Evaluation of 3D/2D Imaging and Image Processing Techniques for the Monitoring of Seed Imbibition

 , , ,
, , , 
Abstract
:1. Introduction
2. 3D X-Ray Imaging of Seed Imbibition in Soil Conditions
2.1. Acquisition Protocol
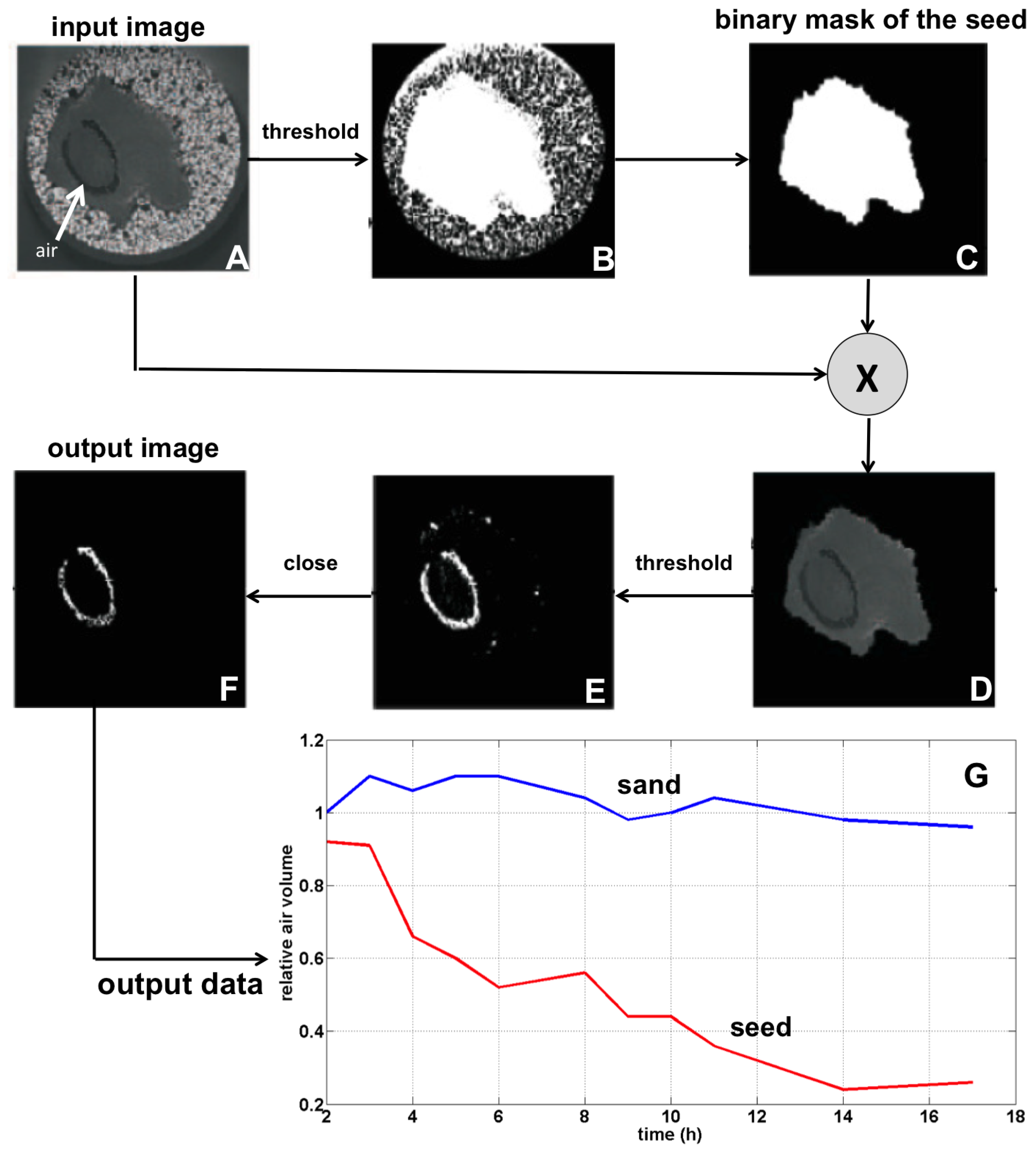
2.2. Image Processing Pipeline
2.3. Illustration
2.4. Interests and Limitations
3. 3D Magnetic Resonance Imaging of Seed Imbibition in Soil Conditions
3.1. Acquisition Protocol
3.2. Image Processing Pipeline
3.3. Illustration
3.4. Interests and Limitations
4. 2D Passive Thermography of Seed Imbibition out of Soil
4.1. Acquisition Protocol
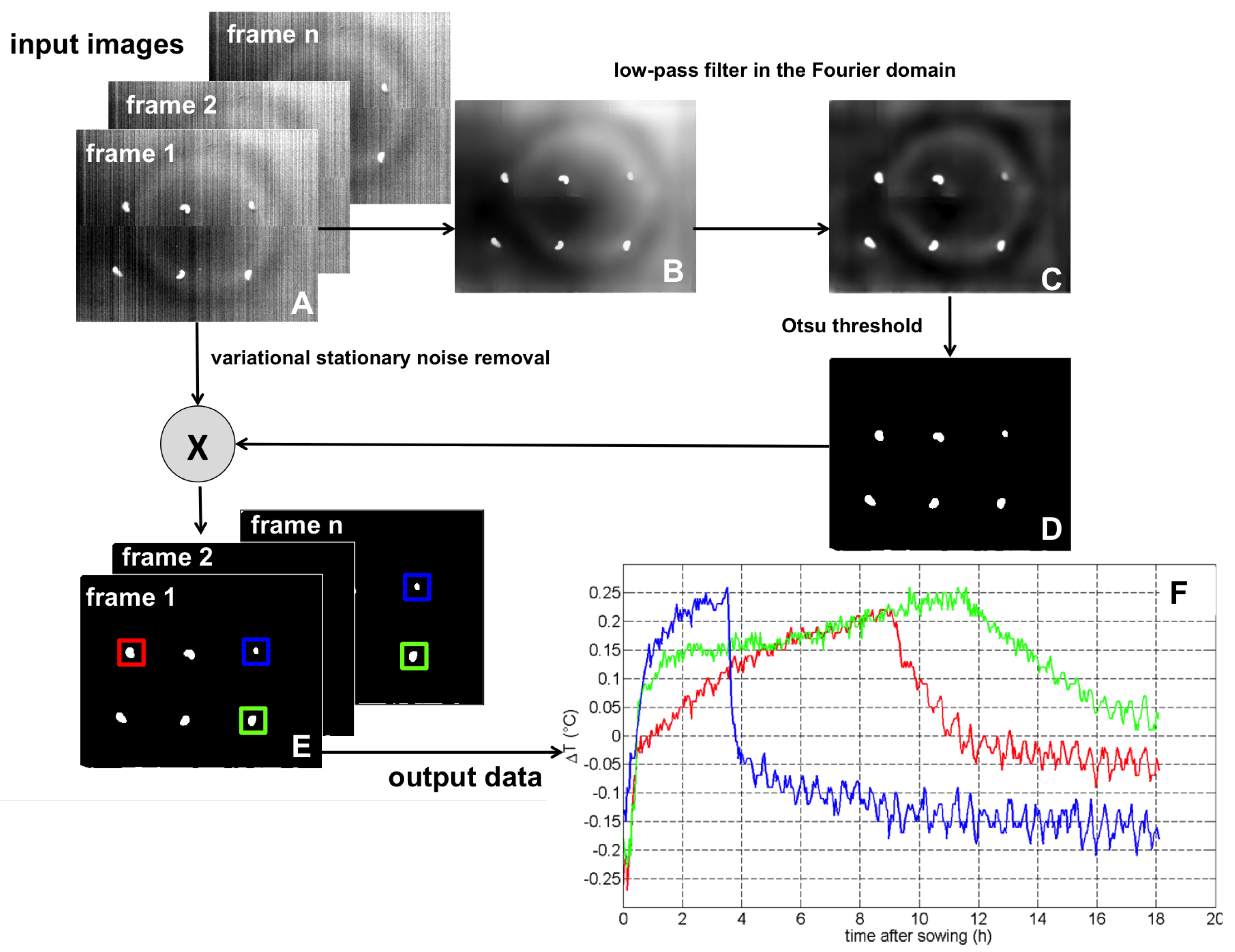
4.2. Image Processing Pipeline
4.3. Illustration
4.4. Interests and Limitations
5. 2D Biospeckle Imaging of Seed Imbibition out of Soil
5.1. Acquisition Protocol
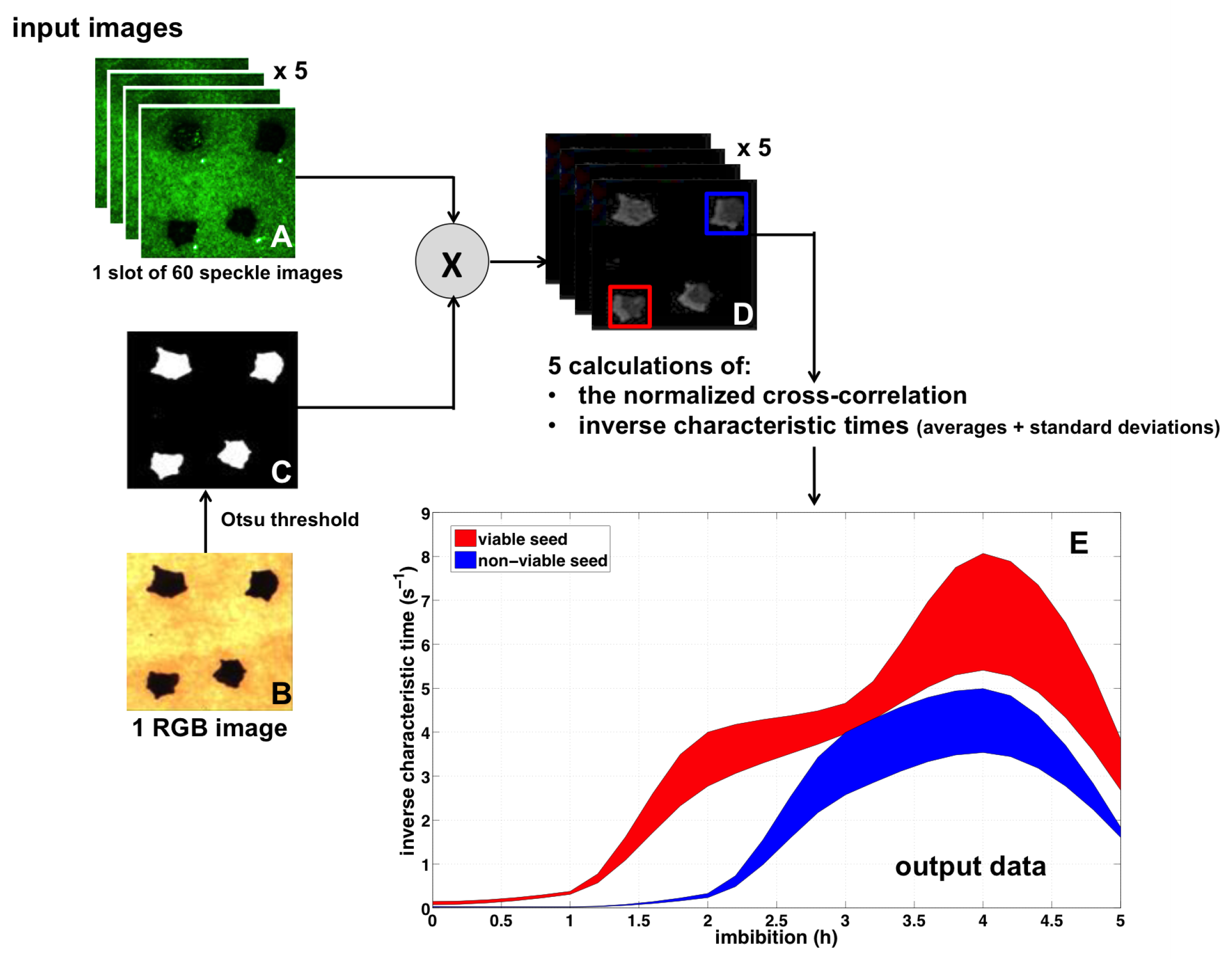
5.2. Image Processing Pipeline
5.3. Illustration
5.4. Interests and Limitations
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bucksch, A.; Burridge, J.; York, L.M.; Das, A.; Nord, E.; Weitz, J.S.; Lynch, J.P. Image-based high-throughput field phenotyping of crop roots. Plant Physiol. 2014, 166, 470–486. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Acosta-Gamboa, L.M.; Huang, X.; Lorence, A. Novel Low Cost 3D Surface Model Reconstruction System for Plant Phenotyping. J. Imaging 2017, 3, 39. [Google Scholar] [CrossRef]
- Mutka, A.M.; Bart, R.S. Image-based phenotyping of plant disease symptoms. Front. Plant Sci. 2015, 5, 734. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, R.; Spalding, E.P.; Ferrier, N.J. A high throughput robot system for machine vision based plant phenotype studies. Mach. Vis. Appl. 2013, 11, 619–636. [Google Scholar] [CrossRef]
