

Biocontrol Potential of Trichoderma asperellum Strain 576 against Exserohilum turcicum in Zea mays

Engineering Research Center of Edible and Medicinal Fungi, Ministry of Education, Jilin Agricultural University, Changchun 130118, China
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(9), 936; https://doi.org/10.3390/jof9090936
Submission received: 29 June 2023
/
Revised: 6 September 2023
/
Accepted: 11 September 2023
/
Published: 16 September 2023
(This article belongs to the Section Fungi in Agriculture and Biotechnology)
Abstract
:Maize is a crucial cereal crop in China, serving both as a staple food and an essential industrial resource. Northern corn leaf blight (NCLB) is a disease of corn caused by a fungus, Exserohilum turcicum (sexual stage Setosphaeria turcica). This study aimed to assess the biocontrol potential of various Trichoderma strains against Exserohilum turcicum 101 in Jilin, China. Through dual culture tests, the Trichoderma strains were categorized into four groups based on their antagonistic abilities. Eleven Trichoderma strains exhibited strong antagonistic behavior, with comparable or faster growth rates than E. turcicum 101. Microscopic observations confirmed that T. asperellum 576 hyphae effectively encircled E. turcicum 101 hyphae, reinforcing their antagonistic behavior. The production of non-volatile and volatile substances by the Trichoderma strains was evaluated, with T. asperellum 576 showing the highest potency in producing non-volatile and volatile substances, leading to an impressive 80.81% and 65.86% inhibition of E. turcicum 101 growth. Remarkably, co-culture suspensions of T. asperellum 576 + E. turcicum 101 and T. atroviride 393 + E. turcicum 101 exhibited strong antifungal activity. Furthermore, the activities of chitinase, β-1.3-glucanase, and cellulase were evaluated using the 3, 5-dinitrosalicylic acid (DNS) method. T. asperellum 576 + E. turcicum 101 displayed stronger cell wall degradation enzyme activity compared to T. atroviride 393 + E. turcicum 101, with values of 8.34 U/mL, 3.42 U/mL, and 7.75 U/mL, respectively. In greenhouse conditions, the application of a 107 spores/mL conidia suspension of T. asperellum 576 significantly enhanced maize seed germination and plant growth while effectively suppressing E. turcicum 101 infection. Maize seedlings inoculated/treated with both E. turcicum 101 and T. asperellum 576 demonstrated substantial improvements compared to those inoculated solely with E. turcicum 101. The T. asperellum 576 treatment involved a 107 spores/mL conidia suspension applied through a combination of foliar spray and soil drench. These findings highlight T. asperellum 576 as a promising biocontrol candidate against northern leaf blight in maize. Its antagonistic behavior, production of inhibitory compounds, and promotion of plant growth all contribute to its potential as an effective biocontrol agent for disease management.
1. Introduction
Maize (Zea mays L.) holds significant agricultural importance as a major crop in China [1]. It thrives in various regions, including the tropics, subtropics, and temperate areas, under both irrigated and rainfed conditions [2,3]. Maize stands out among other grains as a highly nutritious crop, providing essential nutrients for both humans and animals [4,5]. However, the devastating fungal disease Northern corn leaf blight (NCLB) is caused by the pathogen Exserohilum turcicum (Pass.) Leonard and Suggs (Pleosporaceae, Pleosporales, Pleosporomycetidae, Dothideomycetes, Pezizomycotina, Ascomycota, Fungi) [6,7,8] pose a significant threat to maize production. It occurs in all maize-producing regions, from tropical to temperate zones [9]. E. turcicum causes cigar-shaped green-grey lesions on leaves, which become necrotic in later infection stages and may evolve to blight symptoms, leading to high yield losses in maize [10]. Maize yield losses caused by E. turcicum are up to 50% in China from July to August every year. Uncontrolled NCLB can lead to substantial yield losses, documented at up to $2 billion in several countries [11,12]. With the projected increase in soil-borne pathogen species due to global warming, effective control of NCLB becomes even more critical, particularly for Asian countries like China. Traditionally, maize production practices heavily relied on chemical interventions, with three chemicals, metconazole, propiconazole, and prothioconazole, being the primary demethylation inhibitors and fungicides used in the production of corn fields [13]. However, the widespread use of these chemical substances poses risks to human health and the environment [14]. Therefore, it is essential to explore alternative or complementary control mechanisms and strategies to slow down the emergence of resistance in pathogens [15,16,17]. Biocontrol agents offer a promising avenue for crop protection. For instance, three Bacillus isolates demonstrated dominance and significantly reduced the growth rate of E. turcicum [18]. Among the biocontrol agents, Trichoderma strains present several advantages, including their ability to suppress multiple pathogens, their cost-effectiveness, and their promotion of soil fertility [19,20]. By utilizing biocontrol agents, the negative biological and environmental consequences associated with the continuous use of synthetic chemicals can be mitigated [21,22].
Trichoderma strains have gained significant attention as biological control agents (BCAs) since the groundbreaking research conducted by Weindling [23]. One of the key advantages of Trichoderma is its adaptability, as it can thrive in diverse soil types and tolerate a range of temperatures conducive to its growth [24,25]. Trichoderma exerts its protective effects on plants through various mechanisms, including mycoparasitism, antibiosis, promoting plant host resistance, production of plant growth-promoting substances, and modulation of plant hormonal pathways.
Mycoparasitism is a crucial attribute of Trichoderma, wherein it attacks other fungi by infiltrating their structures, killing them, and utilizing the nutrients within their cells [17]. This ability allows Trichoderma to compete effectively with other fungal pathogens for resources and space, thereby limiting their growth [26]. Additionally, Trichoderma exhibits a rapid growth rate, further aiding its competitiveness against plant infections. Antibiosis is another essential mechanism employed by Trichoderma in biological control. Trichoderma has the capacity to produce antimicrobial compounds that can inhibit the growth of pathogenic fungi [27]. These antimicrobial compounds include non-volatile compounds and volatile organic compounds (VOCs) [28]. Non-volatile compounds of Trichoderma have active effects against a range of plant pathogens [29]. Approximately 390 non-volatile compounds from several Trichoderma species have been summarized [30]. VOCs of Trichoderma were shown to inhibit plant pathogens, induce plant resistance, and directly promote plant growth, indicating that VOCs may play a role in the biocontrol activity of Trichoderma spp. [31,32,33]. As far, over 480 VOCs have been detected from Trichoderma species altogether. The detected Trichoderma VOCs comprise simple hydrocarbons, phenols, heterocycles, aldehydes, ketones, thioalcohols, thioesters, and their derivatives [34]. Research by Limdolthamand et al. demonstrated the efficacy of a fresh formulation of Trichoderma harzianum Rifai KUFA0710 in reducing the growth of the fungal pathogen E. turcicum in field trials [35]. This finding underscores the practical applicability of Trichoderma as a BCA in controlling plant diseases.
Furthermore, Trichoderma has been found to promote plant host resistance, offering an additional layer of protection against pathogens [36]. The exact mechanisms by which Trichoderma induces host resistance are not yet fully understood but have been demonstrated in various pathosystems [37]. The ability of Trichoderma to enhance plant defenses can contribute to long-term disease management and reduce the reliance on chemical pesticides.
The existing studies on Trichoderma as a BCA in the pathosystems of maize and E. turcicum are limited. Therefore, the primary objective of our investigation is to evaluate the effectiveness of forty-four strains of Trichoderma against E. turcicum through in vitro, in vivo, and seedling trials. This comprehensive approach aims to provide a more thorough understanding of Trichoderma’s potential as a BCA and its practical application.
2. Materials and Methods
2.1. Fungal Strains and Identification
Forty-four Trichoderma strains were collected from various provinces in China since 2014 (Table 1). The pathogenic strain Exserohilum turcicum was obtained from Jilin Agricultural University. All strains were preserved in the Engineering Research Center of the Chinese Ministry of Education for Edible and Medicinal Fungi at Jilin Agricultural University. The Trichoderma strains were isolated either by single ascospore isolation from fresh stromata of sexual morphs or by direct isolation from asexual morphs on the substrates [38,39]. The species identification of Trichoderma primarily relied on comprehensive morphological characteristics, including observations of stromata, ascus, and ascospores, as well as colony appearance, growth rate, and the structure of conidiophores and conidia. These morphological features were essential in distinguishing different Trichoderma species.
Genomic DNA was extracted from the mycelium of cultures on PDA using a Plant Genomic DNA Extraction Kit (TIANGEN Biosciences, Beijing, China). In cases where certain species presented challenges in identification, we employed molecular analysis of translation elongation factor 1-alpha (TEF1-α) and RNA polymerase II’s second largest subunit (RPB2) as an assisting method [40,41]. PCR products were cycle sequenced on an ABI 3730 XL DNA Sequencer (Applied Biosciences, Foster City, CA, USA) with primers reported by Jaklitsch [42] at Beijing Tianyihuiyuan Bioscience and Technology, China. The strains and the NCBI GenBank accession numbers of DNA sequences used in this work are listed in Table 1.
To support the species identification of Trichoderma strain 576, we conducted a comprehensive phylogenetic analysis. Sequences were assembled, aligned, and manually adjusted when needed with BioEdit 7.0.5.3 [43]. NEXUS files were generated with Clustal X 1.83 [44]. To achieve accurate placement within the phylogenetic framework, we utilized both TEF1-α and RPB2 sequences in our analyses. We selected a set of 18 Trichoderma taxa, including T. thelephoricola 342, as an outgroup taxa.
Maximum likelihood (ML) analysis was performed with RaxmlGUI 2.0 [45], the ML + rapid bootstrap setting, and the GTRGAMMAI substitution model with 1000 bootstrap replicates. Analyses were performed with all characters treated as unordered and unweighted and gaps treated as missing data. Topological confidence of resulted trees was tested by maximum parsimony bootstrap proportions (MLBP) with 1000 replications, each with 10 replicates of random addition of taxa. MLBP greater than 50% is shown at the nodes.
