
Reactivation of Coccidioidomycosis in a Mouse Model of Asymptomatic Controlled Disease
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Arthroconidia
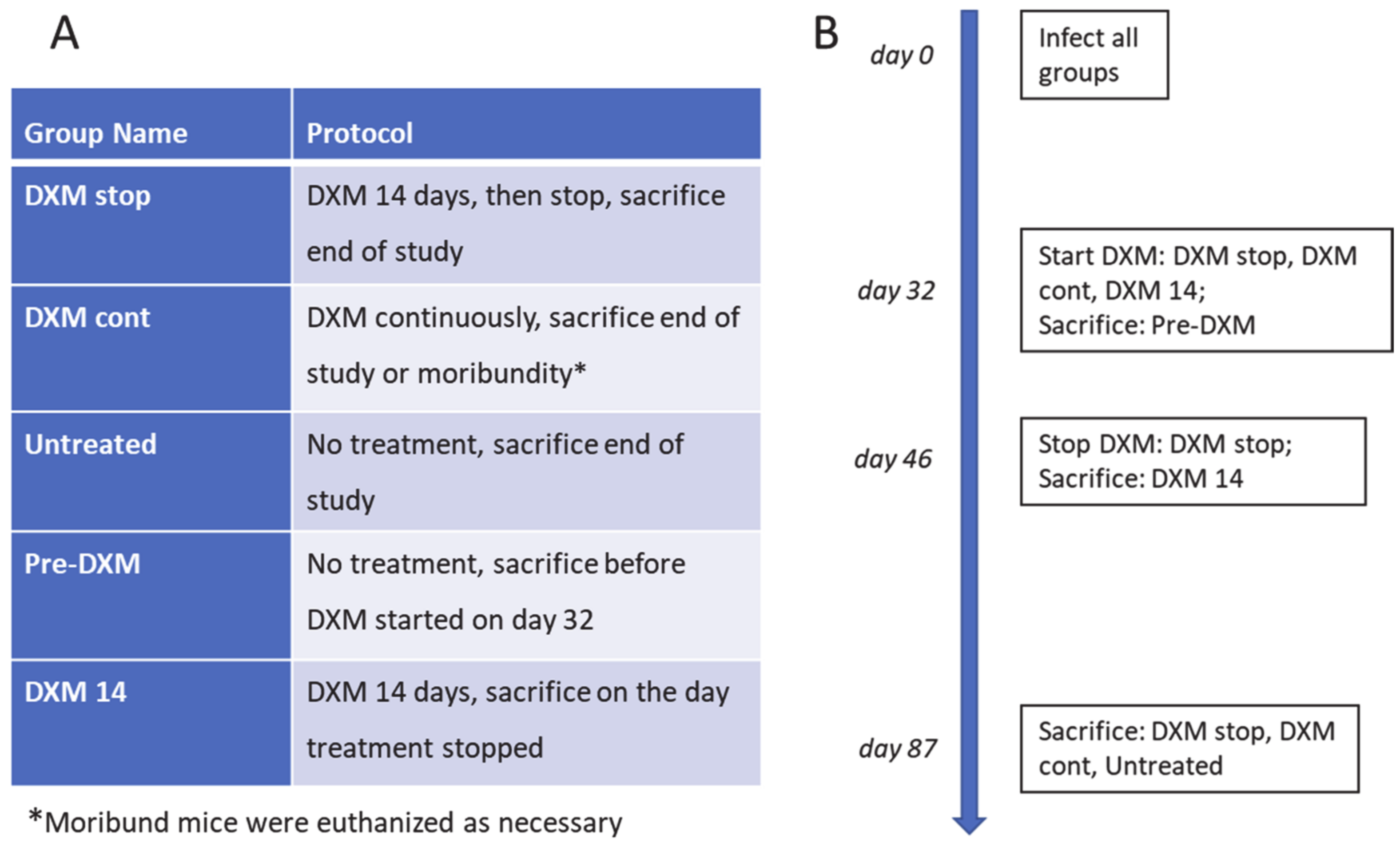
2.3. Mouse Infection and Immunosuppression
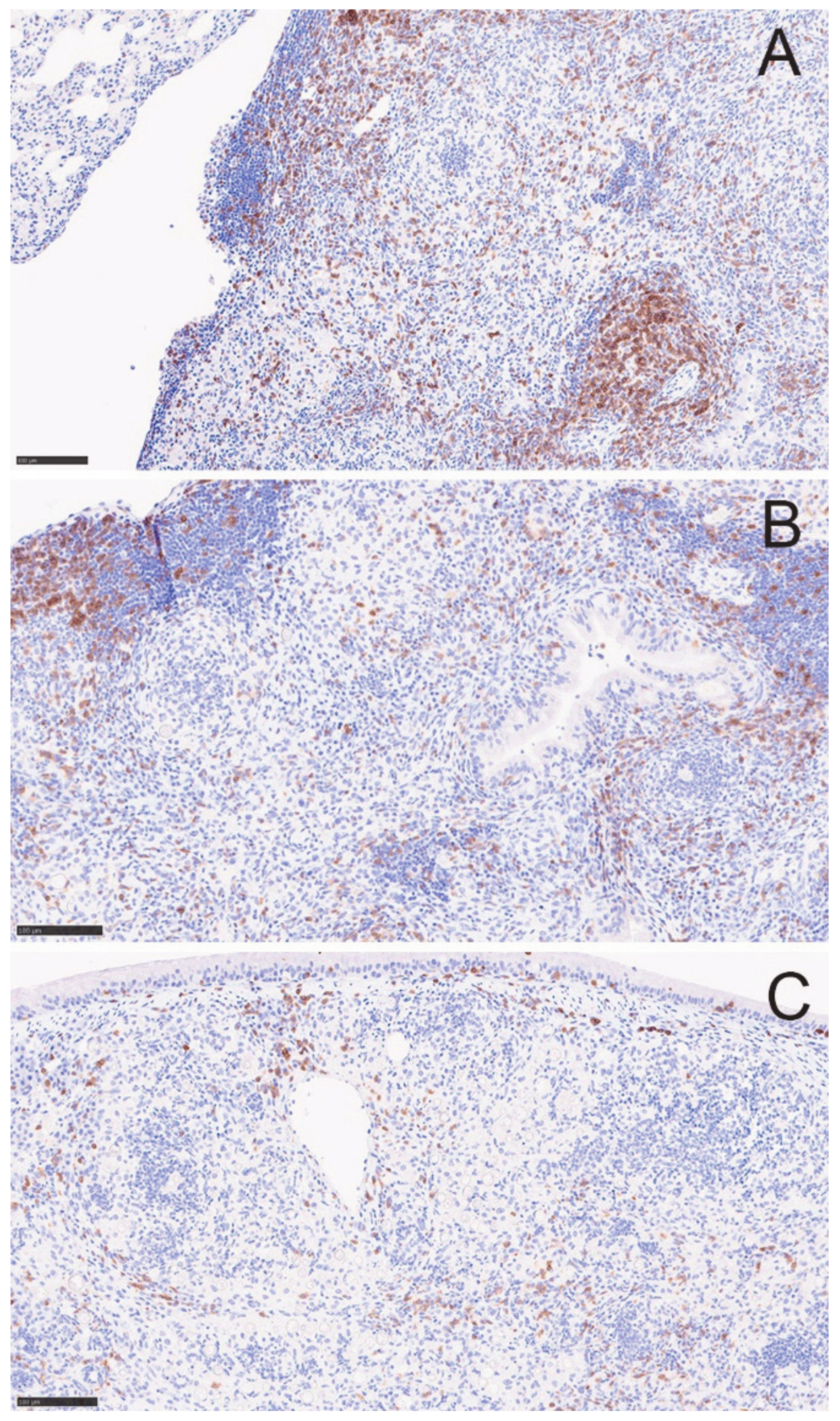
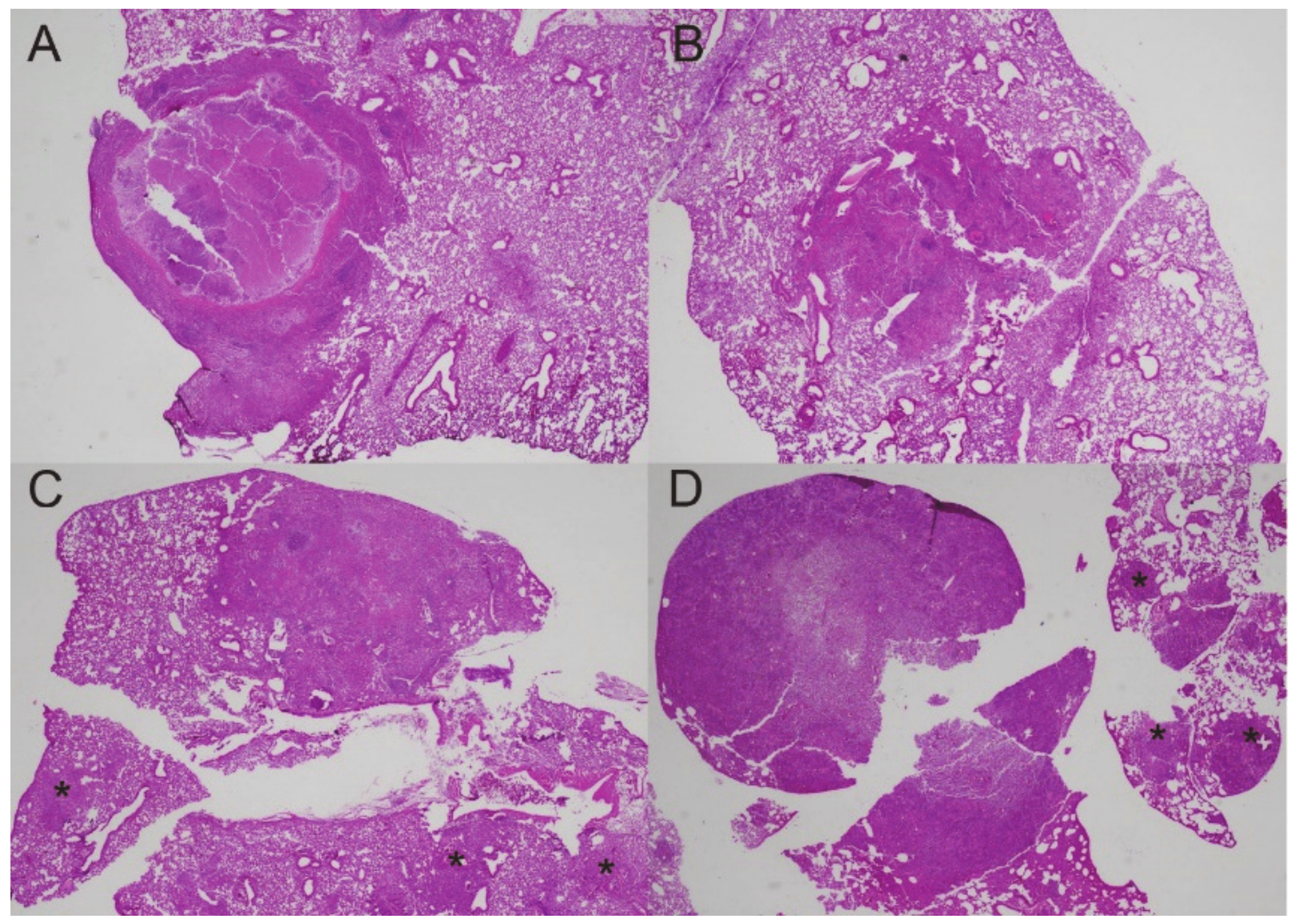
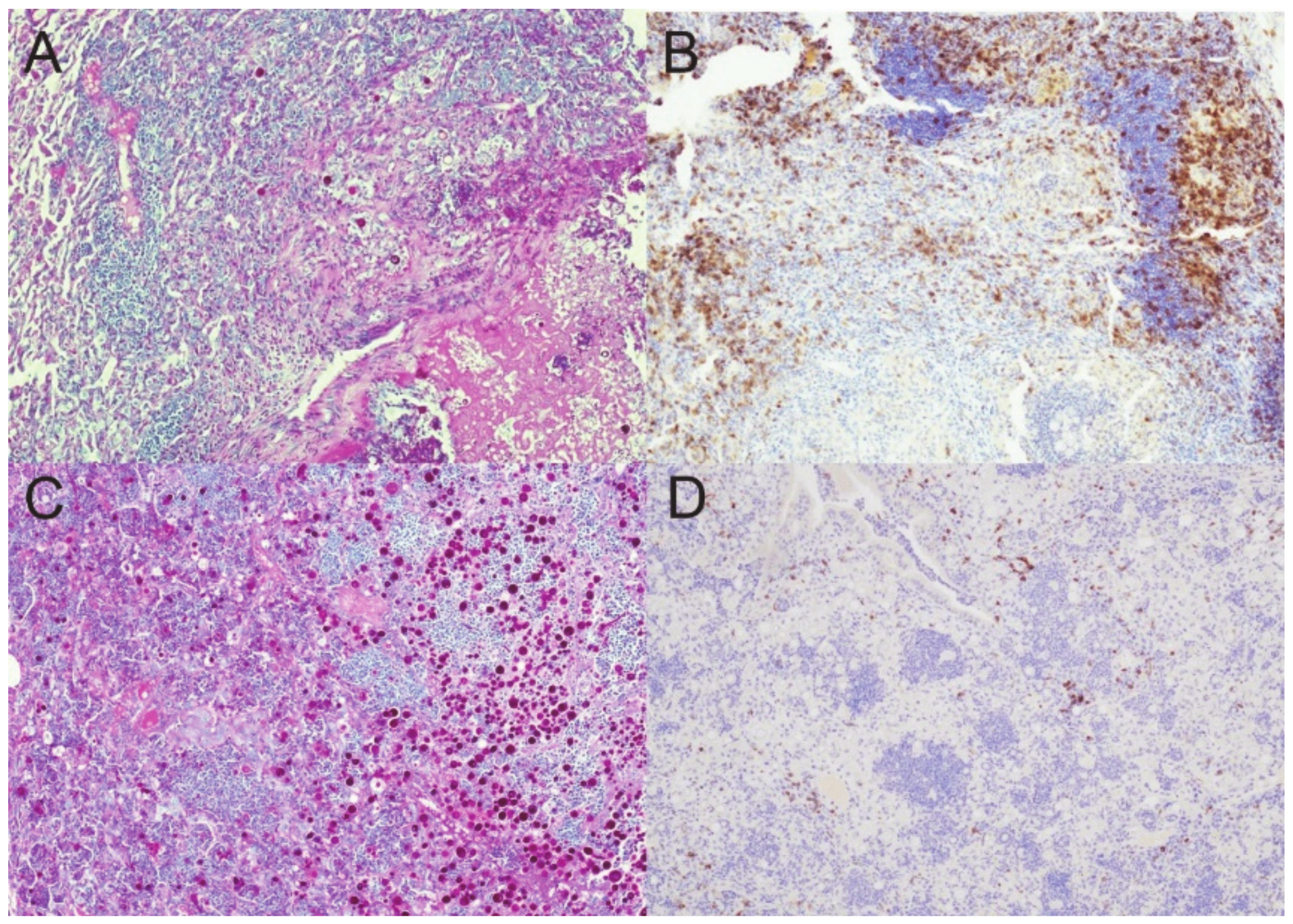
2.4. Histopathology
2.5. Organ Fungal Burden
2.6. Flow Cytometric Characterization of Granulomas
2.7. Statistical Analysis
3. Results
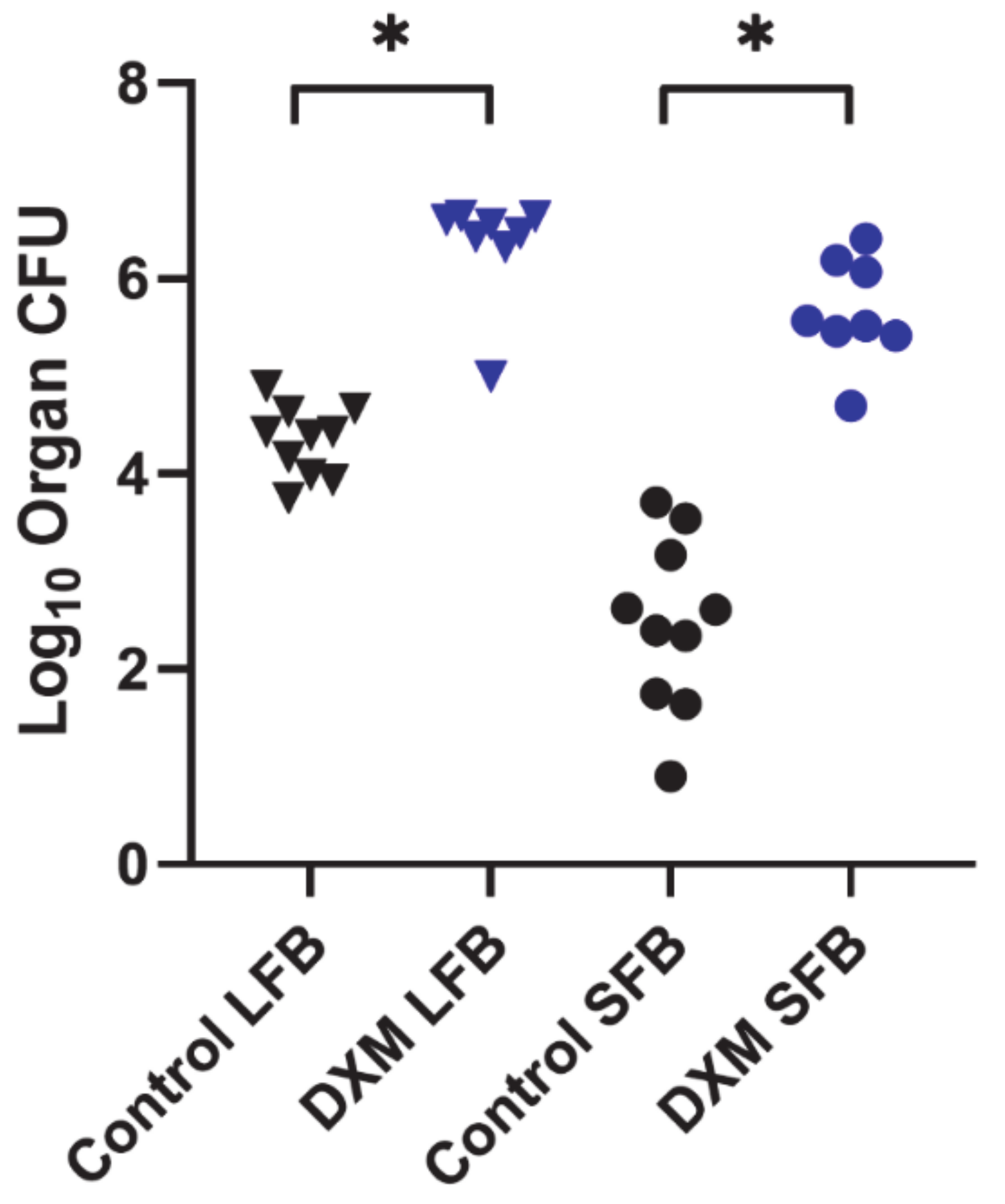
3.1. Fungal Growth, Dissemination, and Mouse Mortality Occur following Administration of DXM
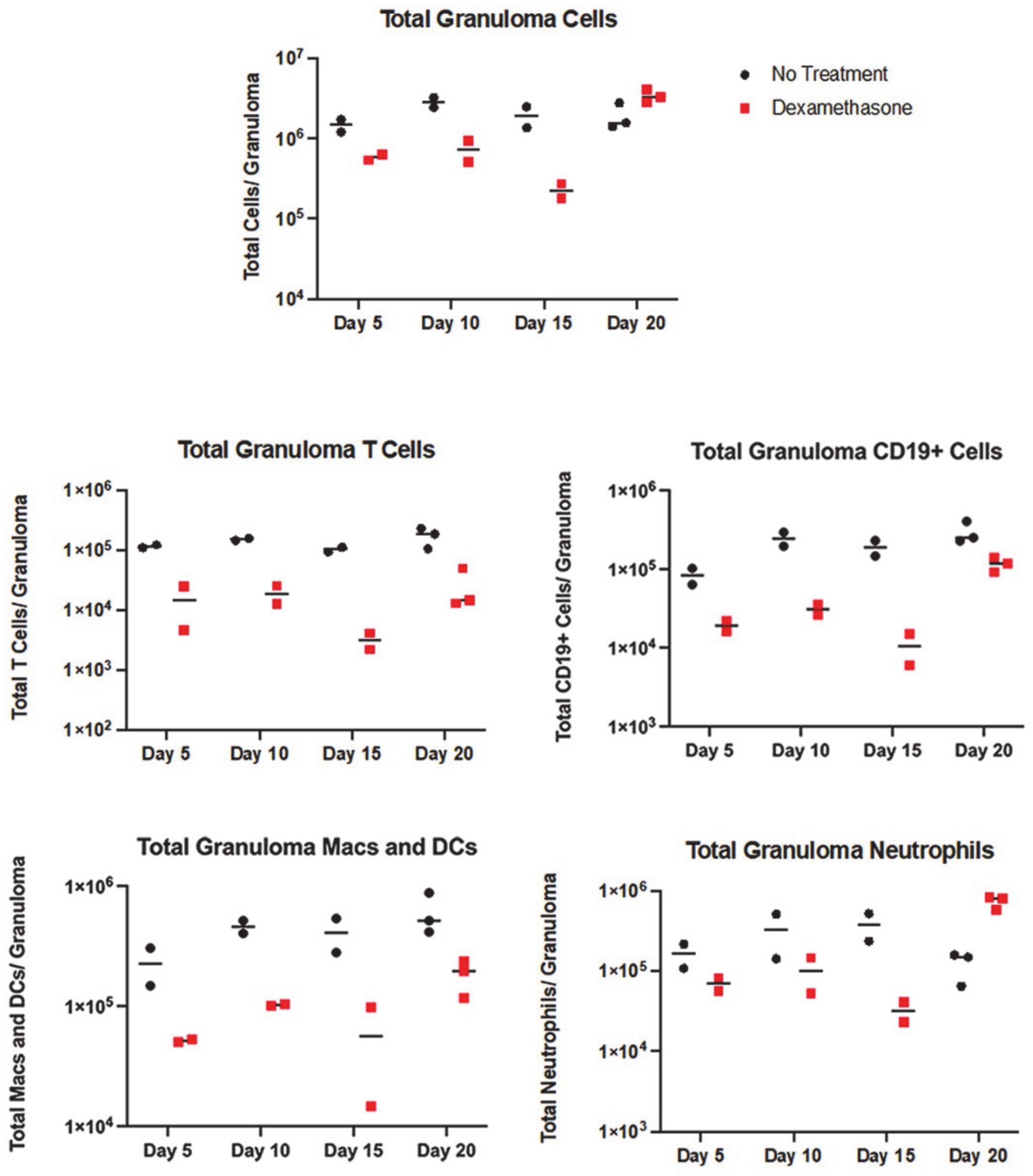
3.2. All Immune Cell Populations Decreased following DXM Treatment
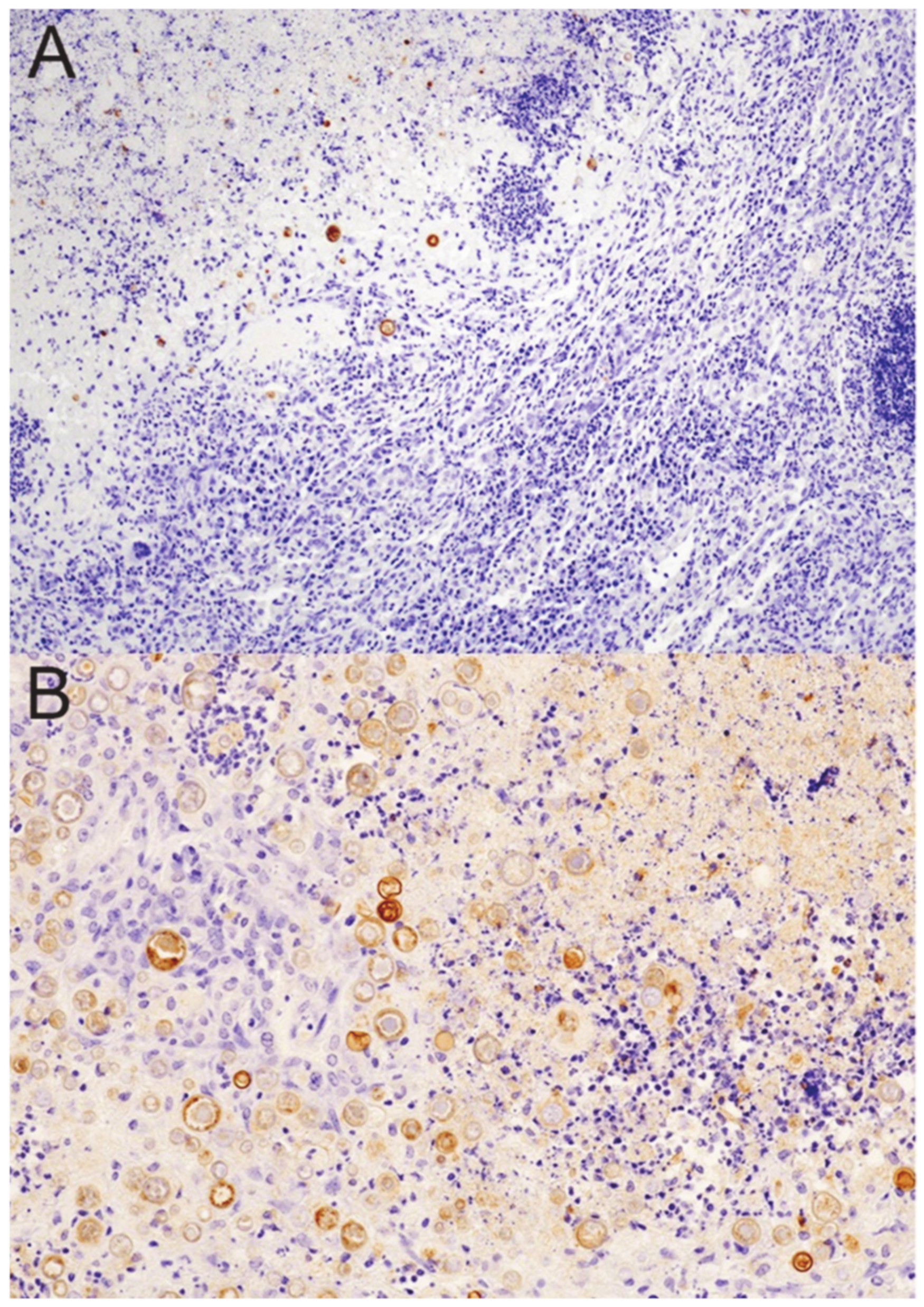
3.3. Granulomas Break down with DXM Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donovan, F.M.; Shubitz, L.; Powell, D.; Orbach, M.; Frelinger, J.; Galgiani, J.N. Early Events in Coccidioidomycosis. Clin. Microbiol. Rev. 2019, 33, e112-19. [Google Scholar] [CrossRef] [PubMed]
