
Alkaliphilic/Alkali-Tolerant Fungi: Molecular, Biochemical, and Biotechnological Aspects
by
 , and
, and
Maikel Gilberto Fernández-López
1,* ,
,
Ramón Alberto Batista-García
2 and
Elva Teresa Aréchiga-Carvajal
1,* 1
Unidad de Manipulación Genética, Laboratorio de Micología y Fitopatología, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, San Nicolás de los Garza 66451, Mexico
2
Centro de Investigación en Dinámica Celular, Instituto de Investigación en Ciencias Básicas y Aplicadas, Universidad Autónoma del Estado de Morelos, Cuernavaca 62209, Mexico
*
Authors to whom correspondence should be addressed.
J. Fungi 2023, 9(6), 652; https://doi.org/10.3390/jof9060652
Submission received: 13 February 2023
/
Revised: 8 May 2023
/
Accepted: 8 May 2023
/
Published: 9 June 2023
(This article belongs to the Special Issue Extremophile Fungi: An Arising Novel Field in Perspective)
Abstract
:Biotechnologist interest in extremophile microorganisms has increased in recent years. Alkaliphilic and alkali-tolerant fungi that resist alkaline pH are among these. Alkaline environments, both terrestrial and aquatic, can be created by nature or by human activities. Aspergillus nidulans and Saccharomyces cerevisiae are the two eukaryotic organisms whose pH-dependent gene regulation has received the most study. In both biological models, the PacC transcription factor activates the Pal/Rim pathway through two successive proteolytic mechanisms. PacC is a repressor of acid-expressed genes and an activator of alkaline-expressed genes when it is in an active state. It appears, however, that these are not the only mechanisms associated with pH adaptations in alkali-tolerant fungi. These fungi produce enzymes that are resistant to harsh conditions, i.e., alkaline pH, and can be used in technological processes, such as in the textile, paper, detergent, food, pharmaceutical, and leather tanning industries, as well as in bioremediation of pollutants. Consequently, it is essential to understand how these fungi maintain intracellular homeostasis and the signaling pathways that activate the physiological mechanisms of alkali resistance in fungi.
1. Introduction
Biotechnological processes require the use of products mainly from mesophilic microorganisms, which thrive well in environmental conditions close to those supported by human cells [1]. However, some organisms do not operate under these conditions and are adapted to extreme environments. Microorganisms that develop and grow under unusual environmental conditions are referred to as extremophiles, a term coined by Macelroy in 1974 [2]. Extremophiles are microorganisms that can grow in extreme physical or geochemical conditions [3], including high and low temperatures, extreme pH or salinity, presence of radiation, extreme pressure, or very high hydrocarbon concentration [4].
Despite the general preference of fungi to grow at neutral or slightly acidic pH, it has been demonstrated that some can also thrive at high pH [5]. These organisms produce alkaline enzymes capable of resisting high pH (proteases, amylases, and lipases) and can be used in the textile, food, pharmaceutical industry, etc. [6,7,8]. Even today, the terms alkaliphilic and alkali-tolerant can be perplexing because this classification is highly dependent on the growth conditions and culture medium composition. Horikoshi (1999) [9] postulated that alkaliphiles are microorganisms that grow optimally or exceptionally well at pH values above 9, typically between 10 and 12, but cannot grow or grow only slowly at the near-neutral pH value of 6.5. Grum-Grzhimaylo et al. (2016) [5] proposed an even more comprehensive classification; they define alkaliphiles as organisms that thrive at pH levels greater than 8. These alkaliphiles can be either obligate (they cannot grow at pH 4–5) or facultative (they can grow at acidic pH). While alkali-tolerant fungi can grow at alkaline pH, their optimal growth occurs at pH levels below pH 8. Depending on their growth profile at different pH, these can be further classified as strongly, moderately, or weakly alkali-tolerant. According to the definition proposed by Grum-Grzhimaylo et al. (2016) [5], we will use the terms alkaliphilic and alkali-tolerant in this review.
Alkaline environments, both terrestrial and aquatic, can be created by nature or by humans. Alkaline environments of anthropogenic origin can be derived from cement manufacturing, indigo dye production, mining operations, paper and pulp production, and food processing effluents, among others [10]. Ca2+ and Mg2+ are indeed initially scarce in natural environments; therefore, the dissolution of atmospheric CO2 coupled with the rapid evaporation of water results in the formation of high pH environments up to pH 12 [1], leaving Na+ as the dominant cation [11]. In calcium-containing systems, however, the accumulated carbonate anion is swiftly involved in the formation of insoluble calcite, CaCO3, and dolomite, MgCa(CO3)2, resulting in an environment that is nearly pH neutral [1]. Nevertheless, each of these ecosystems has its own physicochemical characteristics that impose a unique biodiversity, which is of interest to study fungal adaptative evolution of (poly)extremophilic microorganisms, i.e., alkaliphilic and alkali-tolerant fungi, as well as for its characteristics as a unique source for biomolecules with biotechnological potential.
The pH-dependent signaling system involving the transcription factor PacC/Rim101, which is proteolytically activated by the Pal/Rim signaling pathway, is one of the most thoroughly investigated alkali gene-regulating strategies to modulate metabolism in fungi when grown in alkaline conditions. PacC/Rim101 transcription factor activation activates genes related to alkaline stress and inhibits genes related to growth in the presence of acidic pH [12]. This review aims to synthesize information regarding alkaliphilic and alkali-tolerant fungi, their adaptation mechanisms, related signaling cascades, and the biotechnological significance of the enzymes that can be produced under this stress.
2. Fungal Diversity in Alkaline Environments
It is recognized that abiotic factors inhibit the development of most organisms exposed to extreme habitats. Soda soils (or alkaline soils) with pH values higher than eight are an example of such extreme environments [5]. It could be assumed that the present biota in extreme ecosystems is limited, however, numerous studies have demonstrated otherwise; not only bacterial species but also eukaryotic species of various taxonomic levels have been found [5,13,14,15,16].
In the case of fungi, it is difficult to isolate alkaliphilic and alkali-tolerant strains because, in many instances, neutral or acidic isolation media are used to study the fungal diversity of alkaline soils, thereby decreasing the likelihood of recovering alkaliphilic strains [5]. Steiman et al. (2004) [17] isolated a broad variety of fungi from samples of soil and sediment with pH ranging from 8 to 9.2, which were obtained from Mono Lake (USA). The authors used Malt Extract Agar with a pH = 8 for the isolation, but no buffer to maintain the pH of the culture medium was added. This is not recommended for the isolation of alkaliphilic and/or alkali-tolerant fungi since their metabolic processes can modify the pH of the culture medium. Many of the isolates obtained from Mono Lake are likely alkali-tolerant for the aforementioned reasons. Nonetheless, the environmental conditions present in Mono Lake (USA), such as high alkalinity and salinity, are pertinent to isolating unique phenotypes of alkaliphilic fungi.
When isolating fungi from alkaline environments, the fungal identification was an important methodological challenge for early mycologists, as this was accomplished by analyzing the morphophysiological characteristics of the isolated fungi. The advent of massive sequencing made it feasible to more efficiently classify the different taxonomic groups and reclassify many of the initially isolated species. Many of the identified species in Mono Lake (USA), Iwate Prefecture (Japan), and Jayapura (Indonesia) have been reclassified into other taxonomic groups. Figure 1 shows the biodiversity of fungi in certain alkaline environments.
Despite the fact that alkaline environments inhibit the development of the majority of known mesophilic organisms, diversity bioprospection of these ecosystems has revealed a wide variety of fungi (Figure 1). Species of Acremonium and Sodiomyces have been isolated from different alkaline environments. The species of these two genera are present in fifty percent of the studied environments, and some of these species can thrive at a pH range of 10 and 11 [5]. Both genera have been isolated from soil and water samples; however, the ecosystems where Acremonium sp. was isolated had a pH range of 7 to 11, whereas Sodiomyces sp. was more exclusive of habitats with pH values above 9 due to their obligate alkaliphilicity. Other isolated species include those of the genera Fusarium and Penicillium, which were recovered from 30% to 40% of the environments depicted in Figure 1. Some of the strains from these two genera exhibited a mild to moderate alkali-tolerant profile.
On the other hand, since the sampling site in Jayapura, Indonesia, is a cultivation area, the diversity of alkaliphilic and alkali-tolerant fungi in this region is likely influenced by anthropogenic activities. Some of the fungi isolated from these ecosystems are plant pathogens (Cercospora sp., Cylindrocarpon sp., Paramyrothecium roridum), which is related to the sampling site’s characteristics.
The use of culture techniques has permitted the morpho-physiological study of isolated species and is an essential approach for the analysis of biodiversity. Furthermore, culture techniques are also useful to comprehend the mechanisms of adaptation to extreme pH in alkaliphilic and alkali-tolerant fungi. However, there are numerous non-cultivable fungal species whose presence can only be inferred using metagenomic tools.
Here, we also analyzed several metagenomes deposited in the MG-RAST (https://www.mg-rast.org/ (accessed on 7 May 2023)) to increase our understanding of fungal biodiversity in alkaline environments. Four natural environment metagenomes were chosen: Büdös-szék pan (Hungary), Pantanal (Brazil), Mono Lake (USA), and Lonar Lake (India). The Büdös-szék pan is a typical alkaline aquatic environment (pH = 9.16) in the Pannonian biogeographic steppe [28]. The Brazilian Pantanal biome (specifically in the Nhecolândia sub-region) hosts hundreds of pristine soda lakes with a pH gradient varying between 8.62 and 10.05 [29]. Mono Lake is an alkaline and hypersaline closed-basin lake in central California, just east of the Sierra Nevada Mountains [17]. Water samples were used in the metagenomic analysis of these three sites. The hypersaline and hyperalkaline Lonar Lake in Maharashtra, India, is a natural meteorite lake; the metagenome of this site was derived from sediment with a pH range from 9.16 to 10.16 [30].
Analyzing the relative abundance at the family level, we did not observe a significant difference between metagenomes (Figure 2). However, there are differences between the species identified in these four metagenomes and the isolated species shown in Figure 1. Chaetomiaceae, Cordycipitaceae, Mycosphaerellaceae, Onygenaceae, and Pleosporaceae were among the families recovered by both cultivable and metagenomic methodologies. Approximately 80% of the predicted families in the metagenomes were not recovery by cultivable methods, as shown in Figure 1. These findings demonstrate the significance of conducting biodiversity studies using metagenomic libraries to study a broader spectrum of the fungi present in these extreme environments.
3. Physiology
Most alkaliphiles have an optimal growth pH of around 10, which is the most significant difference from the generally investigated neutrophilic microorganisms; however, their intracellular enzymes have a maximum activity near neutral pH [31]. This evidence suggests that alkaliphilic and alkali-tolerant fungi have evolved mechanisms to overcome the effect of basic pH. Under alkali conditions, it is likely that fungi possess certain mechanisms to alleviate the influence or damage of alkali: H+ proton sequestration in different cellular compartments [32]. In addition, some neutrophilic fungi isolated in soils produce different organic acids to solubilize phosphate salts [33]; a similar mechanism could probably be found in alkaliphilic and alkali-tolerant fungi that contributes to the acidification or neutralization of alkaline soils [34] and morphological arrangements of membranes [35], among others.
Much of the literature that describes the mechanisms involved in alkaline pH resistance is primarily concerned with bacteria and neutrophilic fungi, but these mechanisms may also occur in alkaliphilic and alkali-tolerant fungi. In this section, we will discuss a number of these pieces of evidence and relate them to potential pH homeostasis mechanisms in alkaliphilic and alkali-tolerant fungi.
3.1. Genes, Transcription Factors, and Signaling Mechanism
The eukaryotic organisms in which gene regulation by pH has been most extensively investigated are the ascomycete fungi Aspergillus nidulans (Pal pathway) and Saccharomyces cerevisiae (Rim pathway) [12]. Both models are neutrophilic fungi; however, there are some reasons to use them in studies of pH variation. For instance, mutations that affect pH regulation cause phenotypic variations that are easy to identify and study in mutant lines. In the case of A. nidulans, the ability to detect acid and alkaline phosphatases by a simple colony staining test was decisive [36], while mutations in the Rim genes of S. cerevisiae cause easily detectable phenotypic changes such as poor growth at low temperatures, morphological alterations in the colonies, inefficient sporulation, and defective invasive growth [37].
pH is regulated by a signal transduction pathway that leads to the proteolytic activation of a zinc finger transcription factor called PacC [38,39] (Figure 3). PacC72 possesses 674 amino acids [40,41] and undergoes post-translational modifications when extracellular pH is neutral or alkaline. PacC72 acquires a closed conformation at acidic pH, which is maintained by the interaction of a C-terminal domain with two upstream domains, thereby preventing proteolytic processing. When the pH signal is received, the protein conformation changes to an open state [42]. PacC72 contains a “signaling protease box” that removes roughly 180 C-terminal residues, resulting in ∼495 residues (PacC53) [43]. PacC undergoes a second processing step in which an additional ∼245 C-terminal residue is removed, resulting in the ∼250 residues (PacC27) [40,43,44]. This second proteolysis reaction is pH-independent [43]. PacC27 inhibits acid-expressed genes while activating alkaline-expressed genes [45]. The PacC27 form exposes the DNA-binding domain (DBD) containing three Cys2His2 zinc fingers (core consensus: 5-GCCARG) [45] that is sufficient to drive preferential nuclear localization [46,47]. PacC27 and PacC53, but not PacC72, exhibit marked nuclear localization preference [47]. Finger 1 interacts with finger 2 instead of DNA, whereas residues within fingers 2 and 3 interact with target DNA [41,46,48].
Ascomycota’s Pal/Rim pathway comprises six gene products: Rim21/PalH, Rim8/PalF, Rim20/PalA, Rim13/PalB, Rim23/PalC, and Rim9/PalI [37,38,39,49,50,51]. The products of six genes, palA, palB, palC, palF, palH, and palI, transmit the alkaline extracellular pH signal to PacC [38,39]. Null mutations in any of the six pal genes of the pH signal transduction pathway and in pacC have an acidity-mimicking phenotype [38,39].
The pH signaling begins at a plasma membrane complex consisting of the 7-transmembrane (7TM) domain protein Rim21/PalH, the arrestin Rim8/PalF, and, most likely, the 3-transmembrane domain protein Rim9/PalI [52,53]. PalI is thought to aid in the localization of PalH to the plasma membrane, according to [52]. The C-terminal tail of PalH interacts strongly with the arrestin-like protein PalF, which contains an N-domain and a C-domain that are conserved in the arrestin family and are likely responsible for recognizing and binding to the 7TM receptor. In an alkaline pH, PalF is phosphorylated/ubiquitinated in a palH-dependent and partially PalI-dependent mechanism [53]. Arrestins play key roles in signal transduction by interacting with the cytoplasmic domains of 7TM receptors and are known to participate in endocytosis by connecting them to the endocytic internalization apparatus [54].
