
Exogenous Selenium Improves Physio-Biochemical and Performance of Drought-Stressed Phaseolus vulgaris Seeded in Saline Soil
 , , , , ,
, , , , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site of Experimentation
2.2. Plant Management, Experimental Design, and Treatments
2.3. Irrigation Water Application
2.4. Bean Growth and Green Pods Yield
2.5. Leaf Relative Water Content, Membrane Stability, and Irrigation Use Efficiency
2.6. The Photosynthetic Performance
2.7. Osmolytes and Antioxidative Compounds Quantification
2.8. Statistical Analysis
3. Results
3.1. The Photosynthetic Machinery, Cell Membrane Stability, and Water Status of Bean Plants
3.2. Osmolytes and Antioxidative Compounds of Bean Plants
3.3. Growth and Productivity Parameters of Beans Plants
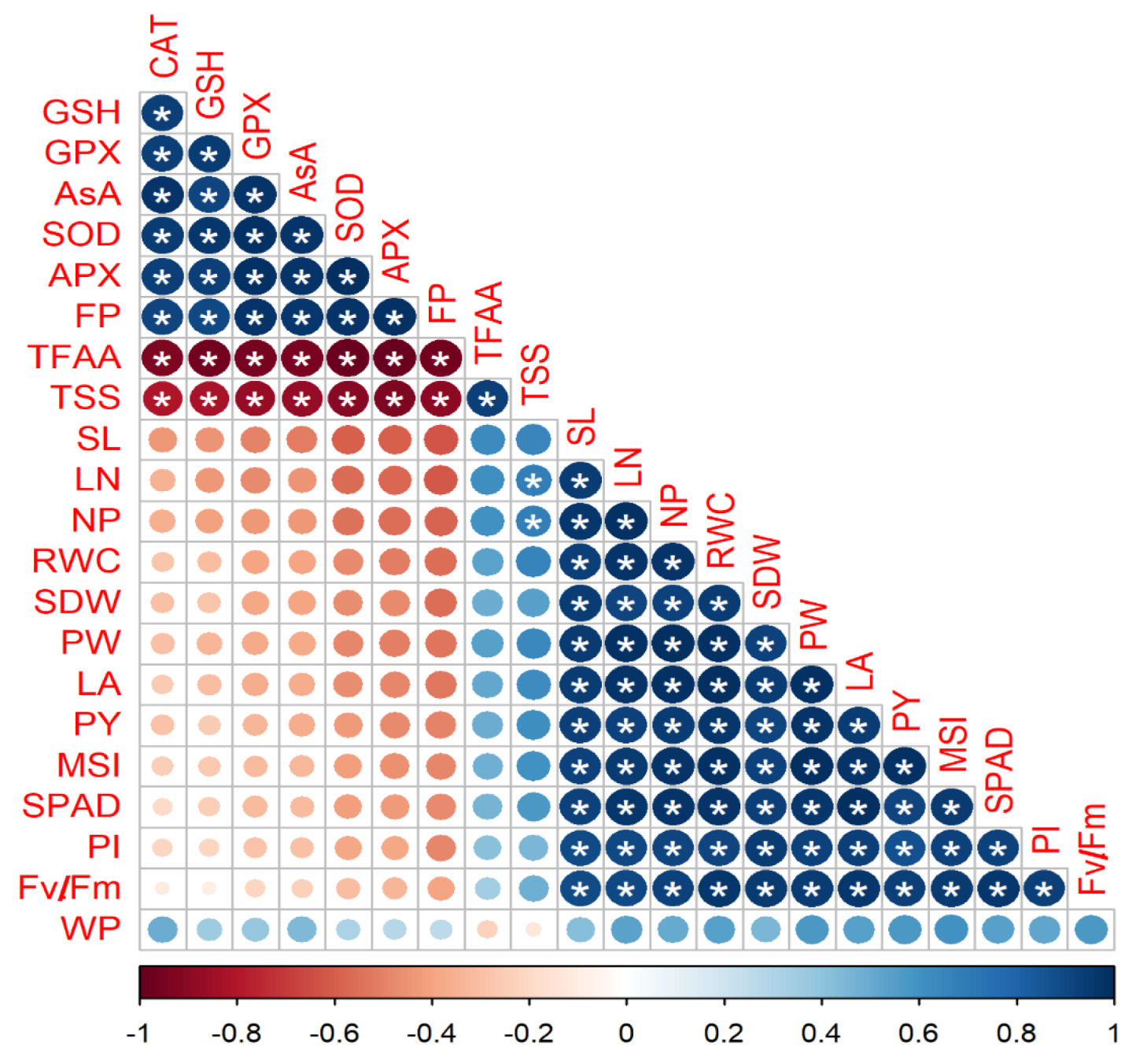
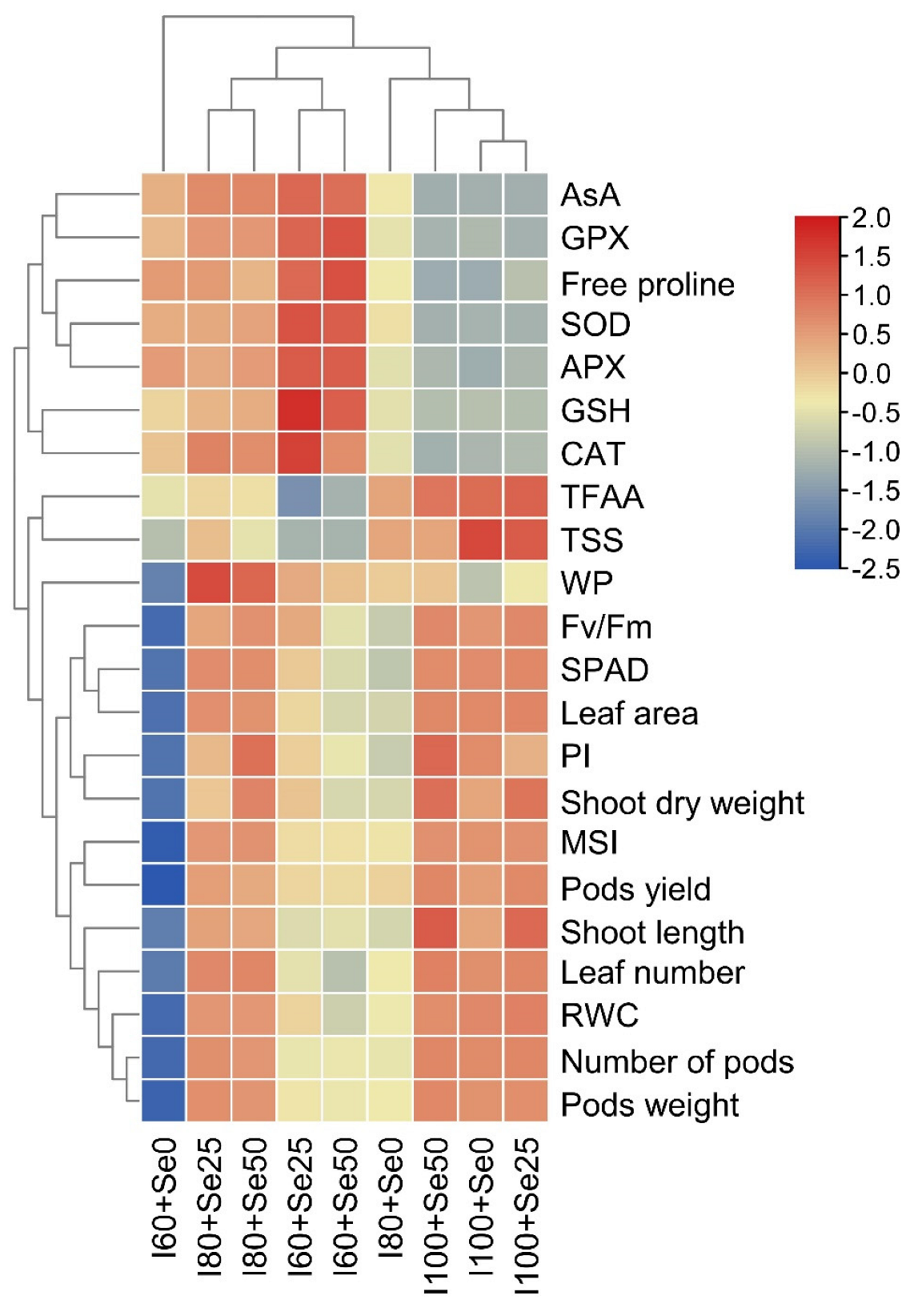
3.4. Relationships
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rady, M.M.; Talaat, N.B.; Abdelhamid, M.T.; Shawky, B.T.; Desoky, E.S.M. Maize (Zea mays L.) grains extract mitigates the deleterious effects of salt stress on common bean (Phaseolus vulgaris L.) growth and physiology. J. Hortic. Sci. Biotechnol. 2019, 94, 777–789. [Google Scholar] [CrossRef]
- Durigon, A.; Evers, J.; Metselaar, K.; van Lier, Q.d.J. Water stress permanently alters shoot architecture in common bean plants. Agronomy 2019, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#data (accessed on 2 September 2022).
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Rady, M.O.A.; Marey, R.A.; Abd El-Mageed, T.A. Exogenously applied proline enhances growth and productivity of drought stressed onion by improving photosynthetic efficiency, water use efficiency and up-regulating osmoprotectants. Sci. Hortic. 2020, 272, 109580. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Shaaban, A.; Abd El-Mageed, S.A.; Semida, W.M.; Rady, M.O.A. Defensive Role in Maize (Zea mays L.) against Drought Stress and Metals-Contaminated Irrigation Water. Silicon 2020, 13, 2165–2176. [Google Scholar] [CrossRef]
- Abdelkhalik, A.; Pascual, B.; Nájera, I.; Baixauli, C.; Pascual-Seva, N. Deficit irrigation as a sustainable practice in improving irrigation water use efficiency in cauliflower under Mediterranean conditions. Agronomy 2019, 9, 732. [Google Scholar] [CrossRef] [Green Version]
- Hammad, S.A.R.; Ali, O.A.M. Physiological and biochemical studies on drought tolerance of wheat plants by application of amino acids and yeast extract. Ann. Agric. Sci. 2014, 59, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Zhu, Y.; Zhao, Y.; Wang, Y.; Li, J.; Wang, Q.; Liu, Y. De novo transcriptome sequencing and analysis of salt-, alkali-, and drought-responsive genes in Sophora alopecuroides. BMC Genomics 2020, 21, 423. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Paul, K.; Pauk, J.; Kondic-Spika, A.; Grausgruber, H.; Allahverdiyev, T.; Sass, L.; Vass, I. Co-occurrence of mild salinity and drought synergistically enhances biomass and grain retardation in wheat. Front. Plant Sci. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Agami, R.A.; Alamri, S.A.M.; Abd El-Mageed, T.A.; Abousekken, M.S.M.; Hashem, M. Salicylic acid and proline enhance water use efficiency, antioxidant defense system and tissues’ anatomy of wheat plants under field deficit irrigation stress. J. Appl. Bot. Food Qual. 2019, 92, 360–370. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.M.; Boriek, S.H.K.; El-mageed, T.A.A.; El-yazal, M.A.S.; Ali, E.F.; Hassan, F.A.S.; Abdelkhalik, A. Exogenous Gibberellic Acid or Dilute Bee Honey Boosts Drought Stress Tolerance in Vicia faba by Rebalancing Osmoprotectants, Antioxidants, Nutrients, and Phytohormones. Plants 2021, 10, 748. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Mageed, T.A.; Semida, W.M. Effect of deficit irrigation and growing seasons on plant water status, fruit yield and water use efficiency of squash under saline soil. Sci. Hortic. 2015, 186, 89–100. [Google Scholar] [CrossRef]
- Abid, M.; Tian, Z.; Ata-Ul-Karim, S.T.; Liu, Y.; Cui, Y.; Zahoor, R.; Jiang, D.; Dai, T. Improved tolerance to post-anthesis drought stress by pre-drought priming at vegetative stages in drought-tolerant and -sensitive wheat cultivars. Plant Physiol. Biochem. 2016, 106, 218–227. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Abdelkhalik, A.; Pascual, B.; Nájera, I.; Domene, M.A.; Baixauli, C.; Pascual-Seva, N. Effects of deficit irrigation on the yield and irrigation water use efficiency of drip-irrigated sweet pepper (Capsicum annuum L.) under Mediterranean conditions. Irrig. Sci. 2020, 38, 89–104. [Google Scholar] [CrossRef]
- Goharrizi, K.J.; Baghizadeh, A.; Kalantar, M.; Fatehi, F. Combined effects of salinity and drought on physiological and biochemical characteristics of pistachio rootstocks. Sci. Hortic. 2020, 261, 108970. [Google Scholar] [CrossRef]
- Sezen, S.M.; Yazar, A.; Akyildiz, A.; Dasgan, H.Y.; Gencel, B. Yield and quality response of drip irrigated green beans under full and deficit irrigation. Sci. Hortic. 2008, 117, 95–102. [Google Scholar] [CrossRef]
- Adda, A.; Regagba, Z.; Latigui, A.; Merah, O. Effect of salt stress on α-amylase activity, sugars mobilization and osmotic potential of Phaseolus vulgaris L. seeds var. ‘Cocorose’ and “Djadida” during germination. J. Biol. Sci. 2014, 14, 370–375. [Google Scholar] [CrossRef] [Green Version]
- Abd El-mageed, T.A.; Abdurrahman, H.A.; El-mageed, S.A.A. Residual acidified biochar modulates growth, physiological responses, and water relations of maize (Zea mays) under heavy metal—contaminated irrigation water. Environ. Sci. Pollut. Res. 2020, 27, 22956–22966. [Google Scholar] [CrossRef] [PubMed]
- Abd El-mageed, T.A.; Abdelkhalik, A.; Abd El-Mageed, S.A.; Semida, W.M. Co-composted Poultry Litter Biochar Enhanced Soil Quality and Eggplant Productivity Under Different Irrigation Regimes. J. Soil Sci. Plant Nutr. 2021. [Google Scholar] [CrossRef]
- Abdelhamid, M.T.; Selim, E.; El-Ghamry, A. Integrated effects of bio and mineral fertilizers and humic substances on growth, yield and nutrient contents of fertigated cowpea (Vigna unguiculata L.) grown on sandy soils. J. Agron. 