
Effect of a Bacillus subtilis plus Yeast Cell Wall Synbiotic on Salmonella Enteritidis Colonization in Ceca of Layer Pullets

1
Quality Technology International, Inc., Elgin, IL 60123, USA
2
Nevysta Laboratory, Iowa State University Research Park, Ames, IA 50010, USA
*
Author to whom correspondence should be addressed.
Poultry 2024, 3(1), 26-35; https://doi.org/10.3390/poultry3010003
Submission received: 22 November 2023
/
Revised: 12 February 2024
/
Accepted: 22 February 2024
/
Published: 26 February 2024
Abstract
:Salmonella Enteritidis (SE) is a major contamination concern in eggs and risk for Salmonellosis in humans. Strains of Bacillus subtilis and yeast cell wall can be used as substitutes for antibiotic substances in feed against Salmonella in poultry. The objective of this study was to assess the effect of BacPack® (Quality Technology International, Inc., Elgin, IL, USA) Q1+1 (BPQ11), a feed additive combination of a Bacillus subtilis strain and Saccharomyces cerevisiae cell wall, on SE cecal colonization in Lohmann LSL pullets. A control group (CON) and a test group (BPQ) were each randomly assigned 100-day-old chicks. CON was fed a corn–soybean meal-based vegetarian mash diet, and BPQ was fed the control diet supplemented with BPQ11 for the duration of the study. At 8 days of age, chicks were orally challenged with a nalidixic acid-resistant SE strain at a dose of 6.3 × 107 colony forming units (CFUs) per bird. At 7, 11, 15, and 19 days post-challenge (DPC), 25 birds per group were euthanized, and their cecal contents were collected and analyzed for SE. SE counts were 6.88, 7.98, 7.79, and 7.50 in CON and 7.18, 7.31, 6.35, and 6.30 log10 CFU/g in BPQ at 7, 11, 15, and 19 DPC, respectively. SE did not differ between CON and BPQ at 7 DPC; however, BPQ had lower (p < 0.0001) SE at 11 (−0.67), 15 (−1.45), and 19 (−1.20 log10 CFU/g) DPC. Results indicate that synbiotic BPQ11 may be a useful dietary pre-harvest tool for SE management in layer birds.
1. Introduction
Nontyphoid Salmonella serovars are the most common bacterial pathogens, causing disease and death worldwide [1,2]. The United States (U.S.) Center for Disease Control (CDC) estimates Salmonella results in more than 1.3 million people infected, 26,000 hospitalized, and 400 dead every year in the U.S. [3]. Infections resulting from consumption of foods contaminated with Salmonella are the second most reported digestive tract diseases in the EU, and more than 50% of the foodborne outbreak sicknesses reported in the EU in 2018 were due to Salmonella [4]. Salmonella is classified into two species, Salmonella bongori and Salmonella enterica. This bacterial genus is also classified into more than 2500 serovars, most of them identified as Salmonella enterica subspecies enterica, including Enteritidis (SE), which is highly widespread and frequently associated with outbreaks of human gastroenteritis [5,6]. While humans can be exposed to SE through consumption of several foods, SE contaminated eggs are considered a major cause of disease in people globally [7,8].
Connection of SE with egg consumption and foodborne disease steered the U.S. Food and Drug Administration (FDA) to issue a regulation in 2009 that required producers of shell eggs to put into effect procedures to prevent eggs from being contaminated with SE on the farm, while they are in storage, or while they are in transport [9]. Despite diligently applied control measures in egg production, SE and eggs contaminated with SE continue to be an international public health concern [10,11]. Such a concern is further compounded by the potential risk of resistance of Salmonella to antibiotics. It appears that the use of antibiotics as feed additives to improve animal growth and efficiency leads to the development of bacterial strains that are resistant to antibiotics [12]. Al-Zenki et al. reported that most of the SE isolates detected in feed, litter, bird ceca, and eggs from a large broiler farm were resistant to many antibiotics including tetracycline and ampicillin [13]. Resistance to antibiotics in various Salmonella species and serovars has also been reported in cattle beef, swine, fresh produce, and seafood [14]. In a recent global overview, Castro-Vargas and coauthors reported that eggs and egg-laying hens had an SE medium prevalence of 40% [15] and stated that SE was the most frequent serotype. They also indicated that ampicillin and nalidixic acid were the two antibiotics for which the greatest level of Salmonella resistance was found within poultry production [15].
Animal welfare concerns have also introduced significant changes in bird management, genetics, and housing in the US over the last decade. While these changes may have implied positive impacts on bird welfare, they may also increase the risk factors for food safety in both pullet and laying chicken operations [16].
The shift away from the routine use of antibiotics in poultry production has resulted in a greater interest in the development and use of substitute feed supplements [17]. Measures to prevent SE contamination in eggs include biosecurity, Salmonella-free replacement pullets, control of rodents and insects, preventing access to animals other than the commercial bird of interest to poultry barns, maintaining clean and disinfected chicken houses, vaccination of hens against Salmonella, refrigeration of eggs immediately after laying, and use of alternative feed additives to antibiotics [18].
