1. Introduction
Histological studies of developing embryos have played a critical role in embryology and developmental biology fields, uncovering cellular and tissue mechanisms of morphogenesis [
1,
2,
3] and providing detailed descriptions of the developmental features of different species [
4,
5]. Light microscopy of tissue sections is also an important tool in correlated fields, such as teratology. Cell and tissue shape and structure may predict function; thus, morphological examinations can identify alterations and explain dysfunctions and diseases. This makes histological observation a valuable device for a more detailed characterization of teratogenicity [
6]. Due to technological advances in microscopy, it is possible to analyze microscopic images in an accurate, objective, repeatable, and quantitative manner, free from both the limits of human vision and the subjectivity of the observer, through computational microscopy [
7]. Notwithstanding its importance, few studies discuss the application of histological assessment in embryology, developmental biology, and associated areas, as well as adaptations in methodologies in favor of the particularities of the embryo specimen.
The chicken (
Gallus gallus) embryo is a funded animal model that provides a great system for studies in embryology, developmental biology, and evolutionary developmental biology (evo-devo) [
8,
9,
10]. Additionally, it has been used for developmental toxicity studies [
11,
12,
13]. These embryos develop quickly, are easily experimentally manipulated, and offer good cost effectiveness [
14]. Furthermore, the chicken embryo development is molecularly and morphologically similar to other vertebrates, most notably during the phylotypic period [
15,
16,
17]. However, despite these advantages, working with embryos is a challenge because of their fragility and reduced size, mostly at the early stages of chick embryos (up to 4 days of incubation). These features make it difficult to obtain proper histological sections with excellent tissue preservation and correct orientation.
The scientific literature demonstrates that several embedding media have been used for routine histology [
18] of animal adult tissues and plant samples, such as paraffin-based resins. However, embryo tissues require more delicate handling and routine procedures usually do not maintain proper preservation of structure and cellular detail [
19]. Embedding media such as polyethylene glycol (PEG) and glycol methacrylate (GMA)-based resins are potential alternatives for embryo histology due to their properties and processing protocols. However, there are no reports in the literature on the use of PEG for the embedding of early chick embryos. Additionally, the application of glycol-methacrylate-based resins is underexplored in this area.
Paraffin is the most common embedding medium for histology. Its usual commercially available formulations are mixtures of long-chain alkanes and plastic polymers, which melt at 56–58 °C [
20]. Paraplast is a commercial resin, composed of highly purified paraffin and plastic polymers, which provides quality sections and is compatible with most routine stains and immunohistochemistry protocols, which melt at 56–57 °C [
21,
22]. Its formulation is known as Paraplast Plus when dimethyl sulfoxide (DMSO) is present, which offers faster infiltration times and favors sectioning (thickness down to 2–4 μm) [
23]. However, due to its insolubility in water, its processing requires transitional solvents such as xylene before infiltration of the tissues [
24]. This step often hardens and shrinks the tissues [
25], especially those of a delicate specimen such as early chick embryos, impairing the final result.
Polyethylene glycols are polymers of ethylene oxide, produced under alkaline catalysis. They are known as “PEG” plus a numerical value, which represents its mean molecular weight [
26]. PEG is a water- and alcohol-miscible embedding medium commonly used in plant histology [
27] and histochemistry [
28], but also applied to animal tissue histology [
29,
30], immunohistochemistry [
30,
31], enzymatic histochemistry, and histofluorescence [
32]. PEG embedding allows a wide range of section thicknesses (1–150 μm) which can be obtained by varying the molecular weights of the PEGs used [
33]. Nevertheless, its use as a single embedding media of animal tissues is poorly discussed and there are no recent studies on this topic. Its low acceptance happened probably because of problems during sectioning at high ambient humidity, and in mounting [
33], due to PEG’s hygroscopic properties [
34]. Additionally, there is no study on its applications for the histology of early stages of embryos, or immunolabeling, although its use seems promising due to the gentler processing when compared with paraffin-based resins. Among the advantages over paraplast is its solubility in water, excluding the need for dehydration for its infiltration into the tissue, in addition to a lower melting temperature.
GMA-based resins, such as historesin, are hydrophilic embedding matrixes that provide good morphological preservation because their protocols do not require clearing agents and high infiltration temperatures [
35]. Several mixtures have been described and different commercial kits are available, in which the proportion of the monomer and other components may vary [
36]. Embedding with GMA-based resins allows a wide range of section thicknesses (0.5–5 μm), depending on the embedding medium and knife used [
36]. Thus, different levels of cell detail, contrast, and sharpness may be explored [
37]. Additionally, GMA does not react with chemical groups in the tissue, which is important for staining methods. Thus, they are compatible with many histological and histochemical protocols used for paraffin sections with some modifications [
38].
To date, there is little information in the literature regarding the ideal method for obtaining precise tissue orientation and optimal morphological preservation of early chick embryo sections. To fill this gap, in this work we tested three different embedding resins (paraplast, polyethylene glycol, and historesin) to find the best protocol for the histology of early stages of chicken embryos. Our findings further the knowledge and optimize the embedding methodology that applies to all areas using the chicken embryo as a model.
