
A Systematic In Silico Investigation of Phytochemicals from Artocarpus Species against Plasmodium falciparum Inhibitors †


Department of Pharmaceutical Sciences and Technology, Birla Institute of Technology, Mesra 835215, India
*
Author to whom correspondence should be addressed.
†
Presented at the 2nd International Electronic Conference on Antibiotics—Drugs for Superbugs: Antibiotic Discovery, Modes of Action and Mechanisms of Resistance, 15–30 June 2022; Available online: https://eca2022.sciforum.net/ .
Med. Sci. Forum 2022, 12(1), 5; https://doi.org/10.3390/eca2022-12712
Published: 15 June 2022
(This article belongs to the Proceedings of The 2nd International Electronic Conference on Antibiotics—Drugs for Superbugs: Antibiotic Discovery, Modes of Action and Mechanisms of Resistance)
Abstract
:Artemisinin-resistant plasmodium strains are becoming increasingly common in malaria patients, posing a serious threat to successful malaria management. Brosimone, a significant polyphenolic ingredient of Artocarpus lakoocha, has previously been shown to have antimalarial activity in vitro. However, research into the precise mechanism of interactions is still in progress. The present study explored molecular modeling research in order to elucidate the likely mechanism of its anti-malarial effect in terms of Falcipain-2 (FP-2) inhibition. Brosimone shows a maximum binding affinity (docking score: −8.1 Kcal/mol) against FP-2 from Plasmodium falciparum, according to our molecular docking analysis of 50 lakoocha bioactive chemicals. For numerous Artocarpus lakoocha polyphenols (ALP), we used in-silico pharmacokinetics and toxicities, leading to critical insights into the mechanism of action of brosimone and other ALPs as potential therapeutic agents (2GHU) against malaria.
1. Introduction
Parasitic diseases are one of the most deadly and widespread infections on the planet, resulting in millions of morbidities and deaths each year [1]. Plasmodium falciparum is the most common vector-borne infectious disease, killing 0.5 million people each year. The single-cell eukaryote has a complicated life cycle and is an obligatory intracellular parasite of hepatocytes and erythrocytes, and infection can lead to a variety of complications, including severe anemia and cerebral malaria. This can result in death; Plasmodium falciparum replicates inside erythrocytes many times during the course of 48 h, resulting in rapid disease progression and exponential expansion. No other pathogen has put a greater selection pressure on the human genome than the most common infectious illness affecting children. There is no effective vaccination; however, there are various curative therapies. Plasmodium falciparum is the most dangerous form of malaria, infecting humans and causing the majority of malaria-related deaths. Despite the fact that the worldwide malaria epidemic has subsided, the World Health Organization predicts that 212 million cases of malaria and 429,000 deaths occurred in 2020 [2,3]. Malaria medications are desperately needed, and traditional malaria treatment procedures could be a viable source of novel antimalarial chemicals [4]. FP-2 and falcipain-3 (FP3) are important papain-family (C1) clan CA trophozoite cysteine proteases that cleave host hemoglobin (native and denatured) and cause erythrocyte rupture in the digestive food vacuole [5,6,7]. FP-2 is the primary haemoglobinase of Plasmodium falciparum, the human malaria parasite. It is currently acquiring clinical significance as a therapeutic target of choice in the fight against malaria [8]. Plasmodium cysteine protease is involved in a number of biological processes, including membrane rupture. Hemoglobin (Hb) is a protein found in red blood cells that distributes oxygen to the human body’s organs and tissues while also transporting carbon dioxide back to the lungs. Hb breakdown, protein trafficking, and host cell invasion are common causes. [9,10]. As a result, the interference of crucial indicator protein FP2 is used to monitor the proliferation of malarial parasites; it is an appealing target for antimalarial drug development [11]. In this research, the well-known “PyRx” docking simulation was employed to conduct an experiment on malaria disease. The present study docked 50 different anti-malaria polyphenols with FP-2 (2GHU.pdb) from the protein data bank in order to find the most effective and efficient one. With a value of −8.1 Kcal/mol, brosimone was discovered to have the lowest binding energy. As a result, it is a viable candidate for antimalarial medication. Based on our research, polyphenols from Artocarpus (Figure 1) [12] have a strong pharmacological potential against a variety of biological targets.