- Straub, M.; Schwartz, L.W. Primary pulmonary arrested lesions of coccidioidomycosis and histoplasmosis. Am. J. Clin. Pathol. 1956, 26, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, M.; Ronaghi, R.; Mills, P.K.; Peterson, M.W. Complications and Yield of Computed Tomography-Guided Transthoracic Core Needle Biopsy of Lung Nodules at a High-Volume Academic Center in an Endemic Coccidioidomycosis Area. Lung 2016, 194, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Deresinski, S.C.; Stevens, D.A. Coccidioidomycosis in compromised hosts. Experience at Stanford University Hospital. Medicine 1975, 54, 377–395. [Google Scholar] [CrossRef]
- Logan, J.L.; Blair, J.E.; Galgiani, J.N. Coccidioidomycosis complicating solid organ transplantation. Semin. Respir. Infect. 2001, 16, 251–256. [Google Scholar] [CrossRef]
- Blair, J.E.; Kusne, S.; Carey, E.J.; Heilman, R.L. The Prevention of Recrudescent Coccidioidomycosis After Solid Organ Transplantation. Transplantation 2007, 83, 1182–1187. [Google Scholar] [CrossRef]
- Huff, D.; Ampel, N.M.; Blair, J.E. Coccidioidomycosis and COVID-19 Infection. An Analysis from a Single Medical Center Within the Coccidioidal Endemic Area. Mycopathologia 2022, 187, 199–204. [Google Scholar] [CrossRef]
- Jones, J.L.; Fleming, P.L.; Ciesielski, C.A.; Hu, D.J.; Kaplan, J.E.; Ward, J.W. Coccidioidomycosis among Persons with AIDS in the United States. J. Infect. Dis. 1995, 171, 961–964. [Google Scholar] [CrossRef]
- Shubitz, L.F.; Powell, D.A.; Butkiewicz, C.D.; Lewis, M.L.; Trinh, H.T.; Frelinger, J.A.; Orbach, M.J.; Galgiani, J.N. A Chronic Murine Disease Model of Coccidioidomycosis Using Coccidioides posadasii, Strain 1038. J. Infect. Dis. 2021, 223, 166–173. [Google Scholar] [CrossRef]
