Synergistic Effects of Nanomedicine Targeting TNFR2 and DNA Demethylation Inhibitor—An Opportunity for Cancer Treatment

 ,
,  ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Implication of TNFR2 in Cancer Development
3. Nanomedicine Applications for Cancer Immunotherapy
4. DNA Demethylation and Immune Evasion
5. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aggarwal, B.B.; Gupta, S.C.; Kim, J.H. Historical perspectives on tumor necrosis factor and its superfamily: 25 years later, a golden journey. Blood 2012, 119, 651–665. [Google Scholar] [CrossRef] [Green Version]
- Sedger, L.M.; McDermott, M.F. TNF and TNF-receptors: From mediators of cell death and inflammation to therapeutic giants–past, present and future. Cytokine Growth Factor Rev. 2014, 25, 453–472. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Azid, N.A.; Boer, J.C.; Lim, J.; Chen, X.; Plebanski, M.; Mohamud, R. The Key Role of TNF-TNFR2 Interactions in the Modulation of Allergic Inflammation: A Review. Front. Immunol. 2018, 9, 2572. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Ham, B.; Fernandez, M.C.; D’Costa, Z.; Brodt, P. The diverse roles of the TNF axis in cancer progression and metastasis. Trends Cancer Res. 2016, 11, 1–27. [Google Scholar] [PubMed]
- Hamilton, K.E.; Simmons, J.G.; Ding, S.; Van Landeghem, L.; Lund, P.K. Cytokine induction of tumor necrosis factor receptor 2 is mediated by STAT3 in colon cancer cells. Mol. Cancer Res. 2011, 9, 1718–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onizawa, M.; Nagaishi, T.; Kanai, T.; Nagano, K.; Oshima, S.; Nemoto, Y.; Yoshioka, A.; Totsuka, T.; Okamoto, R.; Nakamura, T.; et al. Signaling pathway via TNF-alpha/NF-kappaB in intestinal epithelial cells may be directly involved in colitis-associated carcinogenesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G850–G859. [Google Scholar] [CrossRef]
- Cai, X.; Cao, C.; Li, J.; Chen, F.; Zhang, S.; Liu, B.; Zhang, W.; Zhang, X.; Ye, L. Inflammatory factor TNF-alpha promotes the growth of breast cancer via the positive feedback loop of TNFR1/NF-kappaB (and/or p38)/p-STAT3/HBXIP/TNFR1. Oncotarget 2017, 8, 58338–58352. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.; Li, F.; Qin, Z. TNF Receptor 2 Makes Tumor Necrosis Factor a Friend of Tumors. Front. Immunol. 2018, 9, 1170. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Zhao, Z.; Zhao, N. Clinical implications of tumor necrosis factor receptor 2 in breast cancer. Oncol. Lett. 2017, 14, 2393–2398. [Google Scholar] [CrossRef] [Green Version]
- Torrey, H.; Butterworth, J.; Mera, T.; Okubo, Y.; Wang, L.; Baum, D.; Defusco, A.; Plager, S.; Warden, S.; Huang, D. Targeting TNFR2 with antagonistic antibodies inhibits proliferation of ovarian cancer cells and tumor-associated Tregs. Sci. Signal. 2017, 10, eaaf8608. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Min, Y.; Rodgers, Z.; Zhang, L.; Wang, A.Z. Nanomedicine approaches to improve cancer immunotherapy. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2017, 9, e1456. [Google Scholar] [CrossRef] [PubMed]
