

Assessment of the Risk of Exotic Zika Virus Strain Transmission by Aedes aegypti and Culex quinquefasciatus from Senegal Compared to a Native Strain

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
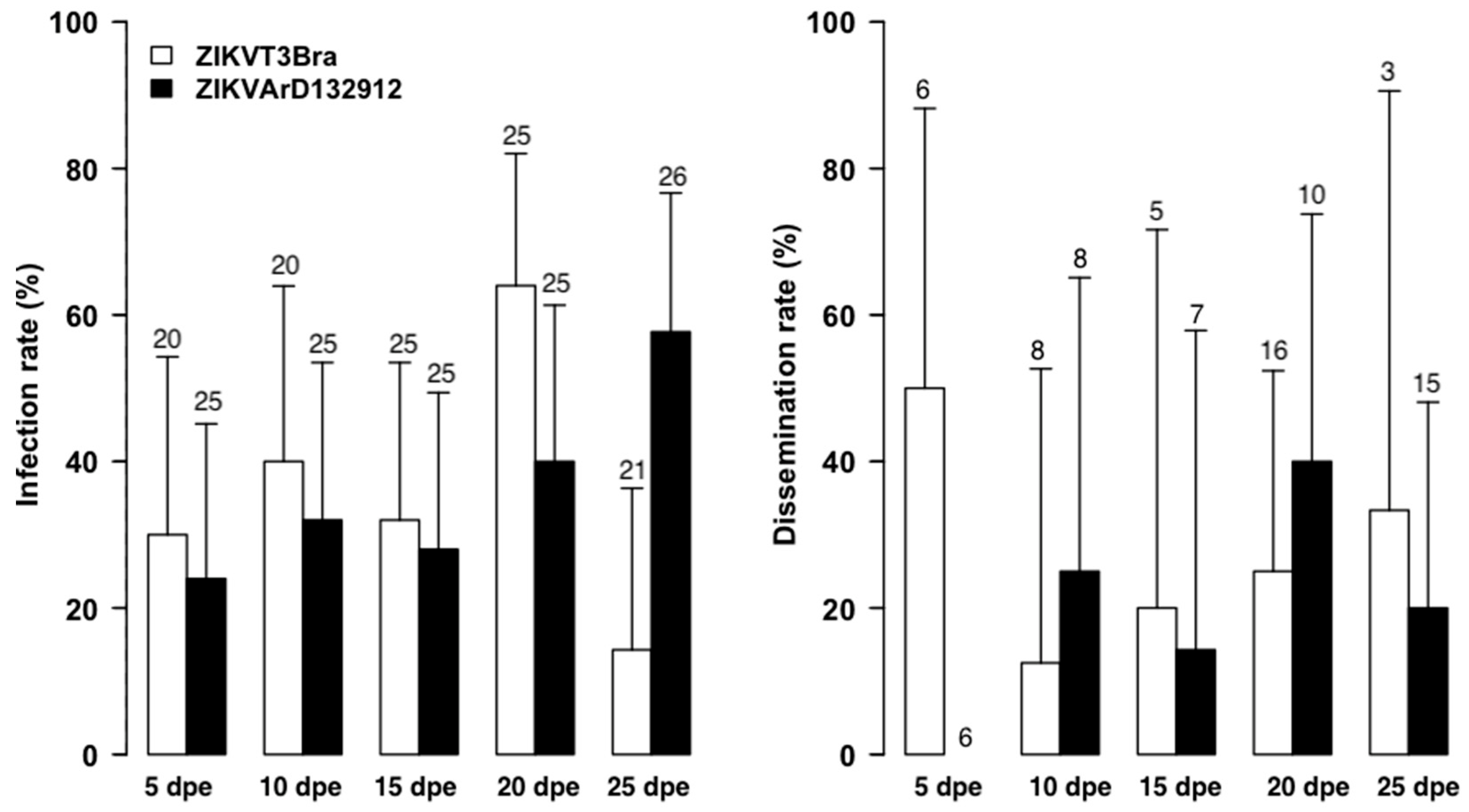
3. Results
3.1. The Zika Senegalese Strain versus Exotic Zika Virus Strains
3.2. Infection by Freshly Harvested versus Frozen ZIKV Stocks
3.3. Vertical Transmission
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dick, G.W.; Cocina, S.F.; Haddow, A.J. Zika Virus. I. Isolations and Serological Specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Simpson, D.I.H. Zika Virus Infection in Man. Trans. R. Soc. Trop. Med. Hyg. 1964, 58, 335–338. [Google Scholar] [CrossRef]
- Beaver, J.T.; Lelutiu, N.; Habib, R.; Skountzou, I. Evolution of Two Major Zika Virus Lineages: Implications for Pathology, Immune Response, and Vaccine Development. Front. Immunol. 2018, 9, 1640. [Google Scholar] [CrossRef] [Green Version]
- Duffy, M.R.; Chen, T.-H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef]
- Grard, G.; Caron, M.; Mombo, I.M.; Nkoghe, D.; Ondo, S.M.; Jiolle, D.; Fontenille, D.; Paupy, C.; Leroy, E.M. Zika Virus in Gabon (Central Africa)—2007: A New Threat from Aedes Albopictus? PLoS Negl. Trop. Dis. 2014, 8, e2681. [Google Scholar] [CrossRef] [Green Version]
- Cao-Lormeau, V.-M.; Roche, C.; Teissier, A.; Robin, E.; Berry, A.-L.; Mallet, H.-P.; Sall, A.A.; Musso, D. Zika Virus, French Polynesia, South Pacific, 2013. Emerg. Infect. Dis. 2014, 20, 1085–1086. [Google Scholar] [CrossRef] [PubMed]
- Campos, G.S.; Bandeira, A.C.; Sardi, S.I. Zika Virus Outbreak, Bahia, Brazil. Emerg. Infect. Dis. 2015, 21, 1885–1886. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C. Prediction and Prevention of Urban Arbovirus Epidemics: A Challenge for the Global Virology Community. Antivir. Res. 2018, 156, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-K.; Lim, J.K.; Yoon, I.-K. An Update on Zika Virus in Asia. Infect. Chemother. 2017, 49, 91–100. [Google Scholar] [CrossRef]
- Spiteri, G.; Sudre, B.; Septfons, A.; Beauté, J.; European Zika Surveillance Network. Surveillance of Zika Virus Infection in the EU/EEA, June 2015 to January 2017. Eurosurveillance 2017, 22, 17. [Google Scholar] [CrossRef] [Green Version]
