
Structure, Stability and Binding Properties of Collagen-Binding Domains from Streptococcus mutans
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification
2.2. Crystallization and Structure Determination
2.3. Collagen-Binding Assay
2.4. Thermal Denaturation
3. Results and Discussion
3.1. Structure of Cnm/CBD
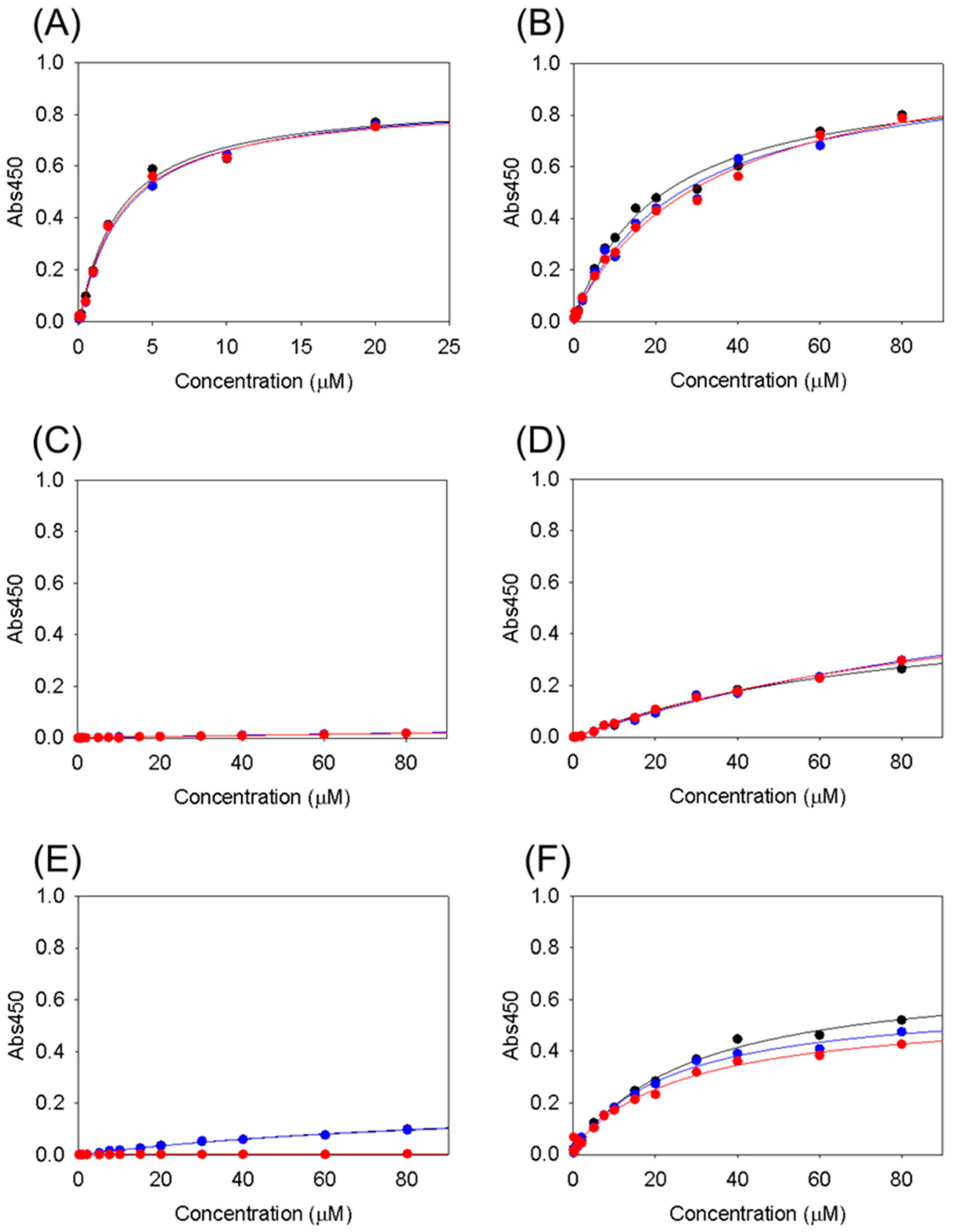
3.2. Collagen-Binding Activity of Cbm/CBD and Its Domain Variants
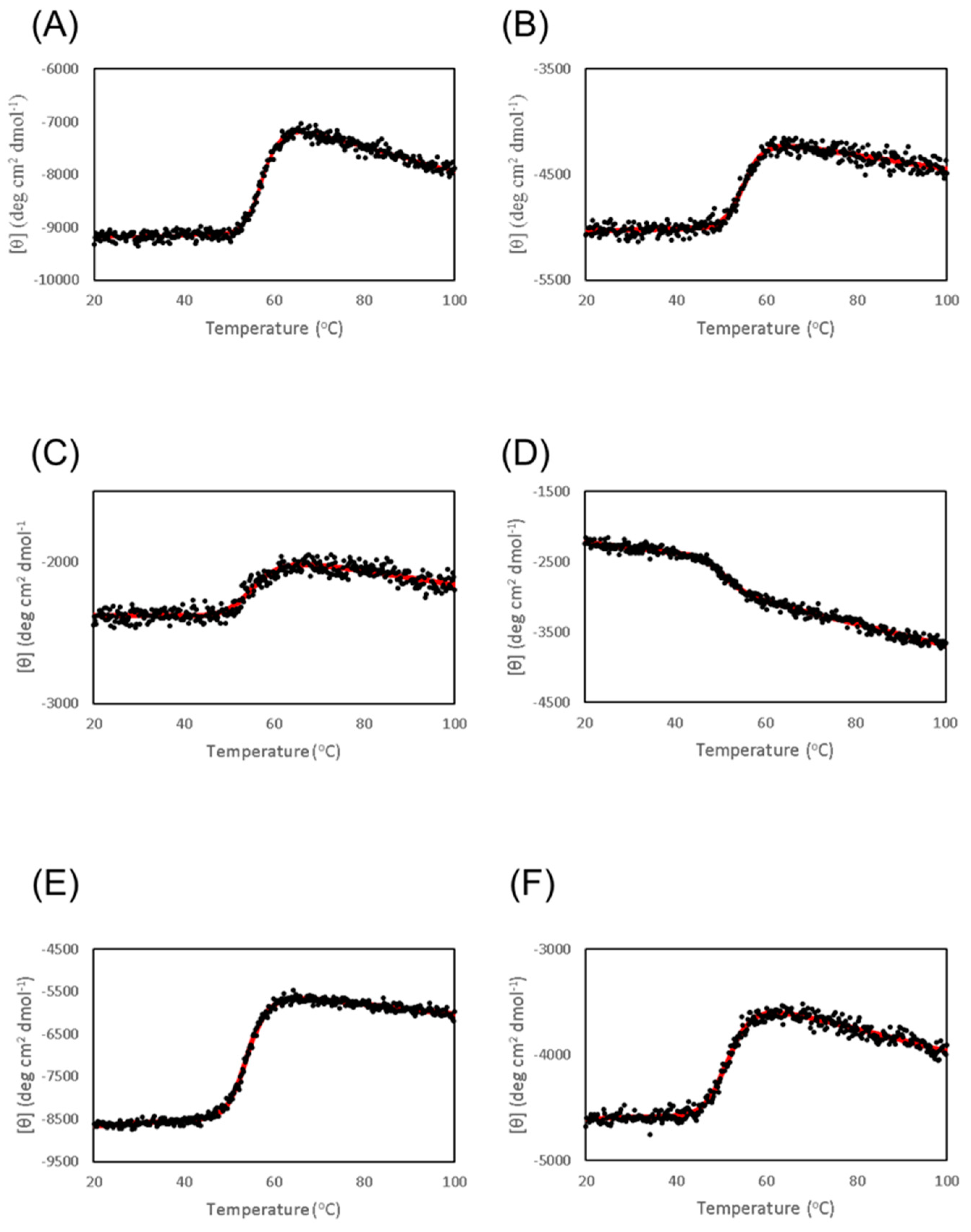
3.3. Thermal Stability of Cbm/CBD and Its Domain Variants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the normal bacterial flora of the oral cavity. J. Clin. Microbiol. 2005, 43, 5721–5732. [Google Scholar] [CrossRef] [PubMed]
- Debelian, G.J.; Olsen, I.; Tronstad, L. Systemic diseases caused by oral microorganisms. Endod. Dent. Traumatol. 1994, 10, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Roberts, G.J.; Lucas, V.S.; Omar, J. Bacterial endocarditis and orthodontics. J. R. Coll. Surg. Edinb. 2000, 45, 141–145. [Google Scholar] [PubMed]
- Seymour, R.A.; Lowry, R.; Whitworth, J.M.; Martin, M.V. Infective endocarditis, dentistry and antibiotic prophylaxis; time for a rethink? Br. Dent. J. 2000, 189, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, S.; Ohara, T.; Ashihara, K.; Izumi, C.; Iwanaga, S.; Eishi, K.; Okita, Y.; Daimon, M.; Kimura, T.; Toyoda, K.; et al. JCS 2017 Guideline on Prevention and Treatment of Infective Endocarditis. Circ. J. 2019, 83, 1767–1809. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.L.; Gordon, S.M.; Shrestha, N.K. Distribution of streptococcal groups causing infective endocarditis: A descriptive study. Diagn. Microbiol. Infect. Dis. 2018, 91, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Nakano, K.; Hokamura, K.; Taniguchi, N.; Wada, K.; Kudo, C.; Nomura, R.; Kojima, A.; Naka, S.; Muranaka, Y.; Thura, M.; et al. The collagen-binding protein of Streptococcus mutans is involved in haemorrhagic stroke. Nat. Commun. 