- Mohamud, R.; Xiang, S.D.; Selomulya, C.; Rolland, J.M.; O’Hehir, R.E.; Hardy, C.L.; Plebanski, M. The effects of engineered nanoparticles on pulmonary immune homeostasis. Drug Metab. Rev. 2014, 46, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Mohamud, R.; LeMasurier, J.S.; Boer, J.C.; Sieow, J.L.; Rolland, J.M.; O’Hehir, R.E.; Hardy, C.L.; Plebanski, M. Synthetic Nanoparticles That Promote Tumor Necrosis Factor Receptor 2 Expressing Regulatory T Cells in the Lung and Resistance to Allergic Airways Inflammation. Front. Immunol. 2017, 8, 1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, C.L.; Lemasurier, J.S.; Mohamud, R.; Yao, J.; Xiang, S.D.; Rolland, J.M.; O’ Hehir, R.E.; Plebanski, M. Differential uptake of nanoparticles and microparticles by pulmonary APC subsets induces discrete immunological imprints. J. Immunol. 2013, 191, 5278–5290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hatamleh, M.A.; Ahmad, S.; Boer, J.C.; Lim, J.; Chen, X.; Plebanski, M.; Mohamud, R. A Perspective Review on the Role of Nanomedicine in the Modulation of TNF-TNFR2 Axis in Breast Cancer Immunotherapy. J. Oncol. 2019, 2019, 13. [Google Scholar] [CrossRef]
- Calvanese, V.; Fernandez, A.F.; Urdinguio, R.G.; Suarez-Alvarez, B.; Mangas, C.; Perez-Garcia, V.; Bueno, C.; Montes, R.; Ramos-Mejia, V.; Martinez-Camblor, P.; et al. A promoter DNA demethylation landscape of human hematopoietic differentiation. Nucleic Acids Res. 2012, 40, 116–131. [Google Scholar] [CrossRef]
- Kitagawa, Y.; Ohkura, N.; Sakaguchi, S. Molecular determinants of regulatory T cell development: The essential roles of epigenetic changes. Front. Immunol. 2013, 4, 106. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, G.; Asemissen, A.; Model, F.; Turbachova, I.; Floess, S.; Liebenberg, V.; Baron, U.; Stauch, D.; Kotsch, K.; Pratschke, J.; et al. Quantitative DNA methylation analysis of FOXP3 as a new method for counting regulatory T cells in peripheral blood and solid tissue. Cancer Res. 2009, 69, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Punit, S.; Dube, P.E.; Liu, C.Y.; Girish, N.; Washington, M.K.; Polk, D.B. Tumor Necrosis Factor Receptor 2 Restricts the Pathogenicity of CD8(+) T Cells in Mice with Colitis. Gastroenterology 2015, 149, 993–1005. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, N.B.; Maker, A.V. Gene-expression profiling to predict responsiveness to immunotherapy. Cancer Gene Ther. 2017, 24, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tartaglia, L.A.; Pennica, D.; Goeddel, D.V. Ligand passing: The 75-kDa tumor necrosis factor (TNF) receptor recruits TNF for signaling by the 55-kDa TNF receptor. J. Biol. Chem. 1993, 268, 18542–18548. [Google Scholar] [PubMed]
- Tartaglia, L.A.; Goeddel, D.V. Two TNF receptors. Immunol. Today 1992, 13, 151–153. [Google Scholar] [CrossRef]
- Liu, Z.-G.; Hsu, H.; Goeddel, D.V.; Karin, M. Dissection of TNF receptor 1 effector functions: JNK activation is not linked to apoptosis while NF-κB activation prevents cell death. Cell 1996, 87, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Du, R.; Wei, F.; Zhao, H.; Yu, J.; Wang, C.; Zhan, Z.; Ding, T.; Ren, X.; Chen, X. Expression of TNFR2 by regulatory T cells in peripheral blood is correlated with clinical pathology of lung cancer patients. Cancer Immunol. Immunother. 2015, 64, 1475–1485. [Google Scholar] [CrossRef]
- Vanamee, E.S.; Faustman, D.L. TNFR2: A Novel Target for Cancer Immunotherapy. Trends Mol. Med. 2017, 23, 1037–1046. [Google Scholar] [CrossRef]