- Pyke, A.T.; Moore, P.R.; Hall-Mendelin, S.; McMahon, J.L.; Harrower, B.J.; Constantino, T.R.; van den Hurk, A.F. Isolation of Zika Virus Imported from Tonga into Australia. PLoS Curr. 2016, 8. [Google Scholar] [CrossRef]
- Hashimoto, T.; Kutsuna, S.; Tajima, S.; Nakayama, E.; Maeki, T.; Taniguchi, S.; Lim, C.-K.; Katanami, Y.; Takeshita, N.; Hayakawa, K.; et al. Importation of Zika Virus from Vietnam to Japan, November 2016. Emerg. Infect. Dis. 2017, 23, 1223–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diallo, D.; Sall, A.A.; Diagne, C.T.; Faye, O.; Faye, O.; Ba, Y.; Hanley, K.A.; Buenemann, M.; Weaver, S.C.; Diallo, M. Zika Virus Emergence in Mosquitoes in Southeastern Senegal, 2011. PLoS ONE 2014, 9, e109442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagbami, A.H. Zika Virus Infections in Nigeria: Virological and Seroepidemiological Investigations in Oyo State. J. Hyg. 1979, 83, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diallo, D.; Dia, I.; Diagne, C.; Gaye, A.; Diallo, M. Emergences of Chikungunya and Zika in Africa. In Chikungunya and Zika Viruses; Academic Press: Cambridge, MA, USA, 2018; pp. 87–133. ISBN 978-0-12-811865-8. [Google Scholar]
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, Chikungunya, and Other Emerging Vector-Borne Viral Diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef]
- Aubry, F.; Dabo, S.; Manet, C.; Filipović, I.; Rose, N.H.; Miot, E.F.; Martynow, D.; Baidaliuk, A.; Merkling, S.H.; Dickson, L.B.; et al. Enhanced Zika Virus Susceptibility of Globally Invasive Aedes Aegypti Populations. Science 2020, 370, 991–996. [Google Scholar] [CrossRef]
- Rose, N.; Dabo, S.; Leal, S.; Sylla, M.; Diagne, C.; Faye, O.; Faye, O.; Sall, A.; Mcbride, C.; Lambrechts, L. Enhanced Mosquito Vectorial Capacity Underlies the Cape Verde Zika Epidemic. PLOS Biol. 2022, 20, e3001864. [Google Scholar] [CrossRef]
- Diagne, C.T.; Diallo, D.; Faye, O.; Ba, Y.; Faye, O.; Gaye, A.; Dia, I.; Faye, O.; Weaver, S.C.; Sall, A.A.; et al. Potential of Selected Senegalese Aedes Spp. Mosquitoes (Diptera: Culicidae) to Transmit Zika Virus. BMC Infect. Dis. 2015, 15, 492. [Google Scholar] [CrossRef] [Green Version]