- Humplík, J.F.; Lazár, D.; Husičková, A.; Spíchal, L. Automated phenotyping of plant shoots using imaging methods for analysis of plant stress responses—A review. Plant Methods 2015, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Dhondt, S.; Wuyts, N.; Inzé, D. Cell to whole-plant phenotyping: The best is yet to come. Trends Plant Sci. 2013, 18, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Paulus, S.; Behmann, J.; Mahlein, A.K.; Plümer, L.; Kuhlmann, H. Low-cost 3D systems: Suitable tools for plant phenotyping. Sensors 2014, 14, 3001–3018. [Google Scholar] [CrossRef] [PubMed]
- Minervini, M.; Scharr, H.; Tsaftaris, S.A. Image analysis: The new bottleneck in plant phenotyping. IEEE Signal Process. Mag. 2015, 32, 126–131. [Google Scholar] [CrossRef]
- Tsaftaris, S.A.; Minervini, M.; Scharr, H. Machine learning for plant phenotyping needs image processing. Trends Plant Sci. 2016, 21, 989–991. [Google Scholar] [CrossRef] [PubMed]
- Dell’Aquila, A. Towards new computer imaging techniques applied to seed quality testing and sorting. Seed Sci. Technol. 2007, 35, 519–538. [Google Scholar] [CrossRef]
- Dell’Aquila, A. Development of novel techniques in conditioning, testing and sorting seed physiological quality. Seed Sci. Technol. 2009, 37, 608–624. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination; Plenum Press: New York, NY, USA, 1994. [Google Scholar]
- Woodstock, L.W. Seed imbibition: A critical perido for successful germination. J. Seed Technol. 1988, 12, 1–15. [Google Scholar]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, L.N.; Fregeau-Reid, J.; Chatson, B.; Blackwell, B. Observations on water distribution in soybean seed during hydration processes using nuclear magnetic resonance imaging. Can. J. Plant Sci. 2002, 82, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Manz, B.; Müller, K.; Kucera, B.; Volke, F.; Leubner-Metzger, G. Water uptake and distribution in germinating tobacco seeds investigated in vivo by nuclear magnetic resonance imaging. Plant Physiol. 2005, 138, 1538–1551. [Google Scholar] [CrossRef] [PubMed]
- Dell’Aquila, A. Pepper seed germination assessed by combined X-radiography and computer-aided imaging analysis. Biol. Plant. 2007, 51, 777–781. [Google Scholar] [CrossRef]