2. Materials and Methods
2.1. Animals
Fertilized unincubated
Gallus gallus eggs were provided by a hatchery in Curitiba, State of Parana (PR), Brazil. A total of approximately 50 eggs were used, from at least three different batches. All procedures were approved by the Animal Use Ethics Committee from the Biological Sciences Sector of Federal University of Parana (CEUA/BIO-UFPR, certificate no 1098;
http://www.bio.ufpr.br/portal/ceua/, accessed on 27 February 2023).
2.2. Incubation
Eggs with unbroken shells were cleaned with 70% ethanol and randomly placed in an incubator (Biochemical Oxygen Demand-BOD incubator/SL-224, SOLAB Cientifica, Piracicaba, Brazil ) with air cells facing upwards. The temperature during the incubation period (72 h) was maintained at 38 ± 0.5 °C, with 60% humidity and constant ventilation [
39].
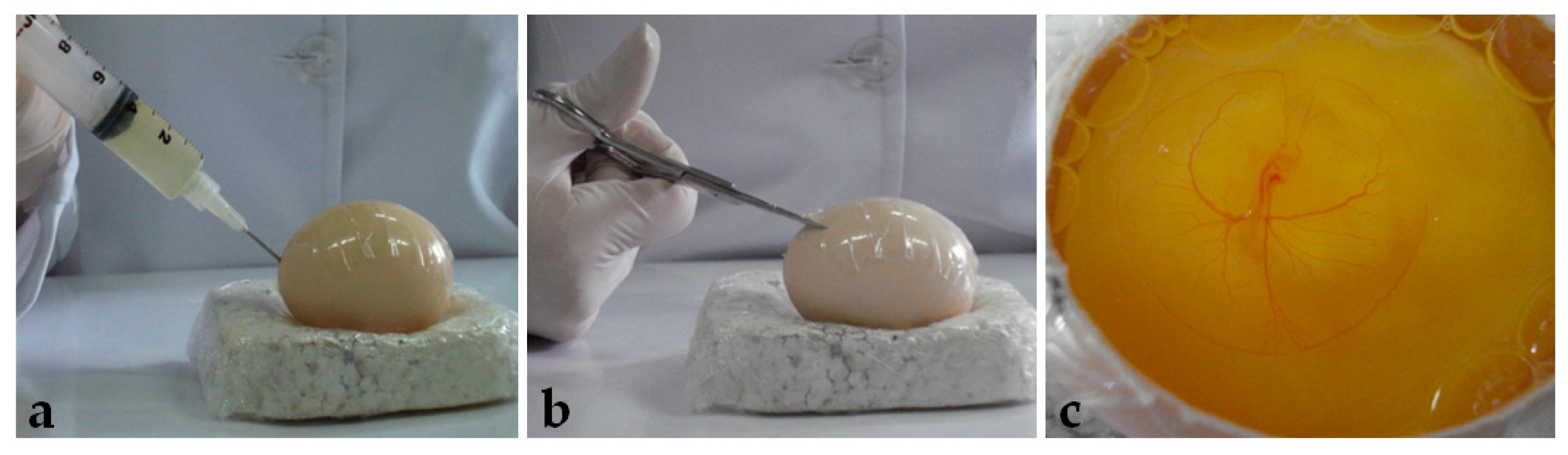
After seventy-two hours of incubation, the eggs were opened using the windowing method described by Korn and Cramer [
40]. Before opening, the egg was turned 90°, so the large surface lied horizontally. After removing the egg from the incubator, adhesive tape was placed on the shell to avoid breaking. Then, 5 mL of albumen was removed with a syringe and needle (
Figure 1a), and, with a scissor (
Figure 1b), a window of approximately 6 cm
2 was opened (
Figure 1c) and embryo viability was determined (
Figure 2).
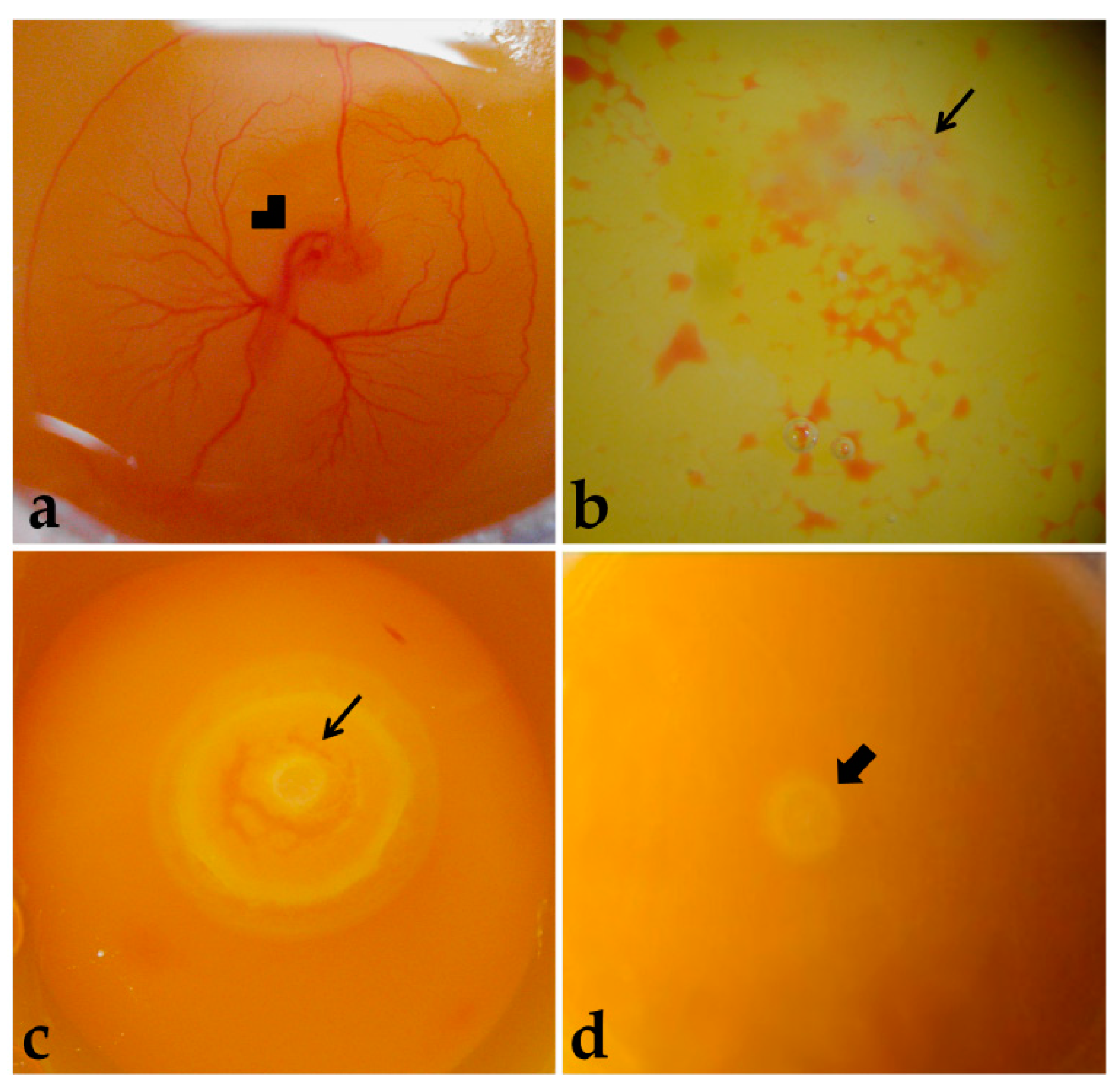
Embryos were considered alive if they were bright rose-colored, presented heartbeats, and had intact extraembryonic blood vessels (
Figure 2a). Dead embryos presented a whitish and opaque vitelline membrane, absence of heartbeats, and non-intact extraembryonic blood vessels (
Figure 2b), or embryonic discs more developed than at laying, meaning that development was resumed with incubation, but was terminated before embryo harvesting (
Figure 2c). Moreover, some embryos were classified as not having resumed development with incubation (
Figure 2d). The dead embryos were discarded.