- Faye, O.; Freire, C.C.M.; Iamarino, A.; Faye, O.; de Oliveira, J.V.C.; Diallo, M.; Zanotto, P.M.A.; Sall, A.A. Molecular Evolution of Zika Virus during Its Emergence in the 20th Century. PLoS Negl. Trop. Dis. 2014, 8, e2636. [Google Scholar] [CrossRef] [Green Version]
- Zanluca, C.; Melo, V.; Mosimann, A.; Santos, G.; Santos, C.; Luz, K. First Report of Autochthonous Transmission of Zika Virus in Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 569–572. [Google Scholar] [CrossRef]
- Buathong, R.; Hermann, L.; Thaisomboonsuk, B.; Rutvisuttinunt, W.; Klungthong, C.; Chinnawirotpisan, P.; Manasatienkij, W.; Nisalak, A.; Fernandez, S.; Yoon, I.-K.; et al. Detection of Zika Virus Infection in Thailand, 2012–2014. Am. J. Trop. Med. Hyg. 2015, 93, 380–383. [Google Scholar] [CrossRef] [Green Version]
- Faye, O.; de Lourdes Monteiro, M.; Vrancken, B.; Prot, M.; Lequime, S.; Diarra, M.; Ndiaye, O.; Valdez, T.; Tavarez, S.; Ramos, J.; et al. Genomic Epidemiology of 2015-2016 Zika Virus Outbreak in Cape Verde. Emerg. Infect. Dis. 2020, 26, 1084–1090. [Google Scholar] [CrossRef]
- Guedes, D.R.; Paiva, M.H.; Donato, M.M.; Barbosa, P.P.; Krokovsky, L.; Rocha, S.W.D.S.; Saraiva, K.L.; Crespo, M.M.; Rezende, T.M.; Wallau, G.L.; et al. Zika Virus Replication in the Mosquito Culex Quinquefasciatus in Brazil. Emerg. Microbes Infect. 2017, 6, e69. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Li, C.; Deng, Y.; Xing, D.; Liu, Q.; Wu, Q.; Sun, A.; Dong, Y.; Cao, W.; Qin, C.; et al. Culex Pipiens Quinquefasciatus: A Potential Vector to Transmit Zika Virus. Emerg. Microbes Infect. 2016, 5, e102. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-J.S.; Ayers, V.B.; Lyons, A.C.; Unlu, I.; Alto, B.W.; Cohnstaedt, L.W.; Higgs, S.; Vanlandingham, D.L. Culex Species Mosquitoes and Zika Virus. Vector Borne Zoonotic Dis. Larchmt. N 2016, 16, 673–676. [Google Scholar] [CrossRef]
- Amraoui, F.; Atyame-Nten, C.; Vega-Rúa, A.; Lourenço-de-Oliveira, R.; Vazeille, M.; Failloux, A.B. Culex Mosquitoes Are Experimentally Unable to Transmit Zika Virus. Eurosurveillance 2016, 21, 30333. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.S.; Campos, S.S.; Ferreira-de-Brito, A.; Miranda, R.M.D.; Silva, K.A.B.D.; Castro, M.G.D.; Raphael, L.M.S.; Brasil, P.; Failloux, A.