- Zhang, T.; Jiao, J.; Jiao, X.; Zhao, L.; Tian, X.; Zhang, Q.; Ma, D.; Cui, B. Aberrant frequency of TNFR2(+) Treg and related cytokines in patients with CIN and cervical cancer. Oncotarget 2017, 9, 5073–5083. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; He, J.; Shirota, H.; Trivett, A.L.; Yang, D.; Klinman, D.M.; Oppenheim, J.J.; Chen, X. Blockade of TNFR2 signaling enhances the immunotherapeutic effect of CpG ODN in a mouse model of colon cancer. Sci. Signal 2018, 11, 1937–9145. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Al-Lamki, R.S. Tumor necrosis factor receptor 2: Its contribution to acute cellular rejection and clear cell renal carcinoma. BioMed Res. Int. 2013, 2013, 82131. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.L.; Wood, K.J. TNFR2: The new Treg switch? Oncoimmunology 2018, 7, e1373236. [Google Scholar] [CrossRef] [Green Version]
- Arcaro, A.; Guerreiro, A.S. The phosphoinositide 3-kinase pathway in human cancer: Genetic alterations and therapeutic implications. Curr. Genom. 2007, 8, 271–306. [Google Scholar] [CrossRef] [PubMed]
- Nur Husna, S.M.; Tan, H.T.; Mohamud, R.; Dyhl-Polk, A.; Wong, K.K. Inhibitors targeting CDK4/6, PARP and PI3K in breast cancer: A review. Ther. Adv. Med. Oncol. 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Wang, J.; Brand, D.D.; Zheng, S.G. Role of TNF–TNF Receptor 2 Signal in Regulatory T Cells and Its Therapeutic Implications. Front. Immunol. 2018, 9, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, A.; Murphy, J.J. STAT5 in Cancer and Immunity. J. Interferon Cytokine Res. 2016, 36, 226–237. [Google Scholar] [CrossRef]
- Taylor, A.; Verhagen, J.; Blaser, K.; Akdis, M.; Akdis, C.A. Mechanisms of immune suppression by interleukin-10 and transforming growth factor-beta: The role of T regulatory cells. Immunology 2006, 117, 433–442. [Google Scholar] [CrossRef]
- Chinen, T.; Kannan, A.K.; Levine, A.G.; Fan, X.; Klein, U.; Zheng, Y.; Gasteiger, G.; Feng, Y.; Fontenot, J.D.; Rudensky, A.Y. An essential role for the IL-2 receptor in Treg cell function. Nat. Immunol. 2016, 17, 1322–1333. [Google Scholar] [CrossRef]
- Holbrook, J.; Lara-Reyna, S.; Jarosz-Griffiths, H.; McDermott, M. Tumor necrosis factor signaling in health and disease. F1000Research 2019, 8, 111. [Google Scholar] [CrossRef]
- Pan, S.; An, P.; Zhang, R.; He, X.; Yin, G.; Min, W. Etk/Bmx as a tumor necrosis factor receptor type 2-specific kinase: Role in endothelial cell migration and angiogenesis. Mol. Cell. Biol. 2002, 22, 7512–7523. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Subleski, J.J.; Kopf, H.; Howard, O.M.; Mannel, D.N.; Oppenheim, J.J. Cutting edge: Expression of TNFR2 defines a maximally suppressive subset of mouse CD4+CD25+FoxP3+ T regulatory cells: Applicability to tumor-infiltrating T regulatory cells. J. Immunol. 2008, 180, 6467–6471. [Google Scholar] [CrossRef] [Green Version]
- Williams, G.S.; Mistry, B.; Guillard, S.; Ulrichsen, J.C.; Sandercock, A.M.; Wang, J.; Gonzalez-Munoz, A.; Parmentier, J.; Black, C.; Soden, J.; et al. Phenotypic screening reveals TNFR2 as a promising target for cancer immunotherapy. Oncotarget 2016, 7, 68278–68291. [Google Scholar] [CrossRef] [Green Version]
- Nikalje, A.P. Nanotechnology and its applications in medicine. Med. Chem. 2015, 5, 81–89. [Google Scholar] [CrossRef]