2.2. Dual-Culture Antagonistic Activity Assay
To assess the antagonistic effect of the Trichoderma strains against E. turcicum 101, a dual culture test was conducted. Based on the observations of Trichoderma’s growth rate, we decided to introduce the E. turcicum 101 plugs onto the dual culture agar two days prior to the addition of Trichoderma. This time interval allowed the E. turcicum 101 isolates to establish themselves before encountering Trichoderma. The procedure involved placing 5-mm diameter fungal plugs of both the Trichoderma strains and E. turcicum 101 on Petri dishes (90 mm diameter) containing potato dextrose agar (PDA), with a distance of 5 cm between them [46]. The Petri dishes were then incubated at 25 °C under a 12-h cycle of light and dark for a period of 15 days. The experimental design followed a completely randomized design, with three replications performed. Each experimental iteration was repeated three times to ensure reliable and consistent results. The growth rate and sporulation of Trichoderma species on E. turcicum 101 were monitored and documented on the fourth, seventh, and fifteenth days after inoculation using a Canon G5 digital camera (Canon, Tokyo, Japan). The findings were analyzed based on the degree of antagonism observed. Zhang et al. utilized four categories to describe the degree of antagonism: strong antagonism, moderate antagonism, weak antagonism, and absence of antagonism [47]. In the case of Trichoderma strains exhibiting strong antagonism, they displayed rapid growth comparable to or even faster than phytopathogenic fungi. Additionally, they formed extensive hyphal coilings around the hyphae of the pathogenic fungi and exhibited abundant sporulation on the colonies of the phytopathogenic fungi within a span of 15 days. Trichoderma strains demonstrating moderate antagonism also exhibited fast growth rates similar to phytopathogenic fungi. They were capable of sporulating on the colonies of phytopathogenic fungi within 15 days, although the sporulation may be comparatively less abundant. On the other hand, Trichoderma strains with weak antagonism tended to grow slower than phytopathogenic fungi. These strains displayed minimal or no sporulation on the phytopathogenic fungal colonies within the 15-day timeframe. Finally, Trichoderma strains lacking antagonism exhibited limited and slow growth, often becoming overgrown by the colonies of phytopathogenic fungi. These categories were used to assess and interpret the results obtained from the dual culture test.
2.3. Effect of Non-Volatile Substances Produced by Trichoderma Strains
The Trichoderma strains and E. turcicum 101 were initially inoculated on PDA and incubated at a temperature of 25 °C. The cellophane filtration membrane method, originally developed by Dennis and Webster, was employed to assess the ability of a subset of Trichoderma strains to generate non-volatile inhibitors [29]. The procedure involved inoculating the Trichoderma strains onto cellophane-covered Petri dishes, which were then incubated for 4 days at 25 °C. After removing the cellophane and mycelium, an E. turcicum 101 inoculum Petri dish was placed in the center of the medium previously occupied by Trichoderma strains. Following a 7-day reincubation at 28 °C, the inhibitory rate of the Trichoderma strains was determined by calculating the difference in diameter between the target colony in the absence and presence of the antagonist. This experiment was repeated three times. The inhibitory rate of the Trichoderma strains was calculated using the provided formula:
where D1 is the diameter of the target colony in the absence of the antagonist, while D2 is the diameter of the target colony in the presence of the antagonist.
Inhibition rate (%) = (D1 − D2)/D1 × 100
2.4. Effect of Volatile Substances Produced by Trichoderma Strains
E. turcicum 101 were initially inoculated on PDA and incubated at a temperature of 25 °C for 3 days. The Trichoderma strains were initially inoculated on PDA and incubated at a temperature of 25 °C for 1 day. After one day, the covers of the Petri dishes were replaced with the bottoms of 3-day-old PDA cultures of E. turcicum 101, as described by Jin and Khalid [48]. The two halves or cultures of the plates were securely taped together using parafilm tape and maintained at 25 °C for a duration of 10 days. In the control group, only E. turcicum 101 was inoculated. To ensure standardized Trichoderma sporulation, all bioassay procedures were conducted under controlled light-limited conditions [49]. Each test was repeated three times to ensure accuracy, and the inhibition of mycelial radial growth was calculated as described previously.
2.5. Fermentation Broth Antagonistic Assays
In order to assess the interaction between Trichoderma strains and E. turcicum 101 and evaluate the mycelial radial growth inhibition, the following experimental procedure was conducted. E. turcicum 101 was cultured on PDA plates at a temperature of 30 °C for 7 days. Meanwhile, Trichoderma strains were grown on PDA plates at 25 °C for 5 days. Circular five mycelia blocks with a diameter of 5 mm were created at the edge of the colonies using a punch. The five mycelia blocks of both Trichoderma strains and E. turcicum 101 were inoculated into sterilized potato dextrose broth (PDB) medium in the same flasks together. The flasks were then placed on a shaker incubator and incubated for 10 days at a temperature of 28 °C with a shaking speed of 150 rpm. From day 2 to day 12 of the fermentation process, 10 mL of the liquid culture was extracted daily. The fermented liquid was filtered using filter paper and subsequently centrifuged at 12,000 rpm for 10 min. Finally, it was filtered with a 0.22 μm filter membrane [50]. The resulting fermented liquid was mixed with solid PDA medium (approximately 60 °C) in a proportion of 1:9 (v/v) and poured into plates. A 5 mm diameter block of E. turcicum 101 was placed in the center of each plate, and the plates were incubated at 25 °C for 5 days. Each treatment was replicated three times. The mycelial radial growth inhibition was calculated as described previously.
2.6. Enzymatic Activity
To evaluate the activities of chitinase, β-1.3-glucanase, and cellulase, a co-culture suspension was prepared as described in 2.5. Colloid chitin, β-1,3-glucan, and Sigma-Aldrich (CMC) were utilized as substrates to measure the respective enzyme activities. The assays for all three enzymes were conducted using the 3,5-dinitrosalicylic acid (DNS) method developed by Miller [51]. For the chitinase assay, the reaction mixture comprised a co-culture supernatant, which was added to 200 mL of 0.5% colloidal chitin in 50 mM sodium acetate buffer (pH 5.0). The mixture was incubated at 37 °C for 1 h. To stop the reaction, 500 µL of DNS reagent was added, followed by boiling for 5 min. In the β-1.3-glucanase assay, the reaction mixture consisted of 100 μL of 0.2 mM sodium acetate buffer (pH 5.5), 50 μL of enzyme extract, and 50 μL of laminarin substrate. The mixture was incubated at 37 °C for 30 min. The reaction was stopped by adding 500 µL of DNS reagent and boiling for 5 min. For the cellulase assay, the reaction mixture included 100 mL of 50 mM sodium acetate buffer (pH 5.0), 50 mL of 1% CMC substrate, and 50 mL of enzyme extract. The mixture was incubated at 50 °C for 60 min. The reaction was terminated by adding 500 µL of DNS reagent, followed by boiling in a water bath for 5 min. The spectrophotometer recorded absorption peaks at 585 nm, 540 nm, and 620 nm to measure the enzymatic activities of chitinase, β-1.3-glucanase, and cellulase, respectively. The experiment was replicated three times for each treatment, and the entire study was repeated at least three times to ensure reliability.
2.7. Effect of T. asperellum 576 on Maize Seed Germination
To assess the germination effects of T. asperellum 576, the maize seeds were subjected to surface disinfection using a 1.5% sodium hypochlorite solution for 5 min [52], followed by placement in clear plastic boxes with lids. Initially, T. asperellum 576 cultures were grown on a PDA medium. After the formation of spores, sterile distilled water was added to the PDA plate. The mycelium was gently rubbed with a sterilized needle to release the spores into a centrifugal tube, which was then collected from the plate using a micropipette to obtain mycelium spores. Using a hematocytometer, the number of conidia was determined, and their concentration was adjusted to 107 spores/mL. The conidial suspension was placed in a centrifugal tube. Each treatment group consisted of 30 seeds: one group was treated with 10 mL of 107 spores/mL T. asperellum 576 conidial suspension, while the control group was treated with 10 mL of sterile distilled water. The seeds were placed on moistened blotter papers inside the plastic boxes after surface disinfection. Subsequently, 10 mL of either T. asperellum 576 conidial suspension or sterile distilled water was poured into each box. The boxes were then incubated at 25 °C with a 16-h light cycle, and the germination progress was monitored every 12 h. The experiment was replicated three times for each treatment, and the entire study was repeated at least three times to ensure reliability. Germination was recorded when the radicle protruded through the seed coat.
2.8. Biocontrol Effect of T. asperellum 576 against E. turcicum 101
Maize seeds used in this study were sourced from the Jilin Provincial Agricultural Science laboratory. The seeds were sown in pots filled with sterile soil consisting of a 1:1 ratio of vermiculite to soil [37]. The pots had dimensions of 14 cm in diameter and 12 cm in height. The seedlings were grown in a greenhouse with controlled conditions, maintaining a temperature of 25 °C and relative humidity of 85%. At the beginning of the experiment, all seedlings were at the four-leaf stage.
To evaluate the biocontrol effect of T. asperellum 576 against E. turcicum 101 under greenhouse conditions, maize seedlings were subjected to four treatments. These included: (T1) plants without T. asperellum 576 and E. turcicum 101, (T2) T. asperellum 576-treated plants, (T3) E. turcicum 101-inoculated plants, and (T4) E. turcicum 101 and T. asperellum 576-inoculated/treated plants. The seedlings were cultivated under controlled conditions with a temperature of 25 °C, a 12-h light/dark cycle, and 60–70% humidity. They were watered every two days for a period of 30 days.
T1: a control group treated with 30 mL of sterile distilled water; T2: seedlings treated with a combination of foliar spray (30 mL) and soil drench (30 mL) application of conidial suspension of T. asperellum 576, containing 107 conidia/mL; T3: seedlings treated with a foliar application of 30 mL conidial suspension of E. turcicum 101, containing 105 conidia/mL, to induce infection; and T4: seedlings treated with a foliar application of 30 mL conidial suspension of E. turcicum 101 and subsequently treated with T. asperellum 576 using a combination of foliar spray (30 mL) and soil drench (30 mL) methods, both with conidial suspension containing 107 conidia/mL. The experiment followed a completely randomized design with a total of sixty seedlings (four treatments × fifteen replicates), and the entire experiment was conducted three times.
After a treatment period of 20 days, the seedlings were visually examined for symptoms or signs of infection by E. turcicum 101 and compared with the control group. Plant height was measured from the base of the seedling to the tip of the tallest shoot using a vernier caliper with an accuracy of 0.1 mm. Stem diameter was measured at the midpoint of the main stem using the same vernier caliper. Fresh weight measurements of stems and roots were recorded using an electronic balance with a precision of 0.01 g immediately after harvesting. Subsequently, the plant samples were dried in an oven at 100 °C for 30 min to remove surface moisture and then dried at 80 °C for 48 h, or until a consistent mass was achieved, to determine the dry weight.
2.9. Statistical Analysis
Statistical analyses were conducted using Minitab® Statistical Software version 18 (IBM Co., New York, NY, USA). The data were analyzed using analysis of variance (ANOVA), followed by Fisher’s least significant difference (LSD) multiple comparison test (p < 0.05) to determine significant differences between treatment groups. The results are presented as means of replicates or independent experiments, and the error bars represent the standard deviation (SD).
3. Results
3.1. Species Identification and Phylogenetic Analysis
Employing a comprehensive analysis method that integrates both morphology and molecular sequences, we successfully identified a total of 44 distinct Trichoderma strains (Table 1). All accompanying plate photographs are available in Supplementary Materials Figure S1. The subsequent phylogenetic analysis involved 18 selected taxa representing major Trichoderma species. The alignment of sequences facilitated the definitive classification of isolate 576 as Trichoderma asperellum, as evident in the phylogenetic tree (Figure 1).
