
Fungi’s Swiss Army Knife: Pleiotropic Effect of Melanin in Fungal Pathogenesis during Cattle Mycosis


1
Maestría en Biotecnología Agropecuaria, Universidad Técnica Particular de Loja, San Cayetano Alto, Calle París s/n, Loja 1101608, Ecuador
2
Museo de Zoología, Universidad Técnica Particular de Loja, San Cayetano Alto, Calle París s/n, Loja 1101608, Ecuador
3
Departamento de Ciencias Biológicas y Agropecuarias, Facultad de Ciencias Exactas y Naturales, Universidad Técnica Particular de Loja, San Cayetano Alto, Calle París s/n, Loja 1101608, Ecuador
*
Authors to whom correspondence should be addressed.
J. Fungi 2023, 9(9), 929; https://doi.org/10.3390/jof9090929
Submission received: 19 July 2023
/
Revised: 5 September 2023
/
Accepted: 8 September 2023
/
Published: 15 September 2023
(This article belongs to the Special Issue Fungal Diseases in Animals, 2nd Edition)
Abstract
:Fungal threats to public health, food security, and biodiversity have escalated, with a significant rise in mycosis cases globally. Around 300 million people suffer from severe fungal diseases annually, while one-third of food crops are decimated by fungi. Vertebrate, including livestock, are also affected. Our limited understanding of fungal virulence mechanisms hampers our ability to prevent and treat cattle mycoses. Here we aim to bridge knowledge gaps in fungal virulence factors and the role of melanin in evading bovine immune responses. We investigate mycosis in bovines employing a PRISMA-based methodology, bioinformatics, and data mining techniques. Our analysis identified 107 fungal species causing mycoses, primarily within the Ascomycota division. Candida, Aspergillus, Malassezia, and Trichophyton were the most prevalent genera. Of these pathogens, 25% produce melanin. Further research is required to explore the involvement of melanin and develop intervention strategies. While the literature on melanin-mediated fungal evasion mechanisms in cattle is lacking, we successfully evaluated the transferability of immunological mechanisms from other model mammals through homology. Bioinformatics enables knowledge transfer and enhances our understanding of mycosis in cattle. This synthesis fills critical information gaps and paves the way for proposing biotechnological strategies to mitigate the impact of mycoses in cattle.
1. Introduction
Fungi have been shaping and influencing life on Earth for over a billion years [1]. Compared to drug-resistant bacterial infections or viral outbreaks, mycoses received relatively little attention, despite posing a significant threat to public health, food biosecurity, and biodiversity [2,3]. Worldwide, over 300 million people suffer from serious fungal diseases, and more than 1.6 million people die from severe fungal infections yearly [4]. Fungi and fungi-like microorganisms (e.g., Oomycetes) destroy a third of all food crops yearly, which would be sufficient to feed 600 million people.
Historically, we perceive the mycoses that occur in animals as low-prevalence diseases. However, the incidence of recalcitrant, recurrent, and massive mycoses has increased sharply during past decades [5]. For example, the discovery of independent strains of the human pathogen Candida auris simultaneously on three continents [6] could be the first documented example of an emerging fungal disease as a consequence of global warming [7].
Mycoses outbreaks are causing mass mortality in several non-human animal species. For example, chytridiomycosis, an infectious disease of amphibians caused by the chytrid fungi Batrachochytrium dendrobatidis and B. salamandrivorans, is considered the most dramatic disease-driven biodiversity decline ever recorded [8]. The nosemosis caused by Nosema apis and N. ceranae determine mass mortality in Apis mellifera bees [9,10]. Infectious fungal diseases contribute significantly to the extinction of chelonians, ranking among the top five causes of decline [11]. For example, Fusarium solani is responsible for mass mortalities by fusariosis in sea turtles’ eggs [12]. The fungal-like pathogen Sphareothecum destruens causes high mortality in some European fish populations [13]. Fungi can be infectious agents that attack mammals too, causing a plethora of diseases. Fungi cause the critical decrease of several bat populations through the white-nose syndrome caused by Pseudogymnoascus destructan [14].
This article focuses on mycoses that occur in mammals, particularly those reported in cattle. Mammal mycoses can range from superficial skin and mucosal infections to invasive or systemic mycoses that affect major internal organs. Systemic mycoses are less frequent than superficial mycoses and can be highly fatal, killing countless farmed and wild mammals [4]. Our limited understanding of the mechanisms involved in fungal pathogenesis and virulence hinder our ability to prevent, treat, and eradicate fungal diseases in cattle.
A growing body of evidence suggests that melanin plays a relevant role in fungal pathogenesis. It seems that melanin has a pleiotropic effect, participating in different evasion mechanisms that pathogenic fungi employ during infection to evade the immune response of their hosts [5]. The main objective of this research is to summarize the virulence factors associated with fungal pathogens and the relevance of melanin in the evasion of the cattle immune response by fungi. Based on an exhaustive bibliographic search, we provide an extensive reference list of the major fungal pathogens that infect cattle (Bos taurus), the main fungal diseases they cause, and their major virulence factors, including those in which melanin is involved. This review synthesizes evidence that can help fill the information gaps associated with this problem and lead to the proposal of biotechnological strategies that reduce the impacts of fungal mycoses in cattle.
2. Materials and Methods
The methodology used in this article was adapted from the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) [15], but includes various bioinformatics and data mining procedures. We considered pathogenic fungi as those fungal species referred in the literature as mycosis-causing agents. We follow the taxonomic scheme of Mycobank [16,17]. The phylogenetic relationships between many Microsporidia and their inclusion within the kingdom of Fungi remain controversial [18]. However, we include Microsporidia in our study because infections caused by this group have historically been referred to as mycoses (i.e., Microsporidiosis) [19].
The analyzes were carried out using the R Studio user interface [20] and the following two R software packages [21]: 1.—‘litsearchr’ allows us to identify keywords without relying on a potentially biased set of pre-selected articles. This process is automated, fast, objective, reproducible, and easy to implement. ‘litsearchr’ is based on text mining and keyword co-occurrence networks [22]. ‘litsearchr’ also contains a set of functions to improve the efficiency of systematic reviews by automatically deduplicating and assembling results from separate databases. 2.—‘quanteda’ is a compilation of software packages for managing and analyzing textual data that allows natural language processing and transformation into structured text. Its capabilities match or exceed those provided by paid, proprietary end-user software applications [23].
2.1. Research Questions
The main research questions addressed in this review are: Q1.—Does research on cattle fungal diseases suffer the same general bias, receiving less attention than viral or bacterial diseases? Q2.—How many and which species of fungi are pathogens of cattle? Q3.—What are the mycoses caused by these pathogens? Q4.—What are the main factors that influence their virulence? Q5.—Which species of those that infect cattle are melanic? and finally Q6.—Which of the virulence factors are associated with the production of melanin? The answers to these questions will allow us to evaluate the pleiotropic effect of melanin on fungal pathogenesis in bovines.
2.2. Naive Search and Early Keywords
Since we do not intend to compare cases, interventions, or results of different quantitative studies, it was not feasible or necessary to use the PICO method (Population, Intervention, Control, Outcome) to establish the conceptual categories of keywords [24]. However, we deconstructed our central research question (i.e., assessing the “Pleiotropic effect of melanin on fungal pathogenesis in bovines”) into four main groups or concepts of keywords. Except for the broad search to compare the relative scientific productivity associated with fungal, bacterial, or viral diseases (Q1), we carried out systematic searches based on combinations of the search terms related to the concepts of fungus, host, melanin, and mycoses. We performed a non-systematic exploration of the bibliography to construct an initial list of search terms that fit into the proposed groups of concepts.
2.3. Obtaining the Final Keyword List
We use the ‘litsearch’ R package [22] to identify additional potential search words derived from the early keywords proposed in the previous section. The list obtained was cleaned manually and any incongruous or imprecise terms were removed. The definitive list of search terms is presented in Table 1.
2.4. Bibliographic Main Searches in Specialized Databases
We pooled the terms associated with our groups of concepts according to each question in search strings based on Boolean operators. To improve accuracy, we restricted our searches so that search terms appeared in titles and abstracts, and only searched three academic databases: Scopus, Web of Science, and PubMed (Table 2).
The required syntax varies slightly depending on the employed database, so search strings need to be adjusted accordingly. We chose the PubMed, Scopus, and Web of Science platforms because they can be used as primary search systems and are particularly well-suited for evidence-based synthesis in the form of reviews [25]. We conducted these searches in January 2022 and restricted them to articles in English. We report the queries applied to the three data bases and the number of items returned in Table 2.
2.5. Creation and Processing of the Corpus
We consolidated the files (*.*bib) that resulted from the searches from each database to confirm and process the corpus with the ‘quanteda’ R package [23]. We automatically removed duplicate records and compiled the year, title, summary, and reference lists for each article in an Excel file. Afterwards, we filtered the articles based on the manual or automatic application of inclusion criteria (Table 3). The selected articles were read and presented as narrative synthesis or tabulated in tables and figures depending on the question.