- Farokhzad, O.C.; Langer, R. Impact of nanotechnology on drug delivery. ACS Nano 2009, 3, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Mirza, A.Z.; Siddiqui, F.A. Nanomedicine and drug delivery: A mini review. Int. Nano Lett. 2014, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.; Lyon, L.A. Soft nanotechnology with soft nanoparticles. Angew. Chem. Int. Ed. 2005, 44, 7686–7708. [Google Scholar] [CrossRef] [PubMed]
- Din, F.U.; Aman, W.; Ullah, I.; Qureshi, O.S.; Mustapha, O.; Shafique, S.; Zeb, A. Effective use of nanocarriers as drug delivery systems for the treatment of selected tumors. Int. J. Nanomed. 2017, 12, 7291–7309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Zhang, L.; Chen, T.; Guo, W.; Bao, X.; Wang, D.; Ren, B.; Wang, H.; Li, Y.; Wang, Y.; et al. Anticancer Effects of Resveratrol-Loaded Solid Lipid Nanoparticles on Human Breast Cancer Cells. Molecules 2017, 22, 1814. [Google Scholar] [CrossRef]
- Ferji, K.; Nouvel, C.C.; Babin, J.; Li, M.-H.; Gaillard, C.D.; Nicol, E.; Chassenieux, C.; Six, J.-L. Polymersomes from amphiphilic glycopolymers containing polymeric liquid crystal grafts. ACS Macro Lett. 2015, 4, 1119–1122. [Google Scholar] [CrossRef]
- Six, J.-L.; Ferji, K. Polymerization induced self-assembly: An opportunity toward the self-assembly of polysaccharide-containing copolymers into high-order morphologies. Polym. Chem. 2019, 10, 45–53. [Google Scholar] [CrossRef]
- Sugano, M.; Egilmez, N.K.; Yokota, S.J.; Chen, F.A.; Harding, J.; Huang, S.K.; Bankert, R.B. Antibody targeting of doxorubicin-loaded liposomes suppresses the growth and metastatic spread of established human lung tumor xenografts in severe combined immunodeficient mice. Cancer Res. 2000, 60, 6942–6949. [Google Scholar]
- Zhong, Q.; Bielski, E.R.; Rodrigues, L.S.; Brown, M.R.; Reineke, J.J.; da Rocha, S.R. Conjugation to Poly(amidoamine) Dendrimers and Pulmonary Delivery Reduce Cardiac Accumulation and Enhance Antitumor Activity of Doxorubicin in Lung Metastasis. Mol. Pharm. 2016, 13, 2363–2375. [Google Scholar] [CrossRef]
- Poon, C.; He, C.; Liu, D.; Lu, K.; Lin, W. Self-assembled nanoscale coordination polymers carrying oxaliplatin and gemcitabine for synergistic combination therapy of pancreatic cancer. J. Control. Release 2015, 201, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafael, D.; Gener, P.; Andrade, F.; Seras-Franzoso, J.; Montero, S.; Fernandez, Y.; Hidalgo, M.; Arango, D.; Sayos, J.; Florindo, H.F.; et al. AKT2 siRNA delivery with amphiphilic-based polymeric micelles show efficacy against cancer stem cells. Drug Deliv. 2018, 25, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Wen, A.M.; Commandeur, U.; Steinmetz, N.F. Presentation of HER2 epitopes using a filamentous plant virus-based vaccination platform. J. Mater. Chem. B 2014, 2, 6249–6258. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, W.; Meng, J.; Chen, S.; Xu, H.; Yang, X.D. Multi-walled carbon nanotubes conjugated to tumor protein enhance the uptake of tumor antigens by human dendritic cells in vitro. Cell Res. 2010, 20, 1170–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Wei, X.; Jing, Y.; Zhou, S. Size Changeable Nanocarriers with Nuclear Targeting for Effectively Overcoming Multidrug Resistance in Cancer Therapy. Adv. Mater. 2015, 27, 6450–6456. [Google Scholar] [CrossRef]