2. Materials and Methods
2.1. Molecular Docking Simulations
For this investigation, a set of 50 ALP is well known. Overall, there are five key processes in molecular docking: protein preparation, ligand preparation, receptor grid construction, ligand docking procedure, and docking results viewing. ‘ChemDraw’ was used to draw all of the necessary structures. The 3D crystal structures of Plasmodium falciparum (PDB ID: 2GHU) were downloaded from the protein data bank (PDB database, https://www.rcsb.org (accessed on 14 January 2022). The binding pocket of 2GHU target enzymes was docked with co-crystallized ligand. The RMSD value was used to assess the reliability of the docking process. The grid parameter that was set up was Center X:47.6593 Y:11.7104 Z:−12.9581 and Dimensions (Angstrom) X:250,000, Y:250,000, Z:250,000. The grid was centered on active binding site residues such as His D:19, Glu:222, ALA:157, TRP:206, CYS:39, etc. ‘PyRx’ was used to execute the docking study, after which Discovery Studio 2020 Visualizer was used to visualize the results [12,13,14].
2.2. In-Silico Drug-Likeness and ADMET (Absorption, Distribution, Metabolism, Excretion, Toxicity) Analysis
A total of 50 ALP were subjected to an in-silico ADME analysis using SWISS tools http://www.swissadme.ch (accessed on 14 January 2022). For the toxicity assessments, we used ‘SMILES’ formats of chemical structures.
3. Result and Discussion
3.1. Molecular Docking Simulations
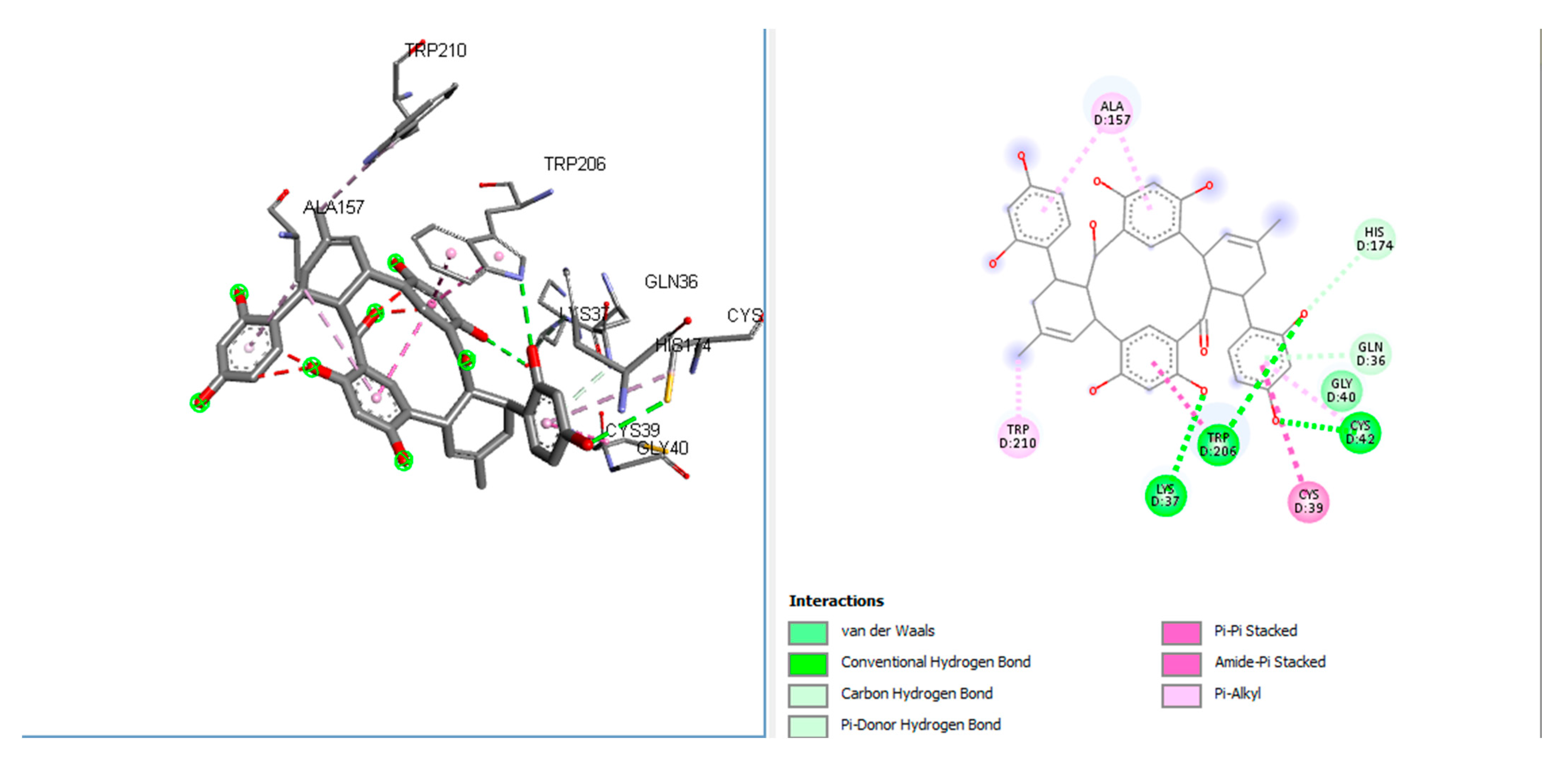
FP-2 is a promising target for malaria treatment. Despite the fact that various FP-2 inhibitors have been clinically proven for the treatment of malaria in recent years, the quest for new active molecules against FP-2 remains difficult. Our research aims to filter several compounds using molecular docking and virtual screening to