26, 1283–1291. [Google Scholar] [CrossRef]
- Asagiri, M.; Takayanagi, H. The molecular understanding of osteoclast differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef]
- Charles, J.F.; Aliprantis, A.O. Osteoclasts: More than ‘bone eaters’. Trends Mol. Med. 2014, 20, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C. Role of cytokines in bone resorption. Bone 1995, 17, 63S–67S. [Google Scholar] [CrossRef]
- Wu, Q.; Zhou, X.; Huang, D.; Ji, Y.; Kang, F. IL-6 Enhances Osteocyte-Mediated Osteoclastogenesis by Promoting JAK2 and RANKL Activity In Vitro. Cell. Physiol. Biochem. 2017, 41, 1360–1369. [Google Scholar] [CrossRef]
- O’Brien, C.A.; Nakashima, T.; Takayanagi, H. Osteocyte control of osteoclastogenesis. Bone 2013, 54, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Maruotti, N.; Corrado, A.; Neve, A.; Cantatore, F.P. Bisphosphonates: Effects on osteoblast. Eur. J. Clin. Pharmacol. 2012, 68, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Bellido, T.; Plotkin, L.I. Novel actions of bisphosphonates in bone: Preservation of osteoblast and osteocyte viability. Bone 2011, 49, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; To, L.B.; Farrugia, A.N.; Findlay, D.M.; Green, J.; Gronthos, S.; Evdokiou, A.; Lynch, K.; Atkins, G.J.; Zannettino, A.C. The nitrogen-containing bisphosphonate, zoledronic acid, increases mineralisation of human bone-derived cells in vitro. Bone 2004, 34, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Kim, J.H.; Abbas, A.A.; Yoon, T.R. Alendronate enhances osteogenic differentiation of bone marrow stromal cells: A preliminary study. Clin. Orthop. Relat. Res. 2009, 467, 3121–3128. [Google Scholar] [CrossRef]
- Wang, C.Z.; Chen, S.M.; Chen, C.H.; Wang, C.K.; Wang, G.J.; Chang, J.K.; Ho, M.L. The effect of the local delivery of alendronate on human adipose-derived stem cell-based bone regeneration. Biomaterials 2010, 31, 8674–8683. [Google Scholar] [CrossRef]
- Kellinsalmi, M.; Mönkkönen, H.; Mönkkönen, J.; Leskelä, H.V.; Parikka, V.; Hämäläinen, M.; Lehenkari, P. In vitro comparison of clodronate, pamidronate and zoledronic acid effects on rat osteoclasts and human stem cell-derived osteoblasts. Basic Clin. Pharmacol. Toxicol. 2005, 97, 382–391. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Y.K.; Harris, S.; Ahuja, S.S.; Bonewald, L.F. MLO-Y4 osteocyte-like cells support osteoclast formation and activation. J. Bone Miner. Res. 2002, 17, 2068–2079. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Onal, M.; Jilka, R.L.; Weinstein, R.S.; Manolagas, S.C.; O’Brien, C.A. Matrix-embedded cells control osteoclast formation. Nat. Med. 2011, 11, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.A.; Gubrij, I.; Lin, S.C.; Saylors, R.L.; Manolagas, S.C. STAT3 activation in stromal/osteoblastic cells is required for induction of the receptor activator of NF-kappaB ligand and stimulation of osteoclastogenesis by gp130-utilizing cytokines or interleukin-1 but not 1,25-dihydroxyvitamin D3 or parathyroid hormone. J. Biol. Chem. 1999, 274, 19301–19308. [Google Scholar] [PubMed]
- Baron, R.; Rawadi, G.; Roman-Roman, S. Wnt signaling: A key regulator of bone mass. Curr. Top. Dev. Biol. 2006, 76, 103–127. [Google Scholar] [PubMed]
- Li, X.; Ominsky, M.S.; Niu, Q.T.; Sun, N.; Daugherty, B.; D’Agostin, D.; Kurahara, C.; Gao, Y.; Cao, J.; Gong, J.; et al. Targeted deletion of the sclerostin gene in mice results in increased bone formation and bone strength. J. Bone Miner. Res. 2008, 23, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Wijenayaka, A.R.; Kogawa, M.; Lim, H.P.; Bonewald, L.F.; Findlay, D.M.; Atkins, G.J. Sclerostin stimulates osteocyte support of osteoclast activity by a RANKL-dependent pathway. PLoS ONE 2011, 6, e25900. [Google Scholar] [CrossRef]
- Kim, H.J.; Prasad, V.; Hyung, S.W.; Lee, Z.H.; Lee, S.W.; Bhargava, A.; Pearce, D.; Lee, Y.; Kim, H.H. Plasma membrane calcium ATPase regulates bone mass by fine-tuning osteoclast differentiation and survival. J. Cell Biol. 2012, 199, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Hart, A. Mann-Whitney test is not just a test of medians: Differences in spread can be important. BMJ 2001, 323, 391–393. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.J.; Kim, H.J.; Choi, Y.; Bae, M.-K.; Hwang, D.S.; Shin, S.-H.; Lee, J.-Y. Zoledronate Enhances Osteocyte-Mediated Osteoclast Differentiation by IL-6/RANKL Axis. Int. J. Mol. Sci. 2019, 20, 1467. https://doi.org/10.3390/ijms20061467
Kim HJ, Kim HJ, Choi Y, Bae M-K, Hwang DS, Shin S-H, Lee J-Y. Zoledronate Enhances Osteocyte-Mediated Osteoclast Differentiation by IL-6/RANKL Axis. International Journal of Molecular Sciences. 2019; 20(6):1467. https://doi.org/10.3390/ijms20061467
Chicago/Turabian StyleKim, Hyung Joon, Ha Jin Kim, YunJeong Choi, Moon-Kyoung Bae, Dae Seok Hwang, Sang-Hun Shin, and Jae-Yeol Lee. 2019. "Zoledronate Enhances Osteocyte-Mediated Osteoclast Differentiation by IL-6/RANKL Axis" International Journal of Molecular Sciences 20, no. 6: 1467. https://doi.org/10.3390/ijms20061467