- Seyfoori, A.; Sarfarazijami, S.; Seyyed Ebrahimi, S.A. pH-responsive carbon nanotube-based hybrid nanogels as the smart anticancer drug carrier. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1437–1443. [Google Scholar] [CrossRef] [Green Version]
- Palucka, K.; Banchereau, J. Cancer immunotherapy via dendritic cells. Nat. Rev. Cancer 2012, 12, 265–277. [Google Scholar] [CrossRef]
- Sehgal, K.; Dhodapkar, K.M.; Dhodapkar, M.V. Targeting human dendritic cells in situ to improve vaccines. Immunol. Lett. 2014, 162, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Zamry, A.A.; Tan, H.T.; Wong, K.K.; Lim, J.; Mohamud, R. Targeting dendritic cells through gold nanoparticles: A review on the cellular uptake and subsequent immunological properties. Mol. Immunol. 2017, 91, 123–133. [Google Scholar] [CrossRef]
- Sehgal, K.; Ragheb, R.; Fahmy, T.M.; Dhodapkar, M.V.; Dhodapkar, K.M. Nanoparticle-mediated combinatorial targeting of multiple human dendritic cell (DC) subsets leads to enhanced T cell activation via IL-15-dependent DC crosstalk. J. Immunol. 2014, 193, 2297–2305. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Miao, L.; Wang, Y.; Xu, Z.; Zhao, Y.; Shen, Y.; Xiang, G.; Huang, L. Curcumin Micelles Remodel Tumor Microenvironment and Enhance Vaccine Activity in an Advanced Melanoma Model. Mol. Ther. 2016, 24, 364–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.Y.; Liu, Y.; Xu, C.F.; Shen, S.; Sun, R.; Du, X.J.; Xia, J.X.; Zhu, Y.H.; Wang, J. Restoring anti-tumor functions of T cells via nanoparticle-mediated immune checkpoint modulation. J. Control. Release 2016, 231, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wang, Y.; Zhang, L.; Huang, L. Nanoparticle-delivered transforming growth factor-beta siRNA enhances vaccination against advanced melanoma by modifying tumor microenvironment. ACS Nano 2014, 8, 3636–3645. [Google Scholar] [CrossRef] [PubMed]
- Roeven, M.W.; Hobo, W.; van der Voort, R.; Fredrix, H.; Norde, W.J.; Teijgeler, K.; Ruiters, M.H.; Schaap, N.; Dolstra, H. Efficient nontoxic delivery of PD-L1 and PD-L2 siRNA into dendritic cell vaccines using the cationic lipid SAINT-18. J. Immunother. 2015, 38, 145–154. [Google Scholar] [CrossRef]
- Nicodemus, C.F. Antibody-based immunotherapy of solid cancers: Progress and possibilities. Immunotherapy 2015, 7, 923–939. [Google Scholar] [CrossRef] [Green Version]
- Conniot, J.; Silva, J.M.; Fernandes, J.G.; Silva, L.C.; Gaspar, R.; Brocchini, S.; Florindo, H.F.; Barata, T.S. Cancer immunotherapy: Nanodelivery approaches for immune cell targeting and tracking. Front. Chem. 2014, 2, 105. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Fang, M.; Zhang, J.; Wang, J.; Song, Y.; Shi, J.; Li, W.; Wu, G.; Ren, J.; Wang, Z.; et al. Hydrogel dual delivered celecoxib and anti-PD-1 synergistically improve antitumor immunity. Oncoimmunology 2016, 5, e1074374. [Google Scholar] [CrossRef]
- Twyman-Saint Victor, C.; Rech, A.J.; Maity, A.; Rengan, R.; Pauken, K.E.; Stelekati, E.; Benci, J.L.; Xu, B.; Dada, H.; Odorizzi, P.M.; et al. Radiation and dual checkpoint blockade activate non-redundant immune mechanisms in cancer. Nature 2015, 520, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Kwong, B.; Gai, S.A.; Elkhader, J.; Wittrup, K.D.; Irvine, D.J. Localized immunotherapy via liposome-anchored Anti-CD137 + IL-2 prevents lethal toxicity and elicits local and systemic antitumor immunity. Cancer Res. 2013, 73, 1547–1558. [Google Scholar] [CrossRef] [Green Version]