3.2. Dual Culture Tests
Among the selected strains, Trichoderma strains 576, 393, 64, 3A, 342, 285, TU2, XZ1-3, 402, MSL-3, and 204 displayed strong antagonistic behavior. These strains exhibited comparable or faster growth rates than E. turcicum 101, formed extensive hyphal coilings around the hyphae of E. turcicum 101, and showed abundant sporulation on the colonies of E. turcicum 101 over a 15-day period (Figure 2). Microscopic observations provided visual evidence of the interaction between T. asperellum 576 and E. turcicum 101, showing the encirclement of E. turcicum 101 hyphae by T. asperellum 576 hyphae (Figure 3). The mycelium of Trichoderma was observed to be entangled in the mycelium of E. turcicum 101. This interaction may imply that Trichoderma parasitizes the mycelium of E. turcicum 101, or it may be part of a competitive process by which Trichoderma inhibits the growth of the pathogen. This observation further supports the antagonistic behavior and potential biocontrol activity of the identified Trichoderma strains.
In addition, twelve Trichoderma strains, namely 110, 224, 295, 375, 99, 421, 539, 581, 593, 526, 578, and XZ9-1, demonstrated a moderate antagonistic response after a 15-day co-culture period. These strains exhibited accelerated growth rates comparable to or surpassing that of E. turcicum 101 but displayed limited sporulation on the colonies of E. turcicum 101 (Supplementary Materials Figure S2). The strong and moderately antagonistic Trichoderma strains hold promise for biocontrol applications and warrant further comprehensive investigation and research.
On the other hand, weakly antagonistic Trichoderma strains had a decelerating effect on the growth of E. turcicum 101. These strains caused limited sporulation of E. turcicum 101 colonies within the 15-day co-culture period (Supplementary Materials Figure S3).
The results also revealed that Trichoderma strains did not exhibit detectable antagonistic behavior towards E. turcicum 101. These strains displayed slower and restricted growth rates compared to E. turcicum 101, suggesting a limited potential for biological control measures (Supplementary Materials Figure S4).
3.3. Antagonistic Effect by Trichoderma Non-Volatile Substances
According to Table 2 and Figure 4, T. asperellum 576 exhibited the highest potency in producing non-volatile compounds, resulting in an inhibition of E. turcicum 101 growth by 80.81%. Additionally, seven strains (393, 421, 110, 3A, XZ9-1, 285, and 539) showed inhibition rates over 50.00%, while six strains (417, XZ1-3, 342, TU2, 64 and 402) exhibited less than 50.00% inhibition against E. turcicum 101. These results provide valuable insights into the antifungal potential of the examined Trichoderma strains and highlight T. asperellum 576 as a particularly promising candidate for further investigation and utilization in biocontrol strategies.
3.4. Fungal Growth Inhibition by Trichoderma Volatile Substances
As shown in Table 3 and Figure 5, T. asperellum 576 displayed the highest inhibitory effect on E. turcicum 101 mycelial growth with a rate of 65.86% compared to CK. Following T. asperellum 576, Trichoderma strains 393, 421, 110, and 3A also showed considerable inhibitory activity, with inhibition rates ranging from 23.10% to 57.54%. Based on these findings, T. asperellum 576 and T. atroviride 393 were selected for further testing and investigation, given their strong potential as biocontrol agents against E. turcicum 101.
3.5. Antifungal Activity of the Co-Culture Suspension against E. turcicum 101
According to Table 4, the co-culture suspensions of T. asperellum 576 + E. turcicum 101 or T. atroviride 393 + E. turcicum 101 exhibited strong antifungal activity against E. turcicum 101, as evidenced by the reduction in mycelial growth over time. All co-culture suspensions demonstrated mycelial inhibition ranging from 12.48% to 64.45%. Specifically, the co-culture suspension of T. asperellum 576 + E. turcicum 101 exhibited effective mycelial growth inhibition of E. turcicum 101 exceeding 50.00% from day 5 to 9. Similarly, the co-culture suspension of T. atroviride 393 + E. turcicum 101 showed effective mycelial growth inhibition exceeding 50.00% from days 6 to 9. These findings highlight the strong antifungal action of these co-culture suspensions, further emphasizing the potential of T. asperellum 576 and T. atroviride 393 as effective agents for inhibiting the growth of E. turcicum 101.
3.6. Activity of Cell Wall Degradation Enzymes
The activity of cell wall degrading enzymes (chitinase, β-1,3-glucanase, and cellulase) in the co-culture suspensions of T. asperellum 576 + E. turcicum 101 or T. atroviride 393 + E. turcicum 101 was investigated. The chitinase activity in the co-culture suspension of T. asperellum 576 + E. turcicum 101 increased gradually until day 6 after treatment and reached its peak at day 9 with a value of 8.34 U/mL. On the other hand, the highest chitinase activity of 2.41 U/mL was observed on day 7 after treatment with the co-culture suspension of T. atroviride 393 + E. turcicum 101 (Figure 6A). The β-1,3-glucanase activity in the co-culture suspension of T. asperellum 576 + E. turcicum 101 also showed a gradual increase until day 3 after treatment and peaked at day 7 with a value of 3.42 U/mL. Similarly, the highest β-1,3-glucanase activity of 2.31 U/mL was detected on day 7 after treatment with the co-culture suspension of T. atroviride 393 + E. turcicum 101 (Figure 6B). As for cellulase activity, the co-culture suspension of T. asperellum 576 + E. turcicum 101 exhibited a gradual increase until day 6 after treatment and reached its peak at day 7 with a value of 7.75 U/mL. The highest cellulase activity of 4.14 U/mL was observed on day 4 after treatment with the co-culture suspension of T. atroviride 393 + E. turcicum 101 (Figure 6C). Comparatively, T. asperellum 576 showed stronger cell wall activities against E. turcicum 101 than T. atroviride 393. Therefore, T. asperellum 576 was selected for in vivo tests.
3.7. Stimulating Germination of Maize Seeds by T. asperellum 576
The impact of T. asperellum 576 conidial suspension on maize seed germination was evaluated compared to an untreated control group. T. asperellum 576 conidial suspension showed a significant promotion of seed germination compared to the inoculation control. As depicted in Figure 7, the seed germination rate of the control group was 50.90%. Among the T. asperellum 576 conidial suspension, there are forty-four maize seeds germinating, with rates reaching 91.00%. Based on these results, T. asperellum 576 was selected for further experiments.
3.8. Effect of T. asperellum 576 on E. turcicum 101 in Maize Seedlings
Under greenhouse conditions, the biocontrol effect of T. asperellum 576 against E. turcicum 101 on maize was evaluated. The performance of T. asperellum 576 in terms of plant growth promotion and suppression of E. turcicum 101 was assessed. These included four treatments: (T1) plants without T. asperellum 576 and E. turcicum 101 (healthy control), (T2) T. asperellum 576-treated plants, (T3) E. turcicum 101-inoculated plants (disease control), and (T4) E. turcicum 101 and T. asperellum 576-inoculated/treated plants.
As shown in Table 5 and Figure 8, after 20 days, the maize seedlings treated with T. asperellum 576 exhibited significant improvements in shoot height, stem diameter, shoot fresh weight, root fresh weight, shoot dry weight, and root dry weight compared to the T1 control plants. The maize seedlings treated with T. asperellum 576 showed increases of 17.14%, 18.18%, 52.55%, 35.76%, 48.50%, and 47.27% in these parameters, respectively, indicating enhanced growth due to the presence of T. asperellum 576.
As shown in Table 5 and Figure 8, the maize seedlings inoculated with both E. turcicum 101 and T. asperellum 576 showed significant enhancements in shoot height, stem diameter, fresh and dry shoot weight, and fresh and dry root weight compared to the maize seedlings inoculated with E. turcicum 101. The promotion rates for these parameters in the maize seedlings inoculated with both E. turcicum 101 and T. asperellum 576 were 8.85%, 15.91%, 46.07%, 69.09%, 71.84%, and 94.52%, respectively. The blight of maize seedlings inoculated with both E. turcicum 101 and T. asperellum 576 was observed to be slighter than the maize seedlings inoculated with E. turcicum 101.
These results indicate that the application of T. asperellum 576 spore suspension effectively prevented the growth of E. turcicum 101 and promoted the growth of maize seedlings under greenhouse conditions.
4. Discussion
4.1. Evidence of Direct Mycoparasitism and Intraspecies Variability in Trichoderma-E. turcicum Interaction
In the dual culture test, a total of 12 strains of Trichoderma exhibited pronounced antagonistic activity. This phenomenon can potentially be attributed to the rapid growth rate of these Trichoderma strains in comparison to E. turcicum 101. T. asperellum 576 demonstrated the highest level of antifungal activity against E. turcicum 101 in the dual culture experiment. Moreover, we also observed significant variations between two Trichoderma strains belonging to the same species. This intraspecies variability aligns with findings reported by El Gamal et al. [53]. In order to comprehensively evaluate the impact of growth rate and mycoparasitism, we employed the evaluation methods described by Zhang et al. [47]. This method does not solely rely on assessing the inhibition percentage but instead considers both factors holistically, categorized as fast, moderate, or slow in comparison to pathogens. Consequently, these varying growth rates among Trichoderma strains may significantly influence the competition for nutrients and space, ultimately leading to the outcompeting of E. turcicum 101.
To gather additional evidence supporting direct mycoparasitism, a comprehensive analysis of microscopic images was conducted. A significant finding emerged wherein T. asperellum 576 hyphae were observed to encircle the hyphae of E. turcicum 101. These images were the mycelium of both T. asperellum 576 and E. turcicum 101. This discovery aligns with earlier research documenting physical contacts between hyphae, the abundant sporulation of Trichoderma on the hyphae of the pathogen, and the development of perforations when Trichoderma species interact with other pathogens [54,55]. Furthermore, the extensive literature confirms the species- and strain-dependent variability in the coiling behavior exhibited by different Trichoderma species. This observation provides additional support for the notion that the interaction between Trichoderma and its host is characterized by specificity [56,57].
4.2. Non-Volatile Metabolites of Trichoderma spp.: Potent Inhibitors of E. turcicum 101 and Antibiosis as a Key Biocontrol Mechanism
The antifungal activity of non-volatile metabolites derived from the tested Trichoderma spp. exhibited a significant growth inhibitory effect against E. turcicum 101. Previous studies have already established T. asperellum as an effective BCA against various plant-pathogenic fungi [58,59,60]. Consistent with these findings, our study demonstrated that T. asperellum 576 displayed strong inhibition of E. turcicum 101 growth (80.81% inhibition), and T. atroviride 393 also exhibited notable effectiveness against E. turcicum 101 (77.68% inhibition). Other Trichoderma species also exhibited varying degrees of radial growth inhibition against the tested pathogens, albeit to a lesser extent. Inhibitory action of these Trichoderma against the test pathogens primarily occurs through the production of non-volatile compounds, emphasizing the central role of antibiosis as the principal biocontrol mechanism [61]. Antibiosis refers to the antagonistic activity mediated by specific or non-specific metabolites of microbial origin, lytic enzymes, volatile compounds, and other toxic substances. It is noteworthy that Trichoderma spp. is known to synthesize various antibiotics, including trichodernin, trichodermol, and herzianolide, among others [62].