Since we did not find any references that described melanin mediated mechanisms of pathogenic evasion by fungi in cattle (Bos taurus), we evaluated if some immunologic mechanisms referred to for other mammals are transferable via homology to cattle. For this, we extracted the name of the host (model, i.e., humane, murine etc.) gene whose expression affects the immune response to fungal infection from selected publications. Then we searched for the amino acid sequence and verified the metabolic pathway of origin in the Kyoto Encyclopedia of Genes and Genomes (KEGG) [26]. Orthologous genes in cattle with a homology (identity) equal to or greater than 75% were afterwards searched for reciprocal BLAST searches performed in pBLast [27].
3. Results
3.1. Patterns of Research
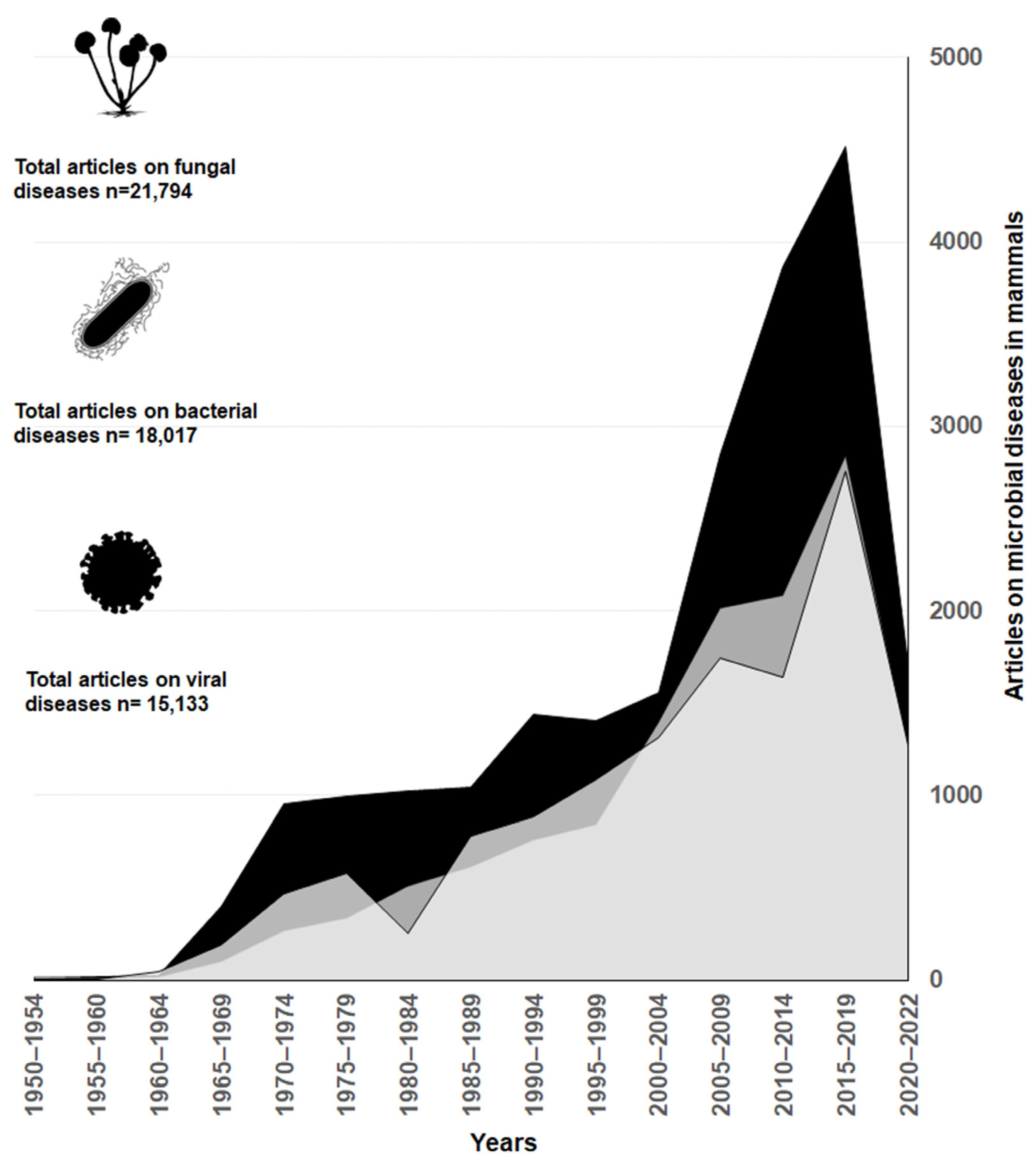
Research of cattle mycoses have advanced faster compared to studies of diseases caused by other microorganisms, such as bacteria and viruses. Overall, mycoses research produced 40% of the articles, while research on bacterial and viral infections represented 32 and 28%, respectively. Starting in the 2000s, there is a marked increase in publications related to fungi diseases compared to bacteria and viruses (Figure 1)
3.2. Diversity of Cattle Fungal Disease
Based on our analysis of nearly 1500 articles on pathogenic fungi and mycosis in cattle, we identified 107 species of fungi causing mycosis in cattle, distributed across five fungal divisions (Ascomycota, Basidiomycota, Microsporidia, Mortierellomycota, and Mucoromycota). Among these divisions, Ascomycota was the best represented, comprising 84 species. The genera with the highest number of species associated with cattle mycosis were Candida (12 species), Aspergillus (9 species), Malassezia (6 species), and Trichophyton (5 species). The detailed diversity of fungi that cause mycosis in bovines is available in Figure S1 and Table S1. The taxonomy used in both is according to Mycobank [16,17]. The Figure S1 is an interactive figure created in Krona [28], arrangement of concentrically nested taxonomic hierarchies from the phylum in the center to the genus in the outer ring.
3.3. Virulence
Each of the 107 species of fungi referred to as pathogenic can be indicated as the etiolant agent of one or more of the 18 (common) mycoses presented in Table 4. A comprehensive list is available in the Supplemental Material (Table S1). At least 40 species are predominant or frequent etiolant agents of any of these diseases. Of the pathogenic fungal species of cattle, 25% biosynthesize melanin (Table 5). Table 6 presents three examples of bovine molecular targets inferred via homology and that are associated with metabolic pathways affected by melanin from pathogenic fungi in other model animals (i.e., house mouse and human).
4. Discussion
The global trend of fungal diseases being relatively less researched compared to other groups of pathogenic microorganisms has been repeatedly mentioned in the literature [2,3]. However, our review reveals that this is not the case in regards to diseases affecting cattle. On the contrary, the evidence presented in Figure 1 shows that the number of articles on fungal diseases is considerably higher than the ones for bacteria and viruses. Thanks to the scientific progress, we now have a better understanding of how specific mycoses affect livestock welfare and production. Some mycoses, such as those causing mycotic abortions and mastitis, have been extensively studied [57,58,59,60,61].
4.1. Diversity of Pathogenic Fungi and Mycoses in Cattle
Rapid evolutionary changes are often associated with antagonistic interactions, such as coevolution between host and pathogen [62]. By definition, pathogens affect the host’s fitness, inducing selection for improved defense mechanisms. Conversely, the improvement of the host’s defense mechanisms will lead to the establishment of more efficient evasion mechanisms by the pathogen [63]. If this antagonism persists for a sufficiently long period, iterative cycles of adaptation and counter-adaptation become an efficient evolutionary force [64]. Numerous examples of rapid evolutionary responses during host-pathogen coevolution exist [65], and resistance to fungal infection in animal hosts could be one of them, although this does not necessarily happen symmetrically.
Some authors hypothesized that the optimization of immune defenses against fungi, including homeothermy, has shaped mammalian evolution [66]. However, it is possible that some virulence factors of pathogenic fungi were acquired as an adaptive response to evade predation or protect against detrimental conditions (i.e., the accidental pathogen hypothesis) [5]. For instance, it is believed that under certain environmental conditions, the extracellular polysaccharide capsule protects Cryptococcus cells from phagocytosis by amoeboid protozoa [67]. Similarly, the deposition of melanin in the cell wall of Cryptococcus protects against thermal and oxidative stress from solar radiation [68]. However, the presence of a capsule and the ability to produce melanin are also two potent virulence factors that make this species a formidable mammalian pathogen.
Compared to bacterial or viral diseases, Systemic fungal diseases are relatively infrequent in non-immunocompromised mammals [66,69]. Mammals have evolved an increased resistance to fungal infections through adaptive immunity [70]. This enables mammals to discriminate between self and non-self and respond rapidly to current and future invaders [71]. Mammals, being homeothermic vertebrates, possess highly sophisticated innate and adaptive immune systems [72]. Mammalian pathogenic fungi must overcome challenges such as surviving within the internal temperatures of mammals, tolerating slightly alkaline environments, adhering to host tissues, and evading host immune responses [73]. As a result, only a small percentage of the described fungal species are pathogens of mammals.