Improved intestinal microbiota, enhanced growth or egg production and efficiency, disease reduction, and inhibition of foodborne pathogens observed in poultry when fed probiotics is most likely a combinatorial effect resulting from short-chain fatty acids and antimicrobials, prevention of physical binding by pathogenic bacteria via competition for adhesion sites on the gastrointestinal tract epithelium, stimulation of the host body’s defenses, and, potentially, other modes of action [19]. Mannan oligosaccharides (MOSs), fructo-oligosaccharides (FOSs), and galacto-oligosaccharides (GOSs) are among the more commonly employed prebiotics in animal and poultry research [17], and other new prebiotic technologies, such as mannabiose (MNB), which are derivatives of MOS, have been shown to be efficacious for use in controlling SE in poultry [20]. Bacillus probiotics [21,22,23] and MOS prebiotics [23,24,25,26,27] have been shown to reduce SE colonization in layer birds. In addition, Bacillus-based probiotics and yeast cell wall-based prebiotics are among the most used feed additives in poultry. Only recently, however, has there been published research, though limited, testing the effects of combinations of Bacillus and MOS on SE colonization in layer birds [23,28,29,30]. In these publications, the synbiotics tested contained different Bacillus probiotics and yeast cell wall prebiotics and used either young pullets or adult hens of either Hy-Line or Dekalb genetics background. Despite years of research focused on substitutes for antibiotics in animal production, more options of feed additives that are effective against Salmonella continue to be of interest to the egg industry. The objective of the study was to assess the effect of BacPack Q1+1 (BPQ11), a combination of a Bacillus subtilis strain and Saccharomyces cerevisiae cell wall, on SE cecal colonization in Lohmann LSL pullets.
2. Materials and Methods
2.1. Experimental Animals
Two-hundred-day-old Lohmann LSL baby chicks were obtained from Hy-line North America, LLC (Goldfield, IA, USA). Chicks received Marek’s disease vaccine at the hatchery and were not exposed to any other viral or bacterial vaccines throughout the study. Pullets were reared in isolation at the Iowa State University AAALAC-accredited animal isolation facility. Pullets were allowed ad libitum access to feed and water throughout the study and were managed as has been prescribed by the Guide for the Care and Use of Agricultural Animals in Research and Teaching [31] in line with the management recommendations for Lohmann LSL pullets [32]. Study protocol (#22-035) was approved by the Institutional Animal Care and Use Committee (IACUC) and the Institutional Biosafety Committee (IBC) of ISU. The behavior and general health condition of the pullets were observed daily.
2.2. Experimental Diets
A corn and soybean meal-based vegetarian mash diet (Table 1) was formulated to meet the standard nutritional specifications for layer pullet chickens [33]. The synbiotic product, BPQ11 (Quality Technology International, Inc., Elgin, IL, USA), was supplemented at 635 g per metric ton of feed. The concentration of the Bacillus subtilis in BPQ11 was 521 × 106 colony forming units (CFUs)/g using a culture count method (ISO 4833-2:2013). The yeast cell wall component contained 32% β-glucans and 24% MOS, measured by UV spectrophotometry using Megazyme (Wicklow, Ireland) enzymatic kit tests, the b-Glucan Assay Kit (K-YBGL), and the D-Mannose/D-Fructose/D-Glucose Assay Kit (K-MANGL) according to the manufacturer’s instructions. Bacteriological analysis was conducted to confirm absence of Salmonella contamination in experimental diets.
2.3. Study Design, Challenge, and Sampling
Day-old chicks were randomly divided into 2 groups fed the control diet or diet supplemented with BPQ11. Each experimental group was reared in a single cage unit measuring 76 cm × 457 cm × 46 cm in a separate but identical isolation room. Cages had wire floor mesh and were raised on stainless steel decks to prevent fecal–oral cycling of the challenge organism.
At 8 days of age, each bird in both groups was orally challenged with 6.3 × 107 CFU of a selected nalidixic acid-resistant strain of SE. Inoculum was prepared fresh on the day of challenge following overnight incubation with shaking (37 °C, 60 rpm). Bacteria concentration was monitored by optical density measurements at 600 nm using GENESYS 50 UV-Vis (Thermo Fisher Scientific, Waltham, MA, USA) and harvested at the end of the log phase of growth.
A total of 25 birds from each group were euthanized by cervical dislocation at 7, 11, 15, and 19 days post-challenge (DPC). Cecal pouches were aseptically collected and transported to the laboratory on ice packs for immediate processing. Contents from all ceca from each treatment and at each cecal sampling period were individually analyzed for SE count and used as replicates in the statistical analysis. Cecal count and prevalence of SE are measurements that address the study’s focus of Salmonella food safety concern. Pullets were housed under Biosafety Level 2 (BSL-2) isolation.
2.4. Environmental Swabs
Chick paper swabs were collected from chick boxes upon chick placement to confirm the absence of SE prior to challenge. Environmental drag swabs were collected from birds’ manure underneath cage units at 7 DPC to confirm occurrence of SE shedding in association with the experimental infection.
Each swab was placed in a sterile sampling bag (Whirl-Pak, Madison, WI, USA), and gloves were changed between samples. Swabs were processed for Salmonella isolation using pre-enrichment in buffered peptone water, enrichment in Tetrathionate Hajna (TTH) broth (Difco Laboratories, Detroit, MI, USA) and plating on Xylose Lysine Tergitol-4 (XLT-4) agar (Difco Laboratories) and Brilliant Green with Novobiocin (BGN) agar (Difco Laboratories). Suspected colonies were further tested in triple sugar iron (TSI) and lysine iron (LI) slants followed by serogrouping using appropriate poly-O and poly-H Salmonella antisera as well as group D-specific antiserum (Mira Vista, Indianapolis, IN, USA).
2.5. Salmonella Enumeration
Enumeration of the challenge SE strain was conducted as described by Girgis et al. [26]. Briefly, the contents of cecal pouches were aseptically squeezed into sterile conical tubes. Sterile saline was added at a ratio of 1:10 weight per volume. Ten-fold serial dilutions were prepared, and standard plate count method was conducted using XLT-4 agar plates containing 25 μg nalidixic acid/mL [34]. Plates were incubated aerobically for 24 h at 37 °C and typical Salmonella colonies were counted. At least 3 randomly selected colonies from countable plate were serologically confirmed to be the SE inoculum strain to validate the accuracy of visual counts.