4.3. Unveiling the Pole of Trichoderma Volatile Substances in Inhibiting E. turcicum 101: A Potential Mechanism for Enhanced Biological Control
An additional potential mechanism that may contribute to the inhibition of E. turcicum 101 is the production of volatile substances. In our study, the results revealed significant suppressive effects, with T. asperellum 576-VOCs demonstrating the highest antifungal properties (65.86% inhibition) against E. turcicum 101. Conversely, T. longibrachiatum 3A displayed the lowest antifungal effect, with only 23.10%. These findings suggest that Trichoderma VOCs possess the potential to provide additional benefits in terms of biological control and promoting plant growth in sustainable agricultural practices. Notably, the inhibition rates of volatile metabolites do not necessarily correlate strongly with those of non-volatile metabolites, as the antagonistic effects involve various factors beyond the strain’s characteristics [63]. Previous research conducted by Amin et al. has shown that Trichoderma-derived VOCs can negatively affect the mycelial growth of various pathogenic fungi, including Alternaria brassicicola, Rhizoctonia solani, and Fusarium oxysporum [64]. Furthermore, several studies have documented the antifungal activity of Trichoderma-derived secondary metabolites. It is plausible that our Trichoderma strains also produce antifungal metabolites that diffuse through the agar, leading to a reduction in the mycelium growth of E. turcicum 101 [65,66]. To gain a deeper understanding, further investigations are required to isolate and purify the individual components present in these extracts and evaluate the spectrum of antifungal activity exhibited by these metabolites. Overall, it is likely that the observed inhibition of E. turcicum 101 is the result of a combination of different mechanisms at play involving both non-volatile and volatile compounds produced by Trichoderma strains.
4.4. Potent Antifungal Activity of T. asperellum 576 in Co-Culture with E. turcicum 101: Implications for Endophytic Environment Establishment
In this study, we conducted experiments using co-culture suspensions of T. asperellum 576 + E. turcicum 101 and T. atroviride 393 + E. turcicum 101 to assess their impact on the growth of E. turcicum 101. Notably, the T. asperellum 576 + E. turcicum 101 co-culture exhibited the most pronounced effect, demonstrating a significant broad-spectrum antifungal activity against E. turcicum 101 on the 7th day, resulting in a remarkable 64.45% inhibition. These findings strongly indicate the potent inhibitory effect of T. asperellum 576 on the growth of E. turcicum 101. Moreover, there is compelling evidence suggesting that the filtrates derived from these co-cultures may contain small plant-derived molecules that contribute to the establishment of an environment resembling a typical endophytic habitat [67]. Such an environment provides favorable conditions for the proliferation of microorganisms exhibiting endophytic behavior, including Trichoderma spp.
4.5. Role of Cell Wall-Degrading Enzymes in the Biocontrol Potential of T. asperellum 576 against E. turcicum 101
Continuing our investigation into the mechanisms involved in the biocontrol of E. turcicum 101, we conducted an evaluation of the chitinase, β-1,3-glucanase, and cellulase activities of T. asperellum 576. These enzymatic activities are known to play a critical role in the mycoparasitic activity exhibited by Trichoderma against pathogens. Our findings reveal that these cell wall-degrading enzymes do not act in isolation but require the concerted action of multiple chitinases to effectively break down fungal cell walls [68]. Trichoderma species have the ability to secrete hydrolytic enzymes that specifically target and degrade the cell walls of host fungi [24]. Extensive research has previously highlighted the remarkable capacity of the Trichoderma genus to produce a diverse range of hydrolytic enzymes and secondary metabolites, which contribute to the suppression of various pathogenic organisms [69]. Our study further demonstrated that the cell wall composition of E. turcicum 101 has a regulatory effect on the activities of T. asperellum 576 associated with mycoparasitism, including the production of β-1,3-glucanases, chitinases, and cellulase. These findings are consistent with the work of Aoki et al. [70], who reported the production and secretion of chitinase by Trichoderma sp. SANA20 in the culture media after three days of incubation. Moreover, our results align with previous studies that have shown significant chitinase and β-1,3-glucanase activities in the cell-free culture filtrate of T. asperellum [71,72,73]. Collectively, these findings suggest that T. asperellum 576 actively produces and secretes cell wall-degrading enzymes, which likely contribute to the suppression of fungal pathogens. The assessment of cell wall enzyme activities can serve as an indicator for determining antagonistic Trichoderma strains, with T. asperellum 576 exhibiting particularly high expression levels among the tested strains. This discovery holds significant importance in the identification of effective mycoparasites for biocontrol strategies.
4.6. Enhancing Maize Seed Germination with T. asperellum 576: Towards Optimal Spore Dose for Targeted Plant Growth Promotion
This study provides empirical support for the effect of applying T. asperellum 576 at a concentration of 107 spores/mL on maize seeds, leading to a notable enhancement in seed germination compared to water-treated maize seeds (88% germination rate). These findings align with the observations made by Singh et al., who documented variations in the spore dose requirements of T. asperellum BHUT8 for promoting the initial growth of different vegetable crops, encompassing seed germination and radicle length [74]. In future investigations, our objective is to develop an innovative approach to determine the precise spore dose of T. asperellum 576 necessary to optimize plant growth on maize seeds. This research endeavor will yield valuable insights into refining the application of T. asperellum 576 for targeted plant growth promotion, specifically tailored to maize cultivation.
4.7. Exploring the Potential of T. asperellum 576 for Plant Growth Promotion and Disease Suppression
Previous research has extensively explored the direct effects of Trichoderma-based biopesticides or Trichoderma spore suspensions on various aspects of plant growth promotion, defense responses, and stress tolerance [75,76,77,78,79,80]. In greenhouse conditions, the application of T. asperellum 576 significantly enhanced the growth of maize seedlings (p < 0.05). These findings align with multiple studies indicating that the application of Trichoderma spp. leads to notable improvements in root length, shoot length, and dry weight compared to control plants [24,37,81]. One of the mechanisms attributed to the plant growth promotion capabilities of plant growth-promoting microorganisms’ strains is the production of growth-promoting hormones such as indole-3-acetic acid (IAA), cytokinins, and gibberellins, which stimulate plant growth [82]. In our study, we demonstrated that the treatment of maize seedlings with T. asperellum 576 resulted in increased fresh and dry weight, as well as lateral root length. These findings strongly suggest that T. asperellum 576 can effectively influence plant growth and dry matter accumulation. We speculate that the observed growth-promoting activity of plants might be mediated by hormones induced by T. asperellum 576. Future studies should investigate the reproducibility of the growth-promoting abilities of T. asperellum 576 under field conditions, following observations made in plants grown under greenhouse conditions with soil application. This line of research will provide valuable insights into the practical applicability of T. asperellum 576 as a plant growth promoter in real-world agricultural settings. Such knowledge is crucial for assessing its potential as a sustainable and effective tool for enhancing crop productivity.
In our investigation, the utilization of both E. turcicum 101 and T. asperellum 576 in treating maize seedlings observed a significant reduction in lesion area and an improved efficacy in controlling the impact of E. turcicum compared to seedlings treated solely with E. turcicum 101. These findings highlight the potential advantages of employing a combined treatment strategy, showcasing the synergistic effects of these two fungal strains in mitigating the detrimental consequences of E. turcicum 101 on maize plants. Interestingly, our study also revealed contrasting outcomes when maize seedlings were co-inoculated with E. turcicum 101 and T. asperellum 576. In this case, we observed a significant detrimental effect on the growth of the seedlings across various measured parameters, in contrast to the treatment with E. turcicum 101 alone. However, it is noteworthy that the maize seedlings subjected to the combined treatment of E. turcicum 101 and T. asperellum 576 displayed substantial improvements in shoot height, stem diameter, shoot fresh weight, root fresh weight, shoot dry weight, and root dry weight when compared to those treated solely with E. turcicum 101. These findings underscore the complexity of interactions among different microbial strains and their effects on plant health and growth. Further investigations are warranted to elucidate the underlying mechanisms driving the observed effects and to optimize the application strategies of these fungal strains for effective disease management in maize cultivation. In the current study, we observed remarkable growth inhibition of E. turcicum 101, the causal agent of northern corn leaf blight, in maize seedlings treated with T. asperellum 576. This effective control of the pathogen resulted in alleviating the symptoms associated with the disease. Previous studies have highlighted the potential of T. asperellum as a promising BCA against plant pathogens, and T. asperellum has been recognized for its capacity to enhance plant growth when introduced to plant roots, as demonstrated in a previous investigation [37,83,84,85,86]. The diverse biocontrol mechanisms demonstrated by Trichoderma spp. are pivotal in their efficacy for plant disease management and growth promotion across diverse contexts. Pathogen infections often induce significant morphological and physiological changes in plants, including the appearance of mosaic symptoms, which are closely linked to alterations in chlorophyll content and subsequent reductions in photosynthesis [87,88]. Consequently, our future investigations will assess changes in chlorophyll pigment content following E. turcicum 101 infection, thereby providing valuable insights into the physiological response of maize seedlings to pathogen-induced stress.
The current study provides compelling evidence supporting the effect of T. asperellum 576 isolates in suppressing the growth of E. turcicum 101 and inhibiting microsclerotia production while simultaneously promoting plant growth. Our findings contribute to a better understanding of the underlying mechanisms involved in the suppression of pathogenic activity by E. turcicum 101. These notable characteristics of the antagonistic strain can be effectively utilized in the development of future formulations and in preliminary experiments for the large-scale production of a bioinoculant containing T. asperellum 576 as the primary active component. We hypothesize that Trichoderma produces components that induce the synthesis of antimicrobial metabolites and cell wall degradation enzymes. This strategic approach can expedite and ensure the high efficiency of biocontrol when applied to maize plants. By synergistically combining the direct effects of pre-existing antimicrobial metabolites and cell wall degradation enzymes produced by T. asperellum 576 with the production of these components at the sites of microbial formulation application, the biocontrol activity can be significantly potentiated.
