Sensory Neuropeptides and their Receptors Participate in Mechano-Regulation of Murine Macrophages

 , , and
, , and
Abstract
:1. Introduction
2. Results
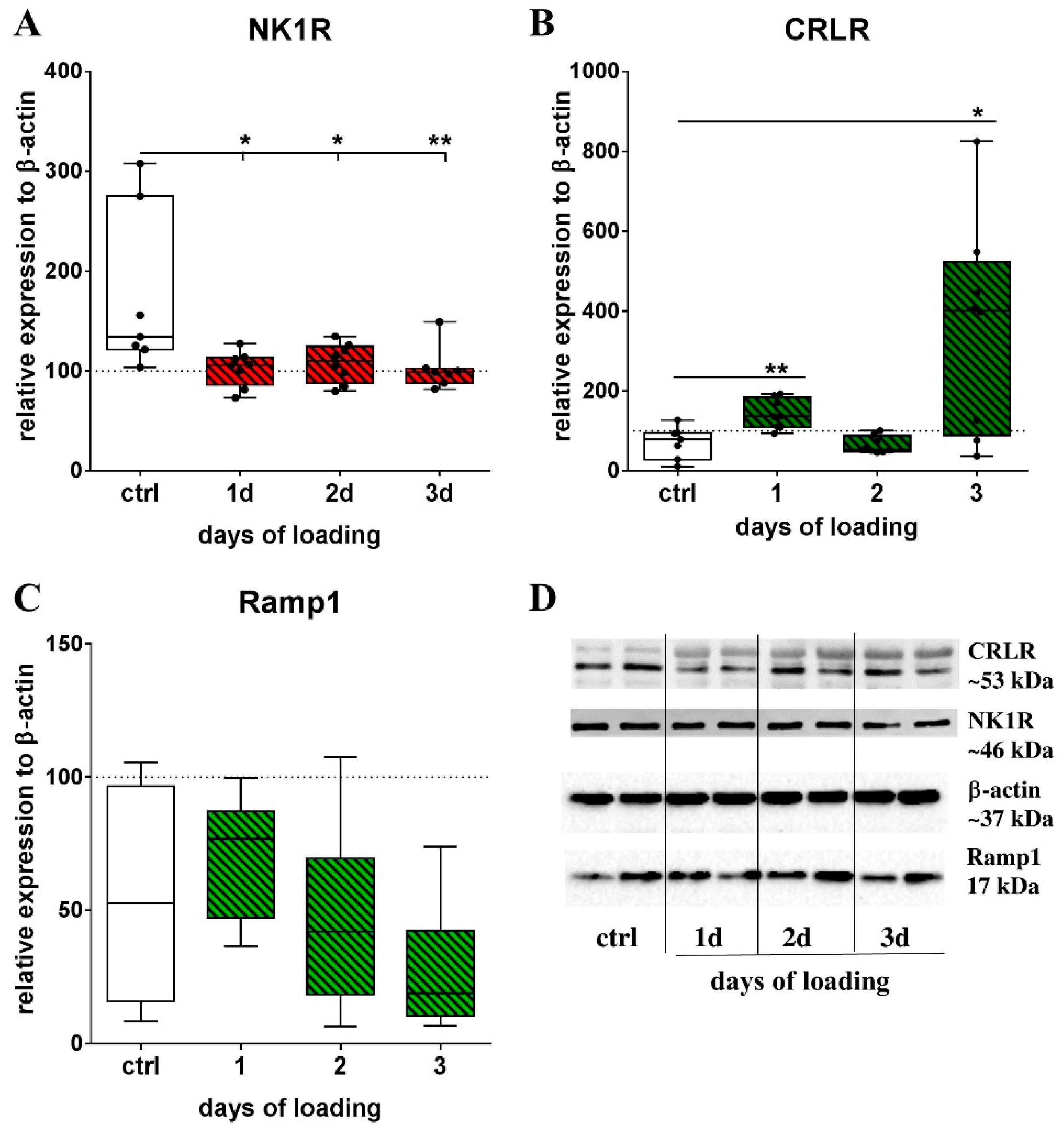
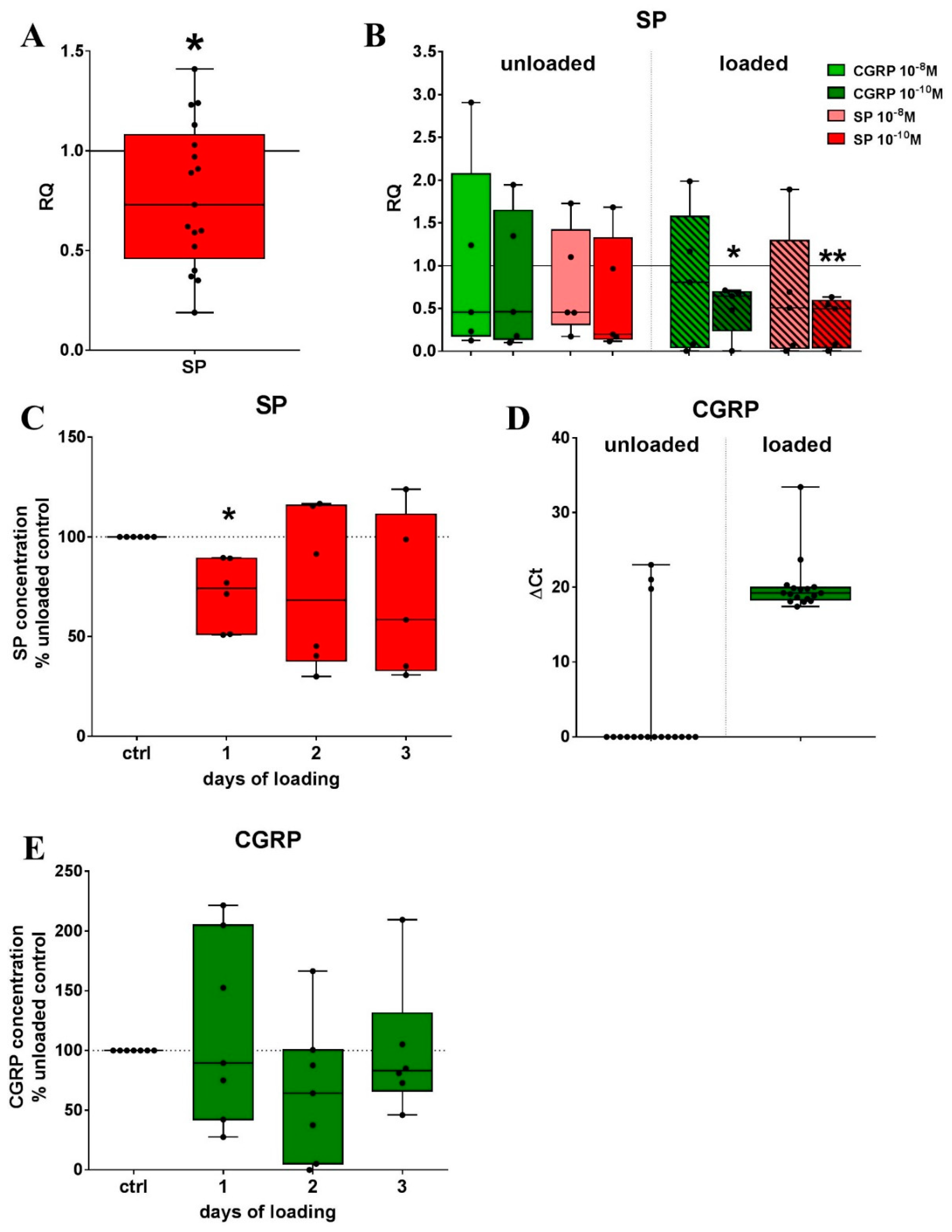
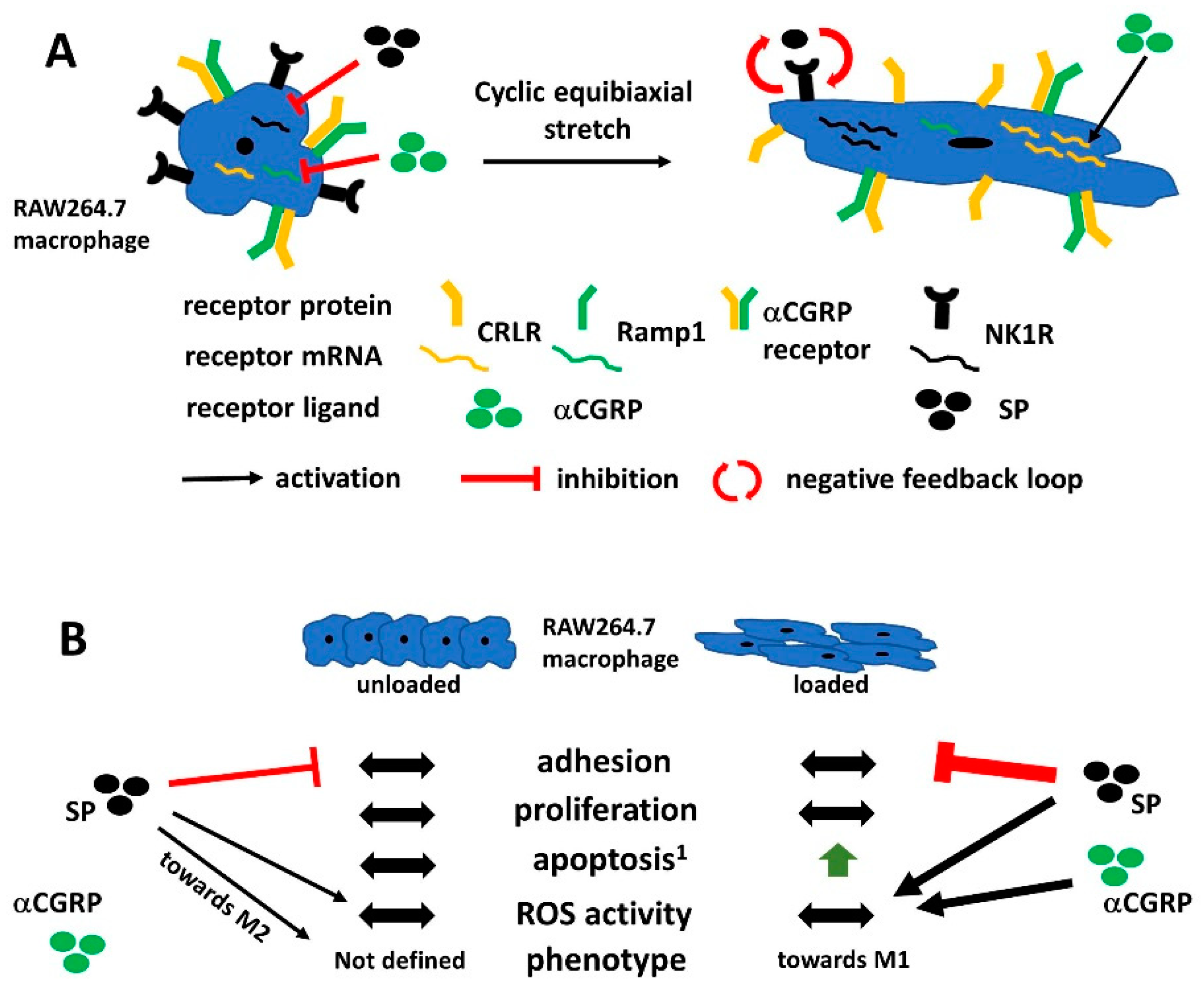
2.1. Mechanical Stretch and Neuropeptide Stimulation Affect Expression of Sensory Neuropeptide Receptor and Endogenous Neuropeptide Expression in RAW264.7 Cells
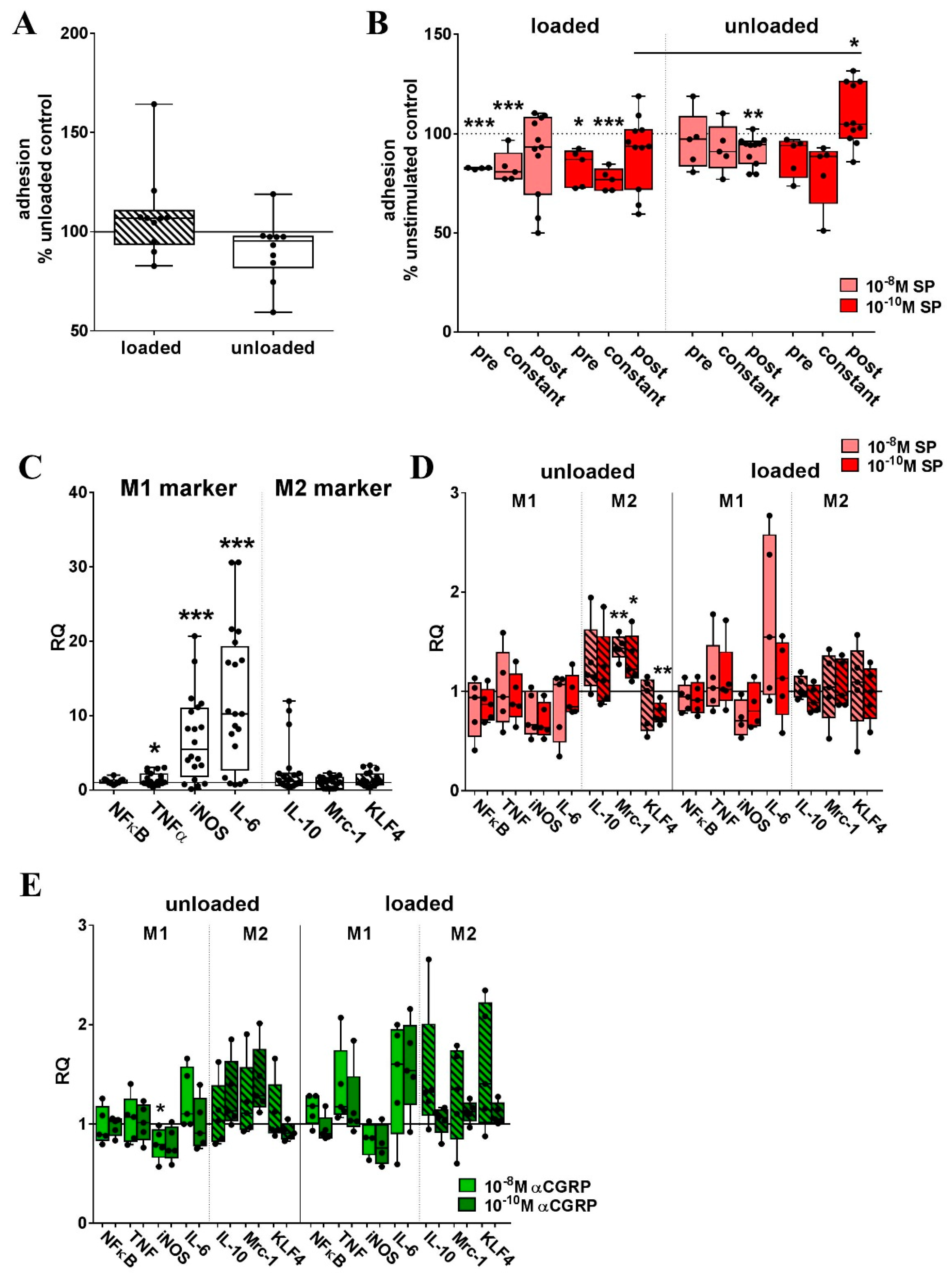
2.2. Metabolic Activity of RAW Cells was Affected by Loading and Sensory Neuropeptide Stimulation
2.3. Mechanical Stretch-Induced M1 Phenotypic Marker Genes in RAW264.7 Cells
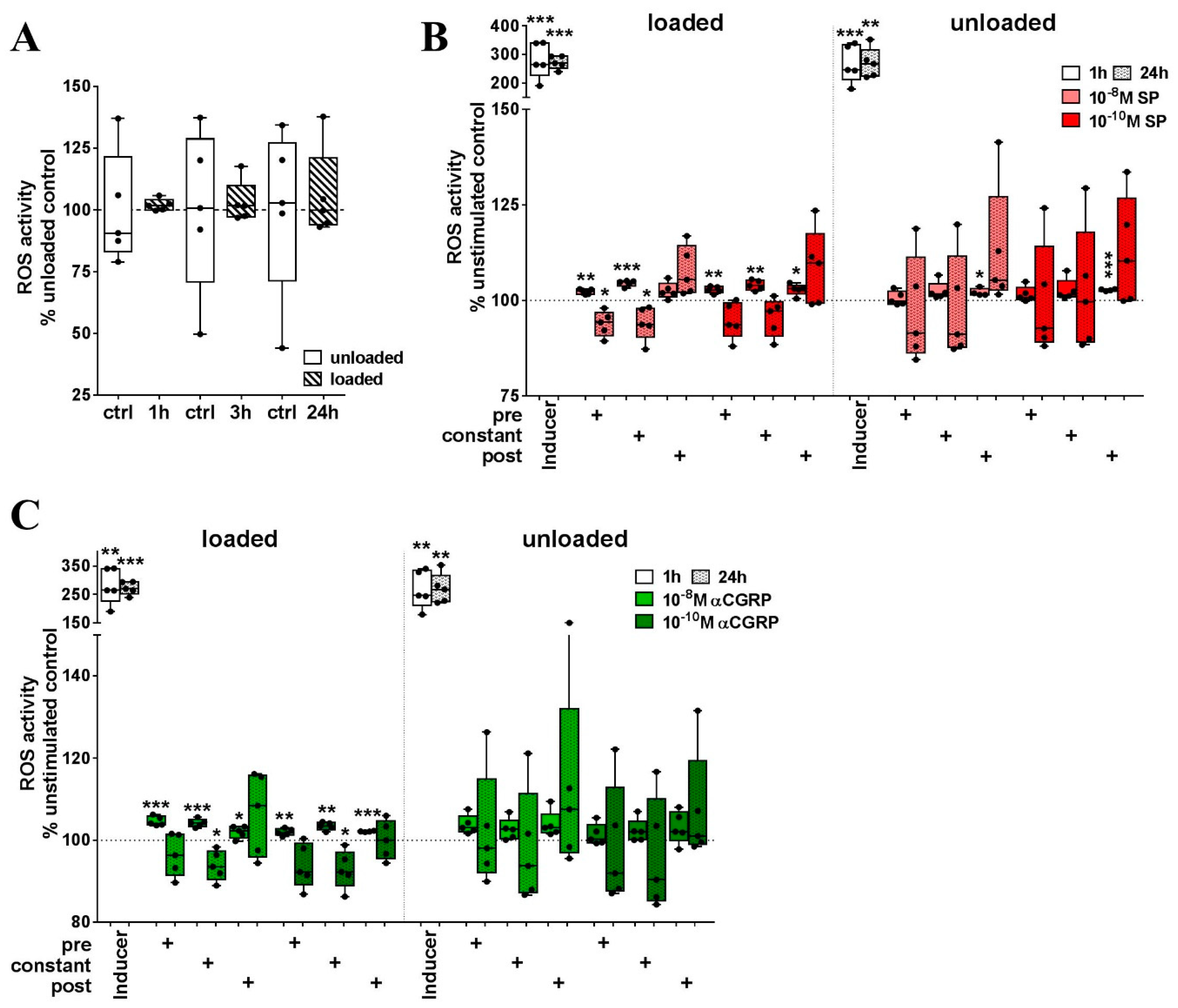
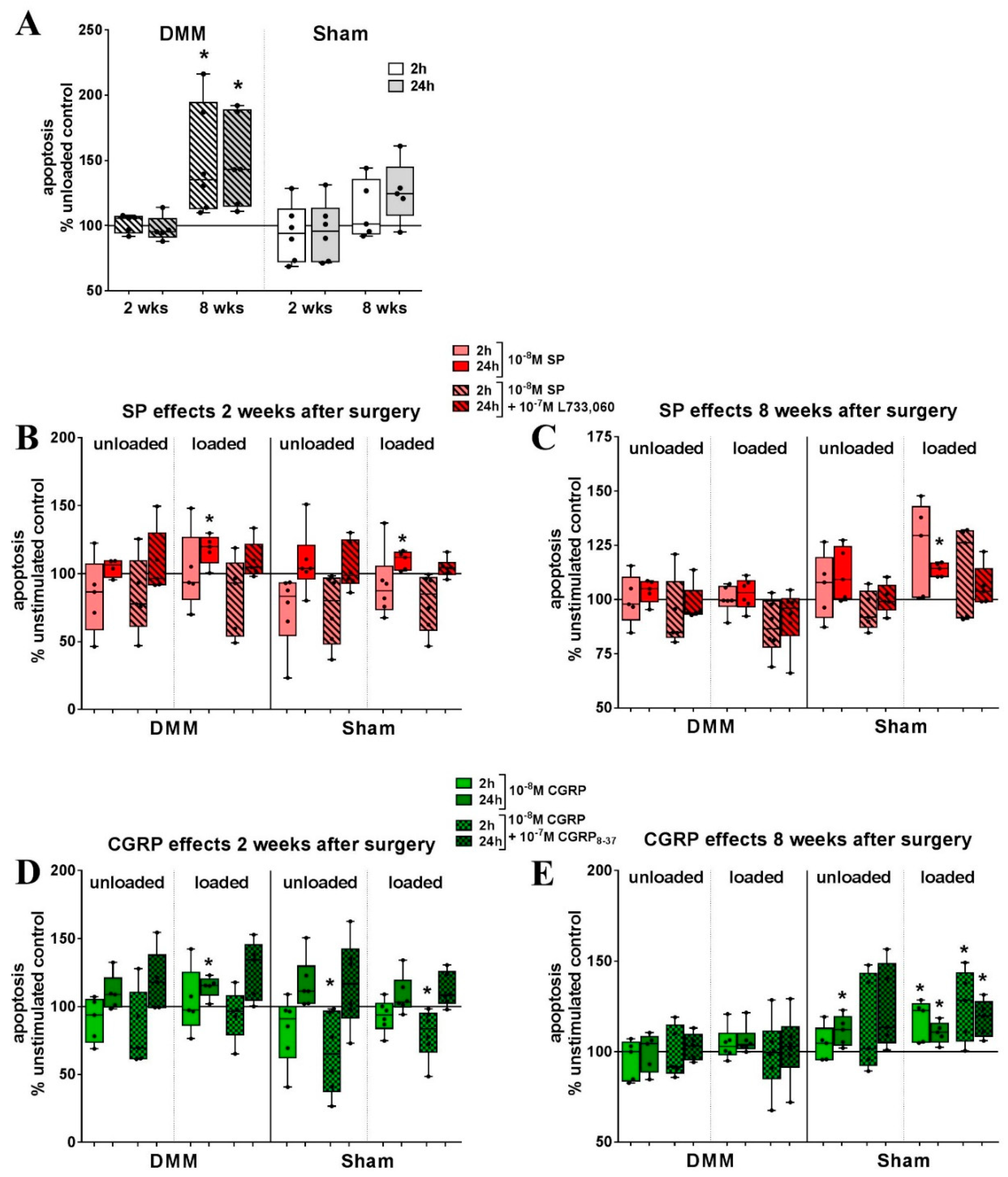
2.4. Loading in the Presence of Neuropeptides but not Loading Alone Evoked a Strong Increase in ROS Activity in RAW264.7 Cells
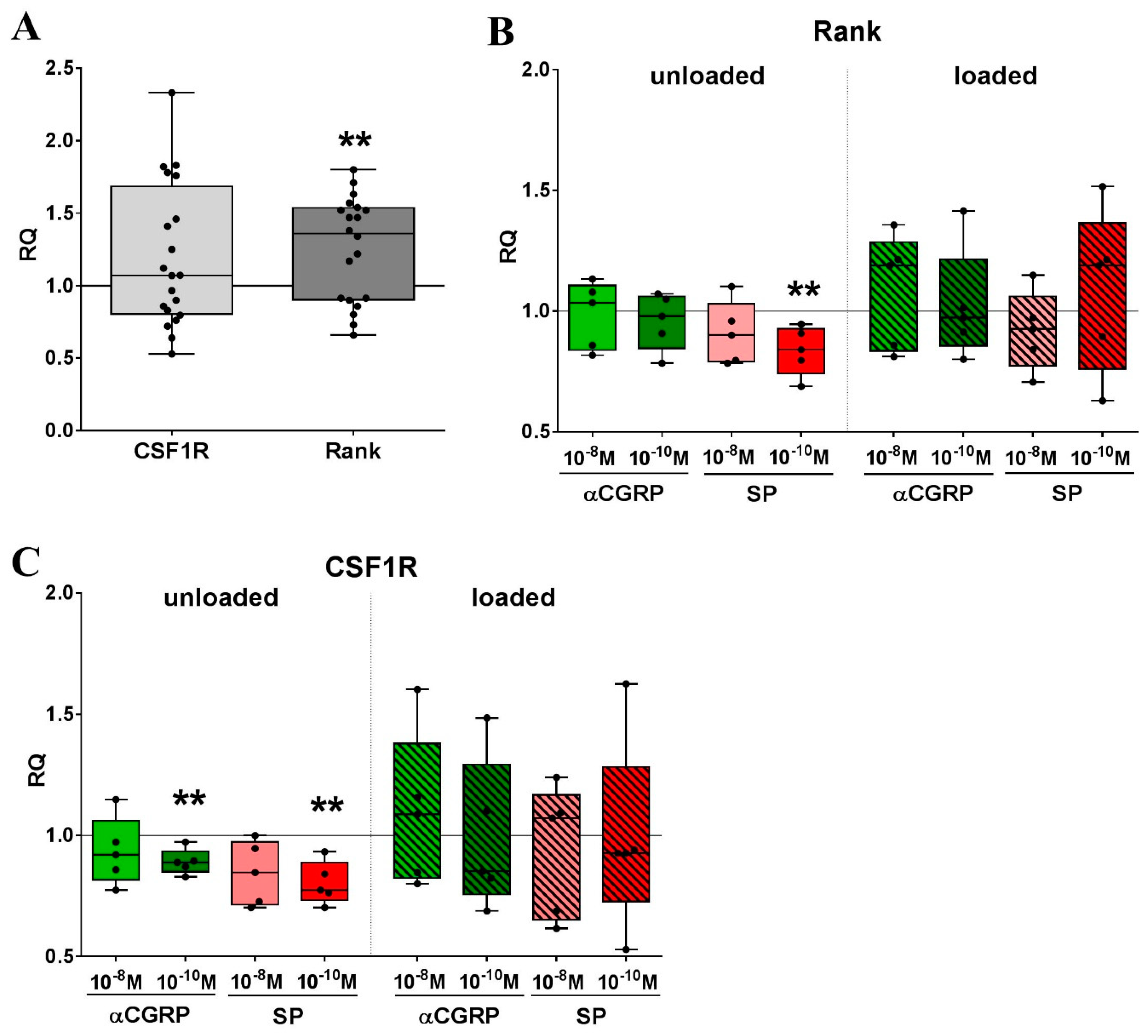
2.5. Mechanical Strain and Neuropeptide Stimulation are Regulators of Osteoclast-related Gene Expression
2.6. Bone Marrow-derived Macrophages from OA Mice Show Differential Reaction to Loading
3. Discussion
4. Materials and Methods
4.1. RAW 264.7 Cells and Cell Culture
4.2. Mechanical Loading
4.3. Animals
4.4. Destabilization of the Medial Meniscus (DMM)
4.5. Isolation of Bone Marrow-derived Macrophages
4.6. RNA Isolation and cDNA Synthesis
4.7. Quantitative Real-time PCR
4.8. Stimulation of RAW264.7 and BMM with SP and αCGRP and Receptor Antagonists
4.9. Caspase 3/7 Apoptosis Assay
4.10. BrdU Incorporation Proliferation Assay
4.11. Crystal Violet Adhesion Assay
4.12. ROS Activity Assay
4.13. Protein Expression of Sensory Neuropeptide Receptors in RAW264.7 Cells
4.14. Production of Endogenous SP and αCGRP
4.15. Osteoclastogenesis of BMM during Mechanical Loading
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SP | Substance P |
| αCGRP | alpha-calcitonin gene-related peptide |
| NK1R | Neurokinin receptor 1 |
| CRLR | Calcitonin receptor-like receptor |
| Ramp1 | Receptor activity-modifying protein 1 |
| BMM | Bone marrow-derived macrophage |
| OA | Osteoarthritis |
| DMM | Destabilized medial meniscus |
| NGF | Nerve growth factor |
| MHCII | Major histocompatibility complex class II |
| IL | Interleukin |
| LPS | Lipopolysaccharide |
| TNF | Tumor necrosis factor |
| TRPV1 | Transient receptor potential cation channel subfamily V member 1 |
| KLF4 | Krüppel-like factor 4 |
| Mrc-1 | Mannose receptor 1 |
| iNOS | Inducible NO synthase |