- Lei, C.; Liu, P.; Chen, B.; Mao, Y.; Engelmann, H.; Shin, Y.; Jaffar, J.; Hellstrom, I.; Liu, J.; Hellstrom, K.E. Local release of highly loaded antibodies from functionalized nanoporous support for cancer immunotherapy. J. Am. Chem. Soc. 2010, 132, 6906–6907. [Google Scholar] [CrossRef]
- Kisseljova, N.P.; Kisseljov, F.L. DNA demethylation and carcinogenesis. Biochemistry 2005, 70, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Szyf, M. The role of DNA hypermethylation and demethylation in cancer and cancer therapy. Curr. Oncol. 2008, 15, 72–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elashi, A.A.; Sasidharan Nair, V.; Taha, R.Z.; Shaath, H.; Elkord, E. DNA methylation of immune checkpoints in the peripheral blood of breast and colorectal cancer patients. Oncoimmunology 2019, 8, e1542918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasidharan Nair, V.; El Salhat, H.; Taha, R.Z.; John, A.; Ali, B.R.; Elkord, E. DNA methylation and repressive H3K9 and H3K27 trimethylation in the promoter regions of PD-1, CTLA-4, TIM-3, LAG-3, TIGIT and PD-L1 genes in human primary breast cancer. Clin. Epigenet. 2018, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.X.; Riggs, A.D. DNA methylation and demethylation in mammals. J. Biol. Chem. 2011, 286, 18347–18353. [Google Scholar] [CrossRef] [Green Version]
- Heninger, E.; Krueger, T.E.; Lang, J.M. Augmenting antitumor immune responses with epigenetic modifying agents. Front. Immunol. 2015, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Guo, M. Epigenetic changes in colorectal cancer. Chin. J. Cancer 2013, 32, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Lao, V.V.; Grady, W.M. Epigenetics and colorectal cancer. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 686–700. [Google Scholar] [CrossRef]
- Issa, J.P. CpG island methylator phenotype in cancer. Nat. Rev. Cancer 2004, 4, 988–993. [Google Scholar] [CrossRef]
- Schlesinger, Y.; Straussman, R.; Keshet, I.; Farkash, S.; Hecht, M.; Zimmerman, J.; Eden, E.; Yakhini, Z.; Ben-Shushan, E.; Reubinoff, B.E.; et al. Polycomb-mediated methylation on Lys27 of histone H3 pre-marks genes for de novo methylation in cancer. Nat. Genet. 2007, 39, 232–236. [Google Scholar] [CrossRef]
- Mohan, N.; Edwards, E.T.; Cupps, T.R.; Oliverio, P.J.; Sandberg, G.; Crayton, H.; Richert, J.R.; Siegel, J.N. Demyelination occurring during anti-tumor necrosis factor alpha therapy for inflammatory arthritides. Arthritis Rheumatol. 2001, 44, 2862–2869. [Google Scholar] [CrossRef]
- Kemanetzoglou, E.; Andreadou, E. CNS Demyelination with TNF-alpha Blockers. Curr. Neurol. Neurosci. Rep. 2017, 17, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
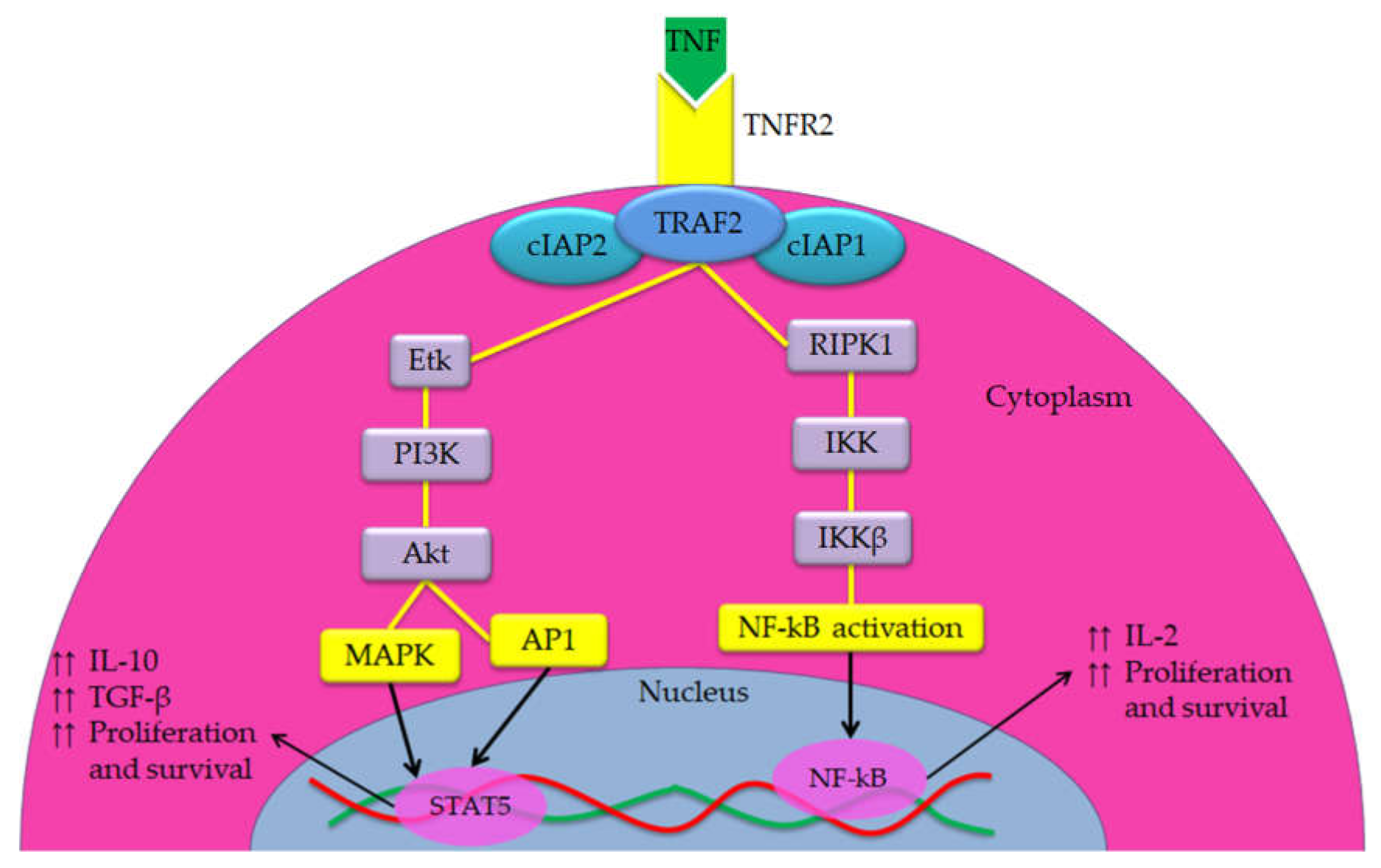{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Samples Used and Diagnosis Assay | Significant Findings |
|---|---|---|
| Yan et al., 2015 [25] |
|
|
| Williams et al., 2016 [40] |
|
|
| Zhang et al., 2017 [27] |
|
|
| Torrey et al., 2017 [11] |
|
|
| Nie et al., 2018 [28] |
|
|
| Nanocarrier Type [Size Range] | Significant Properties | Selected Studies |
|---|---|---|
| Organic nanocarriers | ||