Live embryos were collected and transferred to a Petri dish with PBS, where their extraembryonic membranes were removed. Then, embryos were fixed in 2% paraformaldehyde (in PBS) for 72 h, in a 24-well plate, for further histological examination.
2.3. Embryo Orientation
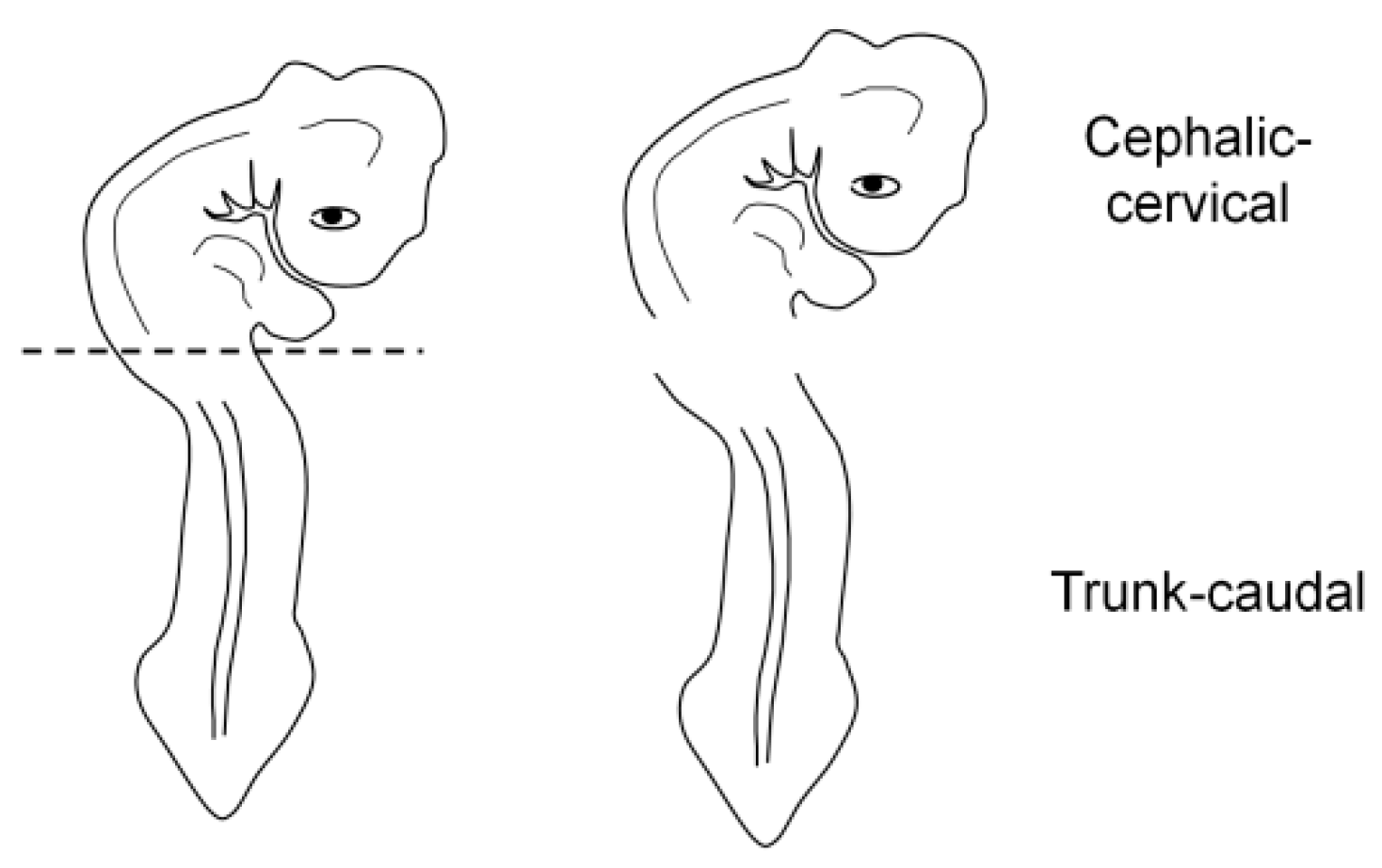
For the correct orientation of the embryo, specimens were embedded as either a whole embryo or a fragment (cephalic-cervical and trunk-caudal region) (
Figure 3). This strategy was adopted due to difficulties at the moment of orientation, as the heavier head tends to sink and elevate the trunk, generating an undesired angle. To obtain the fragments, embryos were separated into regions with a blade, under a stereomicroscope (SZ40, Olympus, Shinjuku, Japan).
2.4. Paraplast
Fixed embryos were washed in PBS for 15 min and dehydrated in 70–95% ethanol, followed by two changes of 100% ethanol, 10–15 min each. The embryos were then cleared in xylene (two changes, 3–5 min each), and infiltrated in Paraplast Plus (Leica Biosystems, Wetzlar, Germany) at 58 °C (three baths, 15–60 min each) [
41]. Then, they were embedded in paraplast and oriented for obtaining transversal sections. Embedding was performed in metallic molds with covering cassettes. Each block contained a whole embryo or an embryo fragment.
Sections of 5 μm were obtained in a semiautomatic microtome (RM 2145, Leica Biosystems, Wetzlar, Germany), using histological disposable steel blades, and placed on glass slides covered with albumin and water. After the distention of the sections on a heated plate, the slides were dried overnight at room temperature. Then, slides were placed on xylene for removing the resin, hydrated in ethanol (100-70%, 3–6 min each) and distilled water, and stained with Harri’s hematoxylin (30 s) and eosin-floxin (30 s) (H&E). Next, sections were dehydrated in ethanol (95% and twice in 100%, 1–3 min each) and ethanol-xylene (1:1—3 min), cleared in xylene (twice for 3 min), and mounted with Permount
TM (Thermo Fisher Scientific, Waltham, EUA) and coverslip. All procedures are detailed in
Table 1 and
Table 2.