| CSF1R | Colony stimulating factor 1 receptor |
| NFκB | Nuclear factor “kappa light-chain enhancer” of activated B cells |
| PCR | Polymerase chain reaction |
References
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- McWhorter, F.Y.; Davis, C.T.; Liu, W.F. Physical and mechanical regulation of macrophage phenotype and function. Cell. Mol. Life Sci. 2015, 72, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Post, M. Invited review: Mechanochemical signal transduction in the fetal lung. J. Appl. Physiol. 2000, 89, 2078–2084. [Google Scholar] [CrossRef] [PubMed]
- Klein-Nulend, J.; Bacabac, R.G.; Mullender, M.G. Mechanobiology of bone tissue. Pathol. Biol. 2005, 53, 576–580. [Google Scholar] [CrossRef]
- Chatterjee, S.; Fujiwara, K.; Perez, N.G.; Ushio-Fukai, M.; Fisher, A.B. Mechanosignaling in the vasculature: Emerging concepts in sensing, transduction and physiological responses. Am. J. Physiol. Heart Circul. Physiol. 2015, 308, H1451–H1462. [Google Scholar] [CrossRef]
- Sanchez-Adams, J.; Leddy, H.A.; McNulty, A.L.; O’Conor, C.J.; Guilak, F. The mechanobiology of articular cartilage: Bearing the burden of osteoarthritis. Curr. Rheumatol. Rep. 2014, 16, 451. [Google Scholar] [CrossRef]
- Burr, D.B.; Gallant, M.A. Bone remodelling in osteoarthritis. Nat. Rev. Rheumatol. 2012, 8, 665–673. [Google Scholar] [CrossRef]
- Reilly, B.D.; Franklin, C.E. Prevention of muscle wasting and osteoporosis: The value of examining novel animal models. J. Exp. Biol. 2016, 219, 2582–2595. [Google Scholar] [CrossRef]
- Kurata, K.; Uemura, T.; Nemoto, A.; Tateishi, T.; Murakami, T.; Higaki, H.; Miura, H.; Iwamoto, Y. Mechanical strain effect on bone-resorbing activity and messenger rna expressions of marker enzymes in isolated osteoclast culture. J. Bone Miner. Res. 2001, 16, 722–730. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Y.; Liu, Y.; Wang, H.; Guo, C.; Zhang, X. Effect of the same mechanical loading on osteogenesis and osteoclastogenesis in vitro. Chin. J. Traumatol. 2015, 18, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Shibata, K.; Yoshimura, Y.; Kikuiri, T.; Hasegawa, T.; Taniguchi, Y.; Deyama, Y.; Suzuki, K.; Iida, J. Effect of the release from mechanical stress on osteoclastogenesis in RAW264.7 cells. Int. J. Mol. Med. 2011, 28, 73–79. [Google Scholar] [PubMed]
- Sinder, B.P.; Pettit, A.R.; McCauley, L.K. Macrophages: Their emerging roles in bone. J. Bone Miner. Res. 2015, 30, 2140–2149. [Google Scholar] [CrossRef] [PubMed]