2.6. Data Analysis
GraphPad Prism software version 9.2.0 (GraphPad Software LLC, San Diego, CA, USA) one-way analysis of variance was used to analyze the data. Tukey’s honest significant difference test was used to determine if there were significant differences among means of experimental groups at p ≤ 0.05. SE count data were log transformed before statistical analysis.
3. Results
Descriptive statistics of SE level in cecal content from CON and BPQ pullets are shown in Table 2.
Cecal SE concentration data were mostly consistent within treatments at each cecal content collection period. Except for the cecal SE data of CON at 7 DPC, which were slightly more variable (STD of 1.63), the remaining data across both treatments and cecal content collection periods had relatively low STD values (0.293 to 0.869).
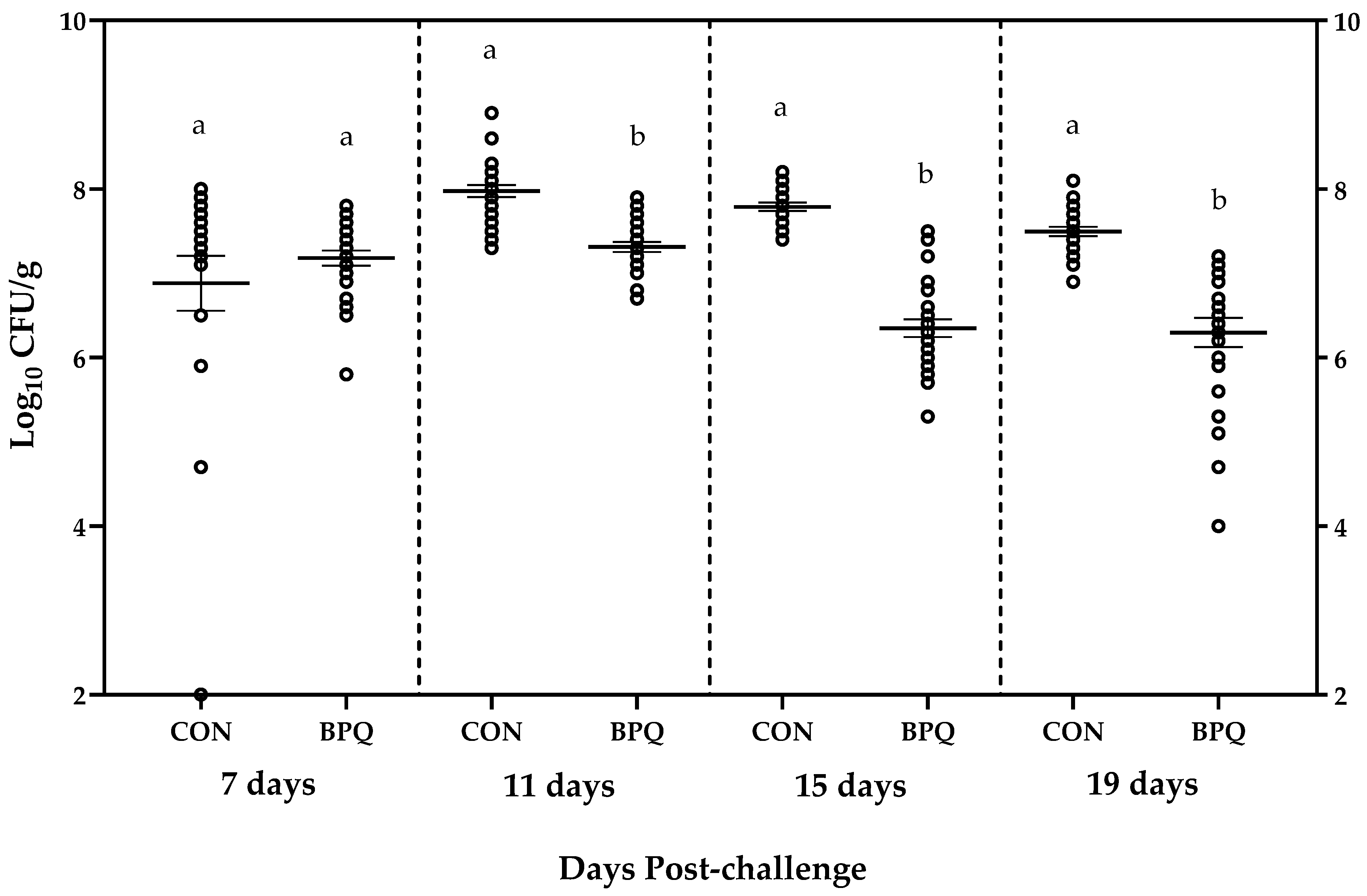
Dot plot of cecal SE level of individual pullets and treatment mean ± SEM for CON and BPQ at each cecal content collection period are presented in Figure 1.
Two birds out of 25 CON pullets at 7 DPC had a cecal SE concentration of 2 log10 CFU/g, which primarily contributed to the higher variability of this data set.
Challenging Lohmann LSL pullets with an SE dose of 6.3 × 107 CFU/bird (7.80 log10 CFU/bird) at 8 days of age resulted in mean SE cecal content concentrations of 6.88, 7.98, 7.79 and 7.50 log10 CFU/g in CON, and 7.18, 7.31, 6.35, and 6.30 log10 CFU/g in BPQ, at 7, 11, 15 and 19 DPC birds, respectively. At 7 DPC, there was no difference (p = 0.379) in SE between CON and BPQ. However, BPQ pullets had a significantly lower (p < 0.0001) SE at 11, 15, and 19 DPC compared to CON. Cecal SE differences between BPQ and CON were 0.67, 1.45, and 1.20 log10 CFU/g at 11, 15, and 19 DPC, respectively.
