
Global Transcriptional Response of Aspergillus niger to Blocked Active Citrate Export through Deletion of the Exporter Gene

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains Media, Growth, and Fermentation Conditions
2.2. Construction and Initial Screening of ΔcitT Knock-Out Strains for Citrate Secretion
2.3. Genome Sequencing of the citT Mutant, Comparative Genomics and Bioinformatics
2.4. Transcriptome and Pathway Enrichment Analysis
3. Results
3.1. Genome Comparison between Strain N402 and the Citrate-Hyperproducing Strain H915-1
3.2. Transcriptional Regulation of the Citrate Exporter Gene
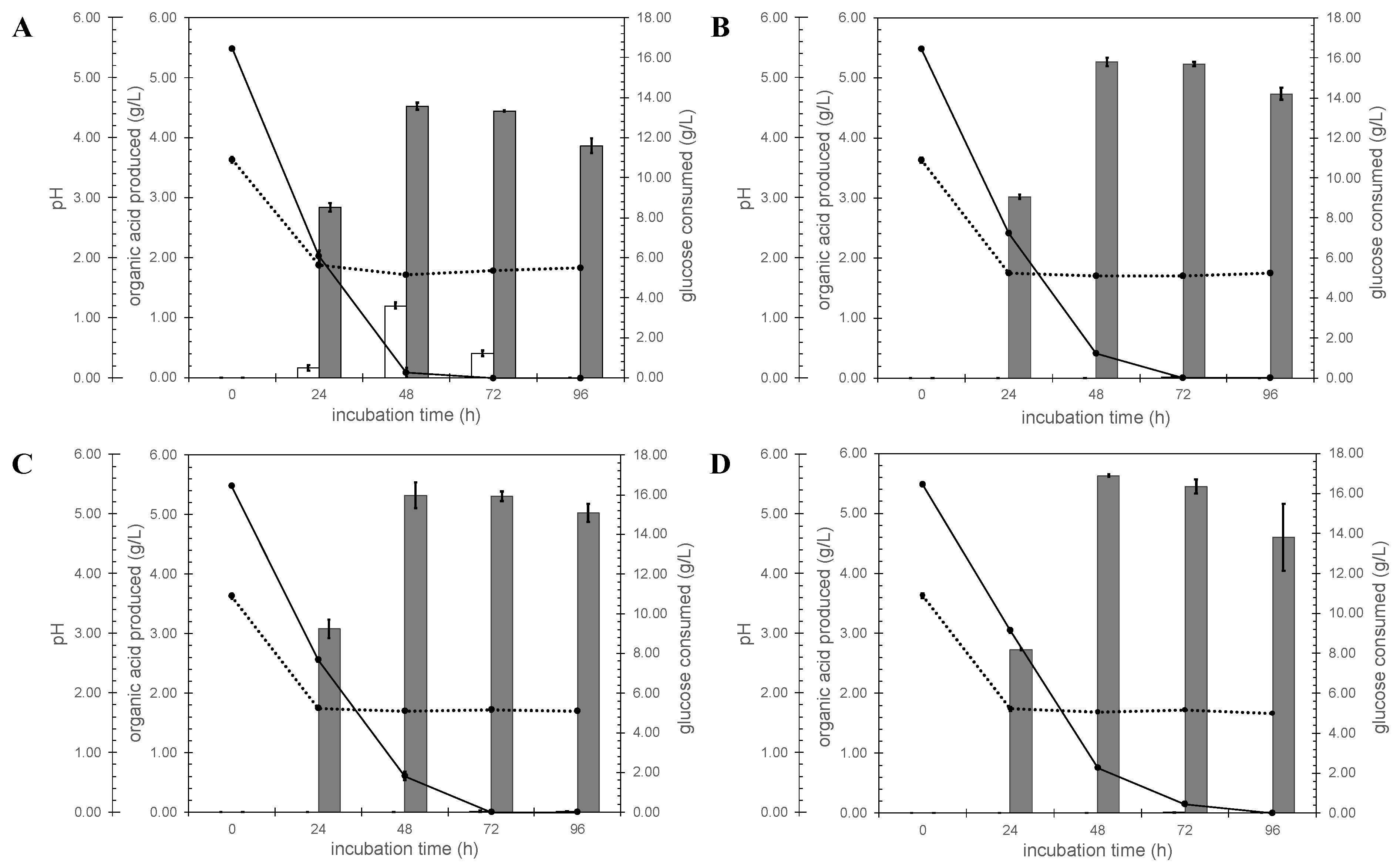
3.3. In Vivo Validation of the A. niger Gene Encoding the Citrate Exporter
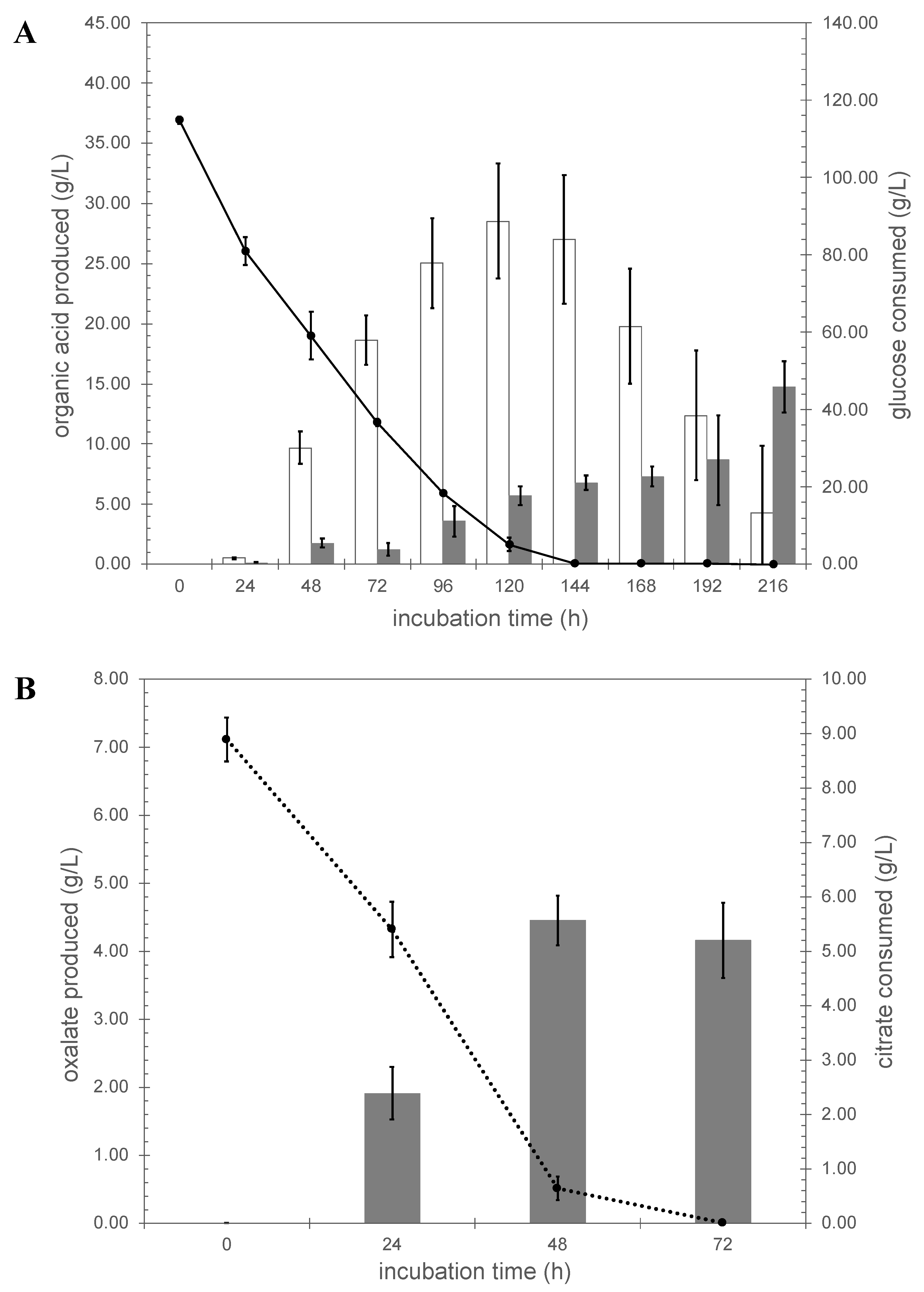
3.4. Organic Acids Production by A. niger Strains N402 and ΔcitT