- Grassel, S.; Muschter, D. Peripheral nerve fibers and their neurotransmitters in osteoarthritis pathology. Int. J. Mol. Sci. 2017, 18, 931. [Google Scholar] [CrossRef] [PubMed]
- Grassel, S.; Muschter, D. Do neuroendocrine peptides and their receptors qualify as novel therapeutic targets in osteoarthritis? Int. J. Mol. Sci. 2018, 19, 367. [Google Scholar] [CrossRef] [PubMed]
- Millward-Sadler, S.J.; Mackenzie, A.; Wright, M.O.; Lee, H.S.; Elliot, K.; Gerrard, L.; Fiskerstrand, C.E.; Salter, D.M.; Quinn, J.P. Tachykinin expression in cartilage and function in human articular chondrocyte mechanotransduction. Arth. Rheum. 2003, 48, 146–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fristad, I.; Vandevska-Radunovic, V.; Fjeld, K.; Wimalawansa, S.J.; Hals Kvinnsland, I. NK1, NK2, NK3 and CGRP1 receptors identified in rat oral soft tissues, and in bone and dental hard tissue cells. Cell Tissue Res. 2003, 311, 383–391. [Google Scholar] [PubMed]
- Ytteborg, E.; Torgersen, J.S.; Pedersen, M.E.; Helland, S.J.; Grisdale-Helland, B.; Takle, H. Exercise induced mechano-sensing and substance P mediated bone modeling in Atlantic salmon. Bone 2013, 53, 259–268. [Google Scholar] [CrossRef]
- Sample, S.J.; Heaton, C.M.; Behan, M.; Bleedorn, J.A.; Racette, M.A.; Hao, Z.; Muir, P. Role of calcitonin gene-related peptide in functional adaptation of the skeleton. PLoS ONE 2014, 9, e113959. [Google Scholar] [CrossRef]
- Heffner, M.A.; Genetos, D.C.; Christiansen, B.A. Bone adaptation to mechanical loading in a mouse model of reduced peripheral sensory nerve function. PLoS ONE 2017, 12, e0187354. [Google Scholar] [CrossRef]
- Niedermair, T.; Schirner, S.; Seebroker, R.; Straub, R.H.; Grassel, S. Substance P modulates bone remodeling properties of murine osteoblasts and osteoclasts. Sci. Rep. 2018, 8, 9199. [Google Scholar] [CrossRef]
- Fernandez, S.; Knopf, M.A.; Bjork, S.K.; McGillis, J.P. Bone marrow-derived macrophages express functional CGRP receptors and respond to CGRP by increasing transcription of c-fos and IL-6 mRNA. Cell. Immunol. 2001, 209, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Tran, A.H.; Paige, C.J. Co-regulated decrease of Neurokinin-1 receptor and hemokinin-1 gene expression in monocytes and macrophages after activation with pro-inflammatory cytokines. J. Neuroimmunol. 2007, 187, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.X.; Zhou, Y.; Zhou, A.Y.; Guan, X.X.; Liu, T.; Yang, H.H.; Xie, H.; Chen, P. Calcitonin gene-related peptide exerts anti-inflammatory property through regulating murine macrophages polarization in vitro. Mol. Immunol. 2017, 91, 105–113. [Google Scholar] [CrossRef]