5. Conclusions
The study demonstrated the potential of T. asperellum 576 as a biocontrol agent against E. turcicum 101 and a growth promoter for maize. T. asperellum 576 showed strong antagonism and inhibitory effects on E. turcicum 101, both through non-volatile and volatile substances. It also exhibited enzymatic activities for degrading fungal cell walls. Additionally, T. asperellum 576 stimulated maize seed germination and enhanced plant growth. Overall, the findings suggest that T. asperellum 576 has promising potential as a biocontrol agent for disease management and as a bioenhancer for promoting the growth of maize.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9090936/s1, Figure S1: Forty-four Trichoderma strains used in this study cultures at 25 °C after 7 days; Figure S2: Trichoderma strains with moderate antagonistic (left) and E. turcicum 101 (right) strains in dual culture. 1–3: 110; 4–6: 224; 7–9: 295; 10–12: 375; 13–15: 99; 16–18: 421; 19–21: 539; 22–24: 581; 25–27: 593; 28–30: 526; 31–33: 578; 34–36: XZ9-1; Figure S3: Trichoderma with weakly antagonistic (left) and E. turcicum 101 (right) strains in dual culture. 1–3: 153; 4–6: 218; 7–9: 219; 10–12: 220; 13–15: 370; 16–18: 439; 19–21: 588; 22–24: 1408; 25–27: L-3; 28–30: 592; Figure S4: Trichoderma without antagonistic (left) and E. turcicum 101 (right) strains in dual culture. 1–3: 115; 4–6: 299; 7–9: 338; 10–12: 376; 13–15: 438; 16–18: 449; 19–21: 509; 22–24: 1283; 25–27: XZ5-2, 28-30: 91.
Author Contributions
Conceptualization, Y.M. and Z.Z.; experimental design and methodology, Y.M. and Z.Z.; performance of practical work, Y.M.; statistical analysis, Y.M., Y.L. (Yetong Li) and S.Y.; validation, Y.M. and Y.L. (Yetong Li); writing—original draft preparation, Y.M.; writing—review and editing, Z.Z.; supervision, Y.L. (Yu Li); project administration, Z.Z.; funding acquisition, Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Science and Technology Development Plan Project of Jilin Province (No. 20230402021GH) and the National Key Research and Development Program of China (grant number 2018YFE0107800).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author, Yukun Ma, expresses her gratitude to Hanxing Gao and Xiaoya An for their valuable contributions and assistance during the experiment.
Conflicts of Interest
The authors state no conflict of interest.
References
- Luo, N.; Meng, Q.; Feng, P.; Qu, Z.; Yu, Y.; Liu, D.L.; Müller, C.; Wang, P. China can be self-sufficient in maize production by 2030 with optimal crop management. Nat. Commun. 2023, 14, 2637. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Bodirsky, B.L.; Dietrich, J.P.; Martinelli, E.; Stenstad, A.; Pradhan, P.; Gabrysch, S.; Mishra, A.; Weindl, I.; Le Mouël, C.; Rolinski, S.; et al. The ongoing nutrition transition thwarts long-term targets for food security, public health and environmental protection. Sci. Rep. 2020, 10, 19778. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Tong, L.; Kang, S.; Du, T.; Ding, R.; Li, S.; Chen, Y. Combination of suitable planting density and nitrogen rate for high yield maize and their source-sink relationship in Northwest China. J. Sci. Food Agric. 2023, 103, 5300–5311. [Google Scholar] [CrossRef] [PubMed]
- Serna-Saldivar, S.O.; Perez Carrillo, E. Chapter 16—Food uses of whole corn and dry-milled fractions. In Corn, 3rd ed.; Serna-Saldivar, S.O., Ed.; AACC International Press: Oxford, UK, 2019; pp. 435–467. [Google Scholar]
- Badu-Apraku, B.; Bankole, F.A.; Ajayo, B.S.; Fakorede, M.A.B.; Akinwale, R.O.; Talabi, A.O.; Bandyopadhyay, R.; Ortega-Beltran, A. Identification of early and extra-early maturing tropical maize inbred lines resistant to Exserohilum turcicum in sub-Saharan Africa. Crop Prot. 2021, 139, 105386. [Google Scholar] [CrossRef]
- Manu, T.G.; Naik, B.G.; Sayipratap, B.R.; Balagar, M.S. Chemical science review and letters efficacy of fungicides, botanicals and bio-agents against Exserohilum turcicum. Chem. Sci. Rev. Lett. 2017, 6, 2100–2107. [Google Scholar]
- Huang, W.; Fang, X.; Wang, H.; Chen, F.; Duan, H.; Bi, Y.; Yu, H. Biosynthesis of AgNPs by Bipolaris maydis and its antifungal effect against Exserohilum turcicum. IET Nanobiotechnol. 2018, 12, 585–590. [Google Scholar] [CrossRef]
- CABI. Crop Protection Compendium: Setosphaeria turcica (Maize Leaf Blight); CABI International CABI: Wallingford, UK, 2022; Volume CABI Compendium. [Google Scholar]
- Navarro, B.L.; Campos, R.D.; Gasparoto, M.C.D.; von Tiedemann, A. In Vitro and In Planta Studies on Temperature Adaptation of Exserohilum turcicum Isolates from Maize in Europe and South America. Pathogens 2021, 10, 154. [Google Scholar] [CrossRef]
- Balint-Kurti, P.J.; Yang, J.Y.; Van Esbroeck, G.; Jung, J.; Smith, M.E. Use of a maize advanced intercross line for mapping of QTL for northern leaf blight resistance and multiple disease resistance. Crop Sci. 2010, 50, 458–466. [Google Scholar] [CrossRef]
- Mueller, D.S.; Wise, K.A.; Sisson, A.J.; Allen, T.W.; Wiebold, W.J. Corn yield loss estimates due to diseases in the United States and Ontario, Canada, from 2016 to 2019. Plant Health Prog. 2020, 21, 238–247. [Google Scholar] [CrossRef]
- Weems, J.D.; Bradley, C.A. Sensitivity of Exserohilum turcicum to demethylation inhibitor fungicides. Crop Prot. 2017, 99, 85–92. [Google Scholar] [CrossRef]
- Mahrous, N.N.; Columbus, M.P.; Southam, G.; Macfie, S.M. Changes in microbial community structure and increased metal bioavailability in a metal-contaminated soil and in the rhizosphere of corn (Zea mays). Rhizosphere 2019, 11, 100169. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Chapter 5—Actinobacteria: Eco-friendly candidates for control of plant diseases in a sustainable manner. In New and Future Developments in Microbial Biotechnology and Bioengineering; Singh, B.P., Gupta, V.K., Passari, A.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 79–91. [Google Scholar]
- Fazeli-Nasab, B.; Shahraki-Mojahed, L.; Piri, R.; Sobhanizadeh, A. 20—Trichoderma: Improving growth and tolerance to biotic and abiotic stresses in plants. In Trends of Applied Microbiology for Sustainable Economy; Soni, R., Suyal, D.C., Yadav, A.N., Goel, R., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 525–564. [Google Scholar]
- Adnan, M.; Islam, W.; Shabbir, A.; Khan, K.A.; Ghramh, H.A.; Huang, Z.; Chen, H.Y.H.; Lu, G.-d. Plant defense against fungal pathogens by antagonistic fungi with Trichoderma in focus. Microb. Pathog. 2019, 129, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Sartori, M.; Nesci, A.; Formento, Á.; Etcheverry, M. Selection of potential biological control of Exserohilum turcicum with epiphytic microorganisms from maize. Rev. Argent. Microbiol. 2015, 47, 62–71. [Google Scholar] [CrossRef]
- Das, P.P.; Singh, K.R.B.; Nagpure, G.; Mansoori, A.; Singh, R.P.; Ghazi, I.A.; Kumar, A.; Singh, J. Plant-soil-microbes: A tripartite interaction for nutrient acquisition and better plant growth for sustainable agricultural practices. Environ. Res. 2022, 214, 113821. [Google Scholar] [CrossRef]
- Emmanuel, O.C.; Babalola, O.O. Productivity and quality of horticultural crops through co-inoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria. Microbiol. Res. 2020, 239, 126569. [Google Scholar] [CrossRef]
- Muhie, S.H. Novel approaches and practices to sustainable agriculture. J. Agric. Food Res. 2022, 10, 100446. [Google Scholar] [CrossRef]
- Ahmad, F.; Saeed, Q.; Shah, S.M.U.; Gondal, M.A.; Mumtaz, S. Chapter 11—Environmental sustainability: Challenges and approaches. In Natural Resources Conservation and Advances for Sustainability; Jhariya, M.K., Meena, R.S., Banerjee, A., Meena, S.N., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 243–270. [Google Scholar]
- Weindling, R. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology 1932, 22, 837–845. [Google Scholar]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Samuels, G.J. Trichoderma: Systematics, the sexual state, and ecology. Phytopathology 2006, 96, 195–206. [Google Scholar] [CrossRef]
- Asad, S.A. Mechanisms of action and biocontrol potential of Trichoderma against fungal plant diseases—A review. Ecol. Complex. 2022, 49, 100978. [Google Scholar] [CrossRef]
- Lin, X.; Tang, Z.; Gan, Y.; Li, Z.; Luo, X.; Gao, C.; Zhao, L.; Chai, L.; Liu, Y. 18-Residue peptaibols produced by the sponge-derived Trichoderma sp. GXIMD 01001. J. Nat. Prod. 2023, 86, 994–1002. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Tang, W.L.; Huang, Q.R.; Li, Y.Z.; Wei, M.L.; Jiang, L.L.; Liu, C.; Yu, X.; Zhu, H.W.; Chen, G.Z.; et al. Trichoderma: A treasure house of structurally diverse secondary metabolites with medicinal importance. Front. Microbiol. 2021, 12, 723828. [Google Scholar] [CrossRef] [PubMed]