Ceca from all 200 pullets tested positive for SE, indicating an SE prevalence of 100% regardless of treatment or cecal content collection period.
4. Discussion
In the past decade, significant changes occurred in housing practices and management systems on behalf of animal welfare concerns. Layers once reared exclusively in cage housing with limited access to environmental sources of infection or contamination are now being reared from hatch in free range or cage-free systems. Both free range and cage-free rearing allow birds greater access to infected droppings from other birds or rodents compared to traditional cages. It is generally known that oral and feces represent the main routes of infection and transmission of Salmonella Enteritidis in chickens. In addition to the ileum and cloaca, cecal pouches are the preferred organs for SE colonization and establishment after it is ingested by chickens [35]. After an SE challenge, SE colonized the ceca of young layer pullets [21,26,29,30,36], layer pullets near sexual maturity [24,25,27,28], and adult laying hens [21,22,23]. In this study, orally infecting layer pullets with an SE dose of 6.3 × 107 CFU at 8 days of age resulted in colonization of the ceca, as evidenced by load and prevalence of SE in the ceca.
BPQ, a synbiotic, consists of yeast cell wall MOS, 1,3/1,6 β-glucans (βG), and a selected Bacillus probiotic. Research has demonstrated that βG [37], MOS [23,38], and Bacillus probiotic [21] decrease SE colonization in chicken gut. Proposed modes of action for MOS, βG, and Bacillus probiotics, with relevance to Salmonella colonization, include acting as an alternative binding site for type-1 fimbria [39], upregulation of the immune system [40], and competitive exclusion [41,42], respectively.
Effects on SE colonization in layer birds infected with SE in response to a dietary synbiotic consisting of yeast cell wall and Bacillus probiotic have been reported recently. Compared to the control, SE count in the ceca was reduced by 0.70 CFU/g, on average, across the trial in synbiotic-fed Dekalb White pullets challenged with 2.1 × 109 CFU/bird SE at 8 days of age [29]. Price et al. reported that the number of SE in the ceca at 7 DPC was numerically reduced (p = 0.14) by 0.78 log10 MPN (most probable number)/g compared to the control when 60-week-old Hy-Line W36 fed synbiotic were inoculated with 7 × 107 CFU SE per hen [23]. In a previous study with BPQ11, no significant differences in cecal SE counts at 7, 10, or 13 DPC were observed between the control and BPQ11-fed Hy-Line W36 pullets challenged with an SE dose of 1.6 × 109 log10 CFU per bird at 3 weeks of age [30].
In the current study, dietary BPQ11 significantly (p < 0.0001) reduced cecal SE count in Lohmann LSL pullets challenged with 6.3 × 107 CFU SE per bird at 8 days of age at 11, 15, and 19 DPC but not at 7 DPC. Breed and age of bird, Bacillus strain and yeast cell wall used, synbiotic inclusion rate, SE challenge dose, number of DPC for cecal content collection, and, potentially, other experimental factors may have accounted for the different effects on SE cecal colonization in layer birds observed among the studies referenced above. Work by Shaji and coworkers noted that SE dose, bird age and genetic strain, as well as Salmonella serotype, can be interacting and influencing factors and may explain differences in SE mitigation studies [43]. The various results from the studies discussed above [23,29,30] and those from the study reported here, all of which evaluated synbiotics consisting of a Bacillus probiotic plus yeast cell wall components, suggest that a more standardized Salmonella challenge model in layer pullets to evaluate similar synbiotics would be of interest.
When comparing cecal SE counts of CON and BPQ in the present study, the SE reduction was higher at 15 (1.45 log10 CFU/g) and 19 (1.20 log10 CFU/g) compared to 11 DPC (0.67 log10 CFU/g). Similarly, a higher SE reduction in ceca from Hy-Line W36 pullets fed synbiotic and challenged with SE was observed at 10 and 14 DPC compared to 3 and 6 DPC [29]. Suganuma and coworkers [29] hypothesized that the greater SE reduction observed at the later stages following the SE challenge likely resulted from the combined actions of yeast cell wall and Bacillus probiotic. MOS is known to act as a substitute attachment area for type-1 fimbriae present in certain pathogenic bacteria, such as E. coli and Salmonella, thus preventing binding to the intestinal wall [39]. Such MOS attachment may primarily account for the SE colonization reduction early in post-SE challenge. The magnitude of the SE reduction may later be augmented by the Bacillus probiotic, possibly via its competitive exclusion mode of action. Ricke [44] and others noted that a bird’s microbiome matures with age, and an immature gastrointestinal microflora system is less adept at defending against foodborne pathogen challenges, especially the serovar Salmonella Enteritidis. Having both a prebiotic and probiotic at this early stage of gut microbiome development should allow birds added defenses from incidental pathogen infections in a typical commercial environment. Suganuma and coworkers [29] also considered the increase in the diversity of bacterial species observed at 10 and 14 DPC, but not at 3 and 6 DPC, as supporting evidence for the YCW and Bacillus probiotic combined effect at the later stages of the study. The individual contribution of each constituent of BPQ11 to the reduction of SE colonization in the ceca of pullets was not assessed in this study. However, the higher SE reduction by BPQ11 at the later stage seen in this study could be hypothesized as resulting from an additive or synergistic effect associated with modes of action of MOS [39], βG [40], and Bacillus probiotic [41,42]. More research is needed to elucidate the proportionality and timing of the effects of Bacillus probiotic and yeast cell wall, when combined in a Salmonella challenge model, on Salmonella colonization of the intestinal tract in layer birds.