PalC is recruited to sites near the plasma membrane in an ambient pH- and 7TM receptor PalH-dependent manner, but its recruitment is PalA-independent. In addition, PalC binds to Endosomal Sorting Complex Required for Transport-III (ESCRT-III) Snf7p/Vps32, indicating a potential role as a link between the upstream and downstream pH signaling complexes [55]. Although multiple studies associate pH signaling proteins with endocytosis, Lucena-Agell et al. (2015) [56] demonstrated by endocytosis block that PacC processing is not significantly affected, indicating that pH signaling proteins probably do not require endocytosis.
pH signal transduction requires that the PalA protein binds to two YPXL/I motifs located at either side of the signaling protease box in PacC72, where Tyr, Pro, and Leu/Ile are crucial for its interactive properties, required for the pH-regulated cleavage of this transcription factor [57]. PalB, a cysteine protease of the calpain family, is probably the signaling protease that causes the first proteolytic processing of PacC72 but not the second proteolytic processing [58].
Data from several research groups strongly suggest that PalA/Rim20 physically interact with certain class E Vps protein complexes, acting at the cytosolic side of endosomes to mediate the sorting of transmembrane proteins into the multivesicular body pathway [57]. The endosomal trafficking pathway plays a fundamental role in the interaction of cells with their environment [59] and has critical implications on signal transduction and cell growth control. In addition, evidence suggests a connection between this pathway and pH-dependent signaling mechanisms [60].
On the other hand, the ESCRTs encompass a set of interacting protein complexes that recognize ubiquitinated membrane proteins (Ub-cargo) and facilitate their sorting into intralumenal vesicles [61]. The ESCRT-I, -II, and -III protein complexes function to create multivesicular bodies for sorting of proteins destined for the lysosome or vacuole [62]. ESCRT-0 and ESCRT-I capture cargo and initiate intralumenal vesicle formation, after which ESCRT-II and, ultimately, ESCRT-III are recruited to the ESCRT apparatus to complete vesicle formation and fission from the endosomal limiting membrane [61].
Xu et al. (2004) [60] determined whether the ESCRT components were required for the catalytic processing of Rim101, which was dependent on ESCRT-III subunits Snf7p and Vps20p but independent of Vps2p and Vps24p. Rim101 processing was also dependent on ESCRT-II subunits Snf8p (Vps22p), Vps25p, and Vps36p and on ESCRT-I subunits Vps23p, Vps28p, and Vps37p. Therefore, the majority of ESCRT subunits are required for Rim101 processing. However, it is likely that the activation of Rim/Pal genes does not require endocytosis [56]. Changes in extracellular pH alter the charge of the inner leaflet of the fungal plasma membrane, thereby modifying the biochemical interactions of the Rim21/PalH pH sensor with the cytoplasmatic membrane, for example in S. cerevisiae [63]. Rim8/PalF decode the alkaline pH signal received by Rim21/PalH, and PalF/Rim8 ubiquitination plays a positive role in pH signaling by recruiting ESCRTs and PalB to sites near the plasma membrane [56].
Some neutrophils have been extensively used as biological models to study pH signaling; however, these studies in alkaliphilic fungi are scarce. Using genomic tools, we can infer the presence of potential adaptations to pH if we demonstrate that alkaliphilic/alkali-tolerant fungi have the genes that encode for the Pal pathway. The genome of the alkaliphilic fungus Sodiomyces alkalinus was sequenced (BioSample: SAMN02745840) [64], and the UniProt annotation [65] shows that this fungus presents practically all the Pal pathway genes (PalI, PalH, PalF, PalC, PacC). Additionally, it contains sequences homologous to V-type proton ATPase. Proton-translocating ATPases (H+-ATPases) are rotary enzymes that couple proton (or Na+) translocation across membranes with ATP synthesis or hydrolysis. Vacuolar or V-type ATPases work in reverse mode by actively pumping protons through membranes using energy derived from ATP hydrolysis [66,67].
Another mechanism associated with pH homeostasis is the Na+/H+ antiporter system, which has been very well studied in bacteria, but very little is known about this response in alkaliphilic fungi. Neutralophilic bacteria utilize the proton electrochemical potential (ΔμH+), a sum of transmembrane pH gradient (ΔpH; inside alkaline) and membrane potential (Δψ; inside negative), in the active transport of solutes, ATP synthesis, and motility [68]. Alkaliphilic bacteria acidify the cytoplasm by a Na+/H+ antiporter system and consequently exhibit ΔpH in the opposite direction to that in neutrophiles [69]. Thus, instead of ΔμH+, alkaliphiles utilize a sodium electrochemical potential (ΔμNa+), a sum of transmembrane sodium gradient (ΔpNa; outside > inside) and Δψ, in the active transport of amino acids and motility [69,70]. Interestingly, S. alkalinus also has a gene that encodes an Na+/H+ exchanger. The presence of orthologs of V-type ATPases and Na+/H+ exchanger gene in S. alkalinus may be indicative of an adaptation mechanism in response to pH.
3.2. Membrane Adaptations
Glycerophospholipids and sphingolipids contribute to the plasma membrane’s asymmetry. Phosphatidylcholine (PC) and complex sphingolipids are predominantly located in the outer leaflet (extracytosolic) of the lipid bilayer. In contrast, phosphatidylserine (PS), phosphatidylethanolamine (PE), and phosphatidylinositol (PI) are restricted to the inner leaflet (cytosolic) [71,72]. The lipid translocases or flippases are responsible for moving the lipids in the membrane and establishing their asymmetry [73]. Among the translocases reported, ABC transporters catalyze flop, which is the movement from the cytosolic to extracytosolic leaflet, whereas P4-type ATPases stimulate flip, the reverse movement [74], and scramblases randomize the lipid distribution between the two leaflets [75]. This asymmetric structure, characterized by a greater number of anionic lipids in the cytosolic leaflet and predominantly neutral lipids in the extracellular leaflet, generates two surfaces with radically distinct electrostatic potentials [76].
There are multiple described mechanisms in fungi for maintaining intracellular homeostasis when the pH is acidic [32]. However, the relationship between plasma membrane lipid asymmetry and alkaline pH in the extracellular environment is poorly understood. Several studies demonstrate that the deletion of translocase genes in yeasts such as Cryptococcus neoformans and S. cerevisiae impairs intracellular traffic and organelle structure maintenance [77,78]. The majority of studies on membrane asymmetry and its relationship with pH are focused on S. cerevisiae, limiting our understanding of the effects of alkaline pH on the membrane in other taxonomic groups of fungi.
In S. cerevisiae, five proteins belonging to the P4-type ATPases family are present: Dnf1, Dnf2, Dnf3, Neo1, and Drs2 [76]. Deletion of the dnf1 and dnf2 genes abolishes ATP-dependent flip of fluorescent-labeled PC, PE, and PS, indicating that Dnf1 and Dnf2 have important roles in the asymmetry stability of plasma membrane glycerophospholipid [73,79]. Dnf1 and Dnf2 require association with a common subunit, Lem3 (also known as Ros3), a member of the Cdc50 family [80,81].
Many ABC transporters implicated in multidrug resistance are present in yeasts. Nevertheless, the presence of a large number of ABC transporters in yeast genomes and the fact that not all ABC members are directly involved in multidrug export indicate that they perform essential physiological functions independent of those related with the presence of drugs [82]. For instance, Pdr5 and Yor1 are believed to be implicated in glycerophospholipids flop in yeast S. cerevisiae [79,83]. pdr5 and yor1 are transcriptionally regulated by Pdr1, and their up-regulation results in a gain of function. The pdr1-3 mutant reduces the accumulation of labeled PE, most likely owing to an increase in efflux [83]. A Pdr5/Yor1-dependent increase in endogenous PE on the cell surface was later confirmed [79].
Although glycerophospholipid translocation mechanisms has been well studied, a similar system for sphingolipid translocation has yet to be disclosed [84]. Rsb1p was identified as a putative sphingoid long-chain base-specific translocase/transporter that mediates flop or efflux of the long-chain base in an ATP-dependent manner [85] (Figure 4). Rsb1p expression may be mediated by alterations in glycerophospholipid asymmetry. In wild-type S. cerevisiae, under normal growth conditions in complex media, the expression level of rsb1 is low, whereas its expression is significantly elevated if the glycerophospholipid asymmetry is disturbed. This alteration can be caused by mutations in genes involved in the flip (i.e., Dnf1, Dnf2, or Lem3) or the flop (i.e., ABC transporters Pdr5 and Yor1) of glycerophospholipids [84]. Interestingly, rsb1 is also induced by Rim13, Rim20, and Rim21. Mutations in any of these genes decrease rsb1 expression levels in pdr5Δ cells, indicating that Rim21, Rim20, and Rim13 regulate the rsb1 expression via Rim101. Therefore, all mutations known to impair Rim101 activation (rim21Δ, dfg16Δ, rim9Δ, rim8Δ, rim13Δ, rim20Δ, ygr122wΔ, vps28Δ, vps25Δ, snf7Δ, and vps20Δ) caused rsb1 expression reduction [73].
Phosphatidylserine is heterogeneously and asymmetrically distributed throughout the cell and constitutes more than 30% of the lipid on the inner leaflet of the plasma membrane; this, combined with its negatively charged head group, confers onto phosphatidylserine a nonpareil ability to direct the recruitment of proteins containing polycationic stretches as well as proteins that possess a specific PS-recognition site [86]. Under normal conditions, in the absence of environmental stress, negatively charged phospholipids are confined to the inner leaflet of the plasma membrane. However, changes in lipid asymmetry can expose these negatively charged phospholipids on the cell surface. The putative sensors of the Rim101 signaling pathway may identify the exposed negative charge similarly to how they identify the charge in alkaline culture medium [73]. Young et al. (2010) [87] demonstrated the role of phospholipids in pH signaling, specifically the function of phosphatidic acid as a cellular pH biosensor. The binding of proteins to the ubiquitous signaling lipid phosphatidic acid was dependent on the intracellular pH and protonation state of the phosphate head group. This evidence demonstrates that membrane asymmetry plays a crucial role in cell signaling and intracellular pH maintenance.
We have previously discussed the significance of membrane asymmetry as a signaling mechanism and adaptation to alkaline pH. However, recent reports indicate that membrane composition may also play a significant role in alkaline pH resistance. For instance, Danilova et al. (2020) [88] evaluated the composition of the cytoplasmic membrane of the haloalkali-tolerant fungus Emericellopsis alkalina at different pH (4.5, 7, 10.2), and they found that changes of the pH had little effect on the composition of membrane lipids. Only an increase in the proportion of phosphatidic acids in neutral conditions (pH 7.0) and an increase in the proportion of sterols in alkaline conditions (pH 10.2) were observed. Phosphatidic acid also increases when growing Sodiomyces magadii, S. alkalinus, and S. tronii at neutral pH, but according to the graphs shown by the authors, the sterols had a different behavior compared to E. alkaline; at alkaline pH, there is a decrease in the proportion of sterols [89,90].
This behavior of little fluctuation in membrane composition in response to pH and variation in phosphatidic acids and sterols was also demonstrated in the alkali-tolerant fungi Acrostalagmus luteoalbus and Chordomyces antarcticus [90]. The behavior of phosphatidic acid at different pH is similar between alkaliphilic and alkali-tolerant fungi, with a higher proportion in neutral conditions but a decrease in sterols at alkaline pH. The differences in sterols between the alkaliphilic/alkali-tolerant fungi (S. magadii, S. alkalinus, S. tronii, A. luteoalbus, and C. antarcticus) with E. alkalina may be because the latter was cultivated under saline conditions (0.4 M NaCl).
The total lipids in the membranes of alkaliphilic and alkali-tolerant fungi is another interesting feature to examine. Alkaliphiles exhibit an increase in total lipids at acidic/neutral pH, whereas alkali-tolerant organisms exhibit an increase at alkaline pH [90]. This is likely because alkaliphilic fungi experience stress in an acidic pH environment, whereas alkali-tolerant fungi thrive in an acidic pH environment. Although there is evidence of little change in the composition of the cytoplasmic membrane in response to pH, these changes in total lipids may indicate that we do not completely comprehend the membrane’s behavior at different pH levels.
3.3. Protein Adaptations
During evolution, organisms achieved viability under extreme conditions either by ‘escaping’ or ‘compensating’ the stress or by enhancing the stability of their cellular inventory [91]. Although a great deal is known about the adaptation of proteins to elevated temperatures [92,93] and salinity [94,95], there are few comparable studies on the effect of pH in fungi. In addition, the majority of the prior research on alkaline pH stability has been conducted on bacterial proteins [10], whereas reports on fungal proteins are scarce and based on studies of enzymatic activities in mesophilic fungi. We are unaware of any enzymatic investigations on alkaliphilic or alkali-tolerant fungi that explain their activity at alkaline pH. It would be premature to discuss the causes within fungi that determine the stability of acidophilic and alkaliphilic proteins. However, research indicates how these potential determinants may be.
The pH activity profiles of enzymes are highly dependent on the pKa of the catalytic residues, which are themselves dependent on the local environment and, thus, on the nature of the amino acids in the vicinity of the catalytic residues [96]. Furthermore, some data indicate a role for hydrogen bonds and salts bridges in protein stabilization [97,98].
It is likely that some amino acids have an important role in catalytic activity and in enzyme stability in alkaline pH. Wang et al. (2005) [99] analyzed the pH profile of endoglucanase III from neutrophilic Trichoderma reesei to determine the effect of substitutions at surface residue N321, where two catalytic residues are located. N321D mutation caused a significant shift in the optimum pH of the enzyme from 4.8 to 4.0, whereas N321H maintained its highest activity in a pH range from 4.6 to 5.5 and more than 84% of its specific activity at pH 6.0. Positive charges near the catalytic cleft probably promote enzyme activity at higher pH, but there is currently no evidence to support this hypothesis in enzymes isolated from alkaliphilic or alkali-tolerant fungi.
Other examples are certain xylanases that are stable in alkaline conditions, which are typically characterized by a decreased number of acidic residues and an increased number of arginine [100]. Substitutions of Ser/Thr surface by arginine residues in Trichoderma reesei endo-1,4-β-xylanase II shifted the activity profile to the alkaline region by 0.5–1.0 pH units [100]. Luo et al. (2012) [101] studied the alkali-tolerant β-mannanases of Humicola insolens Y1, which displays high activity at pH 9.0 and over 10% at pH 10.0, in order to determine the particularities that allowed it to resist alkaline pH. The β-mannanases of H. insolens Y1 have 11% alkaline amino acid residues (Arg and Lys), compared to the acid-active mannanases (3.7–5.3%), and most of the alkaline residues are located on the protein surface. It is probable that the higher pKa values, high frequency of alkaline amino acid residues, and more positively charged residues might be the factors that influence the protein activity and stability under neutral and alkaline conditions [101].
The formation of hydrogen bonds between amide–carboxylate pairs can play an important role in the stability of proteins. Substitutions of carboxyl–carboxylate pairs to amide–carboxylate pairs in Trichoderma reesei cellobiohydrolase showed an increase in protein stability at alkaline pH. This is probably because the carboxyl–carboxylate pairs are negatively charged at alkaline pH, and the strong repulsion causes conformational changes that weaken the stability of the protein. In contrast, amide–carboxylate pairs can maintain the hydrogen bond at different pH [102].