2011, 10, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Soualem, M.; Adda, A.; Belkhodja, M.; Merah, O. Calcium supply reduced effect of salinity on growth in the Mediterranean shrub (Atriplex halimus L.). Life Sci. J. 2014, 11, 278–284. [Google Scholar]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Nawaz, F.; Naeem, M.; Ashraf, M.Y.; Tahir, M.N.; Zulfiqar, B.; Salahuddin, M.; Shabbir, R.N.; Aslam, M. Selenium supplementation affects physiological and biochemical processes to improve fodder yield and quality of maize (Zea mays L.) under water deficit conditions. Front. Plant Sci. 2016, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.M.; Belal, H.E.E.; Gadallah, F.M.; Semida, W.M. Selenium application in two methods promotes drought tolerance in Solanum lycopersicum plant by inducing the antioxidant defense system. Sci. Hortic. 2020, 266, 109290. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H. Effect of exogenous selenium supply on photosynthesis, Na + accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Djanaguiraman, M.; Prasad, P.V.V.; Seppanen, M. Selenium protects sorghum leaves from oxidative damage under high temperature stress by enhancing antioxidant defense system. Plant Physiol. Biochem. 2010, 48, 999–1007. [Google Scholar] [CrossRef]
- Li, H.; Liu, X.; Wassie, M.; Chen, L. Selenium supplementation alleviates cadmium-induced damages in tall fescue through modulating antioxidant system, photosynthesis efficiency, and gene expression. Environ. Sci. Pollut. Res. 2020. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Akbar, A.; Parveen, A.; Rasheed, R.; Hussain, I. Phenological application of selenium differentially improves growth, oxidative defense and ion homeostasis in maize under salinity stress. Plant Physiol. Biochem. 2018, 123, 268–280. [Google Scholar] [CrossRef]
- Semida, W.M.; Abd El-mageed, T.A.; Abdelkhalik, A.; Hemida, K.A.; Abdurrahman, H.A.; Howladar, S.M.; Leilah, A.A.A.; Rady, M.O.A. Selenium Modulates Antioxidant Activity, Osmoprotectants, and Photosynthetic Efficiency of Onion under Saline Soil Conditions. Agronomy 2021, 11, 855. [Google Scholar] [CrossRef]
- Subramanyam, K.; Laing, G.D.; Van Damme, E.J.M. Sodium selenate treatment using a combination of seed priming and foliar spray alleviates salinity stress in rice. Front. Plant Sci. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuznetsov, V.V.; Kholodova, V.P.; Kuznetsov, V.I.V.; Yagodin, B. Selenium regulates the water status of plants exposed to drought. Dokl. Biol. Sci. 2003, 390, 266–268. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Qi, Z.; Li, M.; Jalal, G.; Chu, X.; Zhou, J. Selenium forms and methods of application differentially modulate plant growth, photosynthesis, stress tolerance, selenium content and speciation in Oryza sativa L. Ecotoxicol. Environ. Saf. 2019, 169, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Klute, A. Methods of Soil Analysis: Part 1 Physical and Mineralogical Methods, 2nd ed.; the American Society of Agronomy, Inc.; Soil Science Society of America, Inc.: Madison, WI, USA, 1986. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis Part 2. Chemical and Microbiological Properties; American Society of Agronomy, Inc.: Madison, WI, USA, 1982. [Google Scholar]
- Ponce, V.M.; Pandey, R.P.; Ercan, S. Characterization of drought across climatic spectrum. J. Hydrol. Eng. 2000, 5, 222–224. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Requirements; Irrigation and Drainage Paper No. 56; FAO: Rome, Italy, 1988. [Google Scholar]