- Dennis, C.; Webster, J. Antagonistic properties of species-groups of Trichoderma: I. Production of non-volatile antibiotics. Trans. Br. Mycol. Soc. 1971, 57, 25-IN3. [Google Scholar] [CrossRef]
- Li, M.-F.; Li, G.-H.; Zhang, K.-Q. Non-Volatile Metabolites from Trichoderma spp. Metabolites 2019, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ghirardo, A.; Weber, B.; Schnitzler, J.-P.; Benz, J.P.; Rosenkranz, M. Trichoderma Species Differ in Their Volatile Profiles and in Antagonism Toward Ectomycorrhiza Laccaria bicolor. Front. Microbiol. 2019, 10, 891. [Google Scholar] [CrossRef]
- Kottb, M.; Gigolashvili, T.; Großkinsky, D.K.; Piechulla, B. Trichoderma volatiles effecting Arabidopsis: From inhibition to protection against phytopathogenic fungi. Front. Microbiol. 2015, 6, 995. [Google Scholar] [CrossRef]
- Lee, S.; Behringer, G.; Hung, R.; Bennett, J. Effects of fungal volatile organic compounds on Arabidopsis thaliana growth and gene expression. Fungal Ecol. 2019, 37, 1–9. [Google Scholar] [CrossRef]
- Siddiquee, S. Chapter 11—Recent Advancements on the Role and Analysis of Volatile Compounds (VOCs) from Trichoderma. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 139–175. [Google Scholar]
- Limdolthamand, S.; Songkumarn, P.; Suwannarat, S.; Jantasorn, A.; Dethoup, T. Biocontrol efficacy of endophytic Trichoderma spp. in fresh and dry powder formulations in controlling northern corn leaf blight in sweet corn. Biol. Control 2023, 181, 105217. [Google Scholar] [CrossRef]
- Degani, O.; Rabinovitz, O.; Becher, P.; Gordani, A.; Chen, A. Trichoderma longibrachiatum and Trichoderma asperellum confer growth promotion and protection against late wilt disease in the field. J. Fungi 2021, 7, 444. [Google Scholar] [CrossRef]
- Shang, J.; Liu, B.; Xu, Z. Efficacy of Trichoderma asperellum TC01 against anthracnose and growth promotion of Camellia sinensis seedlings. Biol. Control 2020, 143, 104205. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Samuels, G.J.; Ismaiel, A.; Voglmayr, H. Disentangling the Trichoderma viridescens complex. Persoonia 2013, 31, 112–146. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.X.; Zhuang, W.Y. Trichoderma (Hypocrea) species with green ascospores from China. Persoonia 2015, 34, 113–129. [Google Scholar] [CrossRef]
- Chaverri, P.; Samuels, G.J. Hypocrea/Trichoderma (Ascomycota, Hypocreales, Hypocreaceae): Species with green ascospores. Stud. Mycol. 2003, 48, 1–116. [Google Scholar]
- Jaklitsch, W.M.; Komon, M.; Kubicek, C.P.; Druzhinina, I.S. Hypocrea voglmayrii sp. nov. from the Austrian Alps represents a new phylogenetic clade in Hypocrea/Trichoderma. Mycologia 2005, 97, 1365–1378. [Google Scholar] [CrossRef] [PubMed]
- Jaklitsch, W.M. European species of Hypocrea Part I. The green-spored species. Stud. Mycol. 2009, 63, 1–91. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Bunbury-Blanchette, A.L.; Walker, A.K. Trichoderma species show biocontrol potential in dual culture and greenhouse bioassays against Fusarium basal rot of onion. Biol. Control 2019, 130, 127–135. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Zhuang, W.Y. First step evaluation of Trichoderma antagonism against plant pathogenic fungi in dual culture. Mycosystema 2017, 36, 1251–1259. [Google Scholar]
- Joo, J.H.; Hussein, K.A. Biological control and plant growth promotion properties of volatile organic compound-producing antagonistic Trichoderma spp. Front. Plant Sci. 2022, 13, 897668. [Google Scholar] [CrossRef] [PubMed]
- Di Lelio, I.; Coppola, M.; Comite, E.; Molisso, D.; Lorito, M.; Woo, S.L.; Pennacchio, F.; Rao, R.; Digilio, M.C. Temperature differentially influences the capacity of Trichoderma species to induce plant defense responses in tomato against insect pests. Front. Plant Sci. 2021, 12, 678830. [Google Scholar] [CrossRef] [PubMed]
- Sha, R.; Jiang, L.; Meng, Q.; Zhang, G.; Song, Z. Producing cell-free culture broth of rhamnolipids as a cost-effective fungicide against plant pathogens. J. Basic Microbiol. 2012, 52, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 420–428. [Google Scholar] [CrossRef]
- Singh, D.B.; Gupta, R.K.; Singh, R.; Patil, R. Minimal processing of papaya for quality maintenance and shelf life. Acta Hortic. 2010, 851, 579–590. [Google Scholar] [CrossRef]
- EI-gamal, d.n.; Atalla, S.; El-Mohamedy, R.R.S. Improvement in potential of enzymes from Chaetomium globosum and Trichoderma harzianum using different agricultural wastes and its applications. Biosci. Res. 2018, 15, 3977–3987. [Google Scholar]
- Almeida, F.B.; Cerqueira, F.M.; Silva Rdo, N.; Ulhoa, C.J.; Lima, A.L. Mycoparasitism studies of Trichoderma harzianum strains against Rhizoctonia solani: Evaluation of coiling and hydrolytic enzyme production. Biotechnol. Lett. 2007, 29, 1189–1193. [Google Scholar] [CrossRef]
- Huang, X.; Chen, L.; Ran, W.; Shen, Q.; Yang, X. Trichoderma harzianum strain SQR-T37 and its bio-organic fertilizer could control Rhizoctonia solani damping-off disease in cucumber seedlings mainly by the mycoparasitism. Appl. Microbiol. Biotechnol. 2011, 91, 741–755. [Google Scholar] [CrossRef]
- Monteiro, V.N.; do Nascimento Silva, R.; Steindorff, A.S.; Costa, F.T.; Noronha, E.F.; Ricart, C.A.; de Sousa, M.V.; Vainstein, M.H.; Ulhoa, C.J. New insights in Trichoderma harzianum antagonism of fungal plant pathogens by secreted protein analysis. Curr. Microbiol. 2010, 61, 298–305. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuang, W.-Y. Trichoderma brevicrassum strain TC967 with capacities of diminishing cucumber disease caused by Rhizoctonia solani and promoting plant growth. Biol. Control 2020, 142, 104151. [Google Scholar] [CrossRef]
- Pradhan, D.A.; Bagagoni, P.; Makandar, R. Assessing rhizosphere Trichoderma asperellum strains for root colonizing and antagonistic competencies against Fusarium wilt through molecular and biochemical responses in castor. Biol. Control 2023, 184, 105280. [Google Scholar] [CrossRef]
- Abbas, A.; Mubeen, M.; Zheng, H.; Sohail, M.A.; Shakeel, Q.; Solanki, M.K.; Iftikhar, Y.; Sharma, S.; Kashyap, B.K.; Hussain, S.; et al. Trichoderma spp. genes involved in the biocontrol activity against Rhizoctonia solani. Front. Microbiol. 2022, 13, 884469. [Google Scholar] [CrossRef] [PubMed]
- Mbarga, J.B.; Begoude, B.A.D.; Ambang, Z.; Meboma, M.; Kuate, J.; Ewbank, W.; Hoopen, G.M.t. Field testing an oil-based Trichoderma asperellum formulation for the biological control of cacao black pod disease, caused by Phytophthora megakarya. Crop Prot. 2020, 132, 105134. [Google Scholar] [CrossRef]
- Shanmugam, V.; Varma, A.S. Effect of native antagonists against Pythium aphanidermatum, the casual organism of rhizome rot of ginger. J. Mycol. Pl. Pathol. 1999, 29, 375–379. [Google Scholar]
- Kucuk, C.; Kivanc, M. In vitro antifungal activity of strains of Trichoderma harzianum. Turk. J. Biol. 2004, 28, 111–115. [Google Scholar]
- Lee, J.; Huh, N.; Hong, J.H.; Kim, B.S.; Kim, G.-H.; Kim, J.-J. The antagonistic properties of Trichoderma spp. inhabiting woods for potential biological control of wood-damaging fungi. Holzforschung 2012, 66, 883–887. [Google Scholar] [CrossRef]
- Amin, F.; Razdan, V.; Mohiddin, F.; Bhat, K.; Sheikh, P. Effect of volatile metabolites of Trichoderma species against seven fungal plant pathogens In-Vitro. J. Phytol. 2010, 2, 34–37. [Google Scholar]
- Reino, J.L.; Guerrero, R.F.; Hernández-Galán, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.; Woo, S.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Ruocco, M.; Lanzuise, S.; et al. Trichoderma secondary metabolites active on plants and fungal pathogens. Open Mycol. J. 2014, 8, 127–139. [Google Scholar] [CrossRef]
- Murphy, B.; Batke, S.; Doohan, F.; Hodkinson, T. Media manipulations and the culture of beneficial fungal root endophytes. Int. J. Biol. 2015, 7, 94–102. [Google Scholar] [CrossRef]
- Carsolio, C.; Benhamou, N.; Haran, S.; Cortés, C.; Gutiérrez, A.; Chet, I.; Herrera-Estrella, A. Role of the Trichoderma harzianum endochitinase gene, ech42, in mycoparasitism. Appl. Environ. Microbiol. 1999, 65, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Daguerre, Y.; Siegel, K.; Edel-Hermann, V.; Steinberg, C. Fungal proteins and genes associated with biocontrol mechanisms of soil-borne pathogens: A review. Fungal Biol. Rev. 2014, 28, 97–125. [Google Scholar] [CrossRef]
- Ano, A.; Takayanagi, T.; Uchibori, T.; Okuda, T.; Yokotsuka, K. Characterization of a class III chitinase from Vitis vinifera cv. Koshu. J. Biosci. Bioeng. 2003, 95, 645–647. [Google Scholar] [CrossRef] [PubMed]
- Bech, L.; Busk, P.; Lange, L. Cell wall degrading enzymes in Trichoderma asperellum grown on wheat bran. Fungal Genom. Biol. 2014, 4, 1000116. [Google Scholar]