- Assas, B.M.; Pennock, J.I.; Miyan, J.A. Calcitonin gene-related peptide is a key neurotransmitter in the neuro-immune axis. Front. Neurosci. 2014, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Chai, J.; Zhang, S.; Ding, L.; Yan, P.; Du, W.; Yang, Z. CGRP may regulate bone metabolism through stimulating osteoblast differentiation and inhibiting osteoclast formation. Mol. Med. Rep. 2016, 13, 3977–3984. [Google Scholar] [CrossRef]
- Leal, E.C.; Carvalho, E.; Tellechea, A.; Kafanas, A.; Tecilazich, F.; Kearney, C.; Kuchibhotla, S.; Auster, M.E.; Kokkotou, E.; Mooney, D.J.; et al. Substance P promotes wound healing in diabetes by modulating inflammation and macrophage phenotype. Am. J. Pathol. 2015, 185, 1638–1648. [Google Scholar] [CrossRef]
- Montana, G.; Lampiasi, N. Substance P induces HO-1 expression in RAW 264.7 cells promoting switch towards M2-like macrophages. PLoS ONE 2016, 11, e0167420. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, R.; Shi, X.; Wei, T.; Halloran, B.P.; Clark, D.J.; Jacobs, C.R.; Kingery, W.S. Substance P stimulates bone marrow stromal cell osteogenic activity, osteoclast differentiation, and resorption activity in vitro. Bone 2009, 45, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.A.; Wahbi, A.H.; Nordlin, K. Neuropeptides modulate a murine monocyte/macrophage cell line capacity for phagocytosis and killing of Leishmania major parasites. Immunopharmacol. Immunotoxicol. 2001, 23, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Backman, L.J.; Fong, G.; Andersson, G.; Scott, A.; Danielson, P. Substance P is a mechanoresponsive, autocrine regulator of human tenocyte proliferation. PLoS ONE 2011, 6, e27209. [Google Scholar] [CrossRef] [PubMed]
- Cattaruzza, F.; Poole, D.P.; Bunnett, N.W. Arresting inflammation: Contributions of plasma membrane and endosomal signalling to neuropeptide-driven inflammatory disease. Biochem. Soc. Trans. 2013, 41, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Bullock, C.M.; Wookey, P.; Bennett, A.; Mobasheri, A.; Dickerson, I.; Kelly, S. Peripheral calcitonin gene-related peptide receptor activation and mechanical sensitization of the joint in rat models of osteoarthritis pain. Arth. Rheumatol. 2014, 66, 2188–2200. [Google Scholar] [CrossRef] [PubMed]