The current richness of fungi is estimated to be between 1.5 million and 5.1 million species [74], with over 100,000 species and 18,000 genera described. Approximately 625 species have been reported as vertebrate pathogens [61]. Around 325 species of pathogenic fungi have been documented in humans [75], and an even smaller number in other mammals. Only a handful of species are common infectious agents. In this context, it is evident that the ability of fungi to infect mammals is a rare skill [75].
Fungi commonly cause mammal diseases through three mechanisms: allergies, intoxication by mycotoxins or mycoses [76]. Although this is a relatively small group of species, the classification of clinically significant fungi and the diseases they cause have been challenging [77]. In the veterinary literature, mycoses are often classified based on various criteria, such as the anatomical area of the host affected by the mycosis (cutaneous, subcutaneous, or systemic), the predominant causative agent (e.g., Aspergillosis caused by the genus Aspergillus), the route of acquisition (exogenous or endogenous), epidemiology, or based on virulence (i.e., opportunistic or commensal). Additionally, specific names are used in animal production, such as fungal abortions and fungal mastitis [78].
During the elaboration of this review, we found no consensus for the use of any of these classifications. Therefore, for practical purposes, the mycoses are preliminarily classified here by integrating several available classifications in the literature (Table 2). This classification is not entirely appropriate, as it exhibits a certain degree of overlap among different categories since, given the opportunistic nature of most causative agents of mycoses, it is common for the same pathogen to be involved in different diseases. However, it can be useful for readers to gain an understanding of the wide spectrum of pathogenic fungi and mycoses that can affect cattle.
4.2. Main Virulence Factors of Mammalian Pathogenic Fungi
The concept of virulence represents an emergent property observed in all pathogens. It arises from the intricate interplay of multiple intrinsic morphological, physiological, metabolic, and molecular characteristics of both host and pathogen [66,79]. In the case of fungal infections, successful colonization of host tissues and subversion of the host’s immune response are pivotal. The ability to adhere to host tissues and secrete tissue-degrading enzymes, which elude the host’s immune defenses, are crucial factors in fungal pathogenesis [60]. These evasion and resistance mechanisms against the immune response are known as virulence factors. While our understanding of fungal pathogenicity and virulence-related traits has significantly advanced [5,80,81], these studies have primarily focused on a limited number of pathogenic species, predominantly in humans, murine models (e.g., laboratory mice Mus musculus) [82], and invertebrates (e.g., Galleria mellonella) [83]. Unfortunately, the knowledge about fungal pathogenesis and virulence factors in bovine infections is limited.
Aspergillus, Candida, Cryptococcus, Fonsecaea, Histoplasma, Paracoccidioides, and Sporothrix are among the most extensively studied genera of mammalian pathogenic fungi that commonly infect cattle [61]. These fungi possess intrinsic characteristics or virulence factors that enable successful attacks on mammals. For example, Aspergillus species demonstrate a remarkable dispersal capacity, tolerance to a wide range of environmental conditions, and the production of potent immunosuppressive and cytotoxic mycotoxins, including aflatoxins, gliotoxin, and ochratoxin. Aspergillus fumigatus, like other fungi, possesses dozens of genes that produce secondary metabolites and virulence-associated proteins, facilitating their establishment in stressful or demanding environments such as the mammalian body [60,80,84]. Similarly, Cryptococcus exhibits key virulence traits such as tolerance to oxidative stress conferred by melanin production through lactase, the formation of a protective extracellular polysaccharide capsule, and a wide range of thermal tolerance [5,48,81,85,86,87].
In Cryptococcus neoformans [88,89] and other pathogenic fungi as Candida auris [90], C. glabrata [91] the generational aging play a role in the virulence increasing resistance to antifungals and phagocytic attacks. The older phenotypes exhibit thickened cell walls. The fungal cell wall is essential for yeast viability and pathogenesis and its thickening seems to have increased levels of all major components, increased intracellular trafficking as well as the alteration of vacuole morphology and pH homeostasis [92]. Thicker walls could also imply an increase in melanin deposition, an important virulence factor in fungi.
4.3. Melanin as a Virulence Factor in Fungal Pathogenesis
Melanin is the generic name for a family of structurally complex, insoluble pigments, ranging in color from dark brown to black [93]. Melanin originates from the polymerization of indolic and phenolic compounds. The structural conformation of melanin has not been established. Its macromolecular nature based on indole precursors and its amorphous properties hinders its crystallographic determination. Melanin is widely represented across all kingdoms, from bacteria to chordates [94,95]. In the fungal kingdom, melanization occurs in all phyla, but proportionally in only a few species. Some fungal species are constitutively melanized, known as dematiaceous, and for them melanization is an obligatory condition. On the other hand, facultative melanized fungi synthetize melanin only during certain stages of their development, in response to specific environmental stimuli (e.g., the presence of phenolic melanin precursors or temperature changes) [93].
Various fungal genera produce melanin. In filamentous fungi, such as Alternaria, Armillaria, Aspergillus, Auricularia, Cladosporium, Epicoccum, Eurotium, Magnapothe, Ochroconis, Penicillium, Phomopsis, Sporothrix, Stachybotrys, and Wangiella, have been found to contain melanin. Melanin production has also occurred in yeast, including Candida albicans, Cryptococcus neoformans, Hormoconis resinae, and Kluyveromyces marxianus [5]. While most assays for melanin production are conducted on strains isolated from the environment, humans, murine, or invertebrates’ models such as Galleria, it is noteworthy that approximately 35% of fungal species causing mycosis in livestock are melanic species. Certain melanized fungi are the primary agents responsible for mycosis in cattle (Table 5).
Melanin plays a significant role in the pathogenesis of mammalian fungal agents such as Aspergillus fumigatus [54], Cryptococcus neoformans [48], Histoplasma capsulatum [39], and Paracoccidioides brasiliensis [39]. Melanin can be considered the Swiss army knife of fungi. Its presence enhances fungal survival in adverse environmental conditions, providing resistance against radiation and enzymatic degradation. A growing body of evidence suggests that melanin makes fungi more resistant to antifungal compounds and other innate immune mechanisms such as phagocytosis [94,96,97]. Overall, melanin exhibits pleiotropic effects, participating in various evasion mechanisms employed by fungal pathogens during specific stages of host immune responses [5]. The primary virulence mechanism associated with melanin is its ability to prevent damage from reactive oxygen species. Melanin neutralizes free radicals, attenuating oxidative stress that could be generated by the oxidative defenses of the immune system or by the environment [98].
Normally, the recognition of any pathogenic cell by the host triggers a response called oxidative burst generated by the innate immune system [75] Immune effector cells such as macrophages, monocytes, and neutrophils release reactive oxygen species (ROS), including superoxide and peroxide ions, as well as nitric oxide [97]. These chemicals are highly cytotoxic and are usually effective in destroying foreign cells. However, melanin is capable of neutralizing oxidative bombardment, protecting the melanotic pathogenic fungi [75].
Melanin can also sequester antifungal drugs as well as the antifungal secondary metabolites [97] as amphotericin B and caspofungin in humans [39] or Fluconazole in cattle [57]. By being located in the cell wall, melanin keeps antifungal compounds in the extracellular space, preventing their cytotoxic effects. Melanin also strongly bind to peptides, immobilizing them, which is a convenient skill for preventing or masking complement deposition on the cell surface [99]. Fungal melanin by itself is an immunogen capable of triggering antibody release in murine models [100]. Resistance to antimicrobial agents has significant implications for morbidity, mortality, and healthcare expenses, as resistant strains are responsible for the majority of infections in both animals and humans [101,102]. However, research on resistance to antifungal agents has historically received less attention than studies focused on understanding antibacterial resistance [103]. Several factors contribute to this disparity. The principal reason for this is the relatively recent recognition of fungal agents as significant pathogens in animals and humans [2,3]. The available information on resistance to fungi in Cattle seems scarce. There are a limited number of antifungal agents licensed for use in animals; however, more and more of those available for the treatment of mycoses in humans are used by veterinarians [104]. In this context, it is certainly to be expected that with the increased use of different antifungal drugs, the number of clinical fungal pathogens that become resistant to these drugs will increase [105].
The melanin of the fungus Fonsecaea monophora inhibits the expression of the i-NOS gene in activated mouse macrophages, regulating the production of nitric oxide (NO) and allowing the fungus to avoid oxidative burst [56]. The release of NO by effector cells of the immune system is a consequence of interferon type II signaling associated with macrophage activation. The release of reactive oxygen species by activated macrophages is one of the principal immune mechanisms used to inhibit the propagation of F. monophora [56]. This behavior has also been observed in the related species F. pedrosoi [34] and in other fungi such as Cryptococcus neoformans [106], Histoplasma capsulatum [107], and Sporothrix schenckii [108].