2011, 2, 485. [Google Scholar] [CrossRef] [PubMed]
- Inenaga, C.; Hokamura, K.; Nakano, K.; Nomura, R.; Naka, S.; Ohashi, T.; Ooshima, T.; Kuriyama, N.; Hamasaki, T.; Wada, K.; et al. A Potential new risk factor for stroke: Streptococcus mutans with collagen-binding protein. World Neurosurg. 2018, 113, e77–e81. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Okamoto, K.; Kagami, A.; Yamamoto, Y.; Igarashi, T.; Kizaki, H. Streptococcus mutans strains harboring collagen-binding adhesin. J. Dent. Res. 2004, 83, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Nomura, R.; Nakano, K.; Naka, S.; Nemoto, H.; Masuda, K.; Lapirattanakul, J.; Alaluusua, S.; Matsumoto, M.; Kawabata, S.; Ooshima, T. Identification and characterization of a collagen-binding protein, Cbm, in Streptococcus mutans. Mol. Oral Microbiol. 2012, 27, 308–323. [Google Scholar] [CrossRef] [PubMed]
- Avilés-Reyes, A.; Miller, J.H.; Lemos, J.A.; Abranches, J. Collagen-binding proteins of Streptococcus mutans and related streptococci. Mol. Oral. Microbiol. 2017, 32, 89–106. [Google Scholar] [CrossRef]
- Matsumoto-Nakano, M.; Fujita, K.; Ooshima, T. Comparison of glucan-binding proteins in cariogenicity of Streptococcus mutans. Oral Microbiol. Immunol. 2007, 22, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Nomura, R.; Otsugu, M.; Naka, S.; Teramoto, N.; Kojima, A.; Muranaka, Y.; Matsumoto-Nakano, M.; Ooshima, T.; Nakano, K. Contribution of the interaction of Streptococcus mutans serotype k strains with fibrinogen to the pathogenicity of infective endocarditis. Infect. Immun. 