find novel anti-FP-2 inhibitors. The molecular docking analysis of 50 ALP on an anti-malarial target suggested that bromosine had a high affinity towards 2GHU. The docking interactions depicted that this compound had interactions with GLY A:40, HIS A:174, CYS A: 42, ALA A:157, LYS D:37 A:85, GLN D:36, TRP A: 206 amino acids with three conventional hydrogen bonds at the receptor site of the target 2GHU (Figure 2) [15]. HIS B:174 formed a carbon-hydrogen bond. Van der Waals interactions were observed for GLY D:40. A Pi-donor hydrogen bond was observed for GLN D:36. Pi-alkyl interactions were also observed for TRP D:210 and ALA D:157. Pi-pi amide-pi-pi stacking was observed for CYS D:39. Table 1 and Table 2 give better insights into the interaction profiles of the studied ALP from the docking analysis of 50 bioactive on 2GHU. Furthermore, the top three best docked hits are rutin (docking score: −7.4kcal/mol), lakoochanone (docking score −8 kcal/mol) and brosimone (docking score −8.1 kcal/mol). To determine the most effective and efficient compound, 50 distinct polyphenols were docked with FP-2 (2GHU) from the protein data bank. With a binding energy of −8.1 Kcal/mol, brosimone was determined to have the lowest binding energy. As a result, it is a viable candidate for anti-malaria treatment.
3.2. In-Silico ADMET Studies
Cytochrome P450 (CYPs) enzymes are essential enzymes involved in a variety of metabolic processes. Table 3 shows the in-silico computed ADMET (absorption, distribution, metabolism, excretion, and toxicity) for the body attributes of the top three best-docked hits. Rutin, lakoochanone, and brosimone, three ALPs, had positive human intestinal absorption profiles, a negative blood-brain barrier passage, were non-carcinogenic, non-AMES hazardous, and had class IV acute oral toxicity profiles.