In Cryptococcus neoformans, those phenotypes that produce melanin faster are more-frequent targets of nonlytic exocytosis [109]. Nonlytic exocytosis is a phenomenon where previously internalized microbes are extruded from host phagocytes, allowing the survival of both cell types [110]. The rapid melanin kinetics is mediated by the laccase enzyme, but its influence on nonlytic exocytosis was unknown. The recent experimental evidence suggests that is probably due to higher laccase production, because fungi lacking this enzyme are nonlytically exocytosed less often [109]. This mechanism happens not only in the interactions of Cryptococcus spp. and other fungal cells with phagocytes, spanning diverse organisms, including mammalian, avian, and fish macrophages, but also in ameboid predators [67]. So, the laccase can be recognized as a novel fungal virulence factor [109,111]. The discovery that laccase could affect nonlytic exocytosis adds one more melanin-independent role for this crucial enzyme in fungi virulence.
The melanin of Aspergillus fumigatus inhibits the assembly and activity of the vacuolar ATPase, a proton pump dependent on ATP type V present in the alveolar phagocytes of the host, preventing the acidification of phagolysosomes and contributes to the pathogenicity of A. fumigatus [54]. Additionally, the melanin in the conidia of A. fumigatus inhibits the intrinsic apoptosis pathway after phagocytosis by macrophages [112].
In Histoplasma capsulatum, melanin induces the formation of granulomas inside mammalian macrophages. Hypoxia-inducible factor 1α (HIF-1α) Infection regulates H. capsulatum in mammals, being a key player in macrophage-mediated innate immunity. HIF-1α creates a hostile environment for yeast cells within macrophages by disrupting the pathogen’s ability to induce host cell autophagy [55].
During this review, the absence of publications describing fungal pathogenic evasion mechanisms in cattle (Bos taurus) suggests an asymmetry in knowledge production. Model animals such as mice or humans are often better studied, leading to a bias that constantly affects various knowledge domains. However, this bias can be significantly and rapidly reduced through the creative utilization of bioinformatic methodologies. The V-type proton ATPases, the HIF-1-alpha and the Nitric Oxide Synthase, inducible (iNOS) are examples of bovine molecular targets inferred via homology and that are associated with metabolic pathways affected by melanin from pathogenic fungi Aspergillus fumigatus [54], Histoplasma capsulatum [55] and Fonsecaea monophorain [56] from other model animals (i.e., house mouse and human). This is a technique that requires further exploration and experimentation, but we feel it should be addressed very immediately.
5. Conclusions
Understanding how melanin affects the virulence of pathogenic fungi is a priority issue in veterinary medicine and animal production. Melanin biosynthesis increases the survival of bovine pathogenic melanic fungi. Melanin confers resistance under environmental (non-pathogenic) conditions, but also virulence to pathogenic fungi in the host. Melanin generates fungus virulence in multiple ways, including by neutralizing reactive oxygen species, sequestering antifungal secondary metabolites, and blocking phagocytosis. However, we still do not know much about its potential as a virulence factor. Based on the evidence presented here, it is clear that despite its relevance in veterinary medicine and animal production, it is still necessary to investigate several aspects of fungal pathogenesis in cattle. Such is the case of virulence mechanisms in which melanin is involved or possible pathways of metabolic regulation of fungal pathogenesis based on biotechnologies aimed at limiting or blocking melanogenesis.
A growing body of research is oriented towards identifying molecular targets associated with melanogenesis. These studies primarily utilize murine and human models. As evidenced here, we can extend this information to different taxonomic groups, such as bovines. For this, it is advisable to generate precise information on the metabolic routes that the host activates in response to the fungus so we can establish homologies efficiently. Studies based on bioinformatics tools allow the generation of synthetic knowledge quickly and consistently with the available evidence, providing a better understanding and management of diseases in which melanin is the main virulence factor.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9090929/s1, Figure S1: Diversity of fungi that cause mycosis in bovines Table S1: Diversity of pathogenic fungi of cattle.
Author Contributions
Conceptualization, V.R. and A.S.; Data curation, V.R. and C.K.; Formal analysis, V.R. and A.S.; Investigation, V.R.; Methodology, V.R.; Resources, V.R., C.K., L.R.S. and A.S.; Supervision, A.S.; Writing—original draft, V.R.; Writing—review & editing, V.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this research are available within the main text of the article or in the Supplemental Material.
Acknowledgments
We would like to express our gratefulness for the support and contributions that made this manuscript possible. We extend our appreciation to the Universidad Técnica Particular de Loja (UTPL) for providing the opportunity for VR to pursue and successfully complete the Master’s Degree in Biotechnology. This manuscript serves as a partial requirement for VR’s degree, and we are grateful for the academic foundation and resources provided by UTPL during his postgraduate program consecution. We are also deeply thankful to the Italo-Ecuadorian Fund for Sustainable Development (FIEDS) and UTPL for granting the scholarship that made VR’s studies endeavors financially feasible. Their support has been instrumental in the realization of this work. We extend our sincere thanks to Arturo Casadevall (Johns Hopkins University, USA), Catalina Rey (Francisco de Miranda National Experimental University, Venezuela), Jorge Ordoñez (Central University of Venezuela, Venezuela), and Alexandra Elbakya (Sci-Hub, Russia) for their generous assistance in obtaining bibliographical references. Additionally, we would like to express our gratitude to Diana Szekely for her insightful comments on the R script and earlier versions of this manuscript. Her feedback and suggestions have significantly contributed to the refinement of our research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gonçalves, S.C.; Haelewaters, D.; Furci, G.; Mueller, G.M. Include All Fungi in Biodiversity Goals. Science 2021, 373, 403. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Nosanchuk, J.D. Fungal Diseases as Neglected Pathogens: A Wake-up Call to Public Health Officials. PLoS Neglected Trop. Dis. 2020, 14, e0007964. [Google Scholar] [CrossRef] [PubMed]
- Seyedmousavi, S.; Guillot, J.; Tolooe, A.; Verweij, P.E.; de Hoog, G.S. Neglected Fungal Zoonoses: Hidden Threats to Man and Animals. Clin. Microbiol. Infect. 2015, 21, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden Killers: Human Fungal Infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.; Murray, D.; Scott, A.; Fu, C.; Averette, A.F.; Sun, S.; Heitman, J.; Magwene, P.M. Pleiotropy and Epistasis within and between Signaling Pathways Defines the Genetic Architecture of Fungal Virulence. PLoS Genet. 2021, 17, e1009313. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida Auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef]