2014, 82, 5223–5234. [Google Scholar] [CrossRef] [PubMed]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar] [PubMed]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Xu, Y.; Liang, X.; Keene, D.R.; Höök, A.; Gurusiddappa, S.; Höök, M.; Narayana, S.V. A ‘Collagen Hug’ model for Staphylococcus aureus CNA binding to collagen. EMBO J. 2005, 24, 4224–4236. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ponnuraj, K.; Xu, Y.; Ganesh, V.K.; Sillanpää, J.; Murray, B.E.; Narayana, S.V.; Höök, M. The Enterococcus faecalis MSCRAMM ACE binds its ligand by the Collagen Hug model. J. Biol. Chem. 2007, 282, 19629–19637. [Google Scholar] [CrossRef] [PubMed]
- Herman-Bausier, P.; Valotteau, C.; Pietrocola, G.; Rindi, S.; Alsteens, D.; Foster, T.J.; Speziale, P.; Dufrêne, Y.F. Mechanical strength and inhibition of the Staphylococcus aureus collagen-binding protein Cna. mBio 2016, 7, e01529-16. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Variant | Residue Numbers | |
|---|---|---|
| Cnm | Cnm/CBD (N1-N2~) | 30–325 |
| Cbm | Cbm/CBD (N1-N2~) | 31–330 |
| N1-N2 | 31–320 | |
| N1N2~ | 31–163/172–330 | |
| N1 | 31–163 | |
| N2 | 172–320 | |
| N2-N1~ | 172–320/164–171/31–163/321–330 |
| Cnm/CBD | |
|---|---|
| Data collection | |
| Space group | P41212 |
| Unit-cell parameters | |
| a, b, c (Å) | 40.91, 40.91, 364.89 |
| α, β, γ (deg) | 90, 90, 90 |
| Resolution range (Å) | 50.0–1.81 (1.84–1.81) |
| Reflections measured | 518,784 |
| Unique reflections | 53,762 |
| Completeness (%) | 99.4 (98.8) |
| Redundancy | 9.7 (8.2) |
| Rmerge (%) | 9.3 (84.5) |
| Rmeas (%) | 9.8 (90.0) |
| Rpim (%) | 2.9 (29.6) |