According to the evidence, the primary strategies for resisting alkaline pH at the protein level are variations in pKa, the formation of hydrogen bonds between amide–carboxylate pairs, and the presence of alkaline amino acids on the surface of proteins. However, amino acid substitution experiments shifted the optimum enzyme pH by less than one unit of pH towards alkalinity. In addition to the interactions discussed in this section, it is probable that other interactions contribute to the stability required to withstand high pH values.
4. Biotechnological Impact of Alkaliphilic and Alkali-Tolerant Fungi
Extremophiles, uncommon microorganisms that thrive in extreme environments of temperature, pH, salt, sugar concentrations, and atmospheric pressure, have been identified over the past four decades [10]. Extremophile microorganisms provide robust enzymatic and whole-cell biocatalytic systems that are alluring under conditions that limit the efficacy of conventional bioconversions [4]. The extraordinary survival strategies of extremophiles can be linked to the production of extremozymes and other secondary metabolites with numerous biotechnological applications [103,104]. Fungi must be able to generate and secrete a large number of hydrolytic enzymes that degrade plant organic matter (e.g., maize, wheat, or rice straw), such as cellulose, hemicellulose, and lignin, in order to proliferate in soil [34]. In the case of alkaliphilic and alkali-tolerant fungi, they are known to produce alkaline proteases, cellulases, chitinases (Table 1), and other biotechnologically significant metabolic products, such as carotenoids, steroids, antibiotics, and organic acids [9]. Alkali-resistant enzymes isolated from mesophilic fungi also have biotechnological potential, constituting a second relevant group.
In general, microorganisms must exist in environments where the nutrients are primarily macromolecular in nature. These nutrients are inaccessible to microbes unless they are broken down into molecules that they can assimilate. An enzyme secreted by microorganisms breaks down macromolecular nutrients into smaller molecules. Alkaline proteases have proven beneficial in the detergent, food, pharmaceutical, and tanning industries [105].
The most practiced method for the depilation of hides and skins is the lime–sulfide process; because sulfur is toxic, enzymatic depilation has gained more acceptance [106]. The alkaline protease from Aspergillus flavus was obtained by solid-substrate fermentation and its characteristics allow it to be used for this purpose [107]. A similar protease was isolated from Aspergillus clavatus; its high proteolytic activity at alkaline pH, at moderate temperature and stability towards various surfactants and bleaching agents, suggest its suitability for inclusion in liquid detergent compositions [108]. Detergents are chemical compounds used for cleaning purposes. However, stains of blood and soil being proteinaceous in nature are not easily removed by detergent chemicals. Therefore, proteases have an important role in enhancing detergent cleaning performance [10]. Therefore, identifying, isolating, and characterizing alkaline proteases which do not lose their activity in detergents is of interest to biotechnologists.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Review of alkaline enzymes and alkali-resistant enzymes found in the literature.
| Enzyme | Microorganism | pH Enzyme Production | Optimum pH for Activity | Optimum T (°C) for Activity | References |
|---|---|---|---|---|---|
| Alkaline protease | Aspergillus flavus | 7 | Optimum activity in the pH range of 7.5–9.5. | Optimum activity at 42 °C. | [107] |
| Neosartorya clavata (syn. Aspergillus clavatus) | 8 | Optimum activity at pH 8.5. It is active in the pH range of 6.0–11.0. The enzyme is stable after an hour of incubation at pH 8.0–9.0. | The enzyme was active between 40 and 70 °C with an optimum around 50 °C. Stable activity after 1 h treatment at 30 °C. | [108,109] | |
| Sodiomyces alkalinus | 10 | Optimum activity at pH 8. | Optimum activity at 30 °C. | [64] | |
| Emericella usta (syn. Aspergillus ustus) | 9 | Enzyme active in a wide pH range (6.0–10.0), with an optimum at pH 9.0. | Optimum activity at 45 °C. | [110] | |
| Serine protease | Aspergillus parasiticus | - | Optimum activity at pH 8. Stable activity after 1 h treatment in pH 6.0–10.0. | Optimum activity at 40 °C. The enzyme was stable at 40 °C for 1 h incubation but was inactivated at temperatures over 40 °C | [111] |
| Parengyodontium album (syn. Engyodontium album) | 10 | Enzyme active in a wide pH range (6.0–12.0), with an optimum at pH 11.0. | Optimum activity at 60 °C. More than 90% of the maximal activity was conserved between 45 and 65 °C. | [112] | |
| Clonostachys rosea (syn. Gliocladium roseum) | 7–7.5 | Optimum activity in the pH range of 9.0–10.0. | Optimum activity at 60 °C. | [113] | |
| Trichoderma reesei | 6 | Enzyme active in a wide pH range (6.0–11.0) with an optimum at pH 8.0. | Optimum activity at 50 °C. | [114] | |
| Dipeptidyl peptidase 4 | Chordomyces antarcticus | 10 | Optimum activity at pH 7.7. Stable in the pH range of 3.0–12.0. | Optimum activity at 37 °C. | [115] |
| Sodiomyces alkalinus | 10 | Optimum activity at pH 7.3. Stable in the pH range of 5.0–12.0 and retained 23% of its activity after an hour of incubation at pH 13.0 | Optimum activity at 37 °C. | [115] | |
| Trypsin-like protease | Cordyceps militaris | 6 | Optimum activity in the pH range of 8.5–12.0. | Optimum activity at 25 °C. | [116] |
| Xylanase | Penicillium citrinum | - | Optimum activity at pH 8.5. | Optimum activity at 50 °C. | [117] |
| Cladosporium oxysporum | 7–8 | Optimum activity at pH 8.0. Stable activity after 2 h treatment in pH 7.0–8.5. | Optimum activity at 50 °C. Stable activity after 2 h treatment below 55 °C. | [118] | |
| Aspergillus fischeri | 9 | Optimum activity pH 6.0. Retained over 50% of maximum activity at pH 8.0. pH stability ranged from 5.5 to 9.5 with retention of more than 85% of the activity. | The optimum temperature was 60°C. | [119] | |
| Aspergillus fumigatus | 8 | Optimum pH 8. Substantial residual activity at alkaline pH 8–9 (56–88%). | Optimum temperature 50 °C. Substantial residual activity at 60–70 °C (53–75%). | [120] | |
| Amylases | Clavispora lusitaniae | 7 | Optimum activity at pH 11. Enzymes retained nearly 80% of activity after being exposed to various detergent components for 2 h. | Optimum activity at 40 °C. The enzyme retained 45% and 98% of their maximum activity at 4 °C and 25 °C. | [121] |
| Sporormiella minima (syn. Preussia mínima) | Optimum pH 9. | Optimum activity at 25 °C. | [122] | ||
| Endoglucanase B | Aspergillus niger (The gene encoding the enzyme was cloned and expressed in Pichia pastoris) | - | Stable for 2 h at alkaline pH (7–10). | Stable for 3 h at temperatures below 60 °C. | [123] |
| Endoglucanases | Mycothermus thermophilus (syn. Humicola insolens) (The gene encoding the enzyme was cloned and expressed in Aspergillus oryzae) | - | Optimal activity between pH 7 and 8.5. | [124] | |
| Cellobiohydrolase II | - | Optimal activity pH 9. | |||
| Laccase | Albifimbria verrucaria (syn. Myrothecium verrucaria) | 9 | Optimum activity at pH 9.0 and retained more than 80% of the initial activity after 17 h incubation at 30 °C at pH 8–11.5. | Stable for 1 h at temperatures below 50 °C and retained more than 80% of the initial activity. | [125] |
| Lipases | Trichoderma lentiforme (The gene encoding the enzyme was cloned and expressed in Pichia pastoris) | - | Poor activity under acidic conditions and maximum activity at pH 9.5. The enzyme was stable at pH 6.0–9.0, retaining more than 80% of the initial activity after 1 h pre- incubation at 37 °C. | Temperature optimum at 50 °C and the enzyme was relatively stable at 40 °C, retaining more than 60% of the initial activity after 1 h incubation. | [126] |
| Aspergillus carneus | 8 | The purified enzyme could tolerate pH 6.0–12.0 and it was stable in this range for 24 h. The optimum pH was 9.0. | The optimum temperature was 37 °C and the enzyme was active in the range of 5- 90 °C. | [127] | |
| Lasiodiplodia theobromae | 8 | Optimum activity pH 8.0. | Optimum temperature was 30 °C. | [128] | |
| Gliocladium sp. | - | Optimum activity pH 10. | The assay temperature was 40 °C. | [129] | |
| Verticillium sp. | - | The enzyme was fully stable for 30 min at pH 9 at temperatures up to 50 °C. The enzyme was stable throughout the pH range 6–10 at 25 °C for 24 h. | [129] |
The discovery of alkaline cellulases allowed for the formulation of cellulase-containing detergents. There are many bacterial cellulases that, unlike other detergent enzymes, do not degrade or modify the dirt or stains; instead, these enzymes modify the surface of the garment cellulosic fiber to improve the overall washing performance [130]. Filamentous fungi are excellent organisms that can function as cell factories for the production of a variety of products. They are robust and naturally produce efficient enzymes for the decomposition and conversion of biological material [131]; however, most fungal cellulases are active in acidic or neutral conditions [130], hence the importance of isolating and characterizing alkaline cellulases.
Another interesting application of cellulases is in the textile industry. These enzymes have been incorporated into the refining process of cellulose-based textiles in an effort to enhance the feel and appearance of the textiles [132]. Although neutral and acid active cellulases are used in the biostoning process, alkaline active cellulases have been suggested for this process because they prevent backstaining [133]. Endoglucanase B identified in Aspergillus niger was cloned and expressed in Pichia pastoris [123]; the enzyme activity was increased by 2-fold by Triton X-100 and Tween 80. Endoglucanase B is a potential candidate for use in laundry and textile industrial applications due to its favorable properties. The paper recycling process includes alkaline pH and temperatures close to 60 °C [134]; therefore, the use of alkaline and thermostable cellulases could enhance toner removal from wastepaper and facilitate the recycling process.
Similarly, laccases can also be used in the paper industry to decolorize and decompose dyes and phenolic compounds or to remove toxic aromatic compounds in wastewater from various industries [125]. Therefore, laccase from Albifimbria verrucaria (syn. Myrothecium verrucaria) can be used in those scenarios (Table 1).
On other hand, recently, alkaline xylanases have attracted increasing interest in their applications in paper and feed industries, textile processes, enzymatic saccharification, and waste treatments [118]. The modern pulp and paper industry employs various techniques for recovering cellulose fibers from lignocellulosic biomass, the majority of which is derived from wood. This procedure employs alkaline and high-temperature treatments to degrade and solubilize lignin, which must then be eradicated via a chlorine-based bleaching procedure [130]. The use of chlorine results in the formation of highly toxic and mutagenic chlorinated organic by-products [135]. Xylan hydrolysis facilitates lignin removal; therefore, enzymatic treatment permits a substantial reduction in chlorine use [136]. In addition, alkaline xylanase could treat material with a high pH without the need for costly and time-consuming pH adjustments [118]. The xylanases listed in Table 1 have potential industrial applications in textile and paper manufacturing.
Excessive salt accumulation in soil imposes many negative effects on its organic matter decomposition and uptake of available nutrients [137,138]. Some halo- and/or alkaliphilic fungi are promising agents for bioremediation of saline and alkaline soils. In addition, these fungi can be considered as genetic pools that represent a resource for cloning genes related to saline/alkali-resistance or tolerance [139] and organic matter degradation, which can be used to improve or create highly active fungi for soil remediation [34]. Another attractive property of the alkaliphiles is their ability to produce organic acids. Aspergillus glaucus is an haloalkaliphilic fungus that produces a variety of organic acids, including citric acid, oxalic acid, and malic acid [140]; therefore, they can be used for soil mycoremediation [34]. A. glaucus was evaluated for the bioremediation of saline–alkaline soil in the Songnen plain of Northeastern China. This study primarily indicates that the applied amendments mixed with haloalkaliphilic fungi significantly encourage steady growth and yield of rice in comparison with that achieved in the control plot [141].
Another group of less-studied fungal alkaline enzymes and of great importance are starch degrading enzymes [130]. Amylases have applications in starch processing [142], bakery and sugar production [143], desizing in textile industries [144], and detergent manufacturing [145]. The biochemical characteristics of amylases from Clavispora lusitaniae and Sporormiella minima (Table 1) suggest that theses enzymes may find application in the laundry detergent and textile industries [121,122]. The fungal amylases are preferred over other microbial sources because fungi are generally safer, by the hyphal mode of growth, and their good tolerance to low water activity and high osmotic pressure conditions make fungi most efficient for bioconversion of solid substrates [146].
Lipases are a group of enzymes with significant physiological and industrial significance [147]. Lipases catalyze the hydrolysis of triacylglycerols to glycerol and free fatty acids, and in contrast to esterases, they are only active when adsorbed to an oil–water interface [148]. Lipases are prevalent in nature and have been identified in microorganisms, plants, and animals [149,150,151]. Nonetheless, fungal lipases are of particular interest because they are readily produced and advantageous for industrial applications due to their high yields, versatility, and resistance to harsh conditions. Lipases have promising uses in organic chemical processing, detergent formulations, biosurfactant synthesis, the oleochemical industry, the dairy and agrochemical industry, paper manufacturing, nutrition, cosmetics, and pharmaceutical processing [147]. Most of these conditions necessitate detergent-, low-temperature-, and alkaline-pH-resistant lipases; therefore, their isolation and study are necessary.
The properties of alkaline lipases shown in Table 1 allow them to be used in the washing industry. However, the interest in employing lipases to catalyze reverse synthesis reactions (esterification, transesterification) in non-aqueous environments has increased markedly due to their application in the production of biodiesel [152]. Venkatesagowda et al. (2017) [128] evaluated the potential of Lasiodiplodia theobromae lipases, demonstrating their potential for biodiesel production from coconut oil in alkaline conditions.
The screening of extremophiles may also yield other bioactive compounds that can be used in the treatment of non-infectious diseases, including cancer and cardiovascular problems [130]. Rogozhin et al. (2018) [153] studied the ability of the alkaliphilic fungus Emericellopsis alkalina VKPM F1428 to produce antimicrobials. They were able to isolate and identify a Lipopeptaibol Emericellipsin A with antimicrobial activity against fungi and Gram-positive bacteria. They also demonstrated in vitro selective cytotoxic activity against HepG2 and Hela cell lines. Subsequent work on this strain showed that it produces four additional homologues (Emericellipsin B-E) and that Emericellipsin A has activity against clinical isolates of Aspergillus and yeasts [154].