As it was difficult to obtain the precise embryo orientation for sectioning by direct incorporation into paraplast, we chose to perform a pre-embedding tissue immobilization in an agar–gelatin solution before paraplast embedding. This protocol was based on the studies of Buzzel [
42], Ghassemifar and Franzén [
43], Jones and Calabresi [
44], and McClelland et al. [
45], with modifications. Stock solutions of agar (4%) (Sigma-Aldrich, San Luis, EUA) and gelatin (Sigma-Aldrich, San Luis, EUA) (5%) in PBS were prepared to obtain a final solution of agar–gelatin (2 or 4%—2.5 or 5%). This solution was optimized, and the final concentrations chosen were agar at 2% and gelatin at 5%.
Fixed embryos were washed in PBS (twice, for 5 min each), then transferred to the agar–gelatin solution on a drop of the solution on a Petri dish (
Figure 4a) [
45] or a polyethylene mold filled with the solution (
Figure 4b), and oriented for obtaining transversal sections. After polymerization, the blocks were removed from the molds (
Figure 4c), trimmed, and the anterior portion of the block was stained with aqueous eosin 1% (30 s). Then, the blocks were placed into identified histological cassettes and stored in ethanol 70% or NaCl 0.9% solution, for at least 24 h. Paraplast processing was performed as described previously in this section, except for the step with xylene that was replaced for amyl acetate in the processing of some blocks, to test which clarification solution would bring better results.
2.5. Polyethylene Glycol
In PEG 1500 (Synth) processing, two methods were tested, with dehydration (method 1) (M1) and without (method 2) (M2), as described by Wolosewick [
46]. First, fixed embryos were washed in PBS (15 min) and then processed with methods 1 or 2. In processing M1 with ethanol and PEG, the embryos were dehydrated in ethanol (25–100%, 10 min each) and infiltrated in two solutions of PEG and 100% ethanol, in the proportions 1:1 and 2:1, for 30 or 60 min each. Then, samples were transferred to 100% PEG (two changes), for 30 or 60 min (
Table 3).