- Huppert, M.; Sun, S.H.; Gross, A.J. Evaluation of an experimental animal model for testing antifungal substances. Antimicrob. Agents Chemother. 1972, 1, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Healey, M.C. The Immunosuppressive Effects of Dexamethasone Administered in Drinking Water to C57BL/6N Mice Infected with Cryptosporidium parvum. J. Parasitol. 1993, 79, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Shubitz, L.F.; Dial, S.M.; Perrill, R.; Casement, R.; Galgiani, J.N. Vaccine-induced cellular immune responses differ from innate responses in susceptible and resistant strains of mice infected with Coccidioides posadasii. Infect. Immun. 2008, 76, 5553–5564. [Google Scholar] [CrossRef] [PubMed]
- Kligman, A.M.; Mescon, H. The periodic-acid-Schiff stain for the demonstration of fungi in animal tissue. J. Bacteriol. 1950, 60, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Ronchetti, S.; Ricci, E.; Migliorati, G.; Gentili, M.; Riccardi, C. How Glucocorticoids Affect the Neutrophil Life. Int. J. Mol. Sci. 2018, 19, 4090. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Thompson, G.R., 3rd; Hastey, C.J.; Hodge, G.C.; Lunetta, J.M.; Pappagianis, D.; Heinrich, V. Coccidioides Endospores and Spherules Draw Strong Chemotactic, Adhesive, and Phagocytic Responses by Individual Human Neutrophils. PLoS ONE 2015, 10, e0129522. [Google Scholar] [CrossRef] [PubMed]
- Phuah, J.Y.; Mattila, J.T.; Lin, P.L.; Flynn, J.L. Activated B Cells in the Granulomas of Nonhuman Primates Infected with Mycobacterium tuberculosis. Am. J. Pathol. 2012, 181, 508–514. [Google Scholar] [CrossRef]
- Gideon, H.P.; Phuah, J.; Myers, A.J.; Bryson, B.D.; Rodgers, M.A.; Coleman, M.T.; Maiello, P.; Rutledge, T.; Marino, S.; Fortune, S.M.; et al. Variability in tuberculosis granuloma T cell responses exists, but a balance of pro- and anti-inflammatory cytokines is associated with sterilization. PLoS Pathog. 