Kuvarina et al. (2021) [155] studied the antimicrobial-producing potential of 25 alkaliphilic fungal strains belonging to Sodiomyces alkalinus. Some of these strains showed antimicrobial activity against the opportunistic pathogens Aspergillus niger INA 00760, Bacillus subtilis ATCC 6633, and Candida albicans ATCC 2091. In one of these strains, S. alkalinus 8KS17-10, a class II hydrophobin with antifungal activity against non- and clinical pathogenic isolates was purified and characterized [156]. We submitted the S. alkalinus genome (BioSample: SAMN02745840) to Antismash [157] to predict biosynthetic gene clusters and assess their potential as a producer of bioactives. The results showed 24 biosynthetic clusters that may be related to Polyketide synthases, Nonribosomal peptide synthetase, ribosomally synthesized and post-translationally modified peptides, and terpenes (Supplementary Material File S2). Many of these clusters are likely to be novel and their products could have higher activity than current bioactives.
Extremely diverse enzymes are produced by alkaliphilic fungi, although the production and commercialization of these enzymes are currently centered on bacteria. The scientific community remains attracted to the use of fungi for the production of alkaline and alkali-resistant enzymes and their subsequent industrial application, particularly since many of these enzymes are extracellular, which facilitates their purification by filtration. This stimulates the search for alkaliphilic fungi, the investigation of their survival mechanisms, and the identification of pH-resistant exoenzymes.
5. Aspergillus sydowii, a Case Study
We have discussed the significance of alkaliphilic and alkali-tolerant fungi in biotechnology throughout this article. However, it is fascinating to provide a further illustration. A. sydowii, a halophilic fungus capable of expressing enzymes with biotechnological potential, was isolated in our laboratory under salinity conditions [158]. It produced cellulases, xylanases, manganese peroxidase, and esterase. González-Abradelo et al. (2019) [159] evaluated the growth of A. sydowii for the removal of polycyclic aromatic hydrocarbons (PAHs) and pharmaceutical compounds. This strain used benzo-α-pyrene and phenanthrene as the sole carbon source in a mineral medium, biodegrading both PAHs (99% benzo-α-pyrene and 97% phenanthrene). In addition, A. sydowii removed (biodegradation and adsorption) more than 90% of the pharmaceutical compounds evaluated (acetaminophen, mefenamic acid, ketoprofen, indomethacin, and ibuprofen). Alkaline and salty environments could be interrelated [5], hence the significance of determining this strain’s alkaline pH resistance.
A. sydowii’s growth on Mineral Medium Agar [160] with 2% glucose (w/v) was evaluated. The medium was supplemented with various NaCl concentrations (0–1.5 M) and pH (4–11.2). To maintain the pH, buffers with a final concentration of 0.2 mM were added to the mineral medium. The buffer choices for generating different final pH values were Na3C6H5O7/citric acid buffer system for pH 4 and 5, Na2HPO4/NaH2PO4 buffer system for pH 6.2, 7, and 8, Na2CO3/NaHCO3 carbonate buffer system for pH 9.2 and 10, and Na2HPO4/NaOH buffer system for pH 11.2 [5]. To ascertain the growth rate (mm/day), we measured mycelium growth daily. The results show that A. sydowii has a better growth at acid–neutral pH; however, it can resist alkaline pH values, evidenced by its growth up to pH 11.2 (Figure 5). Compared to pH 11.2, growth is significantly inhibited at pH 9.2 and 10; however, we believe that this inhibition is not solely attributable to pH. There are reports of the use of sodium carbonate as an antimicrobial [161], and the Na2CO3/NaHCO3 carbonate system was used to evaluate growth at pH 9.2 and 10. We conclude that A. sydowii is alkali-tolerant under our experimental conditions, particularly if we analyze the growth at 0 M NaCl condition, in which there are no statistically significant differences between pH 7, 8, and 11.2. Adding NaCl to the culture medium decreases the growth rate considerably at pH 11.2, indicating that the combined effect of both stresses (salinity and pH) inhibits the growth of A. sydowii.
Previous evidence shows that A. sydowii is halophilic [162]; however, according to our data, the behavior of A. sydowii is halotolerant. This discrepancy is due to differences in cultivation methods. Jiménez-Gómez et al. (2022) [162] used a rich medium (Yeast Malt Agar) supplemented with 1 M NaCl, and in our growth kinetics, the medium used was Mineral Medium Agar supplemented with glucose and NaCl. This difference causes changes in the specific growth rate of A. sydowii, especially since in the mineral medium the stress conditions are stronger compared to the Yeast Malt Agar.
Additional studies should be carried out on A. sydowii to determine which other xenobiotics it can degrade or to understand the mechanisms that allow it to resist alkaline pH. However, current studies reaffirm that the isolation and characterization of halophilic and alkali-tolerant fungi such as A. sydowii, with the ability to degrade xenobiotics under extreme conditions, opens doors to the development of new methodologies for bioremediation. A. sydowii could be an excellent candidate to study the adaptations involved in both extreme growth conditions, NaCl and alkaline pH.
6. Conclusions
Bacteria have been the primary focus of research into organisms that are resistant to environmental pH changes. Recent research reveals that a vast array of fungi can resist both alkalinity and salinity. In recent years, there has been an increase in interest in these organisms as a source of alkaline enzymes for biotechnological applications. Although alkaline pH signaling mechanisms in alkali-tolerant fungi have been investigated, it is still unclear how these fungi maintain intracellular homeostasis and what properties enable their extracellular enzymes to remain active at alkaline pH.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/jof9060652/s1, File S1: Relative abundance at the family level of the phylum Ascomycota present in the metagenomic samples Büdös-szék pan (MG-RAST ID: mgm4555202.3), Pantanal (MG-RAST ID: mgm4875736.3), Lonar Lake (MG-RAST ID: mgm4863970.3), and Mono Lake (MG-RAST ID: mgm4962892.3). File S2: Biosynthetic gene clusters predicted in the S. alkalinus genome by antiSMASH.
Author Contributions
M.G.F.-L. conceived this work and wrote the first version of this manuscript. All authors contributed to the editing of this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Prodep postdoctoral program (2017), SEP, Government of Mexico.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in both the article and in the Supplementary Materials.
Acknowledgments
M.G.F.-L. received a post-doc fellowship from PRODEP (2017), SEP, Government of Mexico. M.G.F.-L. appreciates the support received from PRODEP, Universidad Autónoma de Nuevo León, Mexico. R.A.B.-G. received a Sabbatical fellowship from Conacyt, Government of Mexico (CVU 389616).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Berlemont, R.; Gerday, C. Extremophiles. In Comprehensive Biotechnology; Moo-Young, M., Ed.; Elsevier, B.V.: Liége, Belgium, 2011; pp. 229–242. ISBN 978-0-08-088504-9. [Google Scholar]
- Macelroy, R.D. Some comments on the evolution of extremophiles. BioSystems 1974, 6, 74–75. [Google Scholar] [CrossRef]
- Yun, S.H.; Lee, Y.G.; Choi, C.-W.; Lee, S.-Y.; Kim, S.I. Proteomic Exploration of Extremophiles. Curr. Biotechnol. 2014, 3, 87–99. [Google Scholar] [CrossRef]
- Peeples, T.L. Bioremediation Using Extremophiles. In Microbial Biodegradation and Bioremediation; Das, S., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 251–268. ISBN 978-0-12-800021-2. [Google Scholar]
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Bondarenko, S.A.; Debets, A.J.M.; Bilanenko, E.N. On the diversity of fungi from soda soils. Fungal Divers. 2016, 76, 27–74. [Google Scholar] [CrossRef] [Green Version]
- Matkawala, F.; Nighojkar, S.; Kumar, A.; Nighojkar, A. A novel thiol-dependent serine protease from Neocosmospora sp. N1. Heliyon 2019, 5, e02246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devi, R.; Revathi, K.; Arunagirinathan, N.; Yogananth, N. Production and Optimization of α-amylase from Aspergillus ochraceus isolated from Marakkanam Saltpans, Tamil Nadu, India. Bull. Environ. Pharmacol. Life Sci. 2022, 1, 997–1002. [Google Scholar]
- Hasnaoui, I.; Dab, A.; Mechri, S.; Abouloifa, H.; Saalaoui, E.; Jaouadi, B.; Noiriel, A.; Asehraou, A.; Abousalham, A. Purification, Biochemical and Kinetic Characterization of a Novel Alkaline sn-1,3-Regioselective Triacylglycerol Lipase from Penicillium crustosum Thom Strain P22 Isolated from Moroccan Olive Mill Wastewater. Int. J. Mol. Sci. 2022, 23, 1920. [Google Scholar] [CrossRef] [PubMed]
- Horikoshi, K. Alkaliphiles: Some applications of their products for biotechnology. Microbiol. Mol. Biol. Rev. 1999, 63, 735–750. [Google Scholar] [CrossRef] [Green Version]
- Kanekar, P.P.; Joshi, A.A.; Kulkarni, S.O.; Borgave, S.B.; Sarnaik, S.S.; Nilegaonkar, S.S.; Kelkar, A.S.; Thombre, R.S. Biotechnological potential of alkaliphilic microorganisms. In Biotechnology and Bioinformatics, Advances and Applications for Bioenergy, Bioremediation, and Biopharmaceutical Research; Hangadurai, D., Sangeetha, J., Eds.; CRC Press Taylor & Francis Group: Toronto, ON, Canada, 2015; pp. 249–280. ISBN 0101001010101. [Google Scholar]
- Jones, B.E.; Grant, W.D.; Duckworth, A.W.; Owenson, G.G. Microbial diversity of soda lakes. Extremophiles 1998, 2, 191–200. [Google Scholar] [CrossRef]
- Peñalva, M.A.; Arst, H.N. Recent Advances in the Characterization of Ambient pH Regulation of Gene Expression in Filamentous Fungi and Yeasts. Annu. Rev. Microbiol. 2004, 58, 425–451. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Muyzer, G. Desulfurispira natronophila gen. nov. sp. nov.: An obligately anaerobic dissimilatory sulfur-reducing bacterium from soda lakes. Extremophiles 2010, 14, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, D.Y.; Panteleeva, A.N.; Tourova, T.P.; Kaparullina, E.N.; Muyzer, G. Natronoflexus pectinivorans gen. nov. sp. nov., an obligately anaerobic and alkaliphilic fermentative member of Bacteroidetes from soda lakes. Extremophiles 2011, 15, 691–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antony, C.P.; Kumaresan, D.; Hunger, S.; Drake, H.L.; Murrell, J.C.; Shouche, Y.S. Microbiology of Lonar Lake and other soda lakes. ISME J. 2013, 7, 468–476. [Google Scholar] [CrossRef]
- Lanzén, A.; Simachew, A.; Gessesse, A.; Chmolowska, D.; Jonassen, I.; Øvreas, L. Surprising Prokaryotic and Eukaryotic Diversity, Community Structure and Biogeography of Ethiopian Soda Lakes. PLoS ONE 2013, 8, e0072577. [Google Scholar] [CrossRef] [PubMed]
- Steiman, R.; Ford, L.; Ducros, V.; Lafond, J.-L.; Guiraud, P. First survey of fungi in hypersaline soil and water of Mono Lake area (California). Antonie Van Leeuwenhoek 2004, 85, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Grum-Grzhimaylo, A.A.; Debets, A.J.M.; van Diepeningen, A.D.; Georgieva, M.L.; Bilanenko, E.N. Sodiomyces alkalinus, a new holomorphic alkaliphilic ascomycete within the Plectosphaerellaceae. Persoonia 2013, 31, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Debets, A.J.M.; Bilanenko, E.N. Are alkalitolerant fungi of the Emericellopsis lineage (Bionectriaceae) of marine origin? IMA Fungus 2013, 4, 213–228. [Google Scholar] [CrossRef]
- Lisichkina, G.A.; Bab’eva, I.P.; Sorokin, D.Y. Alkalitolerant yeasts from natural biotopes. Microbiology 2003, 72, 618–620. [Google Scholar] [CrossRef]
- Georgieva, M.L.; Lebedeva, M.P.; Bilanenko, E.N. Mycelial fungi in saline soils of the western Transbaikal region. Eurasian Soil Sci. 2012, 45, 1159–1168. [Google Scholar] [CrossRef]
- Nagai, K.; Suzuki, K.; Okada, G. Studies on the distribution of alkalophilic and alkali-tolerant soil fungi II: Fungal flora in two limestone caves in Japan. Mycoscience 1998, 39, 293–298. [Google Scholar] [CrossRef]