The prevalence results agree with findings by other researchers who reported lack of a significant effect of the synbiotic used on SE prevalence in 8- or 12-week-old pullets [36], 17-week-old pullets [28], and 60-week-old laying hens [23], respectively. However, in a previous study [30], BPQ11 significantly reduced SE prevalence in ceca by 50% in pullets challenged with SE at 3 weeks of age. In addition to prevalence, Salmonella contamination data also focus on number and serotype [45]. Furthermore, Oscar [45] stipulated that due to the existence of several risk factors associated with Salmonella infection in humans, multiple benchmarks need to be employed to assess food safety of poultry products, not just Salmonella prevalence. SE number in cecal contents in pullets or layers is an accepted and biologically relevant measurement for assessing a feed additive’s ability to reduce SE colonization [21,22,23,24,28,29,30,36]. While SE prevalence was unchanged in the current study, BPQ11 significantly reduced cecal SE load by 0.67, 1.44, and 1.20 log10 CFU/g at 11, 15, and 19 DPC, respectively, after challenging 8-day old pullets with a high dose of SE. As also pointed out by Hofacre and coworkers [24], it is commonly accepted that a 1 log10 change in Salmonella presence within the intestinal tract of poultry is of importance for food safety, especially at near 100% prevalence. In commercial layer operations, 100% flock infection rates would be uncommon and a synbiotic product such as BPQ11 may not only reduce shedding in SE-positive birds but may also be a tool to reduce the likelihood of SE-negative birds from becoming infected, as shown in this lab’s previous work [30]. Preventing Salmonella infection early in the bird’s life is important. Exposure of chickens to SE soon after hatching can lead to infection that lasts through maturity, whereby other birds in the vicinity can become infected or eggs can be contaminated [46]. Evidence has shown that reducing the SE load in the ceca of laying hens will significantly lower the total amount of SE in the barn, ultimately resulting in a lower risk of foodborne disease to humans via a lower level of SE contamination in eggs [47]. Therefore, the outcome of this study shows that the synbiotic BPQ11 can be a useful pre-harvest tool for mitigation of SE infection in laying hens, thereby helping to minimize the threat of Salmonella-caused illness associated with eggs in humans.
Although bird performance was not measured during this disease challenge study, performance and sustainability are also key factors that could be considered in future studies. In previous published work with the probiotic used to manufacture BPQ11, Samper and coauthors [48] noted that laying hens fed diets supplemented with Bacillus subtilis had significantly (p = 0.02) better hen-day egg production (HDEP) compared to hens fed diets not supplemented with Bacillus (86.0% vs. 84.1%, respectively). During the entire study period, from 17 to 70 weeks of age, researchers also noted a 7.1-point improvement (p = 0.06) in the feed conversion ratio (FCR) of hens supplemented with Bacillus in their diets. In the United States, tighter regulations are being implemented by the FDA to limit the use of antibiotics for improving bird productivity and performance and lessen the potential for drug resistant strains of common bacterial pathogens such as Salmonella. Having alternative health products such as BPQ11 that show both mitigating effects to foodborne pathogens, such as Salmonella, and that can improve animal performance and sustainability will be an important tool to the future of the egg industry.
In conclusion, BPQ11, a combination of yeast cell wall and a selected Bacillus probiotic, resulted in a reduced concentration of Salmonella Enteritidis in ceca of Lohmann LSL pullets.
Author Contributions
Conceptualization, M.A. and G.G.; methodology, M.A. and G.G.; investigation, H.M. and G.G.; resources, H.M. and G.G.; data curation, H.M. and G.G.; writing—original draft preparation, M.A.; writing—review and editing, M.A., G.G. and T.L.; project administration, G.G.; funding acquisition, M.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Biosafety Committee and Institutional Animal Care and Use Committee of the Iowa State University system (IACUC #22-035).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data contained within the article.
Acknowledgments
The authors would like to extend appreciation to the Laboratory Animal Resources staff and caretakers at the Iowa State University College of Veterinary Medicine for their daily tasks, care, and oversight for the duration of the project.
Conflicts of Interest
Miloud Araba was employed by Quality Technology International, Inc. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Chen, H.M.; Wang, Y.; Su, L.H.; Chiu, C.H. Nontyphoid Salmonella Infection: Microbiology, Clinical Features, and Antimicrobial Therapy. Pediatr. Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef]
- GBD 2017 Non-Typhoid Salmonella Invasive Disease Collaborators. The global burden of non-typhoidal salmonella invasive disease: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect. Dis. 2019, 19, 1312–1324. [Google Scholar] [CrossRef]
- Center for Disease Control (CDC). Food Safety and Inspection Service; U.S. Department of Agriculture: Honolulu, HI, USA; U.S. Department of Health and Human Services, Centers for Disease Control and Prevention: Atlanta, GA, USA, 2022.