-B.; Bonaldo, M.C.; et al. Culex Quinquefasciatus from Rio de Janeiro Is Not Competent to Transmit the Local Zika Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliota, M.T.; Peinado, S.A.; Osorio, J.E.; Bartholomay, L.C. Culex Pipiens and Aedes Triseriatus Mosquito Susceptibility to Zika Virus. Emerg. Infect. Dis. 2016, 22, 1857–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thangamani, S.; Huang, J.; Hart, C.E.; Guzman, H.; Tesh, R.B. Vertical Transmission of Zika Virus in Aedes Aegypti Mosquitoes. Am. J. Trop. Med. Hyg. 2016, 95, 1169–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaty, B.J.; Tesh, R.B.; Aitken, T.H. Transovarial Transmission of Yellow Fever Virus in Stegomyia Mosquitoes. Am. J. Trop. Med. Hyg. 1980, 29, 125–132. [Google Scholar] [CrossRef]
- Rosen, L.; Shroyer, D.A.; Tesh, R.B.; Freier, J.E.; Lien, J.C. Transovarial Transmission of Dengue Viruses by Mosquitoes: Aedes Albopictus and Aedes Aegypti. Am. J. Trop. Med. Hyg. 1983, 32, 1108–1119. [Google Scholar] [CrossRef] [PubMed]
- Baqar, S.; Hayes, C.G.; Murphy, J.R.; Watts, D.M. Vertical Transmission of West Nile Virus by Culex and Aedes Species Mosquitoes. Am. J. Trop. Med. Hyg. 1993, 48, 757–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weger-Lucarelli, J.; Rückert, C.; Chotiwan, N.; Nguyen, C.; Luna, S.M.G.; Fauver, J.R.; Foy, B.D.; Perera, R.; Black, W.C.; Kading, R.C.; et al. Vector Competence of American Mosquitoes for Three Strains of Zika Virus. PLoS Negl. Trop. Dis. 2016, 10, e0005101. [Google Scholar] [CrossRef] [Green Version]
- Gaye, A.; Diagne, M.M.; Ndiaye, E.H.; Dior Ndione, M.H.; Faye, M.; Talla, C.; Fall, G.; Ba, Y.; Diallo, D.; Dia, I.; et al. Vector Competence of Anthropophilic Mosquitoes for a New Mesonivirus in Senegal. Emerg. Microbes Infect. 2020, 9, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Faye, O.; Faye, O.; Diallo, D.; Diallo, M.; Weidmann, M.; Sall, A.A. Quantitative Real-Time PCR Detection of Zika Virus and Evaluation with Field-Caught Mosquitoes. Virol. J. 2013, 10, 311. [Google Scholar] [CrossRef] [Green Version]
- R: A Language and Environment for Statistical Computing. Available online: https://www.gbif.org/fr/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 4 September 2021).
