
CO2 Emissions in Layered Cranberry Soils under Simulated Warming


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
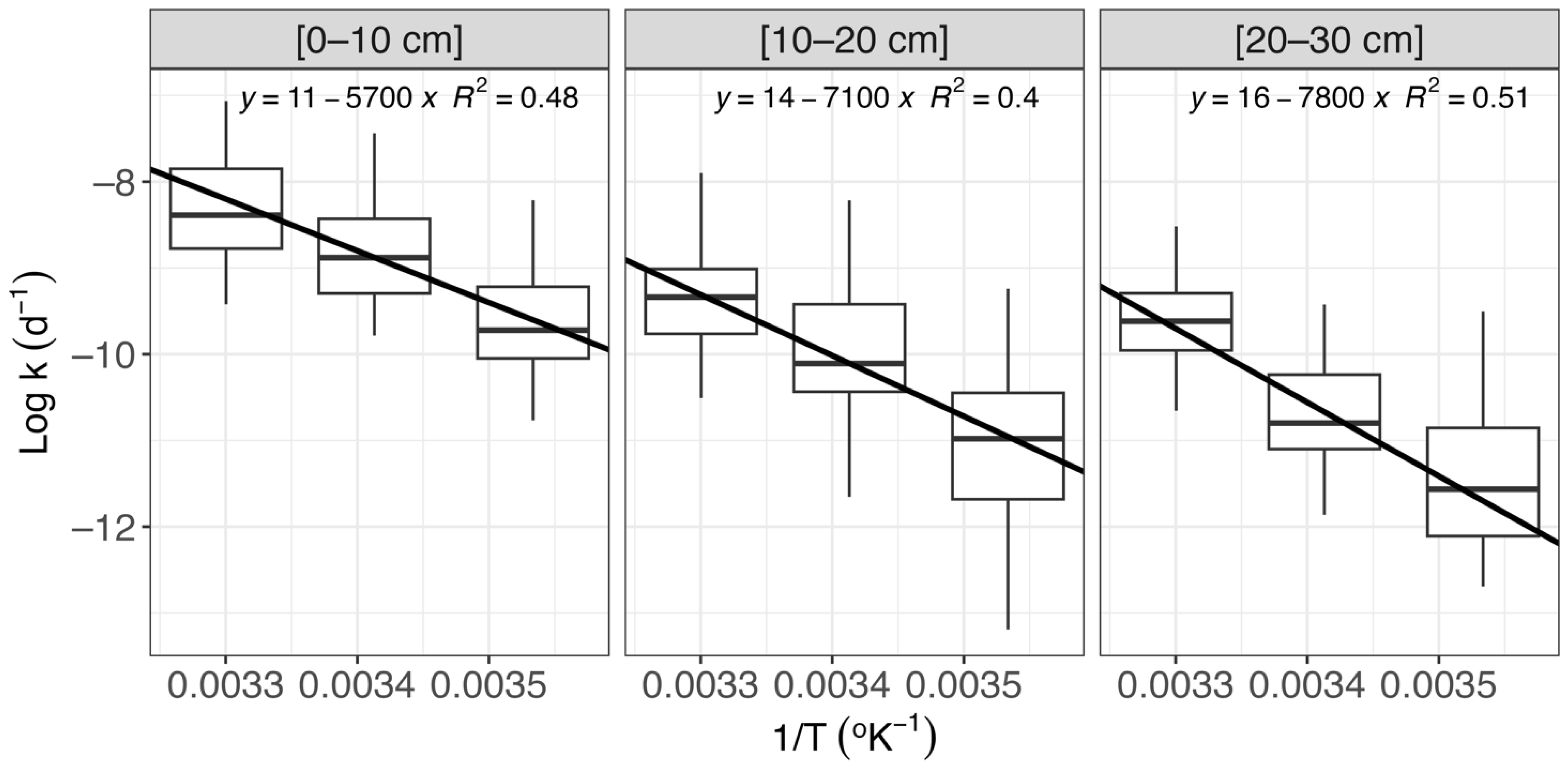
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling and Analysis
2.2. CO2 Emission
2.3. Statistical Analysis
2.3.1. First Order Kinetics
2.3.2. Q10 and Activation Energy
2.3.3. Statistical Analysis
3. Results
3.1. Soil Properties
3.2. CO2 Emission Rate
4. Discussion
4.1. Dependency of CO2 Emission on Soil Depth
4.2. Temperature Sensitivity on CO2 Emission Rates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Post, W.M.; Peng, T.-H.; Emanuel, W.R.; King, A.W.; Dale, V.H.; De Angelis, D.L. The global carbon cycle. Am. Scient. 1990, 78, 310–326. [Google Scholar]
- Lal, R. Soil carbon stocks under present and future climate with specific reference to European ecoregions. Nutr. Cycl. Agroecosyst. 2008, 81, 113–127. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dossou-Yovo, W.; Parent, S.-É.; Ziadi, N.; Parent, É.; Parent, L.E. Tea Bag Index to Assess Carbon Decomposition Rate in Cranberry Agroecosystems. Soil Syst. 2021, 5, 44. [Google Scholar] [CrossRef]
- Angers, D.A.; Chenu, C. Dynamics of soil aggregation and C sequestration. In Soil Processes and the Carbon Cycle; Lal, R., Kimble, J.M., Follett, R.F., Stewart, B.A., Eds.; CRC Press: Boca Raton, FL, USA, 1997; pp. 199–206. [Google Scholar]
- Hassink, J.; Whitmore, A.P.; Kuba, J. Size and density fractionation of soil organic matter and the physical Size and density fractionation of soil organic matter and the physical capacity of soils to protect organic matter. Eur. J. Agron. 1997, 7, 189–199. [Google Scholar] [CrossRef]
- Kennedy, C.D.; Wilderotter, S.; Payne, M.; Buda, A.R.; Kleinman, P.J.A.; Bryant, R.B. A geospatial model to quantify mean thickness of peat in cranberry bogs. Geoderma 2018, 319, 122–131. [Google Scholar] [CrossRef]
- Stackpoole, S.M.; Kosola, K.R.; Workmaster, B.A.A.; Guldan, N.M.; Browne, B.A.; Jackson, R.D. Looking beyond fertilizer: Assessing the contribution of nitrogen from hydrologic inputs and organic matter to plant growth in the cranberry agroecosystem. Nutr. Cycl. Agroecosyst. 2011, 91, 41–54. [Google Scholar] [CrossRef]
- Sandler, H.; DeMoranville, C. Cranberry Production: A Guide for Massachusetts. Summary Edition; University of Massachusetts Cranberry Station: Amherst, MA, USA, 2008; pp. 1–198. [Google Scholar]
- Kosola, K.R.; Workmaster, B.A.A. Mycorrhizal colonization of cranberry: Effects of cultivar, soil type, and leaf litter composition. J. Am. Soc. Hortic. Sci. 2007, 132, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Eilers, K.G.; Debenport, S.; Anderson, S.; Fierer, N. Digging deeper to find unique microbial communities: The strong effect of depth on the structure of bacterial and archaeal communities in soil. Soil Biol. Biochem. 2012, 50, 58–65. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Kramer, C.; Gleixner, G. Soil organic matter in soil depth profiles: Distinct carbon preferences of microbial groups during carbon transformation. Soil Biol. Biochem. 2008, 40, 425–433. [Google Scholar] [CrossRef]