- Semida, W.M.; Abd El-Mageed, T.A.; Mohamed, S.E.; El-Sawah, N.A. Combined effect of deficit irrigation and foliar-applied salicylic acid on physiological responses, yield, and water-use efficiency of onion plants in saline calcareous soil. Arch. Agron. Soil Sci. 2017, 63, 1227–1239. [Google Scholar] [CrossRef]
- Osman, A.S.; Rady, M.M. Effect of humic acid as an additive to growing media to enhance the production of eggplant and tomato transplants. J. Hortic. Sci. Biotechnol. 2014, 89, 237–244. [Google Scholar] [CrossRef]
- Rady, M.M. Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Clark, A.J.; Landolt, W.; Bucher, J.B.; Strasser, R.J. Beech (Fagus sylvatica) response to ozone exposure assessed with a chlorophyll a fluorescence performance index. Environ. Pollut. 2000, 109, 501–507. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 207, 205–207. [Google Scholar] [CrossRef]
- Rosen, H. A modified ninhydrin colorimetric analysis for amino acids. Arch. Biochem. Biophys. 1957, 67, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Van Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Griffith, W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Thomas, R.L.; Jen, J.J.; Morr, C.V. Changes in Soluble and Bound Peroxidase—IAA Oxidase During Tomato Fruit Development. J. Food Sci. 1982, 47, 158–161. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B- and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Khan, Z.; Thounaojam, T.C.; Chowdhury, D.; Upadhyaya, H. The role of selenium and nano selenium on physiological responses in plant: A review. Plant Growth Regul. 2023, 100, 409–433. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalik, A.; Pascual, B.; Nájera, I.; Baixauli, C.; Pascual-Seva, N. Regulated deficit irrigation as a water-saving strategy for onion cultivation in Mediterranean conditions. Agronomy 2019, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Abdou, N.M.; Abdel-razek, M.A.; Abd El-mageed, S.A.; Semida, W.M.; Leilah, A.A.A.; Abd El–Mageed, T.A.; Ali, E.F.; Majrashi, A.; Rady, M.O.A. High Nitrogen Fertilization Modulates Morpho-Physiological Responses, Yield, and Water Productivity of Lowland Rice under Deficit Irrigation. Agronomy 2021, 11, 1291. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Mohamed, G.F.; Abd El-mageed, T.A.; El-Mageed, S.A.A.; Rady, M.M.; Ali, E.F. Foliar Application of Zinc Oxide Nanoparticles Promotes Drought Stress Tolerance in Eggplant (Solanum melongena L.). Plants 2021, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Badal, E.; El-Mageed, T.A.A.; Buesa, I.; Guerra, D.; Bonet, L.; Intrigliolo, D.S. Moderate plant water stress reduces fruit drop of “Rojo Brillante” persimmon (Diospyros kaki) in a Mediterranean climate. Agric. Water Manag. 2013, 119, 154–160. [Google Scholar] [CrossRef]
- Belal, H.E.E.; Rady, M.M.; Gadallah, F.M.; Semida, W.M. Selenium Application in Two Methods Enhances Drought Tolerance By Improving Physiological Attributes in Solanum Lycopersicum Plant. Fayoum J. Agric. Res. Dev. 2020, 34, 113–128. [Google Scholar] [CrossRef]