- Vazeille, M.; Yébakima, A.; Lourenço-de-Oliveira, R.; Andriamahefazafy, B.; Correira, A.; Rodrigues, J.M.; Veiga, A.; Moreira, A.; Leparc-Goffart, I.; Grandadam, M.; et al. Oral Receptivity of Aedes Aegypti from Cape Verde for Yellow Fever, Dengue, and Chikungunya Viruses. Vector Borne Zoonotic Dis. Larchmt. N 2013, 13, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Chevillon, C.; Albright, R.G.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Scott, T.W. Genetic Specificity and Potential for Local Adaptation between Dengue Viruses and Mosquito Vectors. BMC Evol. Biol. 2009, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Li, M.I.; Wong, P.S.J.; Ng, L.C.; Tan, C.H. Oral Susceptibility of Singapore Aedes (Stegomyia) Aegypti (Linnaeus) to Zika Virus. PLoS Negl. Trop. Dis. 2012, 6, e1792. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.-S.J.; Li, M.I.; Chong, C.-S.; Ng, L.-C.; Tan, C.-H. Aedes (Stegomyia) Albopictus (Skuse): A Potential Vector of Zika Virus in Singapore. PLoS Negl. Trop. Dis. 2013, 7, e2348. [Google Scholar] [CrossRef]
- Gaye, A.; Faye, O.; Diagne, C.T.; Faye, O.; Diallo, D.; Weaver, S.C.; Sall, A.A.; Diallo, M. Oral Susceptibility of Aedes Aegypti (Diptera: Culicidae) from Senegal for Dengue Serotypes 1 and 3 Viruses. Trop. Med. Int. Health TM IH 2014, 19, 1355–1359. [Google Scholar] [CrossRef]
- Cheng, G.; Liu, Y.; Wang, P.; Xiao, X. Mosquito Defense Strategies against Viral Infection. Trends Parasitol. 2016, 32, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes Aegypti Toll Pathway Controls Dengue Virus Infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef]
- Saldaña, M.A.; Etebari, K.; Hart, C.E.; Widen, S.G.; Wood, T.G.; Thangamani, S.; Asgari, S.; Hughes, G.L. Zika Virus Alters the MicroRNA Expression Profile and Elicits an RNAi Response in Aedes Aegypti Mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0005760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roundy, C.M.; Azar, S.R.; Rossi, S.L.; Huang, J.H.; Leal, G.; Yun, R.; Fernandez-Salas, I.; Vitek, C.J.; Paploski, I.A.D.; Kitron, U.; et al. Variation in Aedes Aegypti Mosquito Competence for Zika Virus Transmission. Emerg. Infect. Dis. 2017, 23, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Lequime, S.; Dehecq, J.-S.; Matheus, S.; de Laval, F.; Almeras, L.; Briolant, S.; Fontaine, A. Modeling Intra-Mosquito Dynamics of Zika Virus and Its Dose-Dependence Confirms the Low Epidemic Potential of Aedes Albopictus. PLoS Pathog. 2020, 16, e1009068. [Google Scholar] [CrossRef]
- Miller, B.R.; Mitchell, C.J. Genetic Selection of a Flavivirus-Refractory Strain of the Yellow Fever Mosquito Aedes Aegypti. Am. J. Trop. Med. Hyg. 1991, 45, 399–407. [Google Scholar] [CrossRef]
- Tabachnick, W.J.; Wallis, G.P.; Aitken, T.H.G.; Miller, B.R.; Amato, G.D.; Lorenz, L.; Powell, J.R.; Beaty, B.J. Oral Infection of Aedes Aegypti with Yellow Fever Virus: Geographic Variation and Genetic Considerations. Am. J. Trop. Med. Hyg. 1985, 34, 1219–1224. [Google Scholar] [CrossRef]