- Nagai, K.; Sakai, T.; Rantiatmodjo, R.M.; Suzuki, K.; Gams, W.; Okada, G. Studies on the distribution of alkalophilic and alkali-tolerant soil fungi I. Mycoscience 1995, 36, 247–256. [Google Scholar] [CrossRef]
- Sharma, R.; Prakash, O.; Sonawane, M.S.; Nimonkar, Y.; Golellu, P.B.; Sharma, R. Diversity and distribution of phenol oxidase producing fungi from soda lake and description of Curvularia lonarensis sp. nov. Front. Microbiol. 2016, 7, 01847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondarenko, S.A.; Georgieva, M.L.; Bilanenko, E.N. Fungi Inhabiting the Coastal Zone of Lake Magadi. Contemp. Probl. Ecol. 2018, 11, 439–448. [Google Scholar] [CrossRef]
- Orwa, P.; Mugambi, G.; Wekesa, V.; Mwirichia, R. Isolation of haloalkaliphilic fungi from Lake Magadi in Kenya. Heiyon 2020, 6, e02823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elíades, L.A.; Cabello, M.N.; Voget, C.E. Contribution to the study of alkalophilic and alkali-tolerant Ascomycota from Argentina. Darwiniana 2006, 44, 63–74. [Google Scholar]
- Szabó, A.; Korponai, K.; Kerepesi, C.; Somogyi, B.; Vörös, L.; Bartha, D.; Márialigeti, K.; Felföldi, T. Soda pans of the Pannonian steppe harbor unique bacterial communities adapted to multiple extreme conditions. Extremophiles 2017, 21, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Pellegrinetti, T.A.; Cotta, S.R.; Sarmento, H.; Costa, J.S.; Delbaje, E.; Montes, C.R.; Camargo, P.B.; Barbiero, L.; Rezende-Filho, A.T.; Fiore, M.F. Bacterial Communities along Environmental Gradients in Tropical Soda Lakes. Microb. Ecol. 2022; under review. [Google Scholar] [CrossRef]
- Chakraborty, J.; Rajput, V.; Sapkale, V.; Kamble, S.; Dharne, M. Spatio-temporal resolution of taxonomic and functional microbiome of Lonar soda lake of India reveals metabolic potential for bioremediation. Chemosphere 2021, 264, 128574. [Google Scholar] [CrossRef]
- Horikoshi, K. Extremophiles. Where It All Began; First; Springer: Tokyo, Japan, 2016; ISBN 9784431554073. [Google Scholar]
- Bignell, E. The Molecular Basis of pH Sensing, Signaling, and Homeostasis in Fungi. In Advances in Applied Microbiology Vol 79; Sariaslani, S., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 1–18. ISBN 9780123943187. [Google Scholar]
- Scervino, J.M.; Mesa, M.P.; Della Mónica, I.; Recchi, M.; Moreno, N.S.; Godeas, A. Soil fungal isolates produce different organic acid patterns involved in phosphate salts solubilization. Biol. Fertil. Soils 2010, 46, 755–763. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, S.H. Abiostress resistance and cellulose degradation abilities of haloalkaliphilic fungi: Applications for saline–alkaline remediation. Extremophiles 2017, 22, 155–164. [Google Scholar] [CrossRef]
- Khan, S.; Ivey, D.M.; Krulwich, T.A. Membrane ultrastructure of alkaliphilic Bacillus species studied by rapid-freeze electron microscopy. J. Bacteriol. 1992, 174, 5123–5126. [Google Scholar] [CrossRef] [Green Version]
- Tilburn, J.; Arst, H.N.; Peñalva, M.A. Regulation of Gene Expression by Ambient pH. In Cellular and Molecular Biology of Filamentous Fungi; Borkovich, K.A., Ebbole, D.J., Eds.; ASM Press: Washington, DC, USA, 2010; pp. 480–487. ISBN 9781683671299. [Google Scholar]
- Li, W.; Mitchell, A.P. Proteolytic Activation of Rim1p, a Positive Regulator of Yeast Sporulation and Invasive Growth. Genetics 1997, 145, 63–77. [Google Scholar] [CrossRef]
- Caddick, M.X.; Brownlee, A.G.; Arst, H.N. Regulation of gene expression by pH of the growth medium in Aspergillus nidulans. Mol. Gen. Genet. 1986, 203, 346–353. [Google Scholar] [CrossRef]
- Arst, H.N.; Bignell, E.; Tilburn, J. Two new genes involved in signalling ambient pH in Aspergillus nidulans. Mol. Gen. Genet. 1994, 245, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Mingot, J.M.; Tilburn, J.; Diez, E.; Bignell, E.; Orejas, M.; Widdick, D.A.; Sarkar, S.; Brown, C.V.; Caddick, M.X.; Espeso, E.A.; et al. Specificity determinants of proteolytic processing of Aspergillus PacC transcription factor are remote from the processing site, and processing occurs in yeast if pH signalling is bypassed. Mol. Cell. Biol. 1999, 19, 1390–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilburn, J.; Sarkar, S.; Widdick, D.A.; Espeso, E.A.; Orejas, M.; Mungroo, J.; Peñalva, M.A.; Arst, H.N. The Aspergillus PacC zinc finger transcription factor mediates regulation of both acid- and alkaline-expressed genes by ambient pH. EMBO J. 1995, 14, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Espeso, E.A.; Arst, H.N. On the Mechanism by which Alkaline pH Prevents Expression of an Acid-Expressed Gene. Mol. Cell. Biol. 2000, 20, 3355–3363. [Google Scholar] [CrossRef]
- Díez, E.; Álvaro, J.; Espeso, E.A.; Rainbow, L.; Suárez, T.; Tilburn, J.; Arst, H.N.; Peñalva, M.Á. Activation of the Aspergillus PacC zinc finger transcription factor requires two proteolytic steps. EMBO J. 2002, 21, 1350–1359. [Google Scholar] [CrossRef] [Green Version]
- Orejas, M.; Espeso, E.A.; Tilburn, J.; Sarkar, S.; Arst, H.N.; Peñalva, M.A. Activation of the Aspergillus PacC transcription factor in response to alkaline ambient pH requires proteolysis of the carboxy-terminal moiety. Genes Dev. 1995, 9, 1622–1632. [Google Scholar] [CrossRef] [Green Version]
- Peñalva, M.A.; Tilburn, J.; Bignell, E.; Arst, H.N. Ambient pH gene regulation in fungi: Making connections. Cell Press 2008, 16, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Martínez, J.; Brown, C.V.; Díez, E.; Tilburn, J.; Arst, H.N.; Peñalva, M.Á.; Espeso, E.A. Overlap of nuclear localisation signal and specific DNA-binding residues within the zinc finger domain of PacC. J. Mol. Biol. 2003, 334, 667–684. [Google Scholar] [CrossRef]
- Mingot, J.M.; Espeso, E.A.; Díez, E.; Peñalva, M. Ambient pH Signaling Regulates Nuclear Localization of the Aspergillus nidulans PacC Transcription Factor. Mol. Cell. Biol. 2001, 21, 1688–1699. [Google Scholar] [CrossRef] [Green Version]
- Espeso, E.A.; Tilburn, J.; Sánchez-Pulido, L.; Brown, C.V.; Valencia, A.; Arst, H.N.; Peñalva, M.A. Specific DNA recognition by the Aspergillus nidulans three zinc finger transcription factor PacC. J. Mol. Biol. 1997, 274, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Mitchell, A.P. Yeast PalA/AIP1/Alix homolog Rim20p associates with a PEST-like region and is required for its proteolytic cleavage. J. Bacteriol. 2001, 183, 6917–6923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futai, E.; Maeda, T.; Sorimachi, H.; Kitamoto, K.; Ishiura, S.; Suzuki, K. The protease activity of a calpain-like cysteine protease in Saccharomyces cerevisiae is required for alkaline adaptation and sporulation. Mol. Gen. Genet. 1999, 260, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Peñalva, M.A.; Arst, H.N. Regulation of Gene Expression by Ambient pH in Filamentous Fungi and Yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 426–446. [Google Scholar] [CrossRef] [Green Version]
- Calcagno-Pizarelli, A.M.; Negrete-Urtasun, S.; Denison, S.H.; Rudnicka, J.D.; Bussink, H.J.; Múnera-Huertas, T.; Stanton, L.; Hervás-Aguilar, A.; Espeso, E.A.; Tilburn, J.; et al. Establishment of the ambient pH signaling complex in Aspergillus nidulans: PalI assists plasma membrane localization of PalH. Eukaryot. Cell 2007, 6, 2365–2375. [Google Scholar] [CrossRef] [Green Version]
- Herranz, S.; Rodriguez, J.M.; Bussink, H.-J.; Sanchez-Ferrero, J.C.; Arst, H.N.; Peñalva, M.A.; Vincent, O. Arrestin-related proteins mediate pH signaling in fungi. Proc. Natl. Acad. Sci. USA 2005, 102, 12141–12146. [Google Scholar] [CrossRef] [Green Version]
- Lefkowitz, R.J.; Shenoy, S.K. Transduction of receptor signals by β-arrestins. Science 2005, 308, 512–517. [Google Scholar] [CrossRef]
- Galindo, A.; Hervás-Aguilar, A.; Rodríguez-Galán, O.; Vincent, O.; Arst, H.N.; Tilburn, J.; Peñalva, M.A. PalC, one of two Bro1 domain proteins in the fungal pH signalling pathway, localizes to cortical structures and binds Vps32. Traffic 2007, 8, 1346–1364. [Google Scholar] [CrossRef] [Green Version]
- Lucena-Agell, D.; Galindo, A.; Arst, H.N.; Peñalva, M.A. Aspergillus nidulans Ambient pH Signaling Does Not Require Endocytosis. Eukaryot. Cell 2015, 14, 545–553. [Google Scholar] [CrossRef] [Green Version]
- Vincent, O.; Rainbow, L.; Tilburn, J.; Arst, H.N.; Peñalva, M.A. YPXL/I Is a Protein Interaction Motif Recognized by Aspergillus PalA and Its Human Homologue, AIP1/Alix. Mol. Cell. Biol. 2003, 23, 1647–1655. [Google Scholar] [CrossRef] [Green Version]
- Denison, S.H.; Orejas, M.; Arst, H.N. Signaling of ambient pH in Aspergillus involves a cysteine protease. J. Biol. Chem. 1995, 270, 28519–28522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conner, S.D.; Schmid, S.L. Regulated portals of enrty into the cell. Nature 2003, 422, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Smith, F.J.; Subaran, R.; Mitchell, A.P. Multivesicular Body-ESCRT Components Function in pH Response Regulation in Saccharomyces cerevisiae and Candida albicans. Mol. Biol. Cell 2004, 15, 5528–5537. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, C.; Buchkovich, N.J.; Stringer, D.K.; Emr, S.D.; Piper, R.C. Cargo ubiquitination is essential for multivesicular body intralumenal vesicle formation. EMBO Rep. 2012, 13, 331–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henne, W.M.; Buchkovich, N.J.; Emr, S.D. The ESCRT Pathway. Dev. Cell 2011, 21, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Nishino, K.; Obara, K.; Kihara, A. The C-terminal cytosolic region of rim21 senses alterations in plasma membrane lipid composition: Insights into sensing mechanisms for plasma membrane lipid asymmetry. J. Biol. Chem. 2015, 290, 30797–30805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grum-Grzhimaylo, A.A.; Falkoski, D.L.; van den Heuvel, J.; Valero-Jiménez, C.A.; Min, B.; Choi, I.G.; Lipzen, A.; Daum, C.G.; Aanen, D.K.; Tsang, A.; et al. The obligate alkalophilic soda-lake fungus Sodiomyces alkalinus has shifted to a protein diet. Mol. Ecol. 2018, 27, 4808–4819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, T.U. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, 523–531. [Google Scholar] [CrossRef]
- Forgac, M. Vacuolar ATPases: Rotary proton pumps in physiology and pathophysiology. Nat. Rev. Mol. Cell Biol. 2007, 8, 917–929. [Google Scholar] [CrossRef]
- Lee, L.K.; Stewart, A.G.; Donohoe, M.; Bernal, R.A.; Stock, D. The structure of the peripheral stalk of Thermus thermophilus H +-ATPase/synthase. Nat. Struct. Mol. Biol. 2010, 17, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Harold, F.M. Conservation and transformation of energy by bacterial membranes. Bacteriol. Rev. 1972, 36, 172–230. [Google Scholar] [CrossRef] [PubMed]
- Krulwich, T.A. Bioenergetics of alkalophilic bacteria. J. Membr. Biol. 1986, 89, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Kaieda, N.; Wakagi, T.; Koyama, N. Presence of Na+-stimulated V-type ATPase in the membrane of a facultatively anaerobic and halophilic alkaliphile. FEMS Microbiol. Lett. 1998, 167, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Pomorski, T.; Holthuis, J.C.M.; Herrmann, A.; Meer, G. Van Tracking down lipid flippases and their biological functions. J. Cell Sci. 2004, 117, 805–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, M.; Kihara, A.; Igarashi, Y. Lipid Asymmetry of the Eukaryotic Plasma Membrane: Functions and Related Enzymes. Biol. Pharm. Bull. 2006, 29, 1542–1546. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Kihara, A.; Denpoh, A.; Igarashi, Y. The Rim101 Pathway Is Involved in Rsb1 Expression Induced by Altered Lipid Asymmetry. Mol. Biol. Cell 2008, 19, 1922–1931. [Google Scholar] [CrossRef] [Green Version]
- Daleke, D.L. Phospholipid Flippases. J. Biol. Chem. 2007, 282, 821–825. [Google Scholar] [CrossRef] [Green Version]
- Sahu, S.K.; Gummadi, S.N.; Manoj, N.; Aradhyam, G.K. Phospholipid scramblases: An overview. Arch. Biochem. Biophys. 2007, 462, 103–114. [Google Scholar] [CrossRef]