- Rady, M.O.A.; Semida, W.M.; Howladar, S.M.; Abd El-Mageed, T.A. Raised beds modulate physiological responses, yield and water use efficiency of wheat (Triticum aestivum L.) under deficit irrigation. Agric. Water Manag. 2021, 245, 106629. [Google Scholar] [CrossRef]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front. Plant Sci. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Turner, N.C. Turgor maintenance by osmotic adjustment: 40 years of progress. J. Exp. Bot. 2018, 69, 3223–3233. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.O.A.; Semida, W.M.; El-Mageed, T.A.A.; Howladar, S.M.; Shaaban, A. Foliage applied selenium improves photosynthetic efficiency, antioxidant potential and wheat productivity under drought stress. Int. J. Agric. Biol. 2020, 24, 1293–1300. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenomenon. Planta 2020, 251, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alyemeni, M.N.; Ahanger, M.A.; Wijaya, L.; Alam, P.; Bhardwaj, R.; Ahmad, P. Selenium mitigates cadmium-induced oxidative stress in tomato (Solanum lycopersicum L.) plants by modulating chlorophyll fluorescence, osmolyte accumulation, and antioxidant system. Protoplasma 2018, 255, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Nazir, F.; Asgher, M.; Per, T.S.; Khan, N.A. Selenium and sulfur influence ethylene formation and alleviate cadmium-induced oxidative stress by improving proline and glutathione production in wheat. J. Plant Physiol. 2015, 173, 9–18. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Borhannuddin Bhuyan, M.H.M.; Anee, T.I.; Parvin, K.; Nahar, K.; Al Mahmud, J.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Avramova, V.; Abdelgawad, H.; Zhang, Z.; Fotschki, B.; Casadevall, R.; Vergauwen, L.; Knapen, D.; Taleisnik, E.; Guisez, Y.; Asard, H.; et al. Drought induces distinct growth response, protection, and recovery mechanisms in the maize leaf growth zone. Plant Physiol. 2015, 169, 1382–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taha, R.S.; Seleiman, M.F.; Shami, A.; Alhammad, B.A.; Mahdi, A.H.A. Growth and Anatomical Structure, Antioxidant Defense System and Yield of Wheat Grown in Salt-Stressed Soil. Plants 2021, 10, 1–19. [Google Scholar] [CrossRef]
- Desoky, E.M.; Merwad, A.M.; Semida, W.M.; Ibrahim, S.A.; El-saadony, M.T.; Rady, M.M. Heavy metals-resistant bacteria (HM-RB): Potential bioremediators of heavy metals-stressed Spinacia oleracea plant. Ecotoxicol. Environ. Saf. 2020, 198, 110685. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Semida, W.M.; Rady, M.M.; Mohamed, G.F.; Hemida, K.A.; Alhammad, B.A.; Hassan, M.M.; Shami, A. Sequential application of antioxidants rectifies ion imbalance and strengthens antioxidant systems in salt-stressed cucumber. Plants 2020, 9, 1783. [Google Scholar] [CrossRef]
- Proietti, P.; Nasini, L.; Del, D.; Amato, R.D.; Tedeschini, E.; Businelli, D. Selenium protects olive (Olea europaea L.) from drought stress. Sci. Hortic. 2013, 164, 165–171. [Google Scholar] [CrossRef]
- Nassar, R.M.A.; Kamel, H.A.; Ghoniem, A.E.; Alarcón, J.J.; Sekara, A.; Ulrichs, C.; Abdelhamid, M.T. Physiological and anatomical mechanisms in wheat to cope with salt stress induced by seawater. Plants 2020, 9, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rady, M.M.; Mounzer, O.H.; Alarcón, J.J.; Abdelhamid, M.T.; Howladar, S.M. Growth, heavy metal status and yield of salt-stressed wheat (Triticum aestivum L.) plants as affected by the integrated application of bio-, organic and inorganic nitrogen-fertilizers. J. Appl. Bot. Food Qual. 2016, 89, 21–28. [Google Scholar] [CrossRef]