- Bosio, C.F.; Beaty, B.J.; Black, W.C. Quantitative Genetics of Vector Competence for Dengue-2 Virus in Aedes Aegypti. Am. J. Trop. Med. Hyg. 1998, 59, 965–970. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Zika Strain | Blood-Meal Titers (PFU/mL) | Infection Rates | ||
|---|---|---|---|---|
| 5 dpe | 10 dpe | 15 dpe | ||
| ZIKVT3FBra | 2.5 × 104 | 0/30 | 0/30 | 0/37 |
| ZIKVArD132912 | 5 × 106 | 0/20 | 0/20 | 0/28 |
| ZIKVNC | 3.75 × 106 | 0/30 | 0/30 | 0/43 |
| Zika Strains Freshly Harvested from Cells | Blood-Meal Titers (PFU/mL) | Infection Rates | ||
|---|---|---|---|---|
| 15 dpe | 20 dpe | 25 dpe | ||
| ZIKVT3FBra | 7.5 × 104 | 0/35 | 0/35 | 0/40 |
| ZIKVArD132912 | 3 × 104 | 0/30 | 0/30 | 0/25 |
| Zika Strains Frozen a Week | Blood-Meal Titers (PFU/mL) | Infections Rates | |||
|---|---|---|---|---|---|
| 10 dpe | 15 dpe | 20 dpe | 25 dpe | ||
| ZIKVT3FBra | 1.35 × 105 | 0/30 | 0/30 | 0/30 | 0/35 |
| ZIKVArD132912 | 3 × 104 | 0/25 | 0/25 | 0/25 | 0/36 |
| N° Batch | Infection Rates (%) | |
|---|---|---|
| ZikV T3 F BRA | ZikV ArD 132912 | |
| Batch 1 | 3/18 (16.6) | 12/13 (92.3) |
| Batch 2 | 0/15 (00) | 9/15 (60) |
| Batch 3 | 4/14 (28.5) | 11/13 (84.6) |
| Batch 4 | 2/6 (33.3) | 7/9 (77.7) |
| Batch 5 | 3/10 (30) | 11/13 (84.6) |
| Separated females | NA | 20/22 (90.9) |
| Total | 12/63 (19.04%) | 70/85 (82.35%) |
| Days Post-Emergence | Infection Rates of Offspring from Infected Females (%) | ||
|---|---|---|---|
| Eggs from Females in Batches | Eggs from Separated Females | ||
| ZIKV T3FBra | ZIKV ARD132912 | ZIKV ARD132912 | |
| 0 | 0/20 (00) | 0/20 (00) | 0/10 (00) |
| 5 | 0/20 (00) | 0/20 (00) | 0/15 (00) |
| 10 | 0/40 (00) | 0/40 (00) | 0/15 (00) |
| 15 | 0/30 (00) | 0/35 (00) | 0/18 (00) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaye, A.; Fall, C.; Faye, O.; Dupont-Rouzeyrol, M.; Ndiaye, E.H.; Diallo, D.; de Andrade Zanotto, P.M.; Dia, I.; Weaver, S.C.; Diallo, M. Assessment of the Risk of Exotic Zika Virus Strain Transmission by Aedes aegypti and Culex quinquefasciatus from Senegal Compared to a Native Strain. Trop. Med. Infect. Dis. 2023, 8, 130. https://doi.org/10.3390/tropicalmed8020130
Gaye A, Fall C, Faye O, Dupont-Rouzeyrol M, Ndiaye EH, Diallo D, de Andrade Zanotto PM, Dia I, Weaver SC, Diallo M. Assessment of the Risk of Exotic Zika Virus Strain Transmission by Aedes aegypti and Culex quinquefasciatus from Senegal Compared to a Native Strain. Tropical Medicine and Infectious Disease. 2023; 8(2):130. https://doi.org/10.3390/tropicalmed8020130
Chicago/Turabian StyleGaye, Alioune, Cheikh Fall, Oumar Faye, Myrielle Dupont-Rouzeyrol, El Hadji Ndiaye, Diawo Diallo, Paolo Marinho de Andrade Zanotto, Ibrahima Dia, Scott C. Weaver, and Mawlouth Diallo. 2023. "Assessment of the Risk of Exotic Zika Virus Strain Transmission by Aedes aegypti and Culex quinquefasciatus from Senegal Compared to a Native Strain" Tropical Medicine and Infectious Disease 8, no. 2: 130. https://doi.org/10.3390/tropicalmed8020130