| CC1/2 (%) | 94.5 (73.6) |
| I/σ | 27.2 (2.1) |
| Refinement | |
| Resolution limits (Å) | 45.61–1.81 |
| Rwork/Rfree | 0.222/0.247 |
| B-factors (Å2) | |
| Protein | 33.0 |
| Sulfate ion/glycerol/water | 57.7/53.2/39.2 |
| R.m.s deviations from ideal values | |
| Bond lengths (Å)/bond angles (deg) | 0.015/2.01 |
| Ramachandran plot | |
| Favored (%) | 97.21 |
| Allowed (%) | 2.79 |
| Disallowed (%) | 0 |
| KD (μM) | |
|---|---|
| Cbm/CBD (N1-N2~) | 2.8 ± 0.2 |
| N1-N2 | 28 ± 6.5 |
| N1N2~ | ND |
| N1 | ND |
| N2 | ND |
| N2-N1~ | 27 ± 2.9 |
| Tm (°C) | |
|---|---|
| Cbm/CBD (N1-N2~) | 57.6 ± 0.1 |
| N1-N2 | 55.2 ± 0.1 |
| N1N2~ | 56.0 ± 0.3 |
| N1 | 50.7 ± 0.3 |
| N2 | 54.2 ± 0.1 |
| N2-N1~ | 51.9 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishi, A.; Matsui, H.; Hirata, A.; Mukaiyama, A.; Tanaka, S.-i.; Yoshizawa, T.; Matsumura, H.; Nomura, R.; Nakano, K.; Takano, K. Structure, Stability and Binding Properties of Collagen-Binding Domains from Streptococcus mutans. Chemistry 2023, 5, 1911-1920. https://doi.org/10.3390/chemistry5030130
Nishi A, Matsui H, Hirata A, Mukaiyama A, Tanaka S-i, Yoshizawa T, Matsumura H, Nomura R, Nakano K, Takano K. Structure, Stability and Binding Properties of Collagen-Binding Domains from Streptococcus mutans. Chemistry. 2023; 5(3):1911-1920. https://doi.org/10.3390/chemistry5030130
Chicago/Turabian StyleNishi, Akari, Hikaru Matsui, Azumi Hirata, Atsushi Mukaiyama, Shun-ichi Tanaka, Takuya Yoshizawa, Hiroyoshi Matsumura, Ryota Nomura, Kazuhiko Nakano, and Kazufumi Takano. 2023. "Structure, Stability and Binding Properties of Collagen-Binding Domains from Streptococcus mutans" Chemistry 5, no. 3: 1911-1920. https://doi.org/10.3390/chemistry5030130