4. Conclusions
Brosimone has strong interactions with the 2GHU enzyme, according to current research (docking score: −8.1 kcal/mol) (amino acid residues: TRP D:210, LYS D:37, TRP D:206, CYS D:39, CYS D:42, GLY D:40, GLN D:40, HIS D:174, ALA D:157). AMES Toxicity and Carcinogens are not present. Molecular docking analysis was used to explore brosimone’s antimalarial properties against the 2GHU enzyme, based on previous literature reports. Given the benefits of ALP, this research could lead to the development of more effective anti-malarial compounds. Furthermore, brosimone might be evaluated in vitro for antimalarial properties. For numerous ALPs, in-silico pharmacokinetics and toxicities were performed, leading to critical insights into the mechanism of action of brosimone and other ALPs as potential therapeutic agents (2GHU) against malaria.
Author Contributions
All authors have contributed equally. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Will be made available upon reasonable request.
Acknowledgments
The authors would like to thank the Head of the Department of Pharmaceutical Sciences and Technology, BIT, Mesra for providing the research facilities for performing the current study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Momčilović, S.; Cantacessi, C.; Arsić-Arsenijević, V.; Otranto, D.; Tasić-Otašević, S. Rapid diagnosis of parasitic diseases: Current scenario and future needs. Clin. Microbiol. Infect. 2019, 25, 290–309. [Google Scholar] [CrossRef] [PubMed]
- WHO Malaria Report. Available online: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2021 (accessed on 14 January 2022).
- Bouchut, A.; Rotili, D.; Pierrot, C.; Valente, S.; Lafitte, S.; Schultz, J.; Hoglund, U.; Mazzone, R.; Lucidi, A.; Fabrizi, G.; et al. Identification of novel quinazoline derivatives as potent antiplasmodial agents. Eur. J. Med. Chem. 2019, 161, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Benoit-Vical, F. Ethnomedicine in malaria treatment. IDrugs 2005, 8, 45–52. [Google Scholar] [PubMed]
- Shenai, B.R.; Sijwali, P.S.; Singh, A.; Rosenthal, P.J. Characterization of Native and Recombinant Falcipain-2, a Principal Trophozoite Cysteine Protease and Essential Hemoglobinase of plasmodium falciparum. J. Biol. Chem. 2000, 275, 29000–29010. [Google Scholar] [CrossRef] [PubMed]