- Ehuma, O.; Jaiswal, A.K.; Jaiswal, S. Salmonella, food safety and food handling practices. Foods 2021, 10, 907. [Google Scholar] [CrossRef]
- Popa, G.L.; Popa, M.I. Salmonella spp. Infection—A continuous threat worldwide. Germs 2021, 11, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Juniora, C.A. Worldwide epidemiology of Salmonella serovars in animal-based foods: A meta-analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [PubMed]
- Pires, S.M.; Vieira, A.R.; Hald, T.; Cole, D. Source attribution of human salmonellosis: An overview of methods and estimates. Foodborne Pathog. Dis. 2014, 11, 667–676. [Google Scholar] [CrossRef]
- Jackson, B.R.; Griffin, P.M.; Cole, D.; Walsh, K.A.; Chai, S.J. Outbreak-associated Salmonella enterica serotypes and food commodities, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 1239–1244. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Prevention of Salmonella Enteritidis in Shell Eggs During Production, Storage, and Transportation; Final Rule. Fed. Regist. 2009, 74, 33030–33100. [Google Scholar]
- EFSA Panel on Biological Hazards, European Commission. Salmonella control in poultry flocks and its public health impact. EFSA J. 2019, 17, 5596. [Google Scholar] [CrossRef]
- Gast, R.K.; Regmi, P.; Guraya, R.; Jones, D.R.; Anderson, K.E.; Karcher, D.M. Contamination of eggs by Salmonella enteritidis in experimentally infected laying hens of four commercial genetic lines in conventional cages and enriched colony housing. Poult. Sci. 2019, 98, 5023–5027. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K. Use of antibiotics as feed additives: A burning question. Front. Microbiol. 2014, 5, 334. [Google Scholar] [CrossRef]
- Al-Zenki, S.; Al-Nasser, A.; Al-Safar, A.; Alomirah, H.; Al-Haddad, A.; Hendriksen, R.S.; Aarestrup, F.M. Prevalence and antibiotic resistance of Salmonella isolated from a poultry farm and processing plant environment in the state of Kuwait. Foodborne Pathog. Dis. 2007, 4, 367–373. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Venkitanarayanan, K.; Johny, A.K. Antibiotic-Resistant Salmonella in the Food Supply and the Potential Role of Antibiotic Alternatives for Control. Foods 2018, 7, 167. [Google Scholar] [CrossRef]
- Castro-Vargas, R.E.; Herrera-Sanchez, M.P.; Rodriguez-Hernandez, R.; Rondon-Barragan, I.S. Antibiotic resistance in Salmonella spp. Isolated from poultry: A global overview. Vet. World 2020, 13, 2070–2084. [Google Scholar] [CrossRef]
- Sharma, M.; McDaniel, C.; Kiess, A.; Loar, R.; Adhikari, P. Effect of housing environment and hen strain on egg production and egg quality as well as cloacal and eggshell microbiology in laying hens. Poult. Sci. 2022, 101, 101595. [Google Scholar] [CrossRef] [PubMed]
- Ricke, S. Impact of prebiotics on poultry production and food safety. Yale J. Biol. Med. 2018, 91, 151–159. [Google Scholar] [PubMed]
- Trampel, D.W.; Holder, T.D.; Gast, R.K. Integrated farm management to prevent Salmonella Enteritidis contamination in eggs. J. Appl. Poult. Res. 2014, 23, 353–365. [Google Scholar] [CrossRef]
- El Jeni, R.; Dittoe, D.K.; Olson, E.G.; Lourenco, J.; Corcionivoschi, N.; Ricke, S.C.; Callaway, T.R. Probiotics, and potential applications for alternative poultry production systems. Poult. Sc. 2021, 100, 101156. [Google Scholar] [CrossRef] [PubMed]
- Morikoshi, K.; Yokomizo, F. Method of Inhibiting Salmonella in Livestock and Poultry. U.S. Patent 8,999,374B2, 7 April 2015. [Google Scholar]
- Price, P.T.; Gaydos, T.A.; Berghaus, R.D.; Baxter, V.; Hofacre, C.L.; Sims, M.D. Salmonella Enteritidis reduction in layer ceca with a Bacillus probiotic. Vet. World 2020, 13, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Padgett, J.C.; Price, P.T.; Byrd, J.A.; Bailey, C.A. Salmonella Enteritidis control in mature laying hens through dry fed parietal yeast fraction or Bacillus blend probiotic. Int. J. Anim. Sci. Technol. 2021, 5, 1–6. [Google Scholar] [CrossRef]
- Price, P.T.; Gaydos, T.; Legendre, H.; Krehling, J.; Macklin, K.; Padgett, J.C. Production Layer Salmonella Enteritidis Control through Dry Fed Pre & Probiotic Products. Braz. J. Poult. Sci. 2021, 23, 1–6. [Google Scholar]
- Hofacre, C.L.; Berghaus, R.D.; Jalukar, S.; Mathis, G.F.; Smith, J.A. Effect of a yeast cell wall preparation on cecal and ovarian colonization with Salmonella Enteritidis in commercial layers. J. Appl. Poult. Res. 2018, 27, 453–460. [Google Scholar] [CrossRef]
- Girgis, G.; Powell, M.; Youssef, M.; Graugnard, D.E.; King, W.D.; Dawson, K.A. Effects of a mannan-rich yeast cell wall-derived preparation on cecal concentrations and tissue prevalence of Salmonella Enteritidis in layer chickens. PLoS ONE 2020, 15, e0232088. [Google Scholar] [CrossRef] [PubMed]
- Gingerich, E.; Frana, T.; Logue, C.M.; Smith, D.P.; Pavlidis, H.O.; Chaney, W.E. Effect of feeding a postbiotic derived from Saccharomyces cerevisiae fermentation as a preharvest food safety hurdle for reducing Salmonella Enteritidis in the ceca of layer pullets. J. Food Prot. 2021, 84, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Chaney, W.E.; McBride, H.; Girgis, G. Effect of a Saccharomyces cerevisiae Postbiotic Feed Additive on Salmonella Enteritidis Colonization of Cecal and Ovarian Tissues in Directly Challenged and Horizontally Exposed Layer Pullets. Animals 2023, 13, 1186. [Google Scholar] [CrossRef] [PubMed]
- Kimminau, E.A.; Karnezos, T.P.; Berghaus, R.D.; Jones, M.K.; Baxter, J.A.; Hofacre, C.L. Combination of probiotic and prebiotic impacts Salmonella Enteritidis infection in layer hens. J. Appl. Poult. Res. 2021, 30, 100200. [Google Scholar] [CrossRef]
- Suganuma, K.; Hamasaki, T.; Hamaoka, T. Effect of dietary direct-fed microbial and yeast cell walls on cecal digesta microbiota of layer chicks inoculated with nalidixic acid resistant Salmonella Enteritidis. Poult. Sci. 2021, 100, 101385. [Google Scholar] [CrossRef] [PubMed]
- Girgis, G.; McBride, H.; Boyle, B.; Araba, M.; Bodle, B.C.; Lohrmann, T. Effects of a synbiotic combination of Bacillus subtilis and yeast cell wall-derived glucomannan on cecal colonization of Salmonella Enteritidis in layer chickens. J. Appl. Poult. Res. 2022, 31, 100240. [Google Scholar] [CrossRef]
- Guide for the Care and Use of Agricultural Animals in Research and Teaching. 4th ed Federation of Animal Science Societies. 2020. Available online: https://poultryscience.org/files/galleries/AG_Guide_4th_Ed_2020.pdf (accessed on 10 April 2023).