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. On the Emergence of Candida Auris: Climate Change, Azoles, Swamps, and Birds. mBio 2019, 10, 10–1128. [Google Scholar] [CrossRef]
- Fisher, M.C.; Garner, T.W.J. Chytrid Fungi and Global Amphibian Declines. Nat. Rev. Microbiol. 2020, 18, 332–343. [Google Scholar] [CrossRef]
- Iorizzo, M.; Letizia, F.; Ganassi, S.; Testa, B.; Petrarca, S.; Albanese, G.; Di Criscio, D.; De Cristofaro, A. Recent Advances in the Biocontrol of Nosemosis in Honey Bees (Apis mellifera L.). J. Fungi 2022, 8, 424. [Google Scholar] [CrossRef]
- Marín-García, P.J.; Peyre, Y.; Ahuir-Baraja, A.E.; Garijo, M.M.; Llobat, L. The Role of Nosema ceranae (Microsporidia: Nosematidae) in Honey Bee Colony Losses and Current Insights on Treatment. Vet. Sci. 2022, 9, 130. [Google Scholar] [CrossRef]
- Nardoni, S.; Mancianti, F. Mycotic Diseases in Chelonians. J. Fungi 2023, 9, 518. [Google Scholar] [CrossRef]
- Gleason, F.H.; Allerstorfer, M.; Lilje, O. Newly Emerging Diseases of Marine Turtles, Especially Sea Turtle Egg Fusariosis (SEFT), Caused by Species in the Fusarium solani Complex (FSSC). Mycology 2020, 11, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Gozlan, R.; Marshall, W.; Lilje, O.; Jessop, C.; Gleason, F.; Andreou, D. Current Ecological Understanding of Fungal-like Pathogens of Fish: What Lies Beneath? Front. Microbiol. 2014, 5, 62. [Google Scholar] [CrossRef]
- Hoyt, J.R.; Kilpatrick, A.M.; Langwig, K.E. Ecology and Impacts of White-Nose Syndrome on Bats. Nat. Rev. Microbiol. 2021, 19, 196–210. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An Online Initiative to Launch Mycology into the 21st Century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Robert, V.; Vu, D.; Amor, A.B.H.; van de Wiele, N.; Brouwer, C.; Jabas, B.; Szoke, S.; Dridi, A.; Triki, M.; ben Daoud, S.; et al. MycoBank Gearing up for New Horizons. IMA Fungus 2013, 4, 371–379. [Google Scholar] [CrossRef]
- Pang, K.-L.; Hassett, B.T.; Shaumi, A.; Guo, S.-Y.; Sakayaroj, J.; Chiang, M.W.-L.; Yang, C.-H.; Jones, E.B.G. Pathogenic Fungi of Marine Animals: A Taxonomic Perspective. Fungal Biol. Rev. 2021, 38, 92–106. [Google Scholar] [CrossRef]
- Wasson, K.; Peper, R.L. Mammalian Microsporidiosis. Vet. Pathol. 2000, 37, 113–128. [Google Scholar] [CrossRef]
- R Studio Team. RStudio: Integrated Development for R; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Grames, E.M.; Stillman, A.N.; Tingley, M.W.; Elphick, C.S. An Automated Approach to Identifying Search Terms for Systematic Reviews Using Keyword Co-Occurrence Networks. Methods Ecol. Evol. 2019, 10, 1645–1654. [Google Scholar] [CrossRef]
- Benoit, K.; Watanabe, K.; Wang, H.; Nulty, P.; Obeng, A.; Müller, S.; Matsuo, A. Quanteda: An R Package for the Quantitative Analysis of Textual Data. J. Open Source Softw. 2018, 3, 774. [Google Scholar] [CrossRef]
- Speckman, R.A.; Friedly, J.L. Asking Structured, Answerable Clinical Questions Using the Population, Intervention/Comparator, Outcome (PICO) Framework. PMR 2019, 11, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Gusenbauer, M.; Haddaway, N.R. Which Academic Search Systems Are Suitable for Systematic Reviews or Meta-Analyses? Evaluating Retrieval Qualities of Google Scholar, PubMed, and 26 Other Resources. Res. Synth. Methods 2020, 11, 181–217. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating Viruses and Cellular Organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive Metagenomic Visualization in a Web Browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef]
- Du, J.; Wang, X.; Luo, H.; Wang, Y.; Liu, X.; Zhou, X. Epidemiological Investigation of Non-Albicans Candida Species Recovered from Mycotic Mastitis of Cows in Yinchuan, Ningxia of China. BMC Vet. Res. 2018, 14, 251. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Mota, M.; Barros, L.; Dias, M.I.; Ferreira, I.C.F.R.; Piedade, A.P.; Casadevall, A.; Gonçalves, T. Pyomelanin Synthesis in Alternaria Alternata Inhibits DHN-Melanin Synthesis and Decreases Cell Wall Chitin Content and Thickness. Front. Microbiol. 2021, 12, 691433. [Google Scholar] [CrossRef]
- Chongkae, S.; Nosanchuk, J.D.; Pruksaphon, K.; Laliam, A.; Pornsuwan, S.; Youngchim, S. Production of Melanin Pigments in Saprophytic Fungi in Vitro and during Infection. J. Basic Microbiol. 2019, 59, 1092–1104. [Google Scholar] [CrossRef]
- Suri, V.; Pandey, S.; Goyal, N.; Rani, H. Cladophialophora bantiana Brain Abscess with Lymphadenitis. BMJ Case Rep. CP 2021, 14, e246108. [Google Scholar] [CrossRef]
- Romsdahl, J.; Schultzhaus, Z.; Cuomo, C.A.; Dong, H.; Abeyratne-Perera, H.; Hervey, W.J.; Wang, Z. Phenotypic Characterization and Comparative Genomics of the Melanin-Producing Yeast Exophiala lecanii-corni Reveals a Distinct Stress Tolerance Profile and Reduced Ribosomal Genetic Content. J. Fungi 2021, 7, 1078. [Google Scholar] [CrossRef]
- Cunha, M.M.; Franzen, A.J.; Seabra, S.H.; Herbst, M.H.; Vugman, N.V.; Borba, L.P.; de Souza, W.; Rozental, S. Melanin in Fonsecaea pedrosoi: A Trap for Oxidative Radicals. BMC Microbiol. 2010, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Keizer, E.M.; Valdes, I.D.; McCann, B.L.; Bignell, E.M.; Wösten, H.A.B.; de Cock, H. The Protective Role of 1,8-Dihydroxynaphthalene–Melanin on Conidia of the Opportunistic Human Pathogen Aspergillus fumigatus Revisited: No Role in Protection against Hydrogen Peroxide and Superoxides. mSphere 2022, 7, e00874-21. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Yu, Y.; Chen, J.; Yu, B.; Chen, T.; Ying, H.; Zhou, S.; Ouyang, P.; Liu, D.; Chen, Y. Light Signaling Regulates Aspergillus niger Biofilm Formation by Affecting Melanin and Extracellular Polysaccharide Biosynthesis. mBio 2021, 12, e03434-20. [Google Scholar] [CrossRef] [PubMed]
- Palonen, E.K.; Raina, S.; Brandt, A.; Meriluoto, J.; Keshavarz, T.; Soini, J.T. Melanisation of Aspergillus terreus—Is Butyrolactone I Involved in the Regulation of Both DOPA and DHN Types of Pigments in Submerged Culture? Microorganisms 2017, 5, 22. [Google Scholar] [CrossRef]
- Babitskaia, V.G.; Shcherba, V.V.; Filimonova, T.V.; Grigorchuk, E.Z. Melanin pigments of the fungi Paecilomyces variotii and Aspergillus carbonarius. Prikl. Biokhimiia Mikrobiol. 2000, 36, 153–159. [Google Scholar]
- Liu, S.; Youngchim, S.; Zamith-Miranda, D.; Nosanchuk, J.D. Fungal Melanin and the Mammalian Immune System. J. Fungi 2021, 7, 264. [Google Scholar] [CrossRef]
- Youngchim, S.; Pornsuwan, S.; Nosanchuk, J.D.; Dankai, W.; Vanittanakom, N. Melanogenesis in Dermatophyte Species in Vitro and during Infection. Microbiology 2011, 157, 2348–2356. [Google Scholar] [CrossRef]
- Ben Tahar, I.; Kus-Liśkiewicz, M.; Lara, Y.; Javaux, E.; Fickers, P. Characterization of a Nontoxic Pyomelanin Pigment Produced by the Yeast Yarrowia lipolytica. Biotechnol. Prog. 2020, 36, e2912. [Google Scholar] [CrossRef]
- Noguchi, H.; Matsumoto, T.; Kimura, U.; Hiruma, M.; Kano, R.; Yaguchi, T.; Fukushima, S.; Ihn, H. Fungal Melanonychia Caused by Candida parapsilosis Successfully Treated with Oral Fosravuconazole. J. Dermatol. 2019, 46, 911–913. [Google Scholar] [CrossRef]
- Jeon, G.; Kim, Y.; Choi, S.Y.; Kim, Y.-H.; Min, J. Melanin Decolorization by Lysosome-Related Extract in Saccharomyces cerevisiae Modified to Overproduce Glutathione Peroxidase. Appl. Microbiol. Biotechnol. 2021, 105, 8715–8725. [Google Scholar] [CrossRef]
- Vijay, K.; Devi, T.S.; Sree, K.K.; Elgorban, A.M.; Kumar, P.; Govarthanan, M.; Kavitha, T. In Vitro Screening and in Silico Prediction of Antifungal Metabolites from Rhizobacterium Achromobacter kerstersii JKP9. Arch. Microbiol. 2020, 202, 2855–2864. [Google Scholar] [CrossRef] [PubMed]