- Abd El-mageed, T.A.; Gyushi, M.A.H.; Hemida, K.A.; El-Saadony, M.T.; Abd El-Mageed, S.A.; Abdalla, H.; AbuQamar, S.F.; El-Tarabily, K.A.; Abdelkhalik, A. Coapplication of Effective Microorganisms and Nanomagnesium Boosts the Defenses Against Salt Stress in Ipomoea batatas. Front. Plant Sci. 2022, 13, 883274. [Google Scholar] [CrossRef]
- Abdelkhalik, A.; Abd El-mageed, T.A.; Mohamed, I.A.A.; Semida, W.M.; Al-elwany, O.A.A.I.; Ibrahim, I.M.; Hemida, K.A.; El-saadony, M.T.; Abuqamar, S.F.; El-tarabily, K.A. Soil application of effective microorganisms and nitrogen alleviates salt stress in hot pepper (Capsicum annum L.) plants. Front. Plant Sci. 2023, 13, 1079260. [Google Scholar] [CrossRef] [PubMed]
- Martínez-ispizua, E.; Calatayud, Á.; Marsal, J.I.; Cannata, C.; Basile, F.; Abdelkhalik, A.; Soler, S.; Valcárcel, J.V.; Martínez-Cuenca, M.-R. The Nutritional Quality Potential of Microgreens, Baby Leaves, and Adult Lettuce: An Underexploited Nutraceutical Source. Foods 2022, 11, 423. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | EC (dS/m) | pH | OM % | CaCO3 % | Particle Size Distribution | Soil Texture | ρd g.cm−3 | Ksat cm h−1 | Soil Moisture Content at | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sand % | Silt % | Clay % | FC % | WP % | AW % | ||||||||
| 2017 | 6.22 | 7.66 | 1.13 | 4.51 | 74.2 | 12.8 | 13.0 | SL | 1.58 | 2.10 | 21.03 | 10.55 | 10.48 |
| 2018 | 6.18 | 7.70 | 1.15 | 4.42 | 72.6 | 14.5 | 12.9 | SL | 1.55 | 1.96 | 22.2 | 11.4 | 10.8 |
| Month | T. max (°C) | T. min (°C) | T. avg (°C) | RH (%) | U2 ms−1 | Epan mm Day−1 |
|---|---|---|---|---|---|---|
| 2017 | ||||||
| September | 38.3 | 23.6 | 30.95 | 37.0 | 2.1 | 5.85 |
| October | 34.0 | 22.4 | 28.2 | 40.0 | 1.95 | 4.7 |
| November | 27.8 | 15.4 | 21.6 | 41.5 | 2.2 | 2.15 |
| 2018 | ||||||
| September | 37.2 | 22.4 | 31.2 | 38.2 | 1.99 | 5.6 |
| October | 33.2 | 20.3 | 28.6 | 396 | 2.1 | 4.3 |
| November | 26.4 | 14.9 | 20.4 | 41 | 2 | 3.2 |
| Treatments | SPAD | Fv/Fm | PI | MSI | RWC | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | |
| IR | ||||||||||
| I100 | 38.4 ± 0.5a | 38.2 ± 0.4a | 0.82 ± 0.0a | 0.83 ± 0.0a | 2.7 ± 0.2a | 2.9 ± 0.1a | 58.6 ± 1.1a | 59.8 ± 2.4a | 89.0 ± 0.6a | 89.4 ± 1.1a |
| I80 | 34.4 ± 1.1b | 33.7 ± 1.4b | 0.80 ± 0.01ab | 0.81 ± 0.0ab | 2.3 ± 0.3b | 2.7 ± 0.2b | 55.1 ± 2.3b | 55.8 ± 1.9b | 86.2 ± 0.8a | 86.9 ± 1.0a |
| I60 | 27.3 ± 1.7c | 25.9 ± 2.1c | 0.77 ± 0.01b | 0.78 ± 0.05b | 2.0 ± 0.2c | 2.1 ± 0.2c | 42.9 ± 2.1c | 43.4 ± 2.9c | 79.6 ± 0.9b | 80.4 ± 3.3b |
| Se (mg L−1) | ||||||||||
| Se0 | 27.9 ± 2.2c | 27.9 ± 1.8c | 0.77 ± 0.01b | 0.77 ± 0.01b | 1.9 ± 0.1c | 2.3 ± 0.2b | 46.1 ± 2.1b | 46.4 ± 3.3b | 81.7 ± 1.1c | 82.5 ± 1.1b |
| Se25 | 37.4 ± 1.1a | 35.5 ± 0.8a | 0.81 ± 0.00a | 0.83 ± 0.0a | 2.3 ± 0.1b | 2.7 ± 0.01a | 55.3 ± 2.7a | 56.1 ± 1.9a | 87.6 ± 1.1a | 87.4 ± 1.0a |
| Se50 | 34.9 ± 1.0b | 34.5 ± 1.1b | 0.80 ± 0.00a | 0.82 ± 0.0a | 2.8 ± 0.1a | 2.7 ± 0.1a | 55.2 ± 2.7a | 56.5 ± 1.7a | 85.4 ± 1.0b | 86.8 ± 1.0a |
| IR × Se | ||||||||||