- Sample, S.J.; Hao, Z.; Wilson, A.P.; Muir, P. Role of calcitonin gene-related peptide in bone repair after cyclic fatigue loading. PLoS ONE 2011, 6, e20386. [Google Scholar] [CrossRef] [PubMed]
- Hasbak, P.; Sams, A.; Schifter, S.; Longmore, J.; Edvinsson, L. CGRP receptors mediating CGRP-, adrenomedullin- and amylin-induced relaxation in porcine coronary arteries. Characterization with ‘Compound 1’ (WO98/11128), a non-peptide antagonist. Br. J. Pharmacol. 2001, 133, 1405–1413. [Google Scholar] [CrossRef] [Green Version]
- Saxby, D.J.; Lloyd, D.G. Osteoarthritis year in review 2016: Mechanics. Osteoarth. Cartil. 2017, 25, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Muschter, D.; Gottl, C.; Vogel, M.; Grifka, J.; Straub, R.H.; Grassel, S. Reactivity of rat bone marrow-derived macrophages to neurotransmitter stimulation in the context of collagen II-induced arthritis. Arth. Res. Ther. 2015, 17, 169. [Google Scholar] [CrossRef]
- Sager, H.B.; Hulsmans, M.; Lavine, K.J.; Moreira, M.B.; Heidt, T.; Courties, G.; Sun, Y.; Iwamoto, Y.; Tricot, B.; Khan, O.F.; et al. Proliferation and recruitment contribute to myocardial macrophage expansion in chronic heart failure. Circul. Res. 2016, 119, 853–864. [Google Scholar] [CrossRef]
- Adlerz, K.M.; Aranda-Espinoza, H.; Hayenga, H.N. Substrate elasticity regulates the behavior of human monocyte-derived macrophages. Eur. Biophys. J. 2016, 45, 301–309. [Google Scholar] [CrossRef]
- Feng, Y.; Tian, X.Y.; Sun, P.; Cheng, Z.P.; Shi, R.F. Simultaneous study of mechanical stretch-induced cell proliferation and apoptosis on C2C12 myoblasts. Cells Tissues Organs 2018, 205, 189–196. [Google Scholar] [CrossRef]
- Mantella, L.-E.; Quan, A.; Verma, S. Variability in vascular smooth muscle cell stretch-induced responses in 2D culture. Vasc. Cell 2015, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Zhao, D.; Wu, Y.; Xu, C.; Zhang, F. Cyclic stretch induced gene expression of extracellular matrix and adhesion molecules in human periodontal ligament cells. Arch. Oral Biol. 2015, 60, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Opolka, A.; Straub, R.H.; Pasoldt, A.; Grifka, J.; Grassel, S. Substance P and norepinephrine modulate murine chondrocyte proliferation and apoptosis. Arth. Rheum. 2012, 64, 729–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meléndez, G.C.; Manteufel, E.J.; Dehlin, H.M.; Register, T.C.; Levick, S.P. Non-human primate and rat cardiac fibroblasts show similar extracellular matrix-related and cellular adhesion gene responses to substance P. Heart Lung Circul. 