- Balesdent, J.; Basile-Doelsch, I.; Chadoeuf, J.; Cornu, S.; Derrien, D.; Fekiacova, Z.; Hatté, C. Atmosphere–soil carbon transfer as a function of soil depth. Nature 2018, 559, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Beier, C.; Rasmussen, L. Organic matter decomposition in an acidic forest soil in Denmark as measured by the cotton strip assay. Scand. J. For. Res. 1994, 9, 106–114. [Google Scholar] [CrossRef]
- Li, J.; Yan, D.; Pendall, E.; Pei, J.; Noh, N.J.; He, J.S.; Li, B.; Nie, M.; Fang, C. Depth dependence of soil carbon temperature sensitivity across Tibetan permafrost regions. Soil Biol. Biochem. 2018, 126, 82–90. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, M.I.; Chivas, A.R.; Head, J. A latitudinal gradient in carbon turnover times in forest soils. Nature 1996, 381, 143–146. [Google Scholar] [CrossRef]
- Von Lützow, M.; Kögel-Knabner, I. Temperature sensitivity of soil organic matter decomposition-what do we know? Biol. Fertil. Soil 2009, 46, 1–15. [Google Scholar] [CrossRef]
- Lloyd, K.; Madramootoo, C.A.; Edwards, K.P.; Grant, A. Greenhouse gas emissions from selected horticultural production systems in a cold temperate climate. Geoderma 2019, 349, 45–55. [Google Scholar] [CrossRef]
- Lee, C.G.; Suzuki, S.; Inubushi, K. Temperature sensitivity of anaerobic labile soil organic carbon decomposition in brackish marsh. Soil Sci. Plant Nutr. 2018, 64, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Sierra, C.A. Temperature sensitivity of organic matter decomposition in the Arrhenius equation: Some theoretical considerations. Biogeochemistry 2012, 108, 1–15. [Google Scholar] [CrossRef]
- Kowalenko, C.G. Assessment of Leco CNS-2000 analyzer for simultaneously measuring total carbon, nitrogen, and sulphur in soil. Commun. Soil Sci. Plant Anal. 2001, 32, 2065–2078. [Google Scholar] [CrossRef]
- Linn, D.M.; Doran, J.W. Effect of Water-Filled Pore Space on Carbon Dioxide and Nitrous Oxide Production in Tilled and Nontilled Soils. Soil Sci. Soc. Am. J. 1984, 48, 1267–1272. [Google Scholar] [CrossRef] [Green Version]
- Verdonck, O.F.; Cappaert, I.M.; De Boodt, M.F. Physical characterization of horticutural substrates. Acta Hortic. 1978, 178, 191–200. [Google Scholar] [CrossRef]
- Gagnon, B.; Ziadi, N.; Rochette, P.; Chantigny, M.H.; Angers, D.A.; Bertrand, N.; Smith, W.N. Soil-surface carbon dioxide emission following nitrogen fertilization in corn. Can. J. Soil Sci. 2016, 96, 219–232. [Google Scholar] [CrossRef]
- Newton, L.S.J.; Lopes, A.; Spokas, K.; Archer, D.W.; Reicosky, D. First-order decay models to describe soil C-CO2 loss after rotary tillage. Sci. Agric. 2009, 66, 650–657. [Google Scholar]
- Verzani, J. Using R for Introductory Statistics; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Dalgaard, P. Multiple Regression. In Introductory Statistics with R; Springer Science + Business Media, LLC.: Berlin/Heidelberg, Germany, 2008; pp. 185–194. [Google Scholar]