| I100 × Se0 | 38.4 ± 1.4a | 37.9 ± 0.9a | 0.82 ± 0.00a | 0.83 ± 0.0ab | 2.7 ± 0.2b | 2.9 ± 0.1a | 58.4 ± 2.9a | 59.7 ± 2.4a | 88.8 ± 1.4a | 89.4 ± 2.3a |
| I100 × Se25 | 38.8 ± 0.3a | 38.6 ± 0.5a | 0.82 ± 0.00a | 0.84 ± 0.0a | 2.3 ± 0.1d | 2.9 ± 0.1a | 58.8 ± 2.9a | 59.7 ± 1.3a | 89.1 ± 0.7a | 90.1 ± 0.9a |
| I100 × Se50 | 38.0 ± 0.4a | 38.2 ± 0.2a | 0.82 ± 0.01a | 0.84 ± 0.0a | 3.2 ± 0.3a | 2.9 ± 0.2a | 58.6 ± 2.7a | 59.9 ± 0.4a | 88.9 ± 0.6a | 88.7 ± 1.5a |
| I80 × Se0 | 26.2 ± 1.7c | 25.9 ± 1.9c | 0.7800.01ab | 0.77 ± 0.01bc | 1.8 ± 0.2f | 2.3 ± 0.3b | 48.6 ± 1.9b | 48.8 ± 2.2b | 82.9 ± 1.6bc | 83.3 ± 0.4b |
| I80 × Se25 | 38.4 ± 1.2a | 37.9 ± 0.4a | 0.81 ± 0.00a | 0.82 ± 0.0ab | 2.1 ± 0.2e | 2.9 ± 0.2a | 57.8 ± 1.1a | 58.9 ± 2.4a | 87.8 ± 1.2a | 88.7 ± 0.7a |
| I80 × Se50 | 38.6 ± 0.4a | 37.4 ± 0.5a | 0.82 ± 0.00a | 0.83 ± 0.0ab | 3.1 ± 0.3a | 2.9 ± 0.2a | 58.8 ± 2.0a | 59.6 ± 2.4a | 87.8 ± 1.3a | 88.5 ± 2.8a |
| I60 × Se0 | 19.0 ± 1.9d | 20.0 ± 0.2d | 0.73 ± 0.02b | 0.73 ± 0.02c | 1.3 ± 0.2g | 1.72 ± 0.2c | 31.2 ± 2.3c | 30.6 ± 0.9c | 73.5 ± 0.2d | 74.8 ± 0.6c |
| I60 × Se25 | 34.9 ± 0.7b | 29.9 ± 0.8b | 0.81 ± 0.00a | 0.82 ± 0.01ab | 2.5 ± 0.1c | 2.38 ± 0.3b | 49.3 ± 2.4b | 49.6 ± 2.2b | 85.7 ± 2.4ab | 83.4 ± 1.2b |
| I60 × Se50 | 28.0 ± 1.1c | 27.8 ± 1.3bc | 0.77 ± 0.01ab | 0.80 ± 0.0ab | 2.1 ± 0.02e | 2.35 ± 0.3b | 48.3 ± 1.3b | 49.9 ± b | 79.5 ± 1.2c | 83.1 ± 0.2b |
| Treatments | Shoot Length (cm) | Leaf No. Plant−1 | Leaf Area Plant−1 (dm2) | Shoot Dry Weight (g Plant−1) | No. of Pods Plant−1 | Pods Weight Plant−1 | Pods Yield (ton ha−1) | WP (kg pods/m3 of Water) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | |
| IR | ||||||||||||||||
| I100 | 80.3 ± 2.7a | 84.5 ± 3.6a | 28.9 ± 0.44a | 29.30 ± 1.2a | 24.3 ± 0.8a | 23.1 ± 0.8a | 23.9 ± 0.8a | 22.8 ± 0.5a | 32.9 ± 0.9a | 32.3 ± 1.3a | 52.9 ± 1.0a | 52.5 ± 0.9a | 10.6 ± 0.2a | 10.66 ± 0.2a | 2.79 ± 0.1b | 3.55 ± 0.1b |
| I80 | 77.8 ± 2.9b | 77.5 ± 1.8b | 27.6 ± 1.06b | 28.2 ± 0.7b | 22.1 ± 1.4b | 21.8 ± 0.6b | 20.7 ± 1.4b | 21.5 ± 0.6b | 28.3 ± 0.8b | 28.1 ± 0.8b | 48.5 ± 1.4b | 48.5 ± 1.2b | 9.9 ± 0.2b | 9.84 ± 0.2b | 3.28 ± 0.1a | 4.15 ± 0.1a |
| I60 | 71.9 ± 1.4c | 72.6 ± 4.2c | 23.8 ± 0.91c | 23.1 ± 1.4c | 17.8 ± 0.7c | 19.1 ± 1.4c | 17.8 ± 0.7c | 19.4 ± 1.6c | 19.0 ± 1.7c | 18.5 ± 2.4c | 36.4 ± 2.0c | 35.9 ± 1.2c | 8.1 ± 0.3c | 7.97 ± 0.2c | 2.66 ± 0.1c | 3.66 ± 0.2b |
| Se (mg L−1) | ||||||||||||||||
| Se0 | 73.2 ± 2.0b | 74.13 ± 3.5b | 25.1 ± 1.41b | 25.3 ± 0.8c | 19.5 ± 0.8b | 19.2 ± 1.0c | 18.6 ± 0.9c | 19.3 ± 1.3c | 22.5 ± 2.1b | 21.7 ± 3.3b | 40.1 ± 3.6b | 39.6 ± 3.0b | 8.5 ± 0.6b | 8.37 ± 0.7b | 2.52 ± 0.1b | 3.42 ± 0.1b |
| Se25 | 78.2 ± 3.3a | 80.12 ± 2.9a | 27.5 ± 1.67a | 28.1 ± 1.2a | 22.3 ± 1.2a | 23.0 ± 0.7a | 20.9 ± 1.2b | 23.0 ± 0.4a | 29.0 ± 1.3a | 28.7 ± 1.8a | 48.8 ± 3.1a | 49.0 ± 2.5a | 10.2 ± 0.5a | 10.00 ± 0.6a | 3.10 ± 0.1a | 4.02 ± 0.1a |
| Se50 | 78.6 ± 3.3a | 80.42 ± 2.2a | 27.7 ± 1.19a | 27.2 ± 0.4b | 22.4 ± 1.1a | 21.8 ± 0.4b | 22.9 ± 1.2a | 21.5 ± 0.7b | 28.8 ± 1.3a | 28.5 ± 1.3a | 48.8 ± 3.3a | 48.4 ± 3.1a | 9.9 ± 0.7a | 10.10 ± 0.6a | 3.12 ± 0.1a | 3.93 ± 0.1a |
| IR × Se | ||||||||||||||||
| I100 × Se0 | 78.9 ± 1.6ab | 80.13 ± 3.8b | 28.6 ± 0.57a | 29.0 ± 2.1a | 24.1 ± 0.6a | 23.1 ± 1.6a | 21.7 ± 0.6c | 22.5 ± 0.4a | 32.6 ± 2.3ab | 31.8 ± 2.0ab | 52.2 ± 1.1a | 52.0 ± 0.8a | 10.4 ± 0.2ab | 10.23 ± 0.1b | 2.57 ± 0.1c | 3.35 ± 0.1ef |