- Hankins, H.M.; Baldridge, R.D.; Xu, P.; Graham, T.R. Role of Flippases, Scramblases and Transfer Proteins in Phosphatidylserine Subcellular Distribution. Traffic 2015, 16, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, J.; Colombo, A.C.; Zamith-Miranda, D.; Silva, V.K.A.; Allegood, J.C.; Casadevall, A.; Del Poeta, M.; Nosanchuk, J.D.; Kronstad, J.W.; Rodrigues, M.L. The putative flippase Apt1 is required for intracellular membrane architecture and biosynthesis of polysaccharide and lipids in Cryptococcus neoformans. Biochim. Biophys. Acta 2018, 1865, 532–541. [Google Scholar] [CrossRef]
- Mioka, T.; Fujimura-Kamada, K.; Mizugaki, N.; Kishimoto, T.; Sano, T.; Nunome, H.; Williams, D.E.; Andersen, R.J.; Tanaka, K. Phospholipid flippases and Sfk1p, a novel regulator of phospholipid asymmetry, contribute to low permeability of the plasma membrane. Mol. Biol. Cell 2018, 29, 1203–1218. [Google Scholar] [CrossRef] [PubMed]
- Pomorski, T.; Lombardi, R.; Riezman, H.; Devaux, P.F.; van Meer, G.; Holthuis, J.C.M. Drs2p-related P-type ATPases Dnf1p and Dnf2p Are Required for Phospholipid Translocation across the Yeast Plasma Membrane and Serve a Role in Endocytosis. Mol. Biol. Cell 2003, 14, 1240–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Fujimura-Kamada, K.; Furuta, N.; Kato, U.; Umeda, M.; Tanaka, K. Cdc50p, a Protein Required for Polarized Growth, Associates with the Drs2p P-Type ATPase Implicated in Phospholipid Translocation in Saccharomyces cerevisiae. Mol. Biol. Cell 2004, 15, 3418–3432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, N.; Fujimura-Kamada, K.; Saito, K.; Yamamoto, T.; Tanaka, K. Endocytic Recycling in Yeast Is Regulated by Putative Phospholipid Translocases and the Ypt31p/32p–Rcy1p Pathway. Mol. Biol. Cell 2007, 18, 295–312. [Google Scholar] [CrossRef]
- Prasad, R.; Khandelwal, N.K.; Banerjee, A. Yeast ABC Transporters in Lipid Trafficking. Fungal Genet. Biol. 2016, 93, 25–34. [Google Scholar] [CrossRef]
- Decottignies, A.; Grant, A.M.; Nichols, J.W.; De Wet, H.; McIntosh, D.B.; Goffeau, A. ATPase and multidrug transport activities of the overexpressed yeast ABC protein Yor1p. J. Biol. Chem. 1998, 273, 12612–12622. [Google Scholar] [CrossRef] [Green Version]
- Kihara, A.; Igarashi, Y. Cross Talk between Sphingolipids and Glycerophospholipids in the Establishment of Plasma Membrane Asymmetry. Mol. Biol. Cell 2004, 15, 4949–4959. [Google Scholar] [CrossRef] [Green Version]
- Kihara, A.; Igarashi, Y. Identification and Characterization of a Saccharomyces cerevisiae Gene, RSB1, Involved in Sphingoid Long-chain Base Release. J. Biol. Chem. 2002, 277, 30048–30054. [Google Scholar] [CrossRef] [Green Version]
- Kay, J.G.; Grinstein, S. Phosphatidylserine-Mediated Cellular Signaling. In Lipid-Mediated Protein Signaling; Capelluto, D.G.S., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 991, pp. 177–193. ISBN 978-94-007-6330-2. [Google Scholar]
- Young, B.P.; Shin, J.J.H.; Orij, R.; Chao, J.T.; Li, S.C.; Guan, X.L.; Khong, A.; Jan, E.; Wenk, M.R.; Prinz, W.A.; et al. Phosphatidic Acid Is a pH Biosensor that Links Membrane Biogenesis to Metabolism. Science 2010, 329, 1085–1088. [Google Scholar] [CrossRef]
- Danilova, O.A.; Ianutsevich, E.A.; Bondarenko, S.A.; Georgieva, M.L.; Vikchizhanina, D.A.; Groza, N.V.; Bilanenko, E.N.; Tereshina, V.M. Osmolytes and membrane lipids in the adaptation of micromycete Emericellopsis alkalina to ambient pH and sodium chloride. Fungal Biol. 2020, 124, 884–891. [Google Scholar] [CrossRef]
- Bondarenko, S.A.; Ianutsevich, E.A.; Danilova, O.A.; Grum-Grzhimaylo, A.A.; Kotlova, E.R.; Kamzolkina, O.V.; Bilanenko, E.N.; Tereshina, V.M. Membrane lipids and soluble sugars dynamics of the alkaliphilic fungus Sodiomyces tronii in response to ambient pH. Extremophiles 2017, 21, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, S.A.; Ianutsevich, E.A.; Sinitsyna, N.A.; Georgieva, M.L.; Bilanenko, E.N.; Tereshina, B.M. Dynamics of the Cytosol Soluble Carbohydrates and Membrane Lipids in Response to Ambient pH in Alkaliphilic and Alkalitolerant Fungi. Microbiology 2018, 87, 21–32. [Google Scholar] [CrossRef]
- Jaenicke, R.; Böhm, G. The stability of proteins in extreme environments. Curr. Opin. Struct. Biol. 1998, 8, 738–748. [Google Scholar] [CrossRef]
- Finch, A.; Kim, J. Thermophilic Proteins as Versatile Scaffolds for Protein Engineering. Microorganisms 2018, 6, 97. [Google Scholar] [CrossRef] [Green Version]
- Haki, G.D.; Rakshit, S.K. Developments in industrially important thermostable enzymes: A review. Bioresour. Technol. 2003, 89, 17–34. [Google Scholar] [CrossRef]
- Madern, D.; Ebel, C.; Zaccai, G. Halophilic adaptation of enzymes. Extremophiles 2000, 4, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Plemenitaš, A.; Lenassi, M.; Konte, T.; Kejžar, A.; Zajc, J.; Gostinčar, C.; Gunde-Cimerman, N. Adaptation to high salt concentrations in halotolerant/halophilic fungi: A molecular perspective. Front. Microbiol. 2014, 5, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, N.; Shendye, A.; Rao, M. Molecular and Biotechnological Aspects of Xilanases. FEMS Microbiol. Rev. 1999, 23, 411–456. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.D.; Sidhu, G.; Pot, I.; Brayer, G.D.; Withers, S.G.; McIntosh, L.P. Hydrogen bonding and catalysis: A novel explanation for how a single amino acid substitution can change the pH optimum of a glycosidase. J. Mol. Biol. 2000, 299, 255–279. [Google Scholar] [CrossRef] [Green Version]
- Hakulinen, N.; Turunen, O.; Jänis, J.; Leisola, M.; Rouvinen, J. Three-dimensional structures of thermophilic β-1,4-xylanases from Chaetomium thermophilum and Nonomuraea flexuosa: Comparison of twelve xylanases in relation to their thermal stability. Eur. J. Biochem. 2003, 270, 1399–1412. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Liu, X.; Yu, Q.; Zhang, X.; Qu, Y.; Gao, P.; Wang, T. Directed evolution for engineering pH profile of endoglucanase III from Trichoderma reesei. Biomol. Eng. 2005, 22, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Turunen, O.; Vuorio, M.; Fenel, F.; Leisola, M. Engineering of multiple arginines into the Ser/Thr surface of Trichoderma reesei endo-1,4-β-xylanase II increases the thermotolerance and shifts the pH optimum towards alkaline pH. Protein Eng. 2002, 15, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, K.; Huang, H.; Shi, P.; Yang, P.; Yao, B. Gene cloning, expression, and biochemical characterization of an alkali-tolerant β-mannanase from Humicola insolens Y1. J. Ind. Microbiol. Biotechnol. 2012, 39, 547–555. [Google Scholar] [CrossRef]
- Wohlfahrt, G.; Pellikka, T.; Boer, H.; Teeri, T.T.; Koivula, A. Probing pH-dependent functional elements in proteins: Modification of carboxylic acid pairs in Trichoderma reesei cellobiohydrolase ce16A. Biochemistry 2003, 42, 10095–10103. [Google Scholar] [CrossRef] [PubMed]
- Dalmaso, G.Z.L.; Ferreira, D.; Vermelho, A.B. Marine extremophiles a source of hydrolases for biotechnological applications. Mar. Drugs 2015, 13, 1925–1965. [Google Scholar] [CrossRef] [Green Version]
- Dhakar, K.; Pandey, A. Wide pH range tolerance in extremophiles: Towards understanding an important phenomenon for future biotechnology. Appl. Microbiol. Biotechnol. 2016, 100, 2499–2510. [Google Scholar] [CrossRef]
- Kumar, C.G.; Takagi, H. Microbial alkaline proteases: From a bioindustrial viewpoint. Biotechnol. Adv. 1999, 17, 561–594. [Google Scholar] [CrossRef]
- Never, Z.; Cecilia, C.; Suleimain, M.; Madundo, M. Dehairing of animal hides and skins by alkaline proteases of Aspergillus oryzae for efficient processing to leather products in Tanzania. Afr. J. Biotechnol. 2019, 18, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Malathi, S.; Chakraborty, R. Production of alkaline protease by a new Aspergillus flavus isolate under solid-substrate fermentation conditions for use as a depilation agent. Appl. Environ. Microbiol. 1991, 57, 712–716. [Google Scholar] [CrossRef] [Green Version]
- Hajji, M.; Kanoun, S.; Nasri, M.; Gharsallah, N. Purification and characterization of an alkaline serine-protease produced by a new isolated Aspergillus clavatus ES1. Process Biochem. 2007, 42, 791–797. [Google Scholar] [CrossRef]
- Hajji, M.; Rebai, A.; Gharsallah, N.; Nasri, M. Optimization of alkaline protease production by Aspergillus clavatus ES1 in Mirabilis jalapa tuber powder using statistical experimental design. Appl. Microbiol. Biotechnol. 2008, 79, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Damare, S.; Raghukumar, C.; Muraleedharan, U.D.; Raghukumar, S. Deep-sea fungi as a source of alkaline and cold-tolerant proteases. Enzym. Microb. Technol. 2006, 39, 172–181. [Google Scholar] [CrossRef]
- Tunga, R.; Shrivastava, B.; Banerjee, R. Purification and characterization of a protease from solid state cultures of Aspergillus parasiticus. Process Biochem. 2003, 38, 1553–1558. [Google Scholar] [CrossRef]
- Chellappan, S.; Jasmin, C.; Basheer, S.M.; Elyas, K.K.; Bhat, S.G.; Chandrasekaran, M. Production, purification and partial characterization of a novel protease from marine Engyodontium album BTMFS10 under solid state fermentation. Process Biochem. 2006, 41, 956–961. [Google Scholar] [CrossRef]
- Li, J.; Yang, J.; Huang, X.; Zhang, K.Q. Purification and characterization of an extracellular serine protease from Clonostachys rosea and its potential as a pathogenic factor. Process Biochem. 2006, 41, 925–929. [Google Scholar] [CrossRef]
- Dienes, D.; Börjesson, J.; Hägglund, P.; Tjerneld, F.; Lidén, G.; Réczey, K.; Stålbrand, H. Identification of a trypsin-like serine protease from Trichoderma reesei QM9414. Enzym. Microb. Technol. 2007, 40, 1087–1094. [Google Scholar] [CrossRef]
- Alkin, N.; Dunaevsky, Y.; Elpidina, E.; Beljakova, G.; Tereshchenkova, V.; Filippova, I.; Belozersky, M. Proline-specific fungal peptidases: Genomic analysis and identification of secreted dpp4 in alkaliphilic and alkalitolerant fungi. J. Fungi 2021, 7, 744. [Google Scholar] [CrossRef]
- Hattori, M.; Isomura, S.; Yokoyama, E.; Ujita, M.; Hara, A. Extracellular trypsin-like proteases produced by Cordyceps militaris. J. Biosci. Bioeng. 2005, 100, 631–636. [Google Scholar] [CrossRef]
- Dutta, T.; Sengupta, R.; Sahoo, R.; Sinha Ray, S.; Bhattacharjee, A.; Ghosh, S. A novel cellulase free alkaliphilic xylanase from alkali tolerant Penicillium citrinum: Production, purification and characterization. Lett. Appl. Microbiol. 2007, 44, 206–211. [Google Scholar] [CrossRef]
- Guan, G.Q.; Zhao, P.X.; Zhao, J.; Wang, M.J.; Huo, S.H.; Cui, F.J.; Jiang, J.X. Production and Partial Characterization of an Alkaline Xylanase from a Novel Fungus Cladosporium oxysporum. Biomed Res. Int. 2016, 2016, 4575024. [Google Scholar] [CrossRef] [Green Version]
- Chandra Raj, K.; Chandra, T.S. Purification and characterization of xylanase from alkali-tolerant Aspergillus fischeri Fxn1. FEMS Microbiol. Lett. 1996, 145, 457–461. [Google Scholar] [CrossRef]
- Bajaj, B.K.; Abbass, M. Studies on an alkali-thermostable xylanase from Aspergillus fumigatus MA28. 3 Biotech 2011, 1, 161–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjan, K.; Lone, M.A.; Sahay, S. Detergent compatible cold-active alkaline amylases from Clavispora lusitaniae CB13. J. Microbiol. Biotechnol. Food Sci. 2016, 5, 306–310. [Google Scholar] [CrossRef] [Green Version]
- Zaferanloo, B.; Bhattacharjee, S.; Ghorbani, M.M.; Mahon, P.J.; Palombo, E.A. Amylase production by Preussia minima, a fungus of endophytic origin: Optimization of fermentation conditions and analysis of fungal secretome by LC-MS. BMC Microbiol. 2014, 14, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.H.; Wang, H.R.; Yan, T.R. Cloning, purification, and characterization of a heat- and alkaline-stable endoglucanase B from Aspergillus niger BCRC31494. Molecules 2012, 17, 9774–9789. [Google Scholar] [CrossRef] [Green Version]
- Schülein, M. Enzymatic properties of cellulases from Humicola insolens. J. Biotechnol. 1997, 57, 71–81. [Google Scholar] [CrossRef]