- Romans, L.E. Computed Tomography for Technologists: A Comprehensive Text; Wolters Kluwer Health/Lippincott Williams & Wilkins: Alphen aan den Rijn, The Netherlands, 2010. [Google Scholar]
- Foucat, L.; Chavagnat, A.; Renou, J.P. Nuclear magnetic resonance micro-imaging and X-radiography as possible techniques to study seed germination. Sci. Hortic. 1993, 55, 323–331. [Google Scholar] [CrossRef]
- Otsu, N. A threshold selection method from gray-level histograms. Automatica 1975, 11, 23–27. [Google Scholar] [CrossRef]
- Gonzalez, R.C.; Woods, R.E. Digital Image Processing, 3rd ed.; Pearson Prentice Hall: New York, NY, USA, 2008. [Google Scholar]
- Rousseau, D.; Widiez, T.; Tommaso, S.; Rositi, H.; Adrien, J.; Maire, E.; Langer, M.; Olivier, C.; Peyrin, F.; Rogowsky, P. Fast virtual histology using X-ray in-line phase tomography: Application to the 3D anatomy of maize developing seeds. Plant Methods 2015, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Staedler, Y.M.; Masson, D.; Schönenberger, J. Plant tissues in 3D via X-ray tomography: Simple contrasting methods allow high resolution imaging. PLoS ONE 2013, 8, e75295. [Google Scholar] [CrossRef] [PubMed]
- Borisjuk, L.; Rolletschek, H.; Neuberger, T. Surveying the plant’s world by magnetic resonance imaging. Plant J. 2012, 70, 129–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination—Still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Chaerle, L.; Leinonen, I.; Jones, H.G.; der Straeten, D.V. Monitoring and screening plant populations with combined thermal and chlorophyll fluorescence imaging. J. Exp. Bot. 2007, 58, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Belin, E.; Rousseau, D.; Boureau, T.; Caffier, V. Thermography versus chlorophyll fluorescence imaging for detection and quantification of apple scab. Comput. Electron. Agric. 