- Lohmann LSL Lite Management Guide, North America Edition. Available online: https://hylinena.com/wp-content/uploads/2019/10/Lohmann_LSL-Lite16-2.pdf (accessed on 10 April 2023).
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th ed.; The National Academies Press: Washington, DC, USA, 1994.
- Association of Official Analytical Chemists. Official Methods of Analysis; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Turnbull, P.C.; Snoeyenbos, G.H. Experimental Salmonellosis in the chicken. 1. Fate and host response in alimentary canal, liver, and spleen. Avian Dis. 1974, 18, 153–177. [Google Scholar] [CrossRef] [PubMed]
- Krueger, L.A.; Gaydos, T.A.; Sims, M.D.; Spangler, D.A. Avi-Lution supplemented at 250 or 500 mg per kg in feed decreases the abundance of Salmonella Enteritidis in ceca of layer pullets. J. Appl. Poult. Res. 2020, 29, 995–1003. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, Z.; Tian, X.; Guo, Y.; Zhang, H. Yeast β-d-glucans induced antimicrobial peptide expressions against Salmonella infection in broiler chickens. Int. J. Biol. Macromol. 2016, 85, 573–584. [Google Scholar] [CrossRef]
- Chaney, W.E.; Naqvi, S.A.; Gutierrez, M.; Gernat, A.; Johnson, T.J.; Petry, D. Dietary Inclusion of a Saccharomyces cerevisiae-Derived Postbiotic Is Associated with Lower Salmonella enterica Burden in Broiler Chickens on a Commercial Farm in Honduras. Microorganisms 2022, 10, 544. [Google Scholar] [CrossRef]
- Spring, P.; Wenk, C.; Dawson, K.A.; Newman, K.E. The Effects of Dietary Mannanoligosaccharides on cecal parameters and the concentrations of enteric bacteria in the ceca of Salmonella-challenged broiler chicks. Poult. Sci. 2000, 79, 205–211. [Google Scholar] [CrossRef]
- Lowry, V.K.; Farnell, M.B.; Ferro, P.J.; Swaggerty, C.L.; Bahl, A.; Kogut, M.H. Purified beta-glucan as an abiotic feed additive up-regulates the innate immune response in immature chickens against Salmonella enterica serovar Enteritidis. Int. J. Food Microbiol. 2005, 98, 309–318. [Google Scholar] [CrossRef]
- La Ragione, R.M.; Woodward, M.J. Competitive exclusion by Bacillus subtilis of Salmonella enterica serotype Enteritidis and Clostridium perfringens in young chickens. Vet. Microbiol. 2003, 94, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T.; Ashida, N.; Nakagawa, K.; Iwatani, S.; Yamamoto, N. Dietary Bacillus subtilis C-3102 Supplementation Enhances the Exclusion of Salmonella enterica from Chickens. J. Poult. Sci. 2021, 58, 138–145. [Google Scholar] [CrossRef]
- Shaji, S.; Selvaraj, K.; Shanmugasundaram, R. Salmonella Infection in Poultry: A Review on the Pathogen and Control Strategies. Microorganisms 2023, 11, 2814. [Google Scholar] [CrossRef]
- Ricke, S.; Dittoe, D.; Olson, E. Microbiome applications for laying hen performance and egg production. Poult. Sci. 2022, 101, 101784. [Google Scholar] [CrossRef] [PubMed]
- Oscar, T. Salmonella prevalence alone is not a good indicator of poultry food safety. Risk Anal. 2021, 41, 110–130. [Google Scholar] [CrossRef] [PubMed]
- Gast, R.K.; Holt, P.S. Persistence of Salmonella Enteritidis from one day of age until maturity in experimentally infected layer chickens. Poult. Sci. 1998, 77, 1759–1762. [Google Scholar] [CrossRef]
- Price, P.T.; Gaydos, T.; Berghaus, R.; Hofacre, C. Reduction of Salmonella Enteritidis colonization in production layers fed high levels of mannan and beta-glucan. Asian J. Poult. Sci. 2020, 14, 1–5. [Google Scholar] [CrossRef]
- Samper, J.; Araba, M.; Ruano, M.; Lohrmann, T.; Persia, M. Effects of a Direct Fed Microbial Fed from 0 to 70 Weeks of Age on Laying Hen Performance and Egg Quality from 18–70 Weeks of Age. J. Poult. Sci. 2022, 101 (Suppl. S1), 64. [Google Scholar]
Figure 1.
Dot plot of Salmonella Enteritidis (SE) concentration (log10 CFU/g) in cecal contents of pullets collected at 7-, 11-, 15- and 19- days post challenge with SE dose of 6.3 × 107 CFU/bird at 8 days of age. Lines within the plot depict means ± SEM. a,b Means within each cecal content collection period with different letters are significantly different (p < 0.05). CON = pullets fed non-supplemented diet; BPQ = pullets fed diet with added BacPack Q1+1 (BPQ11) at a rate of 635 g per metric ton of feed.