In PEG-only processing (M2), solutions were prepared in distilled water in concentrations (
v/
v) of 25, 50, and 70%. Embryos were infiltrated in these solutions for 15 to 30 min each. Then, they were transferred to 100% PEG (two changes, 15 or 30 min each) (
Table 4).
All processing was performed in a 24-well microplate (histological cassettes can be used as well). At all steps with solutions containing PEG, samples were maintained at 55 °C. After infiltration with PEG, embryos were embedded in paper molds. On a heated plate, the identified mold was filled with PEG and the embryo was properly oriented for obtaining transversal sections. This enables higher polymerization time, especially on cold days, allowing more time for tissue manipulation. Each block contained a whole embryo or an embryo fragment. The blocks were kept in a container with silica, at room temperature or 4 °C for overnight polymerization. Afterward, the blocks were removed from the molds and stored in a container with silica, to avoid humidification and softening. Then, before sectioning, they were trimmed and fixed on wooden supports with previously melted PEG.
Using disposable blades, 5 μm cross sections were produced in a semiautomatic microtome (Leica RM 2145, Leica Biosystems, Wetzlar, Germany). The sections were placed on albumin or chrome gelatin-coated (0.1% or 1%) slides, with or without Triton
TM X-100 (Sigma-Aldrich, San Luis, EUA) solution (0.1% in PBS). After the distention of sections on a heated plate, slides were dried overnight at room temperature. Then, histological sections were hydrated in 70% ethanol and distilled water and stained with Harri’s hematoxylin (30 s) and eosin-floxin (30 s) (
Table 5). After drying, the sections were covered with Permount
TM (Thermo Fisher Scientific, Waltham, EUA, Waltham, MA, USA) and coverslip.
2.6. Historesin
This protocol was based on González Santander et al. [
19], with modifications. The working solution was prepared according to the Historesin Embedding Kit (Leica Biosystems, Wetzlar, Germany) by diluting 5 g of the activator compound in 50 mL of resin (hydroxyethyl methacrylate) and storing at 4 °C. In a 24-well plate, embryos were washed in PBS for 2 h, and dehydrated in ethanol (30%—5 min; 50–90%—10 min; twice in 100%—10 min each). Then, tissues were immersed in pre-infiltration solution (ethanol 100% and working solution, 1:1) for 2 h, and infiltration solution (working solution) for at least 12 h, both at room temperature. Next, each embryo was placed in 600 μL of embedding solution (working solution and hardener compound, 15:1) (
Table 6) in polyethylene molds and orientated to obtain transversal sections (
Figure 5a). To maintain the embryo at the desired position, a cold pack and a heating plate were used to control the initial polymerization time, and tweezers or a wooden pick to hold the embryo in position. Each block was identified and placed in a vacuum desiccator with silica for 48 h for complete polymerization (
Figure 5b). Then, blocks were removed from the molds and stored in a container with silica, to avoid humidification and softening.
For tissue sectioning, two microtomes were used: a semi-automatic one (Leica RM 2145) and a manual one (Spencer 820, Vernon Hills, EUA), to evaluate the best method for holding the block. In the semi-automatic microtome, the block was directly fixed to the support and sectioned with tungsten knives (
Figure 5c). In the manual microtome, the block was fixed to a wooden support with cyanoacrylate glue before being positioned on the microtome and sectioned with non-disposable steel knives (
Figure 5d). Blocks were trimmed until the tissue reached the sectioning surface. Toluidine blue (0.1%) staining was used to confirm the presence of the sample in the sections. After confirmation, sections of 5 μm were obtained, placed on drops of water on clean glass slides, distended, and dried on a heated plate. Then, sections were hydrated in distilled water (3 min) and stained with Harri’s hematoxylin (1–30 min), and aqueous eosin 1% (1–15 min) (
Table 7). After drying, sections were covered with Permount
TM (Thermo Fisher Scientific, Waltham, EUA) and coverslip.
2.7. Analysis
All histological slides were analyzed under a light microscope (Metrimpex Hungary/PZO-Labimex, Studar lab). About 900 slides, including 250 slides of paraplast-only, 150 slides of pre-embedded specimens, 250 slides of PEG samples, and 250 slides of historesin samples. Additionally, about 10,000 sections were analyzed in total. Selected sections were documented under a bright-field photomicroscope (Olympus BX40, DP71 Camera 12.5 megapixels, DPController software, Olympus, Shinjuku, Japan).
The parameters evaluated for considering the best method for embedding early chick embryos were (1) precision of tissue orientation, (2) the preview of the embryos in the blocks, (3) microtomy, (4) contrast in staining, (5) preservation of structures (morphology and presence of artifacts), (6) time, and (7) cost of processing.
Following the examination of the slides, the three embedding methods tested were assigned a score for each evaluated parameter, using an evaluation index developed specifically for this study. The scores ranged from 0 to 5 and are presented in
Table 8. The analysis was qualitative, and the slides were assessed by three observers.