2013, 90, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Chéné, Y.; Belin, E.; Chapeau-Blondeau, F.; Boureau, T.; Caffier, V.; Rousseau, D. Anatomo-functional bimodality imaging for plant phenotyping: An insight through depth imaging coupled to thermal imaging. In Plant Image Analysis: Fundamentals and Applications; Dutta Gupta, S., Ibaraki, Y., Eds.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Wang, X.Z.; Yang, W.P.; Wheaton, A.; Cooley, N.; Moran, B. Automated canopy temperature estimation via infrared thermography: A first step towards automated plant water stress monitoring. Comput. Electron. Agric. 2010, 73, 74–83. [Google Scholar] [CrossRef]
- Baranowski, P.; Mazurek, W.; Walczak, R.T. The use of thermography for pre-sowing evaluation of seed germination capacity. In Proceedings of the International Conference on Quality Chains—An Integrated View on Fruit and Vegetable, Wageningen, The Netherlands, 1 July 2003; Volumes 1 and 2. [Google Scholar]
- Kranner, I.; Kastberger, G.; Hartbauer, M.; Pritchard, H.W. Non-invasive diagnosis of seed viability using infrared thermography. Proc. Natl. Acad. Sci. USA 2010, 107, 3912–3917. [Google Scholar] [CrossRef] [PubMed]
- Belin, E.; Rousseau, D.; Rojas-Varela, J.; Demilly, D.; Wagner, M.H.; Cathala, M.H.; Dürr, C. Thermography as a non-invasive functional imaging for monitoring seedling growth. Comput. Electron. Agric. 2011, 79, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Belin, E.; Rousseau, D.; Benoit, L.; Demilly, D.; Ducournau, S.; Chapeau-Blondeau, F.; Dürr, C. Thermal imaging for evaluation of seedling growth. In Plant Image Analysis: Fundamentals and Applications; Dutta Gupta, S., Ibaraki, Y., Eds.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Budzan, S.; Wyżgolik, R. Noise reduction in thermal images. In International Conference on Computer Vision and Graphics; Springer: Berlin, Germany, 2014; pp. 116–123. [Google Scholar]