2015, 24, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Sloniecka, M.; Le Roux, S.; Zhou, Q.; Danielson, P. Substance P enhances keratocyte migration and neutrophil recruitment through Interleukin-8. Mol. Pharmacol. 2016, 89, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-J.; Yan, H.; Yan, J.; Li, H.; Chen, L.; Han, L.-R.; Yang, X.-F. Substance P promotes the proliferation, but inhibits differentiation and mineralization of osteoblasts from rats with spinal cord injury via RANKL/OPG system. PLoS ONE 2016, 11, e0165063. [Google Scholar] [CrossRef]
- Backman, L.J.; Danielson, P. Akt-mediated anti-apoptotic effects of substance P in Anti-Fas-induced apoptosis of human tenocytes. J. Cell. Mol. Med. 2013, 17, 723–733. [Google Scholar] [CrossRef] [Green Version]
- Gross, K.; Karagiannides, I.; Thomou, T.; Koon, H.W.; Bowe, C.; Kim, H.; Giorgadze, N.; Tchkonia, T.; Pirtskhalava, T.; Kirkland, J.L.; et al. Substance P promotes expansion of human mesenteric preadipocytes through proliferative and antiapoptotic pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1012–G1019. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.E.; Chung, E.; Son, Y. A neuropeptide, Substance-P, directly induces tissue-repairing M2 like macrophages by activating the PI3K/Akt/mTOR pathway even in the presence of IFNgamma. Sci. Rep. 2017, 7, 9417. [Google Scholar] [CrossRef]
- Hao, S.; Meng, J.; Zhang, Y.; Liu, J.; Nie, X.; Wu, F.; Yang, Y.; Wang, C.; Gu, N.; Xu, H. Macrophage phenotypic mechanomodulation of enhancing bone regeneration by superparamagnetic scaffold upon magnetization. Biomaterials 2017, 140, 16–25. [Google Scholar] [CrossRef]
- Baliu-Piqué, M.; Jusek, G.; Holzmann, B. Neuroimmunological communication via CGRP promotes the development of a regulatory phenotype in TLR4-stimulated macrophages. Eur. J. Immunol. 2014, 44, 3708–3716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitsin, S.; Meshki, J.; Winters, A.; Tuluc, F.; Benton, T.D.; Douglas, S.D. Substance P-mediated chemokine production promotes monocyte migration. J. Leukoc. Biol. 2017, 101, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Fukuda, K.; Matsushita, T.; Matsukawa, M.; Hara, F.; Hamanishi, C. Cyclic tensile stretch stimulates the release of reactive oxygen species from osteoblast-like cells. Calcif. Tissue Int. 2005, 76, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yan, J.; He, F.; Zhong, D.; Yang, H.; Pei, M.; Luo, Z.P. Mechanical