- Youngchim, S.; Nosanchuk, J.D.; Pornsuwan, S.; Kajiwara, S.; Vanittanakom, N. The Role of L-DOPA on Melanization and Mycelial Production in Malassezia Furfur. PLoS ONE 2013, 8, e63764. [Google Scholar] [CrossRef] [PubMed]
- Gaitanis, G.; Magiatis, P.; Hantschke, M.; Bassukas, I.D.; Velegraki, A. The Malassezia Genus in Skin and Systemic Diseases. Clin. Microbiol. Rev. 2012, 25, 106–141. [Google Scholar] [CrossRef]
- Brilhante, R.S.N.; da Rocha, M.G.; de Guedes, G.M.M.; de Oliveira, J.S.; dos Santos Araújo, G.; España, J.D.A.; Sales, J.A.; de Aguiar, L.; de Paiva, M.A.N.; de Cordeiro, R.A.; et al. Malassezia pachydermatis from Animals: Planktonic and Biofilm Antifungal Susceptibility and Its Virulence Arsenal. Vet. Microbiol. 2018, 220, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Danesi, P.; Falcaro, C.; Schmertmann, L.J.; de Miranda, L.H.M.; Krockenberger, M.; Malik, R. Cryptococcus in Wildlife and Free-Living Mammals. J. Fungi 2021, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Asadzadeh, M.; Ahmad, S.; Ziauddin, K.; Verghese, S.; Joseph, L. Molecular Identification, Genotypic Heterogeneity and Comparative Pathogenicity of Environmental Isolates of Papiliotrema Laurentii. J. Med. Microbiol. 2020, 69, 1285–1292. [Google Scholar] [CrossRef]
- Figueiredo-Carvalho, M.H.G.; dos Santos, F.B.; Nosanchuk, J.D.; Zancope-Oliveira, R.M.; Almeida-Paes, R. L-Dihydroxyphenylalanine Induces Melanin Production by Members of the Genus Trichosporon. FEMS Yeast Res. 2014, 14, 988–991. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Fernández, E.Z.; Shi, Y.-M.; Grün, P.; Bode, H.B.; Bölker, M. An Unconventional Melanin Biosynthesis Pathway in Ustilago maydis. Appl. Environ. Microbiol. 2021, 87, e01510-20. [Google Scholar] [CrossRef]
- Knudtson, W.U.; Kirkbride, C.A. Fungi Associated with Bovine Abortion in the Northern Plains States (USA). J. Vet. Diagn. Investig. 1992, 4, 181–185. [Google Scholar] [CrossRef]
- Jacobsen, I.D. Animal Models to Study Mucormycosis. J. Fungi 2019, 5, 27. [Google Scholar] [CrossRef]
- Thywißen, A.; Heinekamp, T.; Dahse, H.-M.; Schmaler-Ripcke, J.; Nietsche, S.; Zipfel, P.; Brakhage, A. Conidial Dihydroxynaphthalene Melanin of the Human Pathogenic Fungus Aspergillus fumigatus Interferes with the Host Endocytosis Pathway. Front. Microbiol. 2011, 2, 96. [Google Scholar] [CrossRef]
- Friedrich, D.; Zapf, D.; Lohse, B.; Fecher, R.A.; Deepe, G.S.; Rupp, J. The HIF-1α/LC3-II Axis Impacts Fungal Immunity in Human Macrophages. Infect. Immun. 2019, 87, 10–1128. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Xi, L.; Huang, H.; Hu, Y.; Li, X.; Huang, X.; Lu, S.; Sun, J. Melanin in a Meristematic Mutant of Fonsecaea monophora Inhibits the Production of Nitric Oxide and Th1 Cytokines of Murine Macrophages. Mycopathologia 2013, 175, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Awandkar, S.P.; Kulkarni, M.B.; Agnihotri, A.A.; Chavan, V.G.; Chincholkar, V.V. Novel Fluconazole-Resistant Zoonotic Yeast Isolated from Mastitis. Anim. Biotechnol. 2021, 34, 746–755. [Google Scholar] [CrossRef]
- Elad, D.; Segal, E. Diagnostic Aspects of Veterinary and Human Aspergillosis. Front. Microbiol. 2018, 9, 1303. [Google Scholar] [CrossRef] [PubMed]
- Pesca, C.; Cruciani, D.; Agostini, L.; Rossi, E.; Pavone, S.; Stazi, M.; Mangili, P.; Crotti, S. Simultaneous Detection of Aspergillus nidulans, Aspergillus luchuensis and lichtheimia Sp. in a Bovine Abortion. J. Mycol. Médicale 2020, 30, 100923. [Google Scholar] [CrossRef] [PubMed]
- Seyedmousavi, S.; Guillot, J.; Arné, P.; de Hoog, G.S.; Mouton, J.W.; Melchers, W.J.G.; Verweij, P.E. Aspergillus and Aspergilloses in Wild and Domestic Animals: A Global Health Concern with Parallels to Human Disease. Med. Mycol. 2015, 53, 765–797. [Google Scholar] [CrossRef]
- Carpouron, J.; de Hoog, S.; Gentekaki, E.; Hyde, K. Emerging and Epizootic Fungal Infections in Animals; Seyedmousavi, S., De Hoog, G.S., Guillot, J., Verweij, P.E., Eds.; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-72091-3. [Google Scholar]
- Papkou, A.; Schalkowski, R.; Barg, M.-C.; Koepper, S.; Schulenburg, H. Population Size Impacts Host–Pathogen Coevolution. Proc. R. Soc. B Biol. Sci. 2021, 288, 20212269. [Google Scholar] [CrossRef]
- Masri, L.; Branca, A.; Sheppard, A.E.; Papkou, A.; Laehnemann, D.; Guenther, P.S.; Prahl, S.; Saebelfeld, M.; Hollensteiner, J.; Liesegang, H.; et al. Host–Pathogen Coevolution: The Selective Advantage of Bacillus thuringiensis Virulence and Its Cry Toxin Genes. PLoS Biol. 2015, 13, e1002169. [Google Scholar] [CrossRef]
- Woolhouse, M.E.J.; Webster, J.P.; Domingo, E.; Charlesworth, B.; Levin, B.R. Biological and Biomedical Implications of the Co-Evolution of Pathogens and Their Hosts. Nat. Genet. 2002, 32, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, L.J.; Ashby, B. Coevolutionary Theory of Hosts and Parasites. J. Evol. Biol. 2022, 35, 205–224. [Google Scholar] [CrossRef]
- Casadevall, A. Fungi and the Rise of Mammals. PLoS Pathog. 2012, 8, e1002808. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Casadevall, A. Phagosome Extrusion and Host-Cell Survival after Cryptococcus neoformans Phagocytosis by Macrophages. Curr. Biol. 2006, 16, 2161–2165. [Google Scholar] [CrossRef] [PubMed]
- Rosas, Á.L.; Casadevall, A. Melanization Affects Susceptibility of Cryptococcus neoformans to Heat and Cold. FEMS Microbiol. Lett. 1997, 153, 265–272. [Google Scholar] [CrossRef]
- Biegańska, M.J. Two Fundamentals of Mammalian Defense in Fungal Infections: Endothermy and Innate Antifungal Immunity. Pol. J. Vet. Sci. 2014, 17, 555–567. [Google Scholar] [CrossRef]
- Husband, A.J. Overview of the Mammalian Immune System. In Advances in Nutritional Research: Immunological Properties of Milk; Woodward, B., Draper, H.H., Eds.; Advances in Nutritional Research; Springer: Boston, MA, USA, 2001; pp. 3–14. ISBN 978-1-4615-0661-4. [Google Scholar]
- Köhler, J.R.; Hube, B.; Puccia, R.; Casadevall, A.; Perfect, J.R. Fungi That Infect Humans. Microbiol. Spectr. 2017, 5, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Robert, V.A.; Casadevall, A. Vertebrate Endothermy Restricts Most Fungi as Potential Pathogens. J. Infect. Dis. 2009, 200, 1623–1626. [Google Scholar] [CrossRef]
- Casadevall, A. Fungal Virulence, Vertebrate Endothermy, and Dinosaur Extinction: Is There a Connection? Fungal Genet. Biol. 2005, 42, 98–106. [Google Scholar] [CrossRef]
- Blackwell, M. The Fungi: 1, 2, 3 … 5.1 Million Species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef]
- Casadevall, A. Immunity to Invasive Fungal Diseases. Annu. Rev. Immunol. 2022, 40, 121–141. [Google Scholar] [CrossRef]
- Kappe, R.; Rimek, D. Fungal Diseases. In Antifungal Agents: Advances and Problems; Müller, J., Polak, A., Kappe, R., Rimek, D., Seibold, M., Tintelnot, K., Jucker, E., Eds.; Special Topic; Birkhäuser: Basel, Switzerland, 2003; pp. 13–38. ISBN 978-3-0348-7974-3. [Google Scholar]
- Müller, J.; Polak, A.; Kappe, R.; Rimek, D.; Seibold, M.; Tintelnot, K. Antifungal Agents: Advances and Problems; Jucker, E., Ed.; Birkhäuser: Basel, Switzerland, 2003; ISBN 978-3-7643-6926-2. [Google Scholar]
- Balkema-Buschmann, A.; Fast, C.; Kaatz, M.; Eiden, M.; Ziegler, U.; McIntyre, L.; Keller, M.; Hills, B.; Groschup, M.H. Pathogenesis of Classical and Atypical BSE in Cattle. Prev. Vet. Med. 2011, 102, 112–117. [Google Scholar] [CrossRef]
- Garcia-Solache, M.A.; Casadevall, A. Phylogenetics and Evolution of Virulence in the Kingdom Fungi. In Evolution of Virulence in Eukaryotic Microbes; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; pp. 70–90. ISBN 978-1-118-30816-5. [Google Scholar]