- Moussa, Z.; Alanazi, Y.F.; Khateb, A.M.; Eldadamony, N.M.; Ismail, M.M.; Saber, W.I.A.; Darwish, D.B.E. Domiciliation of Trichoderma asperellum suppresses Globiosporangium ultimum and promotes pea growth, ultrastructure, and metabolic features. Microorganisms 2023, 11, 198. [Google Scholar] [CrossRef]
- Batool, R.; Umer, M.J.; Shabbir, M.Z.; Wang, Y.; Ahmed, M.A.; Guo, J.; He, K.; Zhang, T.; Bai, S.; Chen, J.; et al. Seed Myco-priming improves crop yield and herbivory induced defenses in maize by coordinating antioxidants and Jasmonic acid pathway. BMC Plant Biol. 2022, 22, 554. [Google Scholar] [CrossRef]
- Singh, V.; Upadhyay, R.S.; Sarma, B.K.; Singh, H.B. Trichoderma asperellum spore dose depended modulation of plant growth in vegetable crops. Microbiol. Res. 2016, 193, 74–86. [Google Scholar] [CrossRef]
- Björkman, T.; Blanchard, L.; Harman, G. Growth enhancement of shrunken-2 (sh2) sweet corn by Trichoderma harzianum 1295-22: Effect of environmental stress. J. Am. Soc. Hortic. Sci. 1998, 123, 35–40. [Google Scholar] [CrossRef]
- Batta, Y.A. Effect of treatment with Trichoderma harzianum Rifai formulated in invert emulsion on postharvest decay of apple blue mold. Int. J. Food Microbiol. 2004, 96, 281–288. [Google Scholar] [CrossRef]
- Mastouri, F.; Björkman, T.; Harman, G.E. Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Singh, A.; Chaudhary, A.; Singh, S.; Singh, H.B. Modulation of nutritional and antioxidant potential of seeds and pericarp of pea pods treated with microbial consortium. Food Res. Int. 2014, 64, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Schmoll, M.; Herrera-Estrella, A.; Upadhyay, R.S.; Druzhinina, I.; Tuohy, M. Biotechnology and Biology of Trichoderma; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–549. [Google Scholar]
- Singh, A.; Sarma, B.K.; Singh, H.B.; Upadhyay, R.S. Chapter 40—Trichoderma: A silent worker of plant Rhizosphere. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 533–542. [Google Scholar]
- Abdelkhalek, A.; Hafez, E.; Aboelhana, E. Polyphenolic genes expression pattern and their role in viral resistance in tomato plant infected with Tobacco mosaic virus. Biosci. Res. 2019, 4, 3349–3356. [Google Scholar]
- Rubio, M.B.; Monti, M.M.; Gualtieri, L.; Ruocco, M.; Hermosa, R.; Monte, E. Trichoderma harzianum volatile organic compounds regulated by the THCTF1 transcription factor are involved in antifungal activity and beneficial plant responses. J. Fungi 2023, 9, 654. [Google Scholar] [CrossRef]
- Karuppiah, V.; Zhang, C.; Liu, T.; Li, Y.; Chen, J. Transcriptome Analysis of T. asperellum GDFS 1009 Revealed the Role of MUP1 Gene on the Methionine-Based Induction of Morphogenesis and Biological Control Activity. J. Fungi 2023, 9, 215. [Google Scholar] [CrossRef]
- Saharan, R.; Patil, J.A.; Yadav, S.; Kumar, A.; Goyal, V. The nematicidal potential of novel fungus, Trichoderma asperellum FbMi6 against Meloidogyne incognita. Sci. Rep. 2023, 13, 6603. [Google Scholar] [CrossRef]
- Zapata-Sarmiento, D.H.; Palacios-Pala, E.F.; Rodríguez-Hernández, A.A.; Medina Melchor, D.L.; Rodríguez-Monroy, M.; Sepúlveda-Jiménez, G. Trichoderma asperellum, a potential biological control agent of Stemphylium vesicarium, on onion (Allium cepa L.). Biol. Control 2020, 140, 104105. [Google Scholar] [CrossRef]
- Wu, Q.; Sun, R.; Ni, M.; Yu, J.; Li, Y.; Yu, C.; Dou, K.; Ren, J.; Chen, J. Identification of a novel fungus, Trichoderma asperellum GDFS1009, and comprehensive evaluation of its biocontrol efficacy. PLoS ONE 2017, 12, e0179957. [Google Scholar] [CrossRef]
- Ou, T.; Zhang, M.; Gao, H.; Wang, F.; Xu, W.; Liu, X.; Wang, L.; Wang, R.; Xie, J. Study on the potential for stimulating mulberry growth and drought tolerance of plant growth-promoting fungi. Int. J. Mol. Sci. 2023, 24, 4090. [Google Scholar] [CrossRef]
- Irshad, K.; Shaheed Siddiqui, Z.; Chen, J.; Rao, Y.; Hamna Ansari, H.; Wajid, D.; Nida, K.; Wei, X. Bio-priming with salt tolerant endophytes improved crop tolerance to salt stress via modulating photosystem II and antioxidant activities in a sub-optimal environment. Front. Plant Sci. 2023, 14, 1082480. [Google Scholar] [CrossRef]
Figure 1.
Maximum likelihood tree showing the evolutionary relationships among different Trichoderma species based on translation elongation factor 1 alpha (TEF1-α) gene and the RNA polymerase II second largest subunit (RPB2) sequences. Maximum likelihood bootstrap values above 50% are indicated at nodes. The tree is rooted with Trichoderma thelephoricola 342. The species in this study are indicated in boldface.
Figure 1.
Maximum likelihood tree showing the evolutionary relationships among different Trichoderma species based on translation elongation factor 1 alpha (TEF1-α) gene and the RNA polymerase II second largest subunit (RPB2) sequences. Maximum likelihood bootstrap values above 50% are indicated at nodes. The tree is rooted with Trichoderma thelephoricola 342. The species in this study are indicated in boldface.

Figure 2.
Dual culture assays of Trichoderma strains with strong antagonistic and Exserohilum turcicum 101. The left side shows Trichoderma strains and the right side shows E. turcicum 101. 1–3: 576; 4–6: 393; 7–9: 64; 10–12: 3A; 13–15: 342; 16–18: 285; 19–21: TU2; 22–24: XZ1-3; 25–27: 402; 28–30: MSL-3; 31–33: 204; 34–36: 417. The images for the fourth, seventh, and fifteenth days of co-culture are arranged from left to right.
Figure 2.
Dual culture assays of Trichoderma strains with strong antagonistic and Exserohilum turcicum 101. The left side shows Trichoderma strains and the right side shows E. turcicum 101. 1–3: 576; 4–6: 393; 7–9: 64; 10–12: 3A; 13–15: 342; 16–18: 285; 19–21: TU2; 22–24: XZ1-3; 25–27: 402; 28–30: MSL-3; 31–33: 204; 34–36: 417. The images for the fourth, seventh, and fifteenth days of co-culture are arranged from left to right.

Figure 3.
Observation of the interaction zone between Trichoderma asperellum 576 and Exserohilum turcicum 101. Scale bars = 20 μm.
Figure 3.
Observation of the interaction zone between Trichoderma asperellum 576 and Exserohilum turcicum 101. Scale bars = 20 μm.

Figure 4.
Inhibition effect of non-volatile substances of Trichoderma strains against Exserohilum turcicum 101. (A) 576; (B) 393; (C) 421; (D) 110; (E) 3A; (F) XZ9-1; (G) 285; (H) 539; (I) 417; (J) XZ1-3; (K) 342; (L) TU2; (M) 64; (N) 402; (O) CK. Bars = 10 mm.
Figure 4.
Inhibition effect of non-volatile substances of Trichoderma strains against Exserohilum turcicum 101. (A) 576; (B) 393; (C) 421; (D) 110; (E) 3A; (F) XZ9-1; (G) 285; (H) 539; (I) 417; (J) XZ1-3; (K) 342; (L) TU2; (M) 64; (N) 402; (O) CK. Bars = 10 mm.

Figure 5.
Inhibition effect of volatile substances of Trichoderma strains against Exserohilum turcicum 101. (A) 576; (B) 393; (C) 421; (D) 110; (E) 3A; (F) CK. Bars = 20 mm.
Figure 5.
Inhibition effect of volatile substances of Trichoderma strains against Exserohilum turcicum 101. (A) 576; (B) 393; (C) 421; (D) 110; (E) 3A; (F) CK. Bars = 20 mm.

Figure 6.
Cell wall degrading enzyme activities of the co-culture suspensions of T. asperellum 576 + E. turcicum 101 or T. atroviride 393 + E. turcicum 101. (A) chitinase; (B) β-1,3-glucanase; (C) cellulase. The length of the box represents the significance of the difference (p < 0.05).
Figure 6.
Cell wall degrading enzyme activities of the co-culture suspensions of T. asperellum 576 + E. turcicum 101 or T. atroviride 393 + E. turcicum 101. (A) chitinase; (B) β-1,3-glucanase; (C) cellulase. The length of the box represents the significance of the difference (p < 0.05).
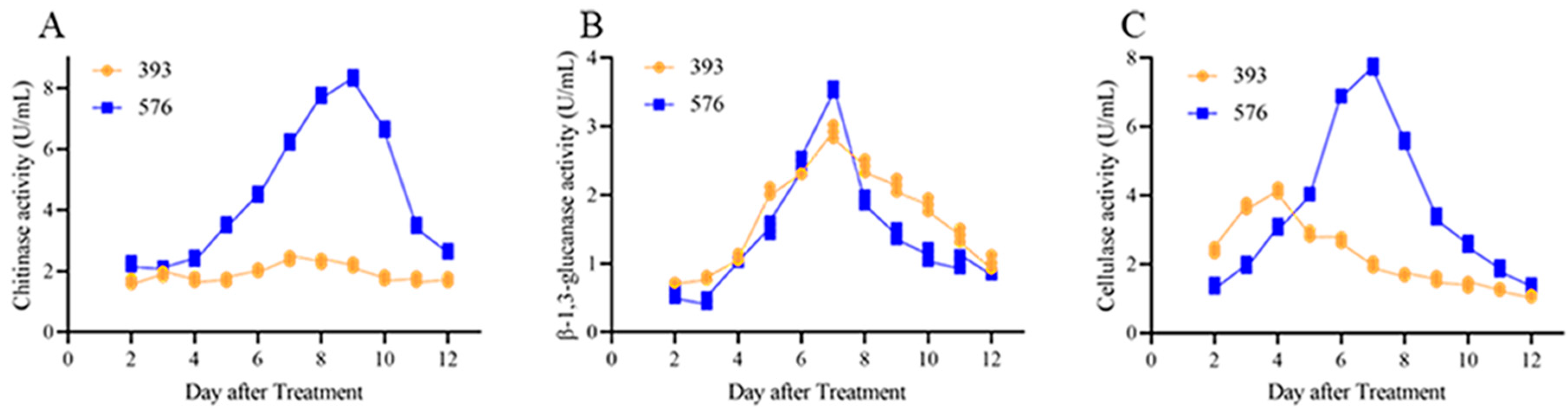
Figure 7.
Effects of Trichoderma asperellum 576 treatment (left) on the germination of maize seeds in comparison with CK (right).
Figure 7.
Effects of Trichoderma asperellum 576 treatment (left) on the germination of maize seeds in comparison with CK (right).

Figure 8.
Effects of Trichoderma asperellum 576 on maize seedlings growth. T1: control plants (plants without T. asperellum 576 or Exserohilum turcicum 101; T2: T. asperellum 576-treated plants; T3: E. turcicum 101-inoculated plants; and T4: E. turcicum 101 and T. asperellum 576-inoculated/treated plants. Bar = 10 cm.
Figure 8.
Effects of Trichoderma asperellum 576 on maize seedlings growth. T1: control plants (plants without T. asperellum 576 or Exserohilum turcicum 101; T2: T. asperellum 576-treated plants; T3: E. turcicum 101-inoculated plants; and T4: E. turcicum 101 and T. asperellum 576-inoculated/treated plants. Bar = 10 cm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Taxonomic information, strain/specimen number, and GenBank accession numbers for TEF1-α and RPB2 genes.
Table 1.