2015, 11, e1004603. [Google Scholar] [CrossRef]
- Huang, Z.; Luo, Q.; Guo, Y.; Chen, J.; Xiong, G.; Peng, Y.; Ye, J.; Li, J. Mycobacterium tuberculosis-Induced Polarization of Human Macrophage Orchestrates the Formation and Development of Tuberculous Granulomas In Vitro. PLoS ONE 2015, 10, e0129744. [Google Scholar] [CrossRef]
- Walzer, P.D.; Runck, J.; Steele, P.; White, M.; Linke, M.J.; Sidman, C.L. Immunodeficient and immunosuppressed mice as models to test anti-Pneumocystis carinii drugs. Antimicrob. Agents Chemother. 1997, 41, 251–258. [Google Scholar] [CrossRef]
- Odio, C.D.; Marciano, B.E.; Galgiani, J.N.; Holland, S.M. Risk Factors for Disseminated Coccidioidomycosis, United States. Emerg. Infect. Dis. 2017, 23, 308–311. [Google Scholar] [CrossRef]
- Keckich, D.W.; Blair, J.E.; Vikram, H.R. Coccidioides Fungemia in Six Patients, with a Review of the Literature. Mycopathologia 2010, 170, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Gideon, H.P.; Hughes, T.K.; Tzouanas, C.N.; Wadsworth, M.H.; Tu, A.A.; Gierahn, T.M.; Peters, J.M.; Hopkins, F.F.; Wei, J.-R.; Kummerlowe, C.; et al. Multimodal profiling of lung granulomas reveals cellular correlates of tuberculosis control. bioRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Modlin, R.L.; Gebhard, J.F.; Taylor, C.R.; Rea, T.H. In situ characterization of T lymphocyte subsets in the reactional states of leprosy. Clin. Exp. Immunol. 1983, 53, 17–24. [Google Scholar] [PubMed]
- Ma, F.; Hughes, T.K.; Teles, R.M.B.; Andrade, P.R.; de Andrade Silva, B.J.; Plazyo, O.; Tsoi, L.C.; Do, T.; Wadsworth, M.H.; Oulee, A.; et al. The cellular architecture of the antimicrobial response network in human leprosy granulomas. Nat. Immunol. 2021, 22, 839–850. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shubitz, L.F.; Powell, D.A.; Dial, S.M.; Butkiewicz, C.D.; Trinh, H.T.; Hsu, A.P.; Buntzman, A.; Frelinger, J.A.; Galgiani, J.N. Reactivation of Coccidioidomycosis in a Mouse Model of Asymptomatic Controlled Disease. J. Fungi 2022, 8, 991. https://doi.org/10.3390/jof8100991
Shubitz LF, Powell DA, Dial SM, Butkiewicz CD, Trinh HT, Hsu AP, Buntzman A, Frelinger JA, Galgiani JN. Reactivation of Coccidioidomycosis in a Mouse Model of Asymptomatic Controlled Disease. Journal of Fungi. 2022; 8(10):991. https://doi.org/10.3390/jof8100991
Chicago/Turabian StyleShubitz, Lisa F., Daniel A. Powell, Sharon M. Dial, Christine D. Butkiewicz, Hien T. Trinh, Amy P. Hsu, Adam Buntzman, Jeffrey A. Frelinger, and John N. Galgiani. 2022. "Reactivation of Coccidioidomycosis in a Mouse Model of Asymptomatic Controlled Disease" Journal of Fungi 8, no. 10: 991. https://doi.org/10.3390/jof8100991