- Cai, A.; Feng, W.; Zhang, W.; Xu, M. Climate, soil texture, and soil types affect the contributions of fine-fraction-stabilized carbon to total soil organic carbon in different land uses across China. J. Environ. Manag. 2016, 172, 2–9. [Google Scholar] [CrossRef]
- Fissore, C.; Jurgensen, M.F.; Pickens, J.; Miller, C.; Page-Dumroese, D.; Giardina, C.P. Role of soil texture, clay mineralogy, location, and temperature in coarse wood decomposition-A mesocosm experiment. Ecosphere 2016, 7, e01605. [Google Scholar] [CrossRef]
- McInerney, M.; Bolger, T. Temperature, wetting cycles and soil texture effects on carbon and nitrogen dynamics in stabilized earthworm casts. Soil Biol. Biochem. 2000, 32, 335–349. [Google Scholar] [CrossRef]
- Sugihara, S.; Funakawa, S.; Kilasara, M.; Kosaki, T. Effects of land management on CO2 flux and soil C stock in two Tanzanian croplands with contrasting soil texture. Soil Biol. Biochem. 2012, 46, 1–9. [Google Scholar] [CrossRef]
- Ngao, J.; Epron, D.; Delpierre, N.; Bréda, N.; Granier, A.; Longdoz, B. Spatial variability of soil CO2 efflux linked to soil parameters and ecosystem characteristics in a temperate beech forest. Agric. For. Meteorol. 2012, 154–155, 136. [Google Scholar] [CrossRef]
- De Graaff, M.A.; Jastrow, J.D.; Gillette, S.; Johns, A.; Wullschleger, S.D. Differential priming of soil carbon driven by soil depth and root impacts on carbon availability. Soil Biol. Biochem. 2014, 69, 147–156. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Hobley, E.U.; Wilson, B. The depth distribution of organic carbon in the soils of eastern Australia. Ecosphere 2016, 7, e01214. [Google Scholar] [CrossRef] [Green Version]
- Bonan, G.B.; Hartman, M.D.; Parton, W.J.; Wieder, W.R. Evaluating litter decomposition in earth system models with long-term litterbag experiments: An example using the Community Land Model version 4 (CLM4). Glob. Change Biol. 2013, 19, 957–974. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, H.C.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Van Bodegom, P. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Duboc, O.; Zehetner, F.; Djukic, I.; Tatzber, M.; Berger, T.W.; Gerzabek, M.H. Decomposition of European beech and black pine foliar litter along an Alpine elevation gradient: Mass loss and molecular characteristics. Geoderma 2012, 189, 522–531. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Schrumpf, M.; Kaiser, K.; Guggenberger, G.; Persson, T.; Kögel-Knabner, I.; Schulze, E.-D. Storage and stability of organic carbon in soils as related to depth, occlusion within aggregates, and attachment to minerals. Biogeosciences 2013, 10, 1675–1691. [Google Scholar] [CrossRef] [Green Version]
- Bosatta, E.; Ågren, G.I. Soil organic matter quality interpreted thermodynamically. Soil Biol. Biochem. 1999, 31, 1889–1891. [Google Scholar] [CrossRef]