- Sulistyaningdyah, W.T.; Ogawa, J.; Tanaka, H.; Maeda, C.; Shimizu, S. Characterization of alkaliphilic laccase activity in the culture supernatant of Myrothecium verrucaria 24G-4 in comparison with bilirubin oxidase. FEMS Microbiol. Lett. 2004, 230, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ma, R.; Li, S.; Gong, M.; Yao, B.; Bai, Y.; Gu, J. An alkaline and surfactant-tolerant lipase from Trichoderma lentiforme ACCC30425 with high application potential in the detergent industry. AMB Express 2018, 8, 95. [Google Scholar] [CrossRef]
- Saxena, R.K.; Davidson, W.S.; Sheoran, A.; Giri, B. Purification and characterization of an alkaline thermostable lipase from Aspergillus carneus. Process Biochem. 2003, 39, 239–247. [Google Scholar] [CrossRef]
- Venkatesagowda, B.; Ponugupaty, E.; Barbosa-Dekker, A.M.; Dekker, R.F.H. The Purification and Characterization of Lipases from Lasiodiplodia theobromae, and Their Immobilization and Use for Biodiesel Production from Coconut Oil. Appl. Biochem. Biotechnol. 2018, 185, 619–640. [Google Scholar] [CrossRef]
- Hirayama, S.; Taira, R.; Borch, K.; Sandal, T.; Halkier, T.; Oxenboll, K.M.; Nielsen, B.R. Alkaline Lipolytic Enzyme. U.S. Patent 6,350,604, 28 February 2002. [Google Scholar]
- Mamo, G.; Mattiasson, B. Alkaliphilic Microorganisms in Biotechnology. In Biotechnology of Extremophiles Advances and Challenges; Rampelotto, P.H., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 243–272. ISBN 978-3-319-13520-5. [Google Scholar]
- Lübeck, M.; Lübeck, P.S. Fungal Cell Factories for Efficient and Sustainable Production of Proteins and Peptides. Microorganisms 2022, 10, 753. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, M.; Ray, R.R. Current Trends in Research and Application of Microbial Cellulases. Res. J. Microbiol. 2011, 6, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Cavaco-Paulo, A.; Morgado, J.; Almeida, L.; Kilburn, D. Indigo Backstaining During Cellulase Washing. Text. Res. J. 1998, 68, 398–401. [Google Scholar] [CrossRef] [Green Version]
- Ibarra, D.; Monte, M.C.; Blanco, A.; Martínez, A.T.; Martínez, M.J. Enzymatic deinking of secondary fibers: Cellulases/hemicellulases versus laccase-mediator system. J. Ind. Microbiol. Biotechnol. 2012, 39, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pokhrel, D.; Viraraghavan, T. Treatment of pulp and paper mill wastewater—A review. Sci. Total Environ. 2004, 333, 37–58. [Google Scholar] [CrossRef]
- De Aquino, F.X.; De Sousa, M.V.; Puls, J.; Da Silva, F.G.; Ferreira Filho, E.X. Purification and characterization of a low-molecular-weight xylanase produced by Acrophialophora nainiana. Curr. Microbiol. 1999, 38, 18–21. [Google Scholar] [CrossRef]
- Karlen, D.L.; Andrews, S.S.; Wienhold, B.J.; Zobeck, T.M. Soil Quality Assessment: Past, Present and Future. J. Integr. Biosci. 2014, 6, 3–14. [Google Scholar]
- Rietz, D.N.; Haynes, R.J. Effects of irrigation-induced salinity and sodicity on soil microbial activity. Soil Biol. Biochem. 2003, 35, 845–854. [Google Scholar] [CrossRef]
- Serrano, R.; Gaxiola, R. Microbial Models and Salt Stress Tolerance in Plants. CRC. Crit. Rev. Plant Sci. 1994, 13, 121–138. [Google Scholar] [CrossRef]
- Liu, X.; Liu, J.; Wei, Y.; Tian, Y.; Fan, F.; Pan, H.; Zhang, S.H. Isolation, identification and biologic characteristics of an extreme halotolerant Aspergillus sp. J. Jilin Univ. 2011, 49, 548–552. [Google Scholar]
- Shi, Y.; Zhu, J. Saline–Alkali Soil Improvement Fertilizer and Preparation Method and Use Method Thereof. Chinese Patent CN 105237293 A, 13 January 2016. [Google Scholar]
- Jain, D.; Katyal, P. Optimization of gluco-amylase production from Aspergillus spp. for its use in saccharification of liquefied corn starch. 3 Biotech 2018, 8, 101. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Guilfoyle, E.; Stinson, J.; Skogerson, L. Sugar-Producing and Texture-Improving Bakery Methods and Products Formed Therefrom. United States Patent Application Publication US2018/0242598A1, 30 August 2018. [Google Scholar]
- Mojsov, K.; Andronikov, D.; Janevski, A.; Jordeva, S.; Kertakova, M.; Golomeova, S.; Gaber, S.; Ignjatov, I. Production and application of α-amylase enzyme in textile industry. Tekst. Ind. 2018, 66, 23–28. [Google Scholar]
- Bartelme, M.; Marquardt, J.; Man, V.F.; Leafblad, B. Fast Dissolving Solid Detergent. United States Patent US 10005986, 26 June 2018. [Google Scholar]
- Singh, S.; Singh, S.; Bali, V.; Sharma, L.; Mangla, J. Production of Fungal Amylases Using Cheap, Readily Available Agriresidues, for Potential Application in Textile Industry. Biomed Res. Int. 2014, 2014, 215748. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Chisti, Y.; Chand, U. Production, purification, characterization, and applications of lipases. Biotechnol. Adv. 2001, 19, 627–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verger, R. “Interfacial activation” of lipases: Facts and artifacts. Trends Biotechnol. 1997, 15, 32–38. [Google Scholar] [CrossRef]
- Azeem, F.; Afzal, M.; Javed, S.; Riaz, M.; Kouser, A.; Hussain, S.; Siddique, M.H.; Rasul, I.; Nadeem, H. Bacterial lipases: A review on purification and characterization. Prog. Biophys. Mol. Biol. 2018, 132, 23–34. [Google Scholar] [CrossRef]
- Villeneuve, P. Plant lipases and their applications in oils and fats modification. Eur. J. Lipid Sci. Technol. 2003, 105, 308–317. [Google Scholar] [CrossRef]
- Casas-Godoy, L.; Gasteazoro, F.; Duquesne, S.; Bordes, F.; Marty, A.; Sandoval, G. Lipases: An Overview. In Lipases and Phospholipases: Methods and Protocols, Methods in Molecular Biology; Sandoval, G., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1835, pp. 3–38. ISBN 9781493986729. [Google Scholar]
- Su, F.; Peng, C.; Li, G.L.; Xu, L.; Yan, Y.J. Biodiesel production from woody oil catalyzed by Candida rugosa lipase in ionic liquid. Renew. Energy 2016, 90, 329–335. [Google Scholar] [CrossRef]
- Rogozhin, E.A.; Sadykova, V.S.; Baranova, A.A.; Vasilchenko, A.S.; Lushpa, V.A.; Mineev, K.S.; Georgieva, M.L.; Kul’ko, A.B.; Krasheninnikov, M.E.; Lyundup, A.V.; et al. A novel lipopeptaibol Emericellipsin A with antimicrobial and antitumor activity produced by the extremophilic fungus Emericellopsis alkalina. Molecules 2018, 23, 2785. [Google Scholar] [CrossRef] [Green Version]
- Kuvarina, A.E.; Gavryushina, I.A.; Kulko, A.B.; Ivanov, I.A.; Rogozhin, E.A.; Georgieva, M.L.; Sadykova, V.S. The Emericellipsins A-E from an alkalophilic fungus Emericellopsis alkalina show potent activity against multidrug-resistant pathogenic fungi. J. Fungi 2021, 7, 153. [Google Scholar] [CrossRef]
- Kuvarina, A.E.; Georgieva, M.L.; Rogozhin, E.A.; Kulko, A.B.; Gavryushina, I.A.; Sadykova, V.S. Antimicrobial Potential of the Alkalophilic Fungus Sodiomyces alkalinus and Selection of Strains–Producers of New Antimicotic Compound. Appl. Biochem. Microbiol. 2021, 57, 86–93. [Google Scholar] [CrossRef]
- Kuvarina, A.E.; Rogozhin, E.A.; Sykonnikov, M.A.; Timofeeva, A.V.; Serebryakova, M.V.; Fedorova, N.V.; Kokaeva, L.Y.; Efimenko, T.A.; Georgieva, M.L.; Sadykova, V.S. Isolation and Characterization of a Novel Hydrophobin, Sa-HFB1, with Antifungal Activity from an Alkaliphilic Fungus, Sodiomyces alkalinus. J. Fungi 2022, 8, 659. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. AntiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista-García, R.A.; Balcázar-López, E.; Miranda-Miranda, E.; Sánchez-Reyes, A.; Cuervo-Soto, L.; Aceves-Zamudio, D.; Atriztán-Hernández, K.; Morales-Herrera, C.; Rodríguez-Hernández, R.; Folch-Mallol, J. Characterization of lignocellulolytic activities from a moderate halophile strain of Aspergillus caesiellus isolated from a sugarcane bagasse fermentation. PLoS ONE 2014, 9, e0105893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Abradelo, D.; Pérez-Llano, Y.; Peidro-Guzmán, H.; Sánchez-Carbente, M.d.R.; Folch-Mallol, J.L.; Aranda, E.; Vaidyanathan, V.K.; Cabana, H.; Gunde-Cimerman, N.; Batista-García, R.A. First demonstration that ascomycetous halophilic fungi (Aspergillus sydowii and Aspergillus destruens) are useful in xenobiotic mycoremediation under high salinity conditions. Bioresour. Technol. 2019, 279, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Holliday, R. The genetics of Ustilago maydis. Genet. Res. 1961, 2, 204–230. [Google Scholar] [CrossRef]
- Cao, X.; An, T.; Fu, W.; Zhang, J.; Zhao, H.; Li, D.; Jin, X.; Liu, B. Genome-Wide Identification of Cellular Pathways and Key Genes that Respond to Sodium Bicarbonate Stress in Saccharomyces cerevisiae. Front. Microbiol. 2022, 13, 831973. [Google Scholar] [CrossRef]
- Jiménez-Gómez, I.; Valdés-Muñoz, G.; Moreno-Ulloa, A.; Pérez-Llano, Y.; Moreno-Perlín, T.; Silva-Jiménez, H.; Barreto-Curiel, F.; Sánchez-Carbente, M.d.R.; Folch-Mallol, J.L.; Gunde-Cimerman, N.; et al. Surviving in the Brine: A Multi-Omics Approach for Understanding the Physiology of the Halophile Fungus Aspergillus sydowii at Saturated NaCl Concentration. Front. Microbiol. 2022, 13, 840408. [Google Scholar] [CrossRef]
Figure 1.
Distribution of alkaline environments around the world (colored circles). Fungi species isolated from alkaline ecosystems are shown. Species nomenclature was updated according to the Mycobank Database. The species name given in the cited article is enclosed in brackets. The color of the circles and the frame that encloses the names of the species indicate the average pH of the isolation sites. The following references were used for the figure: Kulunda steppe [5,18,19]; Aral Lake [5,19]; Ararat Valley [5,20]; Trans-Baikal [5,19,20,21]; Kunkur steppe, Choibalsan area, and Bayan-Zang [5,18]; Ulan-Nur Lake [5]; Barun-Undzy Lake and Natron Lake [18]; Iwate [22]; Jayapura [23]; Lonar Lake [24]; Magadi Lake [5,25,26]; Mono Lake [17]; 20 Km SE from Magdalena City [27]. Studies [5,18,19,24,25,26] used morphophysiological and molecular genetic characterization as taxonomic criteria, while studies [17,20,21,22,23,27] used only morphophysiological characterization for fungal identification.
Figure 1.
Distribution of alkaline environments around the world (colored circles). Fungi species isolated from alkaline ecosystems are shown. Species nomenclature was updated according to the Mycobank Database. The species name given in the cited article is enclosed in brackets. The color of the circles and the frame that encloses the names of the species indicate the average pH of the isolation sites. The following references were used for the figure: Kulunda steppe [5,18,19]; Aral Lake [5,19]; Ararat Valley [5,20]; Trans-Baikal [5,19,20,21]; Kunkur steppe, Choibalsan area, and Bayan-Zang [5,18]; Ulan-Nur Lake [5]; Barun-Undzy Lake and Natron Lake [18]; Iwate [22]; Jayapura [23]; Lonar Lake [24]; Magadi Lake [5,25,26]; Mono Lake [17]; 20 Km SE from Magdalena City [27]. Studies [5,18,19,24,25,26] used morphophysiological and molecular genetic characterization as taxonomic criteria, while studies [17,20,21,22,23,27] used only morphophysiological characterization for fungal identification.
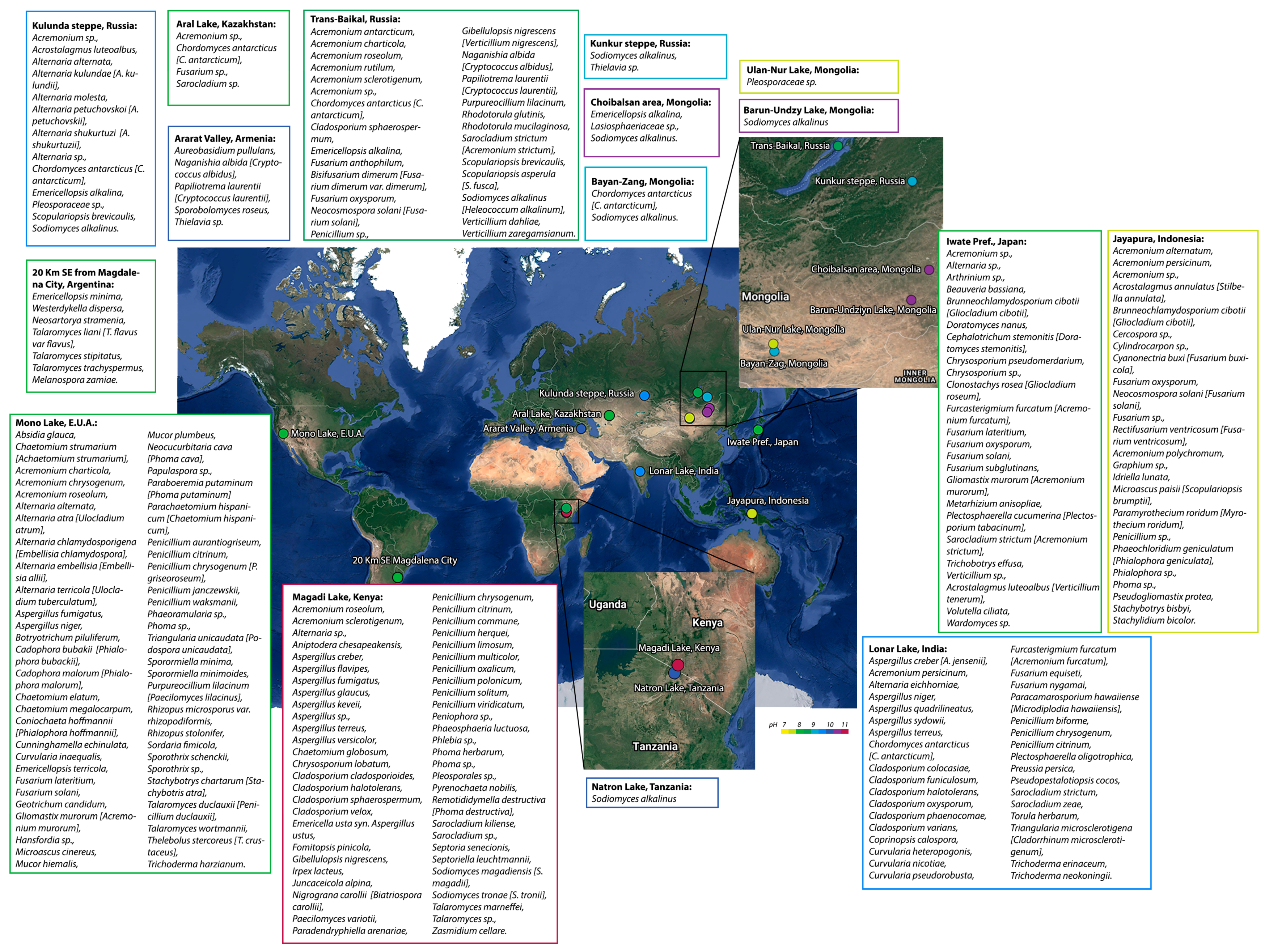
Figure 2.
Relative Family abundance of Ascomycota for the four metagenomes: Büdös-szék pan (MG-RAST ID: mgm4555202.3), Pantanal (MG-RAST ID: mgm4875736.3), Lonar Lake (MG-RAST ID: mgm4863970.3), and Mono Lake (MG-RAST ID: mgm4962892.3). Abundance analysis was performed in MG-RAST (www.mg-rast.org (accessed on 7 May 2023)) with the default values that are in the program. Abundance values are in the Supplementary Material File S1.
Figure 2.
Relative Family abundance of Ascomycota for the four metagenomes: Büdös-szék pan (MG-RAST ID: mgm4555202.3), Pantanal (MG-RAST ID: mgm4875736.3), Lonar Lake (MG-RAST ID: mgm4863970.3), and Mono Lake (MG-RAST ID: mgm4962892.3). Abundance analysis was performed in MG-RAST (www.mg-rast.org (accessed on 7 May 2023)) with the default values that are in the program. Abundance values are in the Supplementary Material File S1.
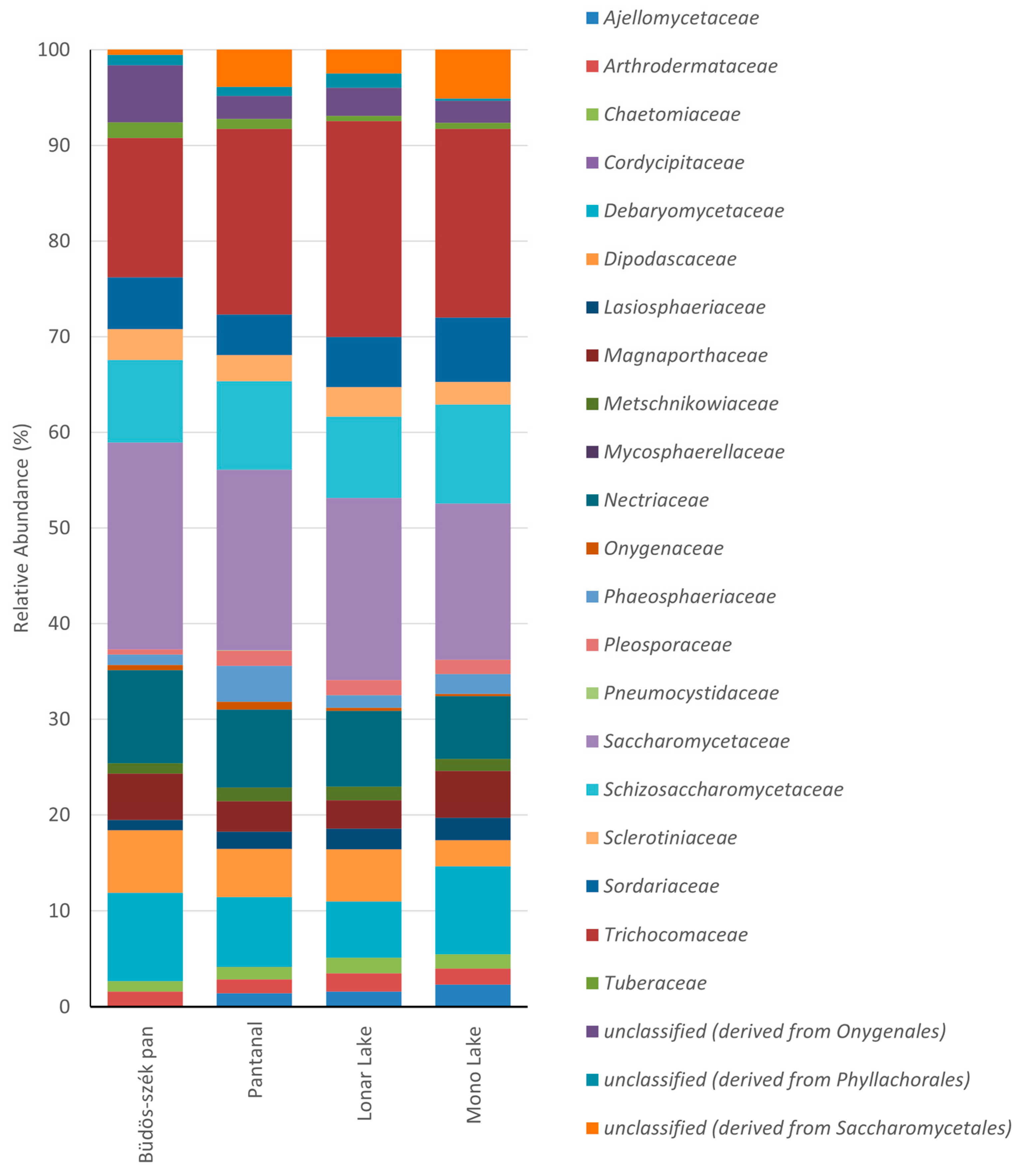
Figure 3.