| Solid lipid nanoparticles (SLNs) [50–200 nm] | Economical large-scale production, high drug payload, better stability and easy to handle, improved bioavailability of poorly water-soluble drugs, as well as lack of biotoxicity. | In 2017, Wang et al. have designed resveratrol-loaded SLNs (Res-SLNs) to treat breast cancer cells. They showed that Res-SLNs significantly exhibited the inhibitory effects on cancer cells proliferation, invasion and migration, compared to controls [46]. |
| Liposomes and polymersomes [30 nm–110 nm] | Enhanced delivery of drugs, preventing early degradation of the encapsulated drug, cost-effective formulations of expensive drugs and efficient treatment, improved performance features of the product, protection of the active drug from environmental factors, as well as reduced systemic toxicity. Weak mechanical properties of liposomes may be enhanced by using polymeric liposomes called polymersomes [47]. Possibility to encapsulate hydrophobic drugs in the lipidic bilayer, as well as hydrophilic drugs in the hydrophilic core [48]. | Doxorubicin (DOX)-loaded specific monoclonal antibodies conjugated to liposomes were used to treat lung tumor in mice. The results showed significant suppression of tumor growth, metastatic spread and increased the survival rate of the tumor-bearing mice compared to controls [49]. |
| Dendrimers [1.5–14.5 nm] | The most utilized nanocarrier owing to their incomparable characteristics, including the increased number of branching, distinctive molecular weight, monodispersed macromolecules, multivalency and spherical shapes. | In treating of lung metastasis mouse model, as indicated by increased survival rates and decreased tumor burden, DOX conjugated to carboxyl-terminated poly(amidoamine) dendrimers (PAMAM) was more effective than DOX delivered intravenously [50]. |
| Polymeric nanoparticles (PNPs) [10–200 nm] | Two types:
PNPs are highly versatile based on a wide range of polymers from synthetic and natural sources. The modification of the physicochemical properties of the polymers used to produce PNPs can accurately control the degradation of PNPs and drug release. | Nanoscale coordination polymer-1 (NCP-1) has been used for simultaneous delivery of Oxaliplatin and Gemcitabine monophosphate. NCP-1 particles effectively avoided uptake by the mononuclear phagocyte system (MPS), which is resulted in potently delivery for both drugs and thus a strong synergistic therapeutic effect was observed against pancreatic cancer cells by inhibiting tumor growth [51]. |
| Polymeric micelles (PMs) [10–100 nm] | PMs allow hydrophobic drugs to be entrapped into their cores which enhance their water solubility. The hydrophilic shell of PMs promotes their stability and their circulation times in blood by preventing the recognition and subsequent uptake of it by the reticule endothelial system. | Pluronic micelles combined with polyplexes spontaneously were used as amphiphilic-based gene delivery system with two breast cancer cell lines. This system has been formed by electrostatic interaction between cationic polyethyleneimine and anionic siRNA against AKT2. After treatment, a significant reduction was observed on cell invasion capacity, as well as a significant inhibition of mammosphere formation [52]. |