- Sijwali, P.S.; Shenai, B.R.; Gut, J.; Singh, A.; Rosenthal, P.J. Expression and characterization of the Plasmodium falciparum haemoglobinase falcipain-3. Biol. Chem. J. 2001, 360, 481–489. [Google Scholar] [CrossRef]
- Hogg, T.; Nagarajan, K.; Herzberg, S.; Chen, L.; Shen, X.; Jiang, H.; Wecke, M.; Blohmke, C.; Hilgenfeld, R.; Schmidt, C.L. Structural and functional characterization of Falcipain-2, a hemoglobinase from the malarial parasite Plasmodium falciparum. J. Biol. Chem. 2006, 281, 25425–25437. [Google Scholar] [CrossRef] [PubMed]
- Rajguru, T.; Bora, D.; Modi, M.K. Identification of promising inhibitors for Plasmodium haemoglobinase Falcipain-2, using virtual screening, molecular docking, and MD Simulation. J. Mol. Struct. 2022, 1248, 131427. [Google Scholar] [CrossRef]
- Lehmann, C.; Heitmann, A.; Mishra, S.; Burda, P.C.; Singer, M.; Prado, M.; Niklaus, L.; Lacroix, C.; Ménard, R.; Frischknecht, F.; et al. A cysteine protease inhibitor of plasmodium berghei is essential for exo-erythrocytic development. PLoS Pathog. 2014, 10, e1004336. [Google Scholar] [CrossRef] [PubMed]
- Melo, P.M.; Maluf, S.E.C.; Azevedo, M.F.; Paschoalin, T.; Budu, A.; Bagnaresi, P.; Henrique-Silva, F.; Soares-Costa, A.; Gazarini, M.L.; Carmona, A.K. Inhibition of Plasmodium falciparum cysteine proteases by the sugarcane cystatin CaneCPI-4. Parasitol. Int. 2018, 67, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Fagnidi, Y.K.H.; Toi, B.; Megnassan, E.; Frecer, V.; Miertuš, S. In silico design of Plasmodium falciparum cysteine protease falcipain 2 inhibitors with favorable pharmacokinetic profile. J. Anal. Pharm. Res. 2018, 7, 298–309. [Google Scholar]
- Mali, S.N.; Pandey, A. Balanced QSAR and molecular modeling to identify structural requirements of imidazopyridine analogues as anti-infective agents against trypanosomiases. J. Comput. Biophys. Chem. 2022, 21, 83–114. [Google Scholar] [CrossRef]
- Mali, S.N.; Pandey, A. Molecular modeling studies on 2, 4-disubstituted imidazopyridines as anti-malarials: Atom-based 3D-QSAR, molecular docking, virtual screening, in-silico ADMET and theoretical analysis. J. Comput. Biophys. Chem. 2021, 20, 267–282. [Google Scholar] [CrossRef]
- Mali, S.N.; Pandey, A.; Thorat, B.R.; Lai, C.H. Multiple 3D-and 2D-quantitative structure–activity relationship models (QSAR), theoretical study and molecular modeling to identify structural requirements of imidazopyridine analogues as anti-infective agents against tuberculosis. Struct. Chem. 2022, 33, 679–694. [Google Scholar] [CrossRef]