- Bultman, K.M.; Kowalski, C.H.; Cramer, R.A. Aspergillus fumigatus Virulence through the Lens of Transcription Factors. Med. Mycol. 2017, 55, 24–38. [Google Scholar] [CrossRef]
- Zaragoza, O. Basic Principles of the Virulence of Cryptococcus. Virulence 2019, 10, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Hohl, T.M. Overview of Vertebrate Animal Models of Fungal Infection. J. Immunol. Methods 2014, 410, 100–112. [Google Scholar] [CrossRef]
- Torres, M.; Pinzón, E.N.; Rey, F.M.; Martinez, H.; Parra Giraldo, C.M.; Celis Ramírez, A.M. Galleria mellonella as a Novelty in Vivo Model of Host-Pathogen Interaction for Malassezia furfur CBS 1878 and Malassezia pachydermatis CBS 1879. Front. Cell. Infect. Microbiol. 2020, 10, 199. [Google Scholar] [CrossRef]
- Brown, N.A.; Goldman, G.H. The Contribution of Aspergillus fumigatus Stress Responses to Virulence and Antifungal Resistance. J. Microbiol. 2016, 54, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Steenbergen, J.N.; Nosanchuk, J.D. ‘Ready Made’ Virulence and ‘Dual Use’ Virulence Factors in Pathogenic Environmental Fungi—The Cryptococcus neoformans Paradigm. Curr. Opin. Microbiol. 2003, 6, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; May, R.C. Chapter 5 Virulence in Cryptococcus Species. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2009; Volume 67, pp. 131–190. [Google Scholar]
- de Sousa, H.R.; de Frazão, S.; de Oliveira Júnior, G.P.; Albuquerque, P.; Nicola, A.M. Cryptococcal Virulence in Humans: Learning from Translational Studies with Clinical Isolates. Front. Cell. Infect. Microbiol. 2021, 11, 657502. [Google Scholar] [CrossRef] [PubMed]
- Bouklas, T.; Fries, B.C. Aging: An Emergent Phenotypic Trait That Contributes to the Virulence of Cryptococcus neoformans. Future Microbiol. 2015, 10, 191–197. [Google Scholar] [CrossRef]
- Orner, E.P.; Bhattacharya, S.; Kalenja, K.; Hayden, D.; Del Poeta, M.; Fries, B.C. Cell Wall-Associated Virulence Factors Contribute to Increased Resilience of Old Cryptococcus neoformans Cells. Front. Microbiol. 2019, 10, 2513. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Holowka, T.; Orner, E.P.; Fries, B.C. Gene Duplication Associated with Increased Fluconazole Tolerance in Candida auris Cells of Advanced Generational Age. Sci. Rep. 2019, 9, 5052. [Google Scholar] [CrossRef]
- Bouklas, T.; Alonso-Crisóstomo, L.; Székely, T., Jr.; Diago-Navarro, E.; Orner, E.P.; Smith, K.; Munshi, M.A.; Poeta, M.D.; Balázsi, G.; Fries, B.C. Generational Distribution of a Candida Glabrata Population: Resilient Old Cells Prevail, While Younger Cells Dominate in the Vulnerable Host. PLoS Pathog. 2017, 13, e1006355. [Google Scholar] [CrossRef]
- Silva, V.K.A.; Bhattacharya, S.; Oliveira, N.K.; Savitt, A.G.; Zamith-Miranda, D.; Nosanchuk, J.D.; Fries, B.C. Replicative Aging Remodels the Cell Wall and Is Associated with Increased Intracellular Trafficking in Human Pathogenic Yeasts. mBio 2021, 13, e0019022. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.J.B.; Casadevall, A. Melanin. Curr. Biol. 2020, 30, R142–R143. [Google Scholar] [CrossRef]
- Eisenman, H.C.; Casadevall, A. Synthesis and Assembly of Fungal Melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Watanabe, B.; Matsui, K.; Lumyong, S. Characterization of Melanin and Optimal Conditions for Pigment Production by an Endophytic Fungus, Spissiomyces endophytica SDBR-CMU319. PLoS ONE 2019, 14, e0222187. [Google Scholar] [CrossRef]
- Casadevall, A.; Rosas, A.L.; Nosanchuk, J.D. Melanin and Virulence in Cryptococcus neoformans. Curr. Opin. Microbiol. 2000, 3, 354–358. [Google Scholar] [CrossRef]
- Smith, D.F.Q.; Casadevall, A. The Role of Melanin in Fungal Pathogenesis for Animal Hosts. In Fungal Physiology and Immunopathogenesis; Rodrigues, M.L., Ed.; Current Topics in Microbiology and Immunology; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–30. ISBN 978-3-030-30237-5. [Google Scholar]
- Gessler, N.N.; Egorova, A.S.; Belozerskaya, T.A. Melanin Pigments of Fungi under Extreme Environmental Conditions (Review). Appl. Biochem. Microbiol. 2014, 50, 105–113. [Google Scholar] [CrossRef]
- Camacho, E.; Vij, R.; Chrissian, C.; Prados-Rosales, R.; Gil, D.; O’Meally, R.N.; Cordero, R.J.B.; Cole, R.N.; McCaffery, J.M.; Stark, R.E.; et al. The Structural Unit of Melanin in the Cell Wall of the Fungal Pathogen Cryptococcus Neoformans. J. Biol. Chem. 2019, 294, 10471–10489. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Rosas, A.L.; Casadevall, A. The Antibody Response to Fungal Melanin in Mice. J. Immunol. 1998, 160, 6026–6031. [Google Scholar] [CrossRef] [PubMed]
- Irfan, M.; Almotiri, A.; AlZeyadi, Z.A. Antimicrobial Resistance and Its Drivers—A Review. Antibiotics 2022, 11, 1362. [Google Scholar] [CrossRef]
- Vitiello, A.; Ferrara, F.; Boccellino, M.; Ponzo, A.; Cimmino, C.; Comberiati, E.; Zovi, A.; Clemente, S.; Sabbatucci, M. Antifungal Drug Resistance: An Emergent Health Threat. Biomedicines 2023, 11, 1063. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the Emerging Threat of Antifungal Resistance to Human Health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Bossche, H.V.; Engelen, M.; Rochette, F. Antifungal Agents of Use in Animal Health—Chemical, Biochemical and Pharmacological Aspects. J. Vet. Pharmacol. Ther. 2003, 26, 5–29. [Google Scholar] [CrossRef]
- Bhanderi, B.; Yadav, M.; Roy, A. Antifungal Drug Resistance-Concerns for Veterinarians. Vet. World 2009, 2, 204–207. [Google Scholar] [CrossRef]
- Rossi, G.R.; Cervi, L.A.; García, M.M.; Chiapello, L.S.; Sastre, D.A.; Masih, D.T. Involvement of Nitric Oxide in Protecting Mechanism during Experimental Cryptococcosis. Clin. Immunol. 1999, 90, 256–265. [Google Scholar] [CrossRef]
- Lane, T.E.; Otero, G.C.; Wu-Hsieh, B.A.; Howard, D.H. Expression of Inducible Nitric Oxide Synthase by Stimulated Macrophages Correlates with Their Antihistoplasma Activity. Infect. Immun. 1994, 62, 1478–1479. [Google Scholar] [CrossRef]
- Fernandes, K.S.S.; Coelho, A.L.J.; Bezerra, L.M.L.; Barja-Fidalgo, C. Virulence of Sporothrix schenckii Conidia and Yeast Cells, and Their Susceptibility to Nitric Oxide. Immunology 2000, 101, 563–569. [Google Scholar] [CrossRef]
- de Oliveira Frazão, S.; de Sousa, H.R.; da Silva, L.G.; dos Santos Folha, J.; de Melo Gorgonha, K.C.; de Oliveira, G.P.; Felipe, M.S.S.; Silva-Pereira, I.; Casadevall, A.; Nicola, A.M.; et al. Laccase Affects the Rate of Cryptococcus neoformans Nonlytic Exocytosis from Macrophages. mBio 2020, 11, e02085-20. [Google Scholar] [CrossRef]
- Ma, H.; Croudace, J.E.; Lammas, D.A.; May, R.C. Expulsion of Live Pathogenic Yeast by Macrophages. Curr. Biol. 2006, 16, 2156–2160. [Google Scholar] [CrossRef]
- Cordero, R.J.B.; Camacho, E.; Casadevall, A. Melanization in Cryptococcus neoformans Requires Complex Regulation. mBio 2020, 11, e03313-19. [Google Scholar] [CrossRef] [PubMed]
- Volling, K.; Thywissen, A.; Brakhage, A.A.; Saluz, H.P. Phagocytosis of Melanized Aspergillus Conidia by Macrophages Exerts Cytoprotective Effects by Sustained PI3K/Akt Signalling. Cell. Microbiol. 2011, 13, 1130–1148. [Google Scholar] [CrossRef]
Figure 1.
Number of scientific papers studying the three main pathogen groups that infect cattle. The black polygon corresponds to fungi, the intermediate grey polygon to bacteria and the light grey polygon to viruses. The results are based on an exploratory search in PubMed using the search terms available in the Supplementary Material.