| Virus-based nanoparticles (VNPs) [up to 100 nm] | VNPs emerged based on their easy surface functionalization, availability in a variety of sizes and shapes, in addition to their biocompatibility and morphological uniformity. | To overcome immunological tolerance against human epidermal growth factor receptor 2 (HER2) in breast cancer mouse model, HER2 epitopes were integrated on the plant-produced vaccination platform potato virus X (PVX). The results showed that this carrier stimulated the production of HER2-specific antibodies in the injected mice [53]. |
| Inorganic nanocarriers | ||
| Carbon nanotubes (CNTs) [0.4–100 nm] | Able to cross the cell membrane via endocytosis and subsequently enter into the cell; based on their needle-like shape. With its physicochemical characteristics, CNTs able to carry high drug amounts, structural flexibility and intrinsic stability and appropriate surface functionalization. | In 2010, Sun et al. have conjugated MCF7 breast cancer cells−derived tumor lysate (covalent) to Carboxylated MWNT (CNTs) in vitro. They reported increased antigen uptake by dendritic cells (DCs) and improved the induction of tumor−specific T cell response by DCs, thus enhance the uptake of tumor antigens [54]. |
| Mesoporous silica nanoparticles (MSNs) [2–50 nm] | Owing to their honeycomb-like shape with hundreds of pores, MSNs able to load large drugs amounts. Based on their ease of surface functionalization for targeted and controlled drug delivery, MSNs reduce the toxicity of drugs and promote therapeutic efficacy. | Guo et al. have used MSNs with nuclear targeting in cancer therapy for multidrug resistance (MDR) breast cancer cells. They used a size changeable MSNs able to alter to smaller micelles under specific conditions. This study reported this type of MSNs as a highly effective delivery system for anticancer drugs to the nucleus of MDR cancer cells, directly [55]. |
| Hybrid nanocarriers | A combination of organic and inorganic NPs. This combination successfully employed specific functionalities of both NPs to enhance the selectivity and efficiency of drugs along with high payload sustained and intracellular delivery. | A multifunctional hybrid nanocarrier was developed by merging the properties of pH-sensitive nanogels and multiwall carbon nanotube, to deliver the DOX. This study showed a significant effect of DOX supernatant with this hybrid nanocarrier on the U-87 glioblastoma cancer cells proliferation suppression [56]. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Hatamleh, M.A.I.; E.A.R., E.N.S.; Boer, J.C.; Ferji, K.; Six, J.-L.; Chen, X.; Elkord, E.; Plebanski, M.; Mohamud, R. Synergistic Effects of Nanomedicine Targeting TNFR2 and DNA Demethylation Inhibitor—An Opportunity for Cancer Treatment. Cells 2020, 9, 33. https://doi.org/10.3390/cells9010033
Al-Hatamleh MAI, E.A.R. ENS, Boer JC, Ferji K, Six J-L, Chen X, Elkord E, Plebanski M, Mohamud R. Synergistic Effects of Nanomedicine Targeting TNFR2 and DNA Demethylation Inhibitor—An Opportunity for Cancer Treatment. Cells. 2020; 9(1):33. https://doi.org/10.3390/cells9010033
Chicago/Turabian StyleAl-Hatamleh, Mohammad A. I., Engku Nur Syafirah E.A.R., Jennifer C. Boer, Khalid Ferji, Jean-Luc Six, Xin Chen, Eyad Elkord, Magdalena Plebanski, and Rohimah Mohamud. 2020. "Synergistic Effects of Nanomedicine Targeting TNFR2 and DNA Demethylation Inhibitor—An Opportunity for Cancer Treatment" Cells 9, no. 1: 33. https://doi.org/10.3390/cells9010033