Signal transduction pathway for the activation of the Rim101/PacC transcription factor. The figure was created in Biorender.com (accessed on 7 May 2023).
Figure 3.
Signal transduction pathway for the activation of the Rim101/PacC transcription factor. The figure was created in Biorender.com (accessed on 7 May 2023).
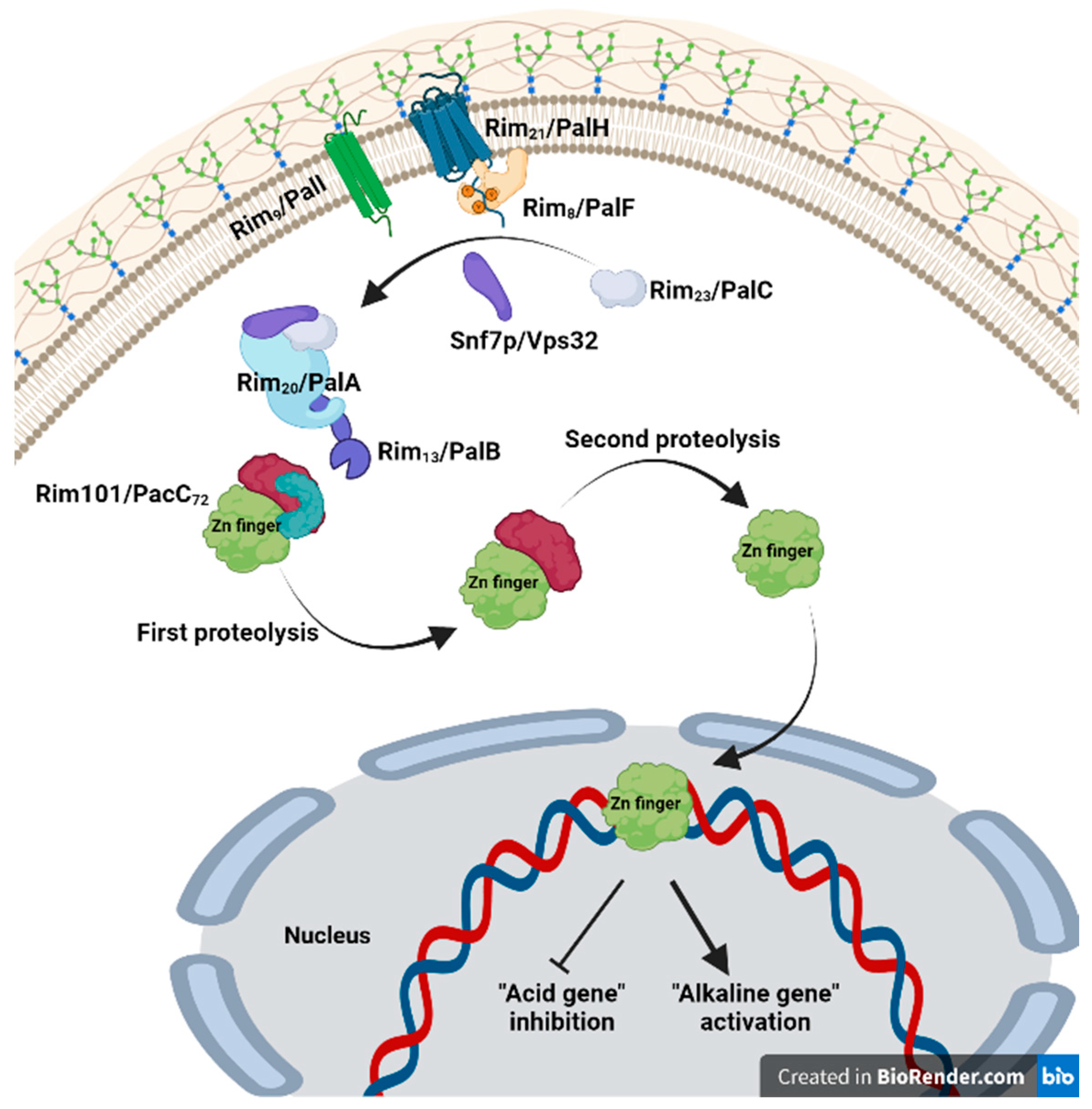
Figure 4.
Effect of pH on the asymmetry of the cytoplasmic membrane of Saccharomyces cerevisiae and signaling mechanisms to conserve lipid distribution in the membrane. The image was designed taking into account the evidence obtained in references [73,84]. The figure was created in Biorender.com (accessed on 7 May 2023).
Figure 4.
Effect of pH on the asymmetry of the cytoplasmic membrane of Saccharomyces cerevisiae and signaling mechanisms to conserve lipid distribution in the membrane. The image was designed taking into account the evidence obtained in references [73,84]. The figure was created in Biorender.com (accessed on 7 May 2023).
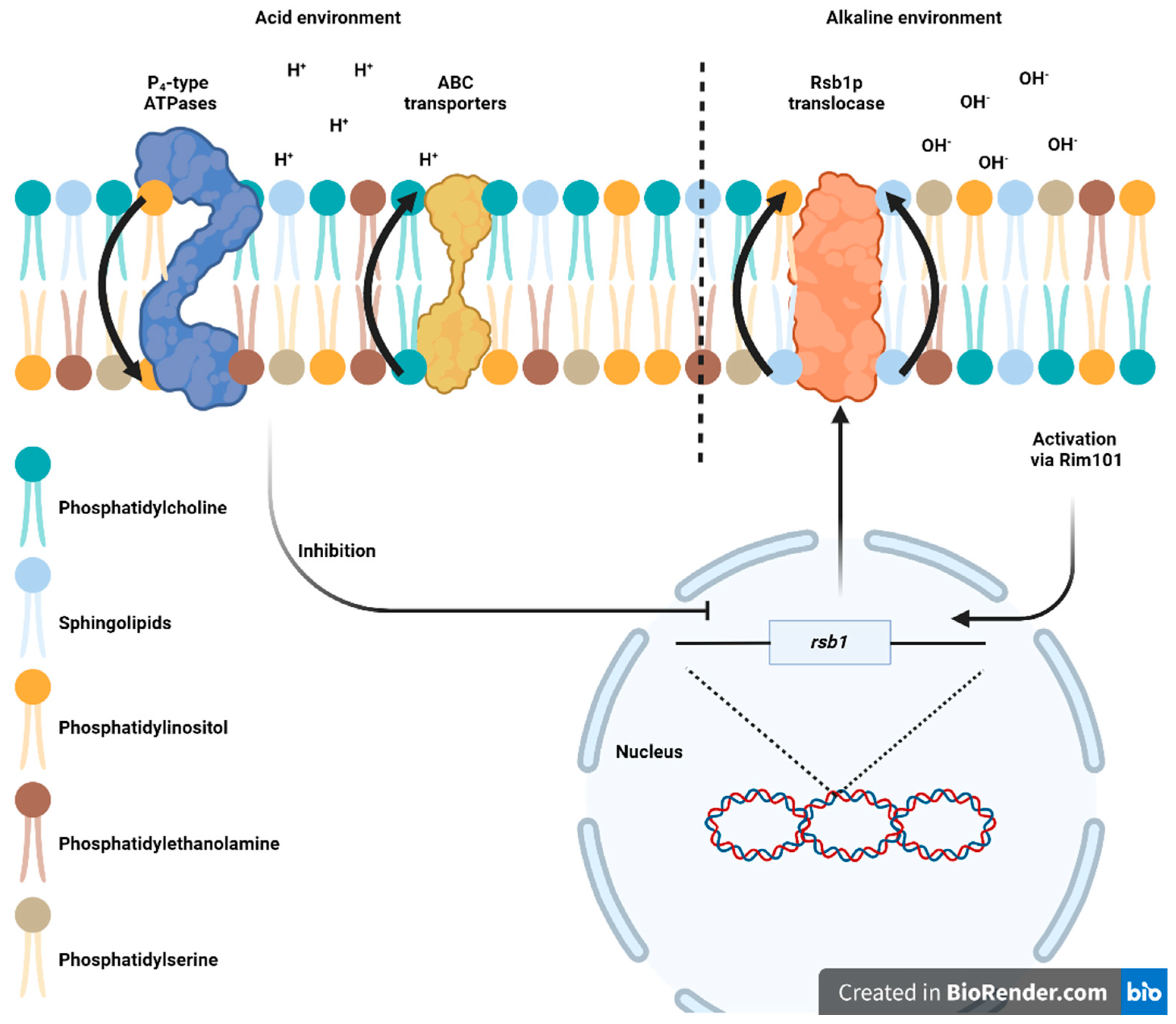
Figure 5.
Effect of pH on the growth rate (mm/d) of Aspergillus sydowii in mineral medium with glucose (2%) and different concentrations of NaCl.
Figure 5.
Effect of pH on the growth rate (mm/d) of Aspergillus sydowii in mineral medium with glucose (2%) and different concentrations of NaCl.
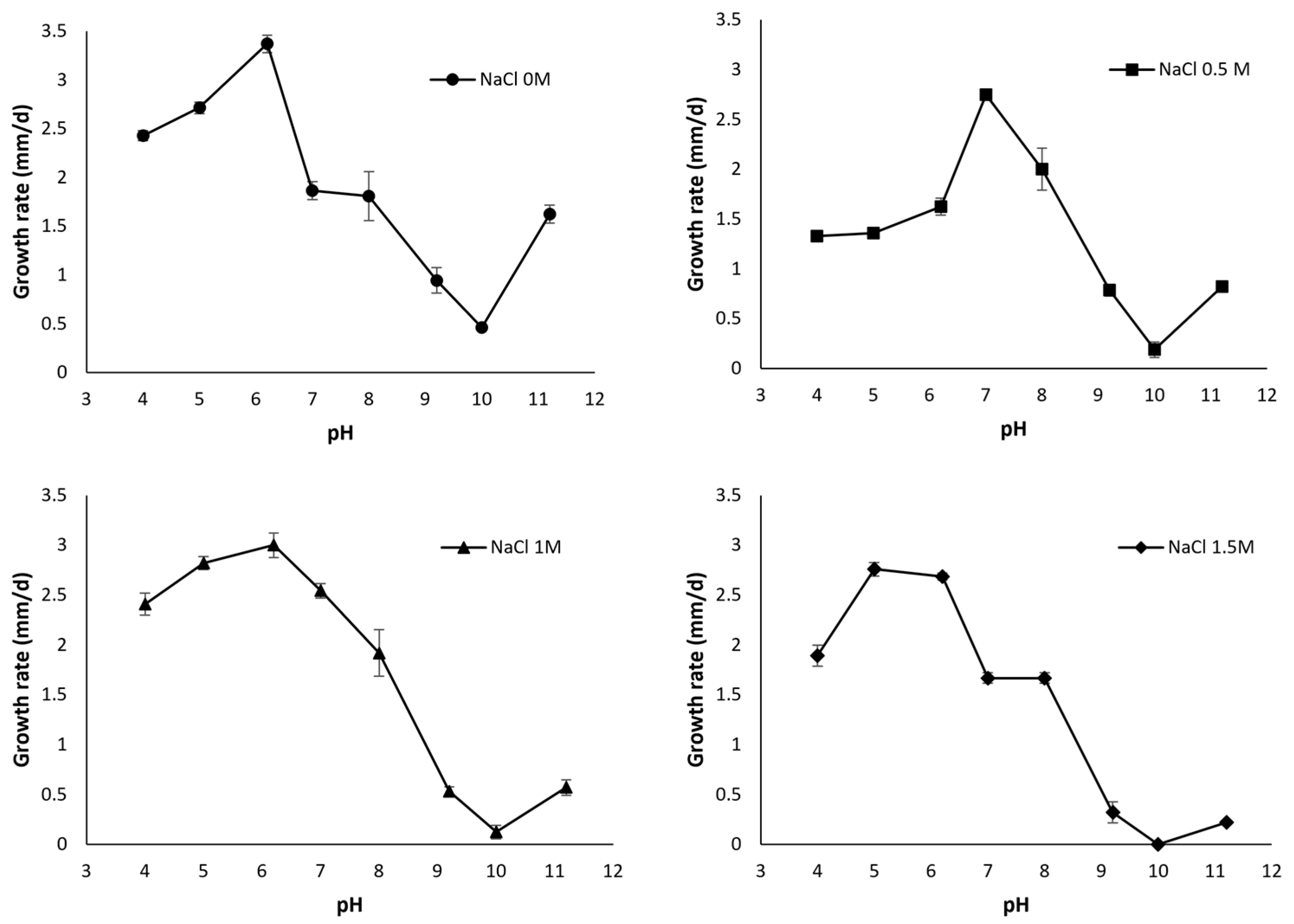
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fernández-López, M.G.; Batista-García, R.A.; Aréchiga-Carvajal, E.T. Alkaliphilic/Alkali-Tolerant Fungi: Molecular, Biochemical, and Biotechnological Aspects. J. Fungi 2023, 9, 652. https://doi.org/10.3390/jof9060652
AMA Style
Fernández-López MG, Batista-García RA, Aréchiga-Carvajal ET. Alkaliphilic/Alkali-Tolerant Fungi: Molecular, Biochemical, and Biotechnological Aspects. Journal of Fungi. 2023; 9(6):652. https://doi.org/10.3390/jof9060652
Chicago/Turabian StyleFernández-López, Maikel Gilberto, Ramón Alberto Batista-García, and Elva Teresa Aréchiga-Carvajal. 2023. "Alkaliphilic/Alkali-Tolerant Fungi: Molecular, Biochemical, and Biotechnological Aspects" Journal of Fungi 9, no. 6: 652. https://doi.org/10.3390/jof9060652
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.