Figure 1.
Number of scientific papers studying the three main pathogen groups that infect cattle. The black polygon corresponds to fungi, the intermediate grey polygon to bacteria and the light grey polygon to viruses. The results are based on an exploratory search in PubMed using the search terms available in the Supplementary Material.


{kind=link}
Table 1.
Key concepts and their associated search terms used in three online data bases.
| Core Concept | Associated Search Terms |
|---|---|
| Fungi | aspergillus OR candida OR cryptococcus OR enterocytozoon OR exophiala OR fonsecaea OR fungal OR fungi OR fungus OR histoplasma OR paracoccidioides OR penicillium OR sporothrix OR talaromyces OR trichophyton OR yeast |
| Host | bovid OR bovine OR bovino OR bullocks OR bulls OR calves OR cattle OR cow OR dairy OR livestock |
| Melanin | melanin OR melanogenesis OR melanotic |
| Mycosis | aspergillosis OR blastomycosis OR candidiasis OR chromoblastomycosis OR coccidioidomycosis OR cryptococcosis OR dermatophytosis OR epizootic OR abortions OR keratomycosis OR lymphangitis OR maduromycosis OR mastitis OR mucormycosis OR mycetoma OR mycotic OR mycosis OR mycoses OR paracoccidioidomycosis OR phaeomycosis OR pithomycotoxicosis OR pythiosis OR ringworm OR sporotrichosis OR zygomycosis |
Table 2.
Research questions and respective searches keywords used in three online data bases.
| Question | Keywords String | Data Base | Processed Articles | |
|---|---|---|---|---|
| Q1 | “fungal AND disease AND cattle” | PubMed | 21,794 18,017 15,133 | 54,944 |
| “bacterial AND disease AND cattle” | ||||
| “viral AND disease AND cattle” | ||||
| Q2–Q4 | “associated with the fungi concept” AND “associated with the host concept” AND “associated with the concept mycosis” | PubMed Scopus WoS | 417 758 238 | 1413 |
| Q5–Q6 | “associated with the fungi concept” AND “ “associated with the concept melanin” | PubMed Scopus WoS | 187 128 120 | 435 |
Table 3.
Articles inclusion criteria to conform the corpus.
| Criteria | Type | |
|---|---|---|
| 1 | At least one of the keywords associated with each search concept present in the title or abstract | Automatic |
| 2 | The article is not duplicated within the corpus | Automatic |
| 3 | The article is in English | Automatic |
| 4 | It was selected by context in R (questions Q5–Q6 only) associated with one of the connecting words, established | Automatic |
| 5 | The article contains some unusual term in capital letters (we did this with the help of the “keep tokens” function and the regular expression “[^[:UPPER:]^]” in R, only questions Q2–Q4) | Automatic |
| 6 | Explicitly mentions a pathogenic fungal species or describes a melanin-associated virulence mechanism | Manual |
Table 4.
Main mycoses encountered in bovines and their causative agents. Taxonomy and references available in Table S1.
Table 4.
Main mycoses encountered in bovines and their causative agents. Taxonomy and references available in Table S1.
| Mycosis Type | Mycosis | Predominant Mycosis-Causing Agent |
|---|---|---|
| Cutaneous, superficial, ringworm or dermatophytosis | Pheomycosis | Alternaria alternata |
| Epizootic Lymphangitis | Trichophyton verrucosum | |
| Mycetoma or maduromycosis | Cochliobolus spiciferus | |
| Curvularia geniculata | ||
| Drechslera rostratum | ||
| Madurella mycetomatis | ||
| Pseudallescheria boydii | ||
| Pythiosis | Pythium insidiosum | |
| Keratomycosis | Mortierella wolfii | |
| Ringworm | Cladophiolophora bantiana | |
| Microsporum canis | ||
| Trichophyton mentagrophytes | ||
| Subcutaneous | Chromoblastomycosis | Fonsecaea monophora |
| Fonsecaea pedrosoi | ||
| Sporotrichosis | Sporothrix schenckii and another ssp. | |
| Systemic, primary, internal, disseminated or deep | Cattle Mycotic abortion | Aspergillus fumigatus |
| Aspergillus nidulans | ||
| Candida albicans | ||
| Emericella nidulan | ||
| Lichtheimia corymbifera | ||
| Mortierella wolfii | ||
| Rhizomucor pusillus | ||
| Rhizopus arrhizus | ||
| Aspergillosis | Aspergillus fumigatus and another ssp. | |
| Blastomycosis | Blastomyces dermatitidis | |
| Candidiasis | Candida albicans and another ssp. | |
| Coccidioidomycosis | Coccidioides immitis | |
| Cryptococcosis | Cryptococcus neoformans | |
| Histoplasmosis | Histoplasma capsulatum | |
| Fungal mastitis | Aspergillus fumigatus | |
| Aureobasidium pullulans | ||
| Candida albicans and another ssp. | ||
| Candida parapsilosis | ||
| Cryptococcus neoformans | ||
| Prototheca zopfii | ||
| Trichosporon mucoides | ||
| Paracoccidioidomycosis | Paracoccidioides brasiliensis | |
| Zygomycosis or Mucormycosis | Syncephalastrum racemosum | |
| Cunninghamella bertholletiae | ||
| Saksenaea vasiformis |
Table 5.
Major mammalian melanistic fungal pathogens infecting cattle, full list of species found in this review available in Table S1.
Table 5.
Major mammalian melanistic fungal pathogens infecting cattle, full list of species found in this review available in Table S1.
| Division | Species |
|---|---|
| Ascomycota | Aureobasidium pullulans [29] |
| Alternaria alternata [30] | |
| Exserohilum rostratum [31] | |
| Cladophialophora bantiana [32] | |
| Exophiala dermatitidis [33] | |
| Fonsecaea pedrosoi [34] | |
| Aspergillus flavus [31] | |
| Aspergillus fumigatus [33] | |
| Aspergillus nidulans [35] | |
| Aspergillus niger [36] | |
| Aspergillus terreus [37] | |
| Paecilomyces variotii [38] | |
| Blastomyces dermatitidis [39] | |
| Histoplasma capsulatum [39] | |
| Paracoccidioides brasiliensis [39] | |
| Trichophyton rubrum [40] | |
| Yarrowia lipolytica [41] | |
| Candida albicans [39] | |
| Candida parapsilosis [42] | |
| Saccharomyces cerevisiae [43] | |
| Fusarium oxysporum [44] | |
| Sporothrix schenckii [39] | |
| Basidiomycota | Malassezia furfur [45] |
| Malassezia obtusa [46] | |
| Malassezia pachydermatis [47] | |
| Malassezia sympodialis [46] | |
| Cryptococcus neoformans [48] | |
| Papiliotrema laurentii [49] | |
| Trichosporon asahii [50] | |
| Ustilago maydis [51] | |
| Mortierellomycota | Actinomortierella wolfii [52] |
| Mucoromycota | Lichtheimia corymbifera [53] |
| Mucor hiemalis [53] | |
| Rhizopus arrhizus [53] | |
| Rhizopus microsporus [53] |
Table 6.
Examples of knowledge transfer by homology of orthologous genes.
| Fungi | Aspergillus fumigatus | Histoplasma capsulatum | Fonsecaea monophora |
| Model | Mus musculus | Homo sapiens | Mus musculus |
| Way | Proton-dependent phagolysosomal acidification type V ATPases | Expression of Hypoxia-inducible Factor 1HFI-1 | Type II interferon signaling associated with macrophage activation |
| Target | V-type proton ATPases | HIF-1-alpha | Nitric Oxide Synthase, inducible (iNOS) |
| Identity B.taurus | 91% | 94% | 88% |
| Source | [54] | [55] | [56] |
| KEGG link | KEGG T01008: 338038 | KEGG T01008: 281814 | KEGG T01008: 282876 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Romero, V.; Kalinhoff, C.; Saa, L.R.; Sánchez, A. Fungi’s Swiss Army Knife: Pleiotropic Effect of Melanin in Fungal Pathogenesis during Cattle Mycosis. J. Fungi 2023, 9, 929. https://doi.org/10.3390/jof9090929
AMA Style
Romero V, Kalinhoff C, Saa LR, Sánchez A. Fungi’s Swiss Army Knife: Pleiotropic Effect of Melanin in Fungal Pathogenesis during Cattle Mycosis. Journal of Fungi. 2023; 9(9):929. https://doi.org/10.3390/jof9090929
Chicago/Turabian StyleRomero, Víctor, Carolina Kalinhoff, Luis Rodrigo Saa, and Aminael Sánchez. 2023. "Fungi’s Swiss Army Knife: Pleiotropic Effect of Melanin in Fungal Pathogenesis during Cattle Mycosis" Journal of Fungi 9, no. 9: 929. https://doi.org/10.3390/jof9090929
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.