Taxonomic information, strain/specimen number, and GenBank accession numbers for TEF1-α and RPB2 genes.
| Taxon | Strain/Specimen | GenBank Accession Number | |
|---|---|---|---|
| TEF1-α | RPB2 | ||
| Trichoderma atroviride | 393 | OR548045 1 | OR548096 |
| T. afroharzianum | XZ9-1 | OR548046 | OR548111 |
| T. breve | 578 | OR548049 | - |
| T. brevicompactum | 592 | OR548083 | OR548105 |
| T. cerinum | XZ1-3 | OR548077 | OR548110 |
| T. chromospermum | 91 | OR548054 | - |
| T. chromospermum | 338 | OR548066 | - |
| T. crassum | 110 | KT149299 | - |
| T. guizhouense | 526 | OR548073 | - |
| T. guizhouense | TU2 | OR548076 | OR548109 |
| T. hamatum | MSL-3 | OR548084 | - |
| T. harzianum | 581 | OR548050 | OR548103 |
| T. koningiopsis | 421 | OR548081 | OR548099 |
| T. koningiopsis | 439 | OR548071 | OR548101 |
| T. linzhiense | 449 | OR548072 | - |
| T. longibrachiatum | 539 | OR548074 | OR548102 |
| T. longibrachiatum | 3a | OR548053 | - |
| T. longifialidicum | 224 | OR548063 | OR548091 |
| T. longipile | L-3 | OR548075 | - |
| T. dorothopsis | 438 | OR548087 | OR548100 |
| T. pararogersonii | 219 | OR548062 | - |
| T. paratroviride | 402 | OR548069 | OR548097 |
| T. paraviridescens | 295 | OR548079 | OR548092 |
| T. petersenii | 509 | OR548047 | - |
| T. pleuroticola | 588 | OR548082 | OR548104 |
| T. polysporum | 1408 | OR548052 | - |
| T. pyramidale | 285 | OR548064 | - |
| T. rodmanii | 299 | OR548065 | - |
| T. rodmanii | 376 | OR548080 | OR548095 |
| T. endophyticum | 99 | KX689257 | OR548108 |
| T. auriculariae | 417 | OR548070 | OR548098 |
| T. sinense | 204 | OR548060 | - |
| T. solum | 375 | OR548068 | OR548094 |
| T. stipitatum | 218 | OR548061 | - |
| T. strictipile | 370 | OR548078 | - |
| T. strictipile | 115 | OR548058 | OR548089 |
| T. thailandicum | 1283 | OR548086 | OR548090 |
| T. thelephoricola | 342 | OR548067 | OR548093 |
| T. tomentosum | 153 | OR548059 | - |
| T. vinosum | XZ5-2 | OR548085 | - |
| T. longibrachiatum | 593 | OR548051 | OR548106 |
| T. zonatum | 220 | MF374809 | MF374806 |
| T. italicum | 64 | OR548054 | OR548107 |
| T. asperellum | 576 | OR548047 | OR548088 |
| T. asperellum | HZA10 | MK850832 | MH647800 |
| T. atroviride | CBS 119499 | FJ860611 | FJ860518 |
| T. breve | HMAS248845 | KY688046 | KY687984 |
| T. compactum | CBS 121218 | KF134798 | KP115276 |
| T. evansil | Dis 282d | EU856319 | FJ150784 |
| T. helicolixii | CBS 133499 | KJ665517 | KJ665278 |
| T. italicum | CBS 132567 | KJ665525 | KJ665282 |
| T. harzianum | CBS 226.95 | AF534621 | AF545549 |
| T. guizhouense | S628 | KJ665511 | KJ665273 |
| T. hamatum | Th23 | OL439486 | OL412667 |
| T. longibrachiatum | CBS 816.68 | AY865640 | DQ087242 |
| T. polysporum | CPK 3131 | FJ860661 | JQ685878 |
| T. rodmanii | CBS 121553 | FJ860687 | FJ860580 |
| T. koningiopsis | GJS 93-20 | DQ284966 | EU241506 |
| T. paratroviride | CBS136489 | KJ665627 | KJ665321 |
| T. brevicompactum | CBS 109720 | OP203936 | OP203935 |
| T. pseudolacteum | TUFC 61490 | JX238493 | JX238478 |
1 Newly generated sequences in this study are indicated in bold, and sequences that are unavailable in GenBank are indicated by “-”.
Table 2.
Inhibition effect of non-volatile substances of Trichoderma strains against Exserohilum turcicum 101.
Table 2.
Inhibition effect of non-volatile substances of Trichoderma strains against Exserohilum turcicum 101.
| Strains | Colony Diameter (mm) | Inhibition Rate (%) |
|---|---|---|
| 576 | 11.18 | 80.81 ± 2.01 a 1 |
| 393 | 13.29 | 77.68 ± 3.87 ab |
| 421 | 15.84 | 73.41 ± 2.70 ab |
| 110 | 18.35 | 69.20 ± 1.23 ab |
| 3A | 20.35 | 65.84 ± 5.60 bc |
| XZ9-1 | 27.64 | 53.60 ± 4.36 cd |
| 285 | 27.95 | 53.07 ± 2.94 d |
| 539 | 27.95 | 53.07 ± 2.94 d |
| 417 | 34.42 | 42.22 ± 2.02 de |
| XZ1-3 | 37.25 | 37.46 ± 2.44 ef |
| 342 | 40.21 | 32.49 ± 3.01 ef |
| TU2 | 40.64 | 31.77 ± 5.10 ef |
| 64 | 43.92 | 26.27 ± 3.30 fg |
| 402 | 44.82 | 24.75 ± 6.14 g |
| CK | 59.56 | - |
1 Means ± SD followed by the same letters do not differ from each other according to the Fisher’s test (p < 0.05).
Table 3.
Inhibition effect of volatile substances of Trichoderma strains against Exserohilum turcicum 101.
Table 3.
Inhibition effect of volatile substances of Trichoderma strains against Exserohilum turcicum 101.
| Strain | Colony Diameter (mm) | Inhibition Rate (%) |
|---|---|---|
| 576 | 14.72 | 65.86 ± 0.27 a 1 |
| 393 | 18.31 | 57.54 ± 0.71 ab |
| 421 | 19.9 | 53.85 ± 0.34 b |
| 110 | 29.12 | 32.47 ± 0.75 c |
| 3A | 33.16 | 23.10 ± 0.83 d |
| CK | 43.12 | - |
1 Means ± SD followed by the same letters do not differ from each other according to the Fisher’s test (p < 0.05).
Table 4.
Inhibitory effect of the co-culture suspension on Exserohilum turcicum 101.
| Day | T. asperellum 576 + E. turcicum 101 | T. atroviride 393 + E. turcicum 101 | ||
|---|---|---|---|---|
| Colony Diameter (mm) | Inhibition Rate (%) | Colony Diameter (mm) | Inhibition Rate (%) | |
| 2 | 48.79 | 12.48 ± 2.69 gh 1 | 41.53 | 25.50 ± 3.53 h |
| 3 | 40.18 | 27.93 ± 4.32 e | 36.86 | 34.12 ± 2.13 fg |
| 4 | 31.07 | 44.26 ± 3.42 cd | 31.90 | 43.02 ± 1.95 e |
| 5 | 28.19 | 50.43 ± 3.63 bc | 30.20 | 46.22 ± 1.41 de |
| 6 | 25.85 | 53.63 ± 0.64 b | 20.94 | 62.50 ± 4.95 a |
| 7 | 19.82 | 64.45 ± 2.52 a | 27.69 | 50.50 ± 0.71 cd |
| 8 | 34.04 | 54.94 ± 3.15 b | 24.08 | 57.00 ± 1.39 b |
| 9 | 44.09 | 50.91 ± 4.23 bc | 25.75 | 54.03 ± 1.88 bc |
| 10 | 46.41 | 39.74 ± 0.84 de | 28.40 | 49.25 ± 2.37 cd |
| 11 | 47.20 | 15.32 ± 0.86 fg | 35.29 | 36.50 ± 2.12 f |
| 12 | 51.84 | 10.71 ± 3.78 h | 38.27 | 31.14 ± 2.83 g |
| CK | 55.74 | - | - | - |
1 Means ± SD followed by the same letters do not differ from each other according to the Fisher’s test (p < 0.05).
Table 5.
Effect of Trichoderma asperellum 576 on maize seedlings growth. T1: control seedlings without T. asperellum 576 or Exserohilum turcicum 101; T2: maize seedlings treated with T. asperellum 576; T3: maize seedlings inoculated with E. turcicum 101; and T4: maize seedlings inoculated/treated with E. turcicum 101 and T. asperellum 576.
Table 5.
Effect of Trichoderma asperellum 576 on maize seedlings growth. T1: control seedlings without T. asperellum 576 or Exserohilum turcicum 101; T2: maize seedlings treated with T. asperellum 576; T3: maize seedlings inoculated with E. turcicum 101; and T4: maize seedlings inoculated/treated with E. turcicum 101 and T. asperellum 576.
| Treatment | Shoot Height (cm) | Stem Diameter (cm) | Fresh Shoot (g) | Fresh Root (g) | Dry Shoot (g) | Dry Root (g) |
|---|---|---|---|---|---|---|
| T1 | 70.00 ± 2.00 b 1 | 0.55 ± 0.03 b | 9.42 ± 1.18 b | 3.44 ± 0.40 b | 2.00 ± 0.15 b | 1.65 ± 0.18 b |
| T2 | 82.00 ± 1.73 a | 0.65 ± 0.00 a | 14.37 ± 1.86 a | 4.67 ± 0.55 a | 2.97 ± 0.26 a | 2.43 ± 0.06 a |
| T3 | 60.33 ± 1.53 c | 0.44 ± 0.05 c | 5.17 ± 1.26 c | 1.65 ± 0.67 c | 1.03 ± 0.49 c | 0.73 ± 0.40 c |
| T4 | 65.67 ± 3.21 b | 0.51± 0.20 b | 7.50 ± 0.50 c | 2.79 ± 0.24 b | 1.77 ± 0.17 b | 1.42 ± 0.19 b |
1 Means ± SD followed by the same letters do not differ from each other according to the Fisher’s test (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ma, Y.; Li, Y.; Yang, S.; Li, Y.; Zhu, Z. Biocontrol Potential of Trichoderma asperellum Strain 576 against Exserohilum turcicum in Zea mays. J. Fungi 2023, 9, 936. https://doi.org/10.3390/jof9090936
AMA Style
Ma Y, Li Y, Yang S, Li Y, Zhu Z. Biocontrol Potential of Trichoderma asperellum Strain 576 against Exserohilum turcicum in Zea mays. Journal of Fungi. 2023; 9(9):936. https://doi.org/10.3390/jof9090936
Chicago/Turabian StyleMa, Yukun, Yetong Li, Shijia Yang, Yu Li, and Zhaoxiang Zhu. 2023. "Biocontrol Potential of Trichoderma asperellum Strain 576 against Exserohilum turcicum in Zea mays" Journal of Fungi 9, no. 9: 936. https://doi.org/10.3390/jof9090936
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.