The precision of tissue orientation was categorized from impossible (score = 0) to excellent (score = 5). The scoring of this parameter was comparative between the resins. The assessment of embryo visualization in the blocks was categorized as impossible (score = 0) or possible (score = 1). Microtomy was ranked from impossible (score = 0) to good (score = 3) based on the time required for sectioning a whole embryo and the quality of resulting sections. The scoring of this parameter was also comparative between the resins. As for the contrast in staining, this was considered weak (score = 0) to excellent (score = 5). Here, the differentiation of cytoplasm, nucleus, nucleolus, and mitotic figures was considered. The tissue preservation was categorized by the maintenance of morphology, which was classified from awful (score = 0) to excellent (score = 5), and the presence of artifacts, from highly frequent (score = 0) to not observed (score = 5). In this parameter, the shape and integrity of embryonic cells and structures (ectoderm, neural tube, notochord, somite, and mesoderm) were considered. The methods with the longer time and higher cost received a lower score (score = 0) and the others were scored until the maximum (score = 5), based on their percentage from the higher time or cost.
4. Discussion
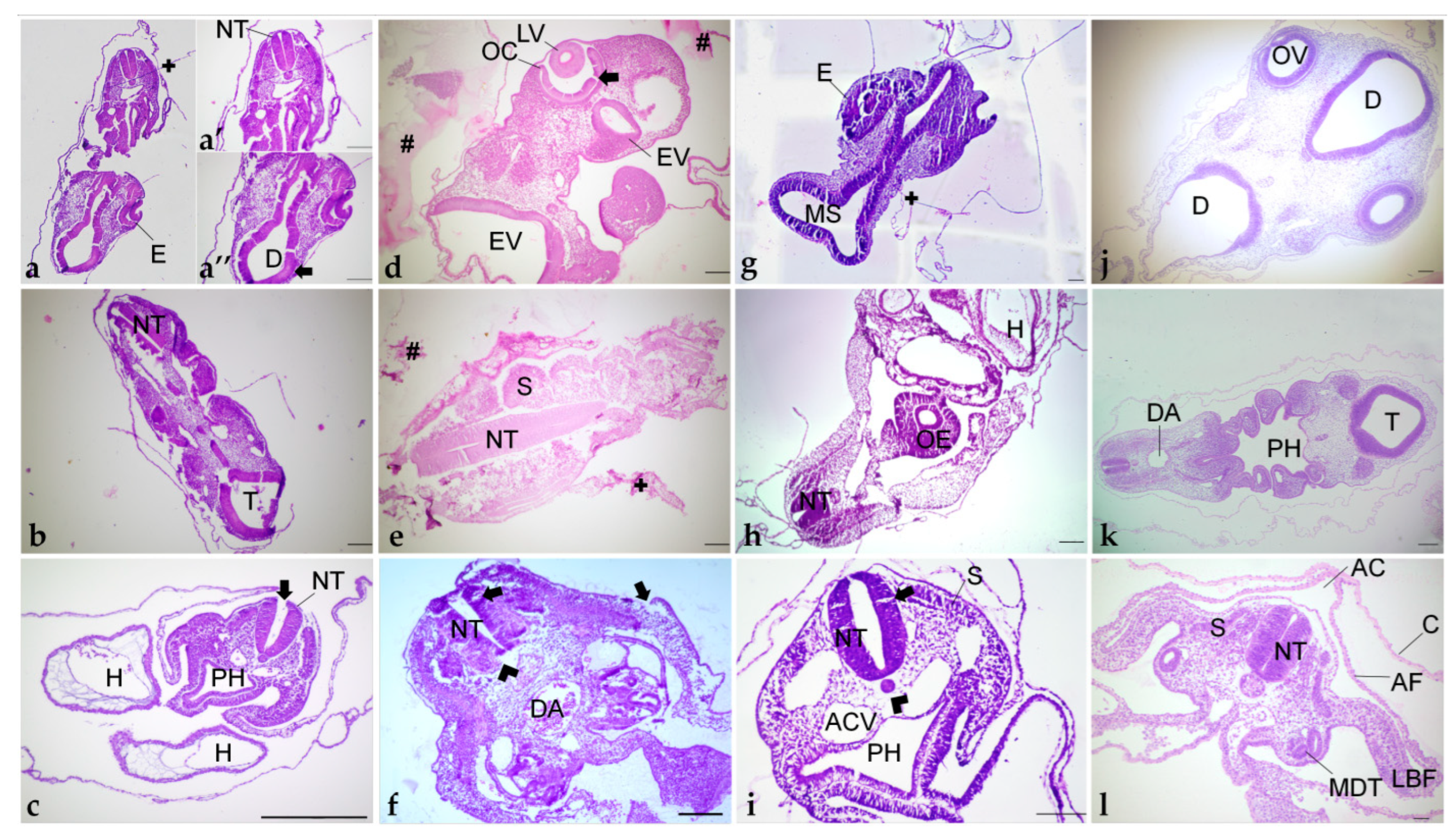
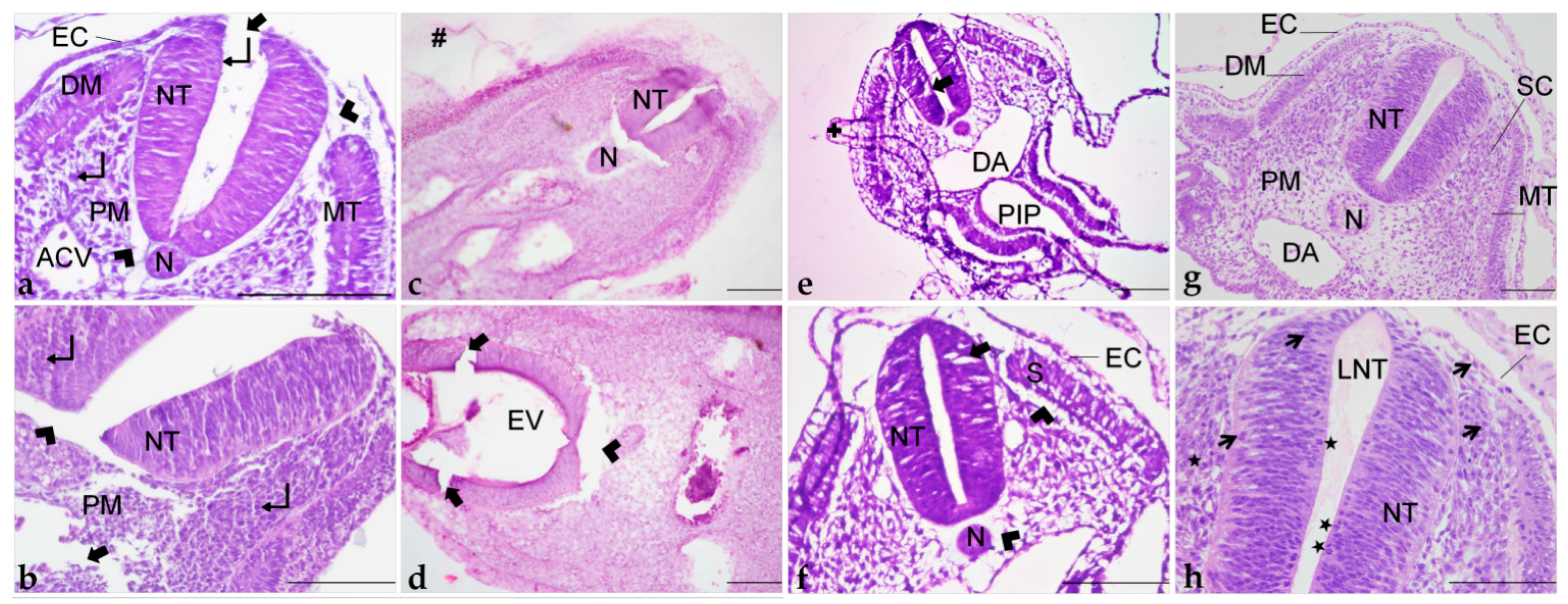
Due to the fragility and reduced size of early chick embryos (up to 4 days of incubation), structural preservation is lost with histological routine processing. Information about the ideal method for this developmental period is still limited. Thus, in this study, we demonstrated how different embedding resins preserve morphological features, such as embryonic structure and cellular details of chick embryo sections.
Tissue orientation is crucial for the demonstration of proper morphology, correct identification of structures, and diagnosis of possible damage [
48,
49,
50], particularly in samples that present different organizations depending on the plane of section, such as embryos. Among the tested resins, paraplast and PEG did not offer appropriate conditions to position the embryo in the mold, due to its quick polymerization which depends only on temperature. Thus, paraplast and PEG embedding require an oven and a heated plate, or embedding station equipment to perform tissue embedding, due to their high melting points. In contrast, historesin allowed embryo positioning in the mold, because it presents a slow polymerization rate at room temperature. This process is dependent on a catalyst (hardener), which reacts in the presence of the activator and generates free radicals that act as polymerization initiators [