| I100 × Se25 | 80.4 ± 1.2a | 86.33 ± 3.1a | 29.0 ± 0.57a | 29.3 ± 0.3a | 24.2 ± 0.6a | 23.3 ± 0.3a | 24.6 ± 0.6ab | 23.1 ± 0.3a | 32.9 ± 0.3a | 32.7 ± 0.8a | 52.7 ± 0.3a | 52.3 ± 1.2a | 10.9 ± 0.1a | 10.63 ± 0.2ab | 2.84 ± 0.0b | 3.54 ± 0.1de |
| I100 × Se50 | 81.6 ± 0.5a | 87.1 ± 3.5a | 29.0 ± 0.52a | 29.7 ± 0.6a | 24.4 ± 1.0a | 22.9 ± 0.6a | 25.4 ± 1.0a | 22.8 ± 0.6a | 33.3 ± 0.8a | 32.3 ± 1.8ab | 53.7 ± 0.3a | 53.3 ± 0.6a | 10.6 ± 0.1b | 11.10 ± 0.1a | 2.95 ± 0.0b | 3.76 ± 0.1c |
| I80 × Se0 | 73.4 ± 0.0c | 74.2 ± 2.3d | 25.1 ± 0.66b | 26.0 ± 0.0b | 18.6 ± 0.1b | 19.7 ± 0.0b | 18.9 ± 0.4d | 19.2 ± 0.0b | 22.2 ± 1.0c | 21.8 ± 0.0c | 41.8 ± 0.6b | 40.8 ± 0.5b | 9.4 ± 0.1cd | 9.13 ± 0.1c | 2.90 ± 0.0b | 3.76 ± 0.1cd |
| I80 × Se25 | 79.8 ± 2.3a | 79.7 ± 1.4bc | 28.8 ± 0.82a | 29.3 ± 0.6a | 23.9 ± 0.4a | 22.9 ± 0.9a | 19.2 ± 0.4d | 22.7 ± 0.9a | 31.7 ± 1.7ab | 31.7 ± 1.4ab | 51.7 ± 0.3a | 53.3 ± 0.3a | 10.4 ± 0.1ab | 10.23 ± 0.1b | 3.55 ± 0.0a | 4.43 ± 0.0a |
| I80 × Se50 | 80.20 ± 0.3a | 78.7 ± 0.0bcd | 29.0 ± 0.88a | 29.3 ± 0.6a | 23.8 ± 0.3a | 22.8 ± 0.8a | 24.0 ± 0.7b | 22.6 ± 0.8a | 31.0 ± 1.1b | 30.9 ± 0.8b | 51.9 ± 1.4a | 51.3 ± 0.3a | 10.0 ± 0.2bc | 10.17 ± 0.4b | 3.40 ± 0.1a | 4.27 ± 0.2ab |
| I60 × Se0 | 67.4 ± 0.6d | 68.0 ± 2.6e | 21.6 ± 1.40c | 20.8 ± 0.0d | 15.9 ± 0.7c | 14.8 ± 0.5c | 15.1 ± 0.4e | 16.1 ± 0.5c | 12.6 ± 0.8d | 11.4 ± 1.0d | 26.3 ± 1.5 | 25.9 ± 0.6c | 5.83 ± 0.3e | 5.73 ± 0.1d | 2.08 ± 0.1d | 3.14 ± 0.1f |
| I60 × Se25 | 74.3 ± 0.0bc | 74.3 ± 2.7d | 24.7 ± 0.82b | 25.7 ± 0.8b | 18.7 ± 0.4b | 22.8 ± 0.9a | 19.0 ± 0.3d | 23.2 ± 0.8a | 22.4 ± 1.5c | 21.7 ± 0.7c | 42.0 ± 0.3b | 41.3 ± 1.6b | 9.2 ± 0.5d | 9.13 ± 0.3c | 2.90 ± 0.2b | 4.08 ± 0.2bc |
| I60 × Se50 | 74.0 ± 1.5c | 75.3 ± 2.7cd | 25.0 ± 0.57b | 22.7 ± 0.3c | 18.8 ± 0.3b | 19.6 ± 0.8b | 19.2 ± 0.3d | 19.0 ± 1.0b | 22.0 ± 0.8c | 22.3 ± 0.8c | 40.9 ± 1.5b | 40.6 ± 0.3b | 9.2 ± 0.1d | 9.03 ± 0.1c | 3.00 ± 0.0b | 3.75 ± 0.0d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semida, W.M.; Abd El-Mageed, T.A.; Gyushi, M.A.H.; Abd El-Mageed, S.A.; Rady, M.M.; Abdelkhalik, A.; Merah, O.; Sabagh, A.E.; El-Metwally, I.M.; Sadak, M.S.; et al. Exogenous Selenium Improves Physio-Biochemical and Performance of Drought-Stressed Phaseolus vulgaris Seeded in Saline Soil. Soil Syst. 2023, 7, 67. https://doi.org/10.3390/soilsystems7030067
Semida WM, Abd El-Mageed TA, Gyushi MAH, Abd El-Mageed SA, Rady MM, Abdelkhalik A, Merah O, Sabagh AE, El-Metwally IM, Sadak MS, et al. Exogenous Selenium Improves Physio-Biochemical and Performance of Drought-Stressed Phaseolus vulgaris Seeded in Saline Soil. Soil Systems. 2023; 7(3):67. https://doi.org/10.3390/soilsystems7030067
Chicago/Turabian StyleSemida, Wael M., Taia A. Abd El-Mageed, Mohammed A. H. Gyushi, Shimaa A. Abd El-Mageed, Mostafa M. Rady, Abdelsattar Abdelkhalik, Othmane Merah, Ayman El Sabagh, Ibrahim M. El-Metwally, Mervat Sh. Sadak, and et al. 2023. "Exogenous Selenium Improves Physio-Biochemical and Performance of Drought-Stressed Phaseolus vulgaris Seeded in Saline Soil" Soil Systems 7, no. 3: 67. https://doi.org/10.3390/soilsystems7030067