- Hicks Pries, C.E.; Sulman, B.N.; West, C.; O’Neill, C.; Poppleton, E.; Porras, R.C.; Castanha, C.; Zhu, B.; Wiedemeier, D.B.; Torn, M.S. Root litter decomposition slows with soil depth. Soil Biol. Biochem. 2018, 125, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakov, Y. Priming effects: Interactions between living and dead organic matter. Soil Biol. Biochem. 2010, 42, 1363–1371. [Google Scholar] [CrossRef]
- St-Luce, M.; Ziadi, N.; Chantigny, M.H.; Braun, J. Long-term effects of tillage and nitrogen fertilization on soil C and N fractions in a corn–soybean rotation. Can. J. Soil Sci. 2022, 102, 277–292. [Google Scholar] [CrossRef]
- Andlar, M.; Rezić, T.; Marđetko, N.; Kracher, D.; Ludwig, R.; Šantek, B. Lignocellulose degradation: An overview of fungi and fungal enzymes involved in lignocellulose degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Steinweg, J.M.; Jagadamma, S.; Frerichs, J.; Mayes, M.A. Activation energy of extracellular enzymes in soils from different biomes. PLoS ONE 2013, 8, e59943. [Google Scholar] [CrossRef] [PubMed]
- Parent, L.E.; Mackenzie, A.F. Rate of Pyrophosphate Hydrolysis in Organic Soils. Can. J. Soil Sci. 1985, 65, 497–506. [Google Scholar] [CrossRef]
- Fierer, N.; Craine, J.M.; McLauchlan, K.K.; Schimel, J.P. Litter quality and the temperature sensitivity of decomposition. Ecology 2005, 86, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Gerke, J. Concepts and misconceptions of humic substances as the stable part of soil organic matter: A review. Agronomy 2018, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Meyer, N.; Welp, G.; Amelung, W. The temperature sensitivity (Q10) of soil respiration: Controlling factors and spatial prediction at regional scale based on environmental soil classes. Glob. Biogeochem. Cycl. 2018, 32, 306–323. [Google Scholar] [CrossRef]
- Ignace, D.D. Determinants of temperature sensitivity of soil respiration with the decline of a foundation species. PLoS ONE 2019, 14, e0223566. [Google Scholar] [CrossRef] [Green Version]
- Meyer, N.; Meyer, H.; Welp, G.; Amelung, W. Soil respiration and its temperature sensitivity (Q10): Rapid acquisition using mid-infrared spectroscopy. Geoderma 2018, 323, 31–40. [Google Scholar] [CrossRef]
- Ward, E.B.; Duguid, M.C.; Kuebbing, S.E.; Lendemer, J.C.; Warren II, R.J.; Bradford, M.A. Ericoid mycorrhizal shrubs alter the relationship between tree mycorrhizal dominance and soil carbon and nitrogen. J. Ecol. 2021, 109, 3524–3540. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dossou-Yovo, W.; Parent, S.-É.; Ziadi, N.; Parent, L.E. CO2 Emissions in Layered Cranberry Soils under Simulated Warming. Soil Syst. 2023, 7, 3. https://doi.org/10.3390/soilsystems7010003
Dossou-Yovo W, Parent S-É, Ziadi N, Parent LE. CO2 Emissions in Layered Cranberry Soils under Simulated Warming. Soil Systems. 2023; 7(1):3. https://doi.org/10.3390/soilsystems7010003
Chicago/Turabian StyleDossou-Yovo, Wilfried, Serge-Étienne Parent, Noura Ziadi, and Léon E. Parent. 2023. "CO2 Emissions in Layered Cranberry Soils under Simulated Warming" Soil Systems 7, no. 1: 3. https://doi.org/10.3390/soilsystems7010003