stretch induces antioxidant responses and osteogenic differentiation in human mesenchymal stem cells through activation of the AMPK-SIRT1 signaling pathway. Free Radic. Biol. Med. 2018, 126, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sui, W.; Li, Y.; Qi, X.; Wang, Y.; Zhou, Q.; Gao, H. Substance P inhibits hyperosmotic stress-induced apoptosis in corneal epithelial cells through the mechanism of Akt activation and reactive oxygen species scavenging via the Neurokinin-1 receptor. PLoS ONE 2016, 11, e0149865. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.X.; Fang, F.; Jiang, J.L.; Xu, F. Extrinsic calcitonin gene-related peptide inhibits hyperoxia-induced alveolar epithelial type II cells apoptosis, oxidative stress, and reactive oxygen species (ROS) production by enhancing notch 1 and Homocysteine-induced endoplasmic reticulum protein (HERP) expression. Med. Sci. Monit. 2017, 23, 5774–5782. [Google Scholar] [PubMed]
- Wang, L.; Shi, X.; Zhao, R.; Halloran, B.P.; Clark, D.J.; Jacobs, C.R.; Kingery, W.S. Calcitonin-gene-related peptide stimulates stromal cell osteogenic differentiation and inhibits RANKL induced NF-kappaB activation, osteoclastogenesis and bone resorption. Bone 2010, 46, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Kadow-Romacker, A.; Hoffmann, J.E.; Duda, G.; Wildemann, B.; Schmidmaier, G. Effect of mechanical stimulation on osteoblast- and osteoclast-like cells in vitro. Cells Tissues Organs 2009, 190, 61–68. [Google Scholar] [CrossRef]
- Li, F.; Sun, X.; Zhao, B.; Ma, J.; Zhang, Y.; Li, S.; Li, Y.; Ma, X. Effects of cyclic tension stress on the apoptosis of osteoclasts in vitro. Exp. Ther. Med. 2015, 9, 1955–1961. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.H.; Sakamoto, H.; Xu, E.C.; Lee, R.T. Biomechanical regulation of human monocyte/macrophage molecular function. Am. J. Pathol. 2000, 156, 1797–1804. [Google Scholar] [CrossRef]
- Yarwood, R.E.; Imlach, W.L.; Lieu, T.; Veldhuis, N.A.; Jensen, D.D.; Klein Herenbrink, C.; Aurelio, L.; Cai, Z.; Christie, M.J.; Poole, D.P.; et al. Endosomal signaling of the receptor for calcitonin gene-related peptide mediates pain transmission. Proc. Natl. Acad. Sci. USA 2017, 114, 12309–12314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasson, S.S.; Blanchet, T.J.; Morris, E.A. The surgical destabilization of the medial meniscus (DMM) model of osteoarthritis in the 129/SvEv mouse. Osteoarth. Cartil. 