- Rogalski, A. Infrared Detectors, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Fehrenbach, J.; Weiss, P.; Lorenzo, C. Variational algorithms to remove stationary noise: Applications to microscopy imaging. IEEE Trans. Image Process. 2012, 21, 4420–4430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieczywek, P.M.; Kurenda, A.; Zdunek, A.; Adamiak, A. The biospeckle method for the investigation of agricultural crops: A review. Opt. Lasers Eng. 2014, 52, 156–158. [Google Scholar]
- Braga Júnior, R.A. When noise became information: State-of-the-art in biospeckle laser. Cienc. Agrotecnologia 2017, 41, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Miquet, E.E.; Darias, J.G.; Otero, I.; Rodriguez, D.; Murialdo, S.; Rabal, H.; Trivi, M. Biospeckle technique for monitoring bacterial colony growth with minimal photo-exposure time associated. In Proceedings of the VI Latin American Congress on Biomedical Engineering CLAIB 2014, Parana, Argentina, 29–31 October 2014; Springer: Cham, Switzerland, 2014; pp. 313–316. [Google Scholar]
- Ramirez-Miquet, E.E.; Cabrera, H.; Grassi, H.C.; Andrades, E.D.J.; Otero, I.; Rodriguez, D.; Darias, J.G. Digital imaging information technology for biospeckle activity assessment relative to bacteria and parasites. Lasers Med. Sci. 2017, 32, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, G.F.; Enes, A.M.; Braga Júnior, R.A.; Dal Fabbro, I.M. Frequency response of biospeckle laser images of bean seeds contaminated by fungi. Biosyst. Eng. 2011, 110, 297–301. [Google Scholar] [CrossRef]
- Rousseau, D.; Caredda, C.; Morille, Y.; Belin, E.; Chapeau-Blondeau, F.; Gindre, D. Low-cost biospeckle imaging applied to the monitoring of seed germination. In Proceedings of the 3rd International Workshop on Image Analysis Methods for the Plant Sciences (IAMPS), Aberystwyst, UK, 15–16 September 2014. [Google Scholar]
- Braga, R.A.; Dal Fabbro, I.M.; Borem, F.M.; Rabelo, G.; Arizaga, R.; Rabal, H.J.; Trivi, M. Assessment of seed viability by laser speckle techniques. Biosyst. Eng. 2003, 86, 287–294. [Google Scholar] [CrossRef]
- Platform PHENOTIC. Available online: http://laris.univ-angers.fr/fr/activites-scientifiques/projets/plateformes-1/plateforme-phenotic-1.html (accessed on 1 June 2018).