51]. The speed of this reaction depends on temperature, which is very high and may turn the block overly hard and brittle, or, if too low, excessively soft [
52]. The ideal conditions for the polymerization of glycol methacrylate (GMA)-based resins, such as historesin, vary greatly according to their composition, such as temperature (from −20 °C to 40 °C), and, in some cases, the need for ultraviolet light and microwaves [
36]. However, there are no studies that describe these conditions for historesin. Another favorable point of historesin was its transparency after polymerization, allowing excellent visualization of the embryo in the block, which facilitates positioning during microtomy. Therefore, it is also used for other small species [
53,
54] or organs [
55,
56]. Despite having different polymerization speeds at room temperature, higher when compared with glycol methacrylate resins, PEG and paraplast allowed similar orientation quality for fragments of the trunk-caudal region.
The pre-embedding in agar–gelatin was found to be an accessible alternative in trying to obtain optimal embryo orientation with paraplast. However, it was not efficient in the processing of our samples. Even after adjustments in the dehydration, clarification, and infiltration times, it did not improve tissue preservation. This fact was not expected, because according to Jones and Calabresi [
44], pre-embedding preparations should not damage tissue processing, as the agar–gelatin solution does not infiltrate into the sample [
42]. Moreover, the use of an agar–gelatin solution should have prevented issues observed when they are used individually, such as sections detaching from slides and poor staining [
44], as was observed in the present study.
Histology is a useful tool in embryology, developmental biology [
3], and correlated fields, including developmental toxicology [
22] and teratology [
57]. Thus, it is clear that the maintenance of morphology is essential to obtain reliable results. During histological processing, embedding matrixes are used to impregnate the tissue, such as paraplast-, PEG-, and GMA-based resins. These matrixes provide support and allow thin sections to be obtained [
21]. As preservation status is the result of the interaction of the specimen with the embedding medium, this parameter will be discussed per resin.
Paraplast did not keep appropriate preservation of chick embryo tissue, and artifacts were frequent. Its tissue processing method requires the use of intermediate solvents because it is not directly soluble in ethanol. This, together with the high temperature needed for infiltration, damaged the fragile samples, and generated artifacts due to tissue hardening and distortion [
58]. These artifacts were accentuated in pre-embedded samples, because the agar–gelatin coat impaired impregnation, which had to be prolonged, resulting in higher shrinkage and crumble. Smith and Warfield [
59] suggested that tissue shrinkage can occur due to the effect of heat on collagen. To reduce these temperature artifacts, Paraplast X-tra could be an alternative, as it has a lower melting point (53–54 °C) [
60]. PEG melts at a similar temperature (55 °C) but also did not yield satisfactory structural preservation.
The use of PEG 1500 did not yield appropriate tissue structural preservation in either method tested. Additionally, method 1 (with dehydration) required about 1 h less than method 2 (without dehydration) to obtain the same section quality. Nevertheless, our work is the first to approach PEG as an embedding medium for early chick embryos and to compare its use with other resins. Despite the low quality of morphology preservation observed in this study, this medium is recommended for immunohistochemical techniques due to the high antigenicity of the tissue sections obtained, including those of soft materials such as early chick embryos [
61]. Additionally, PEG is proper for immunofluorescent labeling, and its preservation of cellular detail is described as superior to frozen sections [
62]. Thus, our data on PEG embedding enrich the limited literature on this matter and may help researchers in exploring its properties in immunodetection techniques.
Acrylic resins are appropriate for embedding fragile, soft, and temperature-sensitive tissues such as embryos, because infiltration and polymerization occur at room temperature, minimizing protein degradation and damage to cellular structures [
36]. Indeed, historesin yielded the best preparation of the embryo sections and high microscopic image resolution, which, according to González Santander et al. [
19], is close to the in vivo state. Furthermore, its hydrophilic properties exclude the need for highly toxic clearing agents [
63], such as xylene, which hardens and shrinks the tissue [
25].
The adhesion of the tissue sections to the slides is crucial to avoid detachment during immunohistochemistry or staining procedures. In some cases, properly cleaned and dried slides are enough. GMA sections attach to slides by heat and water evaporation on a heated plate [
64]. However, paraplast and PEG sections require adhesive coating on slides, such as albumin, gelatin, poly-L-lysine, or 3-aminopropyltriethoxysilane [
65]. Albumin is commonly used with paraplast sectioning, in which adhesion is promoted by heating the slides to 55–60 °C, after placing the sections. This process coagulates the albumin layer, making it insoluble in water and preventing the sections from detaching [
66]. The use of chrome gelatin coating provides charge to the slides, allowing them to attract negatively charged tissue sections [
67]. This method was the most efficient in holding PEG sections to the slides, with a 1% gelatin solution, which provided more cross-linked bonds due to a higher concentration of gelatin. Additionally, Triton X-100 solution increased the adhesion of the sections to the slides, as described by Smithson et al. [
33].
Histological staining is used to enhance tissue features and promote contrast between structures [
68]. Hematoxylins stain nuclear chromatin and other acidic cellular elements [
58]. Among them, Harris’ hematoxylin is the most commonly used combined with eosin [
69] to demonstrate the general histological structure of tissues. In contrast, eosin stains cell cytoplasm and most connective tissue in varying tones and intensities of pink, orange, and red. Among the types of eosins available, eosin Y is the most used and is soluble in ethanol and water [
47], which makes it suitable for GMA processing. Additionally, floxin is often used to provide a darker cytoplasmic color [
58]. In this study, different levels of color and contrast were obtained on the sections of early chick embryos with H&E staining. The nuclear staining was noticeable in the sections of all the used methods, except in those of pre-embedded embryos. On the other hand, eosin staining was less evident in PEG sections. According to Bancroft and Layton [
47], variations in staining may occur due to factors such as fixation, variations in processing steps, section thickness, and excessive hot plate temperatures. Additionally, our results point out that the embedding medium is a factor related to processing that also affects color tones and contrast provided by dyes.
Although only historesin provided excellent tissue preparation for morphological assessment of 72 h chick embryos, the three tested resins have their advantages and disadvantages, depending on the purpose of the study. Thus, when choosing which embedding medium to apply it is important to consider factors such as morphological preservation and antigenicity. Paraplast and PEG are recommended for immunohistochemistry approaches, without the need for precise morphological preservation, and for histological examination of later stages of embryo development, which are less fragile than early stages. GMA-based resins provide precise structure preservation and also allow immunostaining techniques [
70]. Another relevant difference between these three resins is the time of processing. Paraplast and PEG present similar processing times, whereas that of historesin is close to paraplast with pre-embedded samples. Beyond processing, the tissue staining protocol of glycol methacrylate resins is longer as the medium is not previously removed from the sections and slows stain penetration [
71]. However, fewer steps are necessary because ethanol and xylene are not used. In terms of cost, paraplast and PEG resins have a similar price and require the usual equipment of a histology laboratory (stove, embedding station, cassettes, steel blades). In contrast, the historesin kit has a higher cost, as well as the blades, such as tungsten and glass knives, required for sectioning [
36].

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}