2007, 15, 1061–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, J.P.; Ho, W.Z.; Kilpatrick, L.E.; Wang, X.; Tuluc, F.; Korchak, H.M.; Douglas, S.D. Full-length and truncated neurokinin-1 receptor expression and function during monocyte/macrophage differentiation. Proc. Natl. Acad. Sci. USA 2006, 103, 7771–7776. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | FORWARD: 5′-3′ SEQUENZ REVERSE: 5′-3′ SEQUENZ | Amplicon Size (bp) |
|---|---|---|
| Neuropeptides and Receptor Genes | ||
| NK1R | Fwd: ATTGAGTGGCCAGAACATCC Rev: ACTGGCCCACAGTGTAATCC | 135 |
| CRLR | Fwd: GCCAATAACCAGGCCTTAGTG Rev: GCCCATCAGGTAGAGATGGAT | 77 |
| Ramp1 | Fwd: CCTGACTATGGGACTCTCATCC Rev: CGTGCTTGGTGCAGTAAGTG | 139 |
| SP | Fwd: GATGAAGGAGCTGTCCAAGC Rev: GCACAGGAGTCTCTGCTTCC | 102 |
| CGRP | Fwd: TGCAGGACTATATGCAGATGAAA Rev: GGATCTCTTCTGAGCAGTGACA | 91 |
| Osteoclastogenesis-Related Genes | ||
| CSF1R | Fwd: CAGAAGACCCACCTTCCAAC Rev: CTGCTTGGCAGGTTAGCATA | 93 |
| Rank | Fwd: GCTCCTGAAATGTGGACCAT Rev: CACGATGATGTCACCCTTGA | 241 |
| Macrophage Polarization Genes | ||
| NFκB | Fwd: GGCAGCTCTTCTCAAAGCAG Rev: CCACTCCCTCATCTTCTCCA | 107 |
| TNFα | Fwd: GACAGTGACCTGGACTGTGG Rev: GAGACAGAGGCAACCTGACC | 132 |
| iNOS | Fwd: CAAGCACCTTGGAAGAGGAG Rev: AAGGCCAAACACAGCATACC | 149 |
| IL-6 | Fwd: CCGGAGAGGAGACTTCACAG Rev: CAGAATTGCCATTGCACAAC | 134 |
| IL-10 | Fwd: CCAAGCCTTATCGGAAATGA Rev: TTTTCACAGGGGAGAAATCG | 162 |
| Mrc1 | Fwd: AGAAAATGCACAAGAGCAAGC Rev: GGAACATGTGTTCTGCGTTG | 101 |
| KLF4 | Fwd: CCGTCCTTCTCCACGTTC Rev: GAGTTCCTCACGCCAACG | 93 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muschter, D.; Beiderbeck, A.-S.; Späth, T.; Kirschneck, C.; Schröder, A.; Grässel, S. Sensory Neuropeptides and their Receptors Participate in Mechano-Regulation of Murine Macrophages. Int. J. Mol. Sci. 2019, 20, 503. https://doi.org/10.3390/ijms20030503
Muschter D, Beiderbeck A-S, Späth T, Kirschneck C, Schröder A, Grässel S. Sensory Neuropeptides and their Receptors Participate in Mechano-Regulation of Murine Macrophages. International Journal of Molecular Sciences. 2019; 20(3):503. https://doi.org/10.3390/ijms20030503
Chicago/Turabian StyleMuschter, Dominique, Anna-Sophie Beiderbeck, Tanja Späth, Christian Kirschneck, Agnes Schröder, and Susanne Grässel. 2019. "Sensory Neuropeptides and their Receptors Participate in Mechano-Regulation of Murine Macrophages" International Journal of Molecular Sciences 20, no. 3: 503. https://doi.org/10.3390/ijms20030503