- Mali, S.N.; Pandey, A. Unveiling Naturally Occurring Green Tea Polyphenol Epigallocatechin-3-Gallate (EGCG) Targeting Mycobacterium DPRE1 for Anti-Tb Drug Discovery. Eng. Proc. 2021, 11, 31. [Google Scholar]
Figure 1.
Chemical structure of Artocarpus lakoocha polyphenols.
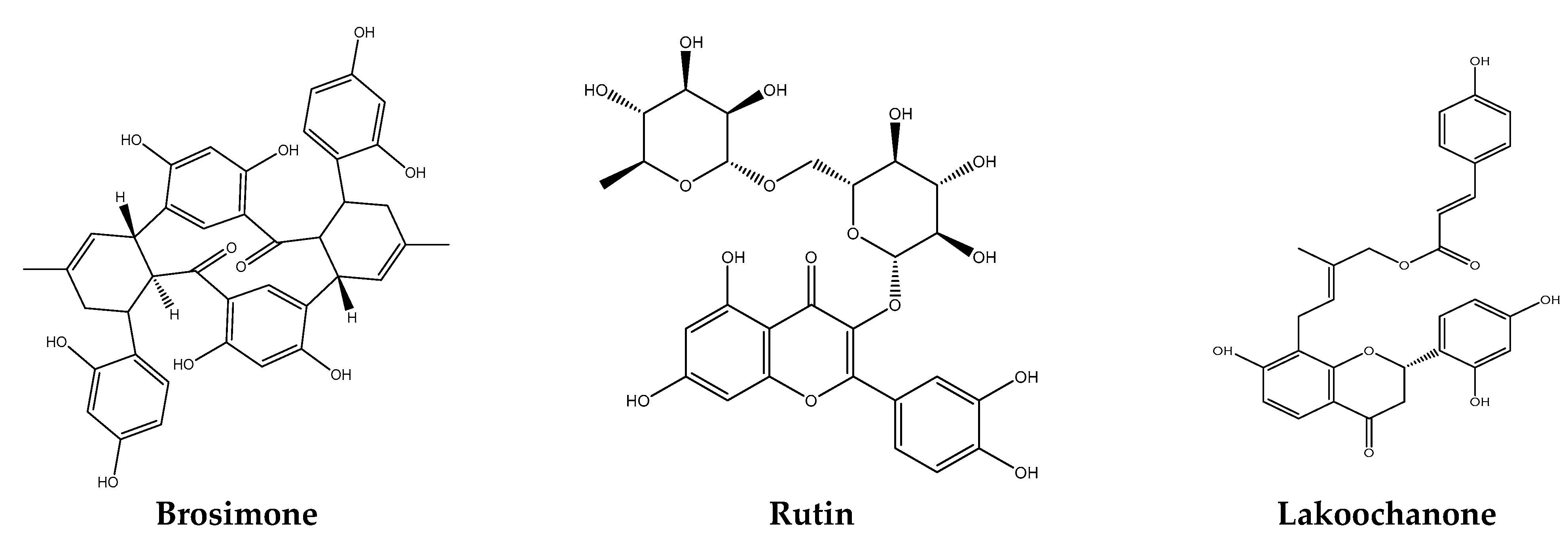
Figure 2.
2D and 3D interaction profiles for best-docked brosimone with 2GHU.

{kind=link}
{kind=link}
Table 1.
Docking interaction energies* of 50 selected bio-active molecules for target protein 2GHU.
| Molecule | PyRx Interaction Energy |
|---|---|
| 1. Brosimone | −8.1 * |
| 2. Lakoochanone | −8 |
| 3. Rutin | −7.4 |
* Docking scores have only been provided for the higher-affinity scored target protein.
Table 2.
The energy contribution of the key residues is computed by docking methodology.
| Sr. No. | Molecules | Residues with Contribution Energy |
|---|---|---|
| 1. | Rutin | HIS D:19, GLU D:222, ALA A:157, TRP A:210, GLN A:209, LYS A:37, ASP A:35, CYS A:39 |
| 2. | Lakoochanone | GLY A:166, ASP A:155, LYS D:135, PHE A:164 |
| 3. | Brosimone | TRP D:210, LYS D:37, TRP D:206, CYS D:39, CYS D:42, GLY D:40, GLN D:40, HIS D:174, ALA D:157 |
Table 3.
In-silico ADMET profiling for top 3 best-docked hits against target 2GHU.
| GI Absorption | BBB Permeant | P-gp Substrate | CYP1A2 Inhibitor | CYP2C19 Inhibitor | CYP2C9 Inhibitor | CYP2D6 Inhibitor | Lipinski | Bioavailability Score | |
|---|---|---|---|---|---|---|---|---|---|
| 1. Lakoochanone | Low | No | No | No | No | Yes | No | Yes | 0.55 |
| 2. Rutin | Low | No | Yes | No | No | No | No | No | 0.17 |
| 3. Brosimone | Low | No | Yes | No | No | No | No | No | 0.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chaurasia, S.; Pandey, A. A Systematic In Silico Investigation of Phytochemicals from Artocarpus Species against Plasmodium falciparum Inhibitors. Med. Sci. Forum 2022, 12, 5. https://doi.org/10.3390/eca2022-12712
AMA Style
Chaurasia S, Pandey A. A Systematic In Silico Investigation of Phytochemicals from Artocarpus Species against Plasmodium falciparum Inhibitors. Medical Sciences Forum. 2022; 12(1):5. https://doi.org/10.3390/eca2022-12712
Chicago/Turabian StyleChaurasia, Surabhi, and Anima Pandey. 2022. "A Systematic In Silico Investigation of Phytochemicals from Artocarpus Species against Plasmodium falciparum Inhibitors" Medical Sciences Forum 12, no. 1: 5. https://doi.org/10.3390/eca2022-12712