Figure 1.
Dot plot of Salmonella Enteritidis (SE) concentration (log10 CFU/g) in cecal contents of pullets collected at 7-, 11-, 15- and 19- days post challenge with SE dose of 6.3 × 107 CFU/bird at 8 days of age. Lines within the plot depict means ± SEM. a,b Means within each cecal content collection period with different letters are significantly different (p < 0.05). CON = pullets fed non-supplemented diet; BPQ = pullets fed diet with added BacPack Q1+1 (BPQ11) at a rate of 635 g per metric ton of feed.


{kind=link}
Table 1.
Formula of basal diet formula fed to pullets in mash form and ad libitum throughout the length of study. Control pullets were fed the basal diet, and the treatment pullets were fed the basal diet supplemented with BackPack Q1+1 (BPQ11) at a rate of 635 g per metric ton of feed.
Table 1.
Formula of basal diet formula fed to pullets in mash form and ad libitum throughout the length of study. Control pullets were fed the basal diet, and the treatment pullets were fed the basal diet supplemented with BackPack Q1+1 (BPQ11) at a rate of 635 g per metric ton of feed.
| Ingredient | Diet, as-Fed Basis |
|---|---|
| Corn, % | 56.9 |
| Soybean meal, % | 33.5 |
| Calcium carbonate, % | 1.95 |
| Monocalcium phosphate (21% p), % | 1.79 |
| Vegetable oil, % | 3.91 |
| Salt, % | 0.50 |
| Choline chloride, % | 0.06 |
| L-lysine, % | 0.68 |
| DL methionine, % | 0.47 |
| Vitamin and mineral premix, % | 0.10 |
| Nutrient | |
| Metabolizable energy, Kcal/Kg | 3054 |
| Crude protein, % | 22.0 |
| Crude fat, % | 5.47 |
| Crude fiber, % | 2.57 |
| Calcium, % | 1.19 |
| Available phosphorus, % | 0.51 |
| Lysine, % | 1.54 |
| Methionine, % | 0.78 |
| Methionine + cysteine, % | 1.16 |
| Sodium, % | 0.25 |
Table 2.
Descriptive statistics for Salmonella Enteritidis (SE) concentration (log10 CFU/g) in cecal contents from pullets at 7, 11, 15, and 19 days post-challenge with SE dose of 6.3 × 107 CFU/bird at 8 days of age. Cecal content was collected from 25 pullets from CON or PBP at each of the 4 collection periods and analyzed for SE level. CON = pullets fed non-supplemented diet; PBQ = pullets fed diet with added BacPack Q1+1 (BPQ11) at a rate of 635 g per metric ton of feed.
Table 2.
Descriptive statistics for Salmonella Enteritidis (SE) concentration (log10 CFU/g) in cecal contents from pullets at 7, 11, 15, and 19 days post-challenge with SE dose of 6.3 × 107 CFU/bird at 8 days of age. Cecal content was collected from 25 pullets from CON or PBP at each of the 4 collection periods and analyzed for SE level. CON = pullets fed non-supplemented diet; PBQ = pullets fed diet with added BacPack Q1+1 (BPQ11) at a rate of 635 g per metric ton of feed.
| Days Post-Challenge | 7 | 11 | 15 | 19 | ||||
|---|---|---|---|---|---|---|---|---|
| Treatment | CON | BPQ | CON | BPQ | CON | BPQ | CON | BPQ |
| Number of pullets | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 |
| Minimum (log10 CFU/g) | 2.0 | 5.8 | 7.3 | 6.7 | 7.4 | 5.3 | 6.9 | 4.0 |
| Maximum (log10 CFU/g) | 8.0 | 7.8 | 8.9 | 7.9 | 8.2 | 7.5 | 8.1 | 7.2 |
| Mean (log10 CFU/g) | 6.88 | 7.18 | 7.98 | 7.31 | 7.79 | 6.35 | 7.50 | 6.30 |
| STD (log10 CFU/g) | 1.63 | 0.45 | 0.363 | 0.293 | 0.246 | 0.521 | 0.276 | 0.869 |
| SEM (log10 CFU/g) | 0.326 | 0.090 | 0.073 | 0.059 | 0.049 | 0.104 | 0.055 | 0.174 |
| BPQ mean—CON mean (log10 CFU/g) | +0.30 | −0.67 | −1.45 | −1.20 | ||||
| p-value | 0.379 | <0.0001 | <0.0001 | <0.0001 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Araba, M.; Girgis, G.; McBride, H.; Lohrmann, T. Effect of a Bacillus subtilis plus Yeast Cell Wall Synbiotic on Salmonella Enteritidis Colonization in Ceca of Layer Pullets. Poultry 2024, 3, 26-35. https://doi.org/10.3390/poultry3010003
AMA Style
Araba M, Girgis G, McBride H, Lohrmann T. Effect of a Bacillus subtilis plus Yeast Cell Wall Synbiotic on Salmonella Enteritidis Colonization in Ceca of Layer Pullets. Poultry. 2024; 3(1):26-35. https://doi.org/10.3390/poultry3010003
Chicago/Turabian StyleAraba, Miloud, George Girgis, Hannah McBride, and Troy Lohrmann. 2024. "Effect of a Bacillus subtilis plus Yeast Cell Wall Synbiotic on Salmonella Enteritidis Colonization in Ceca of Layer Pullets" Poultry 3, no. 1: 26-35. https://doi.org/10.3390/poultry3010003