- Hartley, R.; Zisserman, A. Multiple View Geometry in Computer Vision; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Rousseau, D.; Chéné, Y.; Belin, E.; Semaan, G.; Trigui, G.; Boudehri, K.; Franconi, F.; Chapeau-Blondeau, F. Multiscale imaging of plants: Current approaches and challenges. Plant Methods 2015, 11, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Imaging Technique | Imbibition Trait Measured | Interests | Limitations |
|---|---|---|---|
| X-ray | remaining air volume | 3D, high spatial resolution | low contrast, very expensive costs |
| MRI | imbibed volume | 3D, high water sensitivity | low throughput, low spatial resolution, very expensive costs |
| thermography | thermal signature of biochemical activities | high sensitivity | 2D, expensive costs |
| biospeckle | external deformations | high micrometer sensitivity, very low costs | 2D |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belin, E.; Douarre, C.; Gillard, N.; Franconi, F.; Rojas-Varela, J.; Chapeau-Blondeau, F.; Demilly, D.; Adrien, J.; Maire, E.; Rousseau, D. Evaluation of 3D/2D Imaging and Image Processing Techniques for the Monitoring of Seed Imbibition. J. Imaging 2018, 4, 83. https://doi.org/10.3390/jimaging4070083
Belin E, Douarre C, Gillard N, Franconi F, Rojas-Varela J, Chapeau-Blondeau F, Demilly D, Adrien J, Maire E, Rousseau D. Evaluation of 3D/2D Imaging and Image Processing Techniques for the Monitoring of Seed Imbibition. Journal of Imaging. 2018; 4(7):83. https://doi.org/10.3390/jimaging4070083
Chicago/Turabian StyleBelin, Etienne, Clément Douarre, Nicolas Gillard, Florence Franconi, Julio Rojas-Varela, François Chapeau-Blondeau, Didier Demilly, Jérôme Adrien, Eric Maire, and David Rousseau. 2018. "Evaluation of 3D/2D Imaging and Image Processing Techniques for the Monitoring of Seed Imbibition" Journal of Imaging 4, no. 7: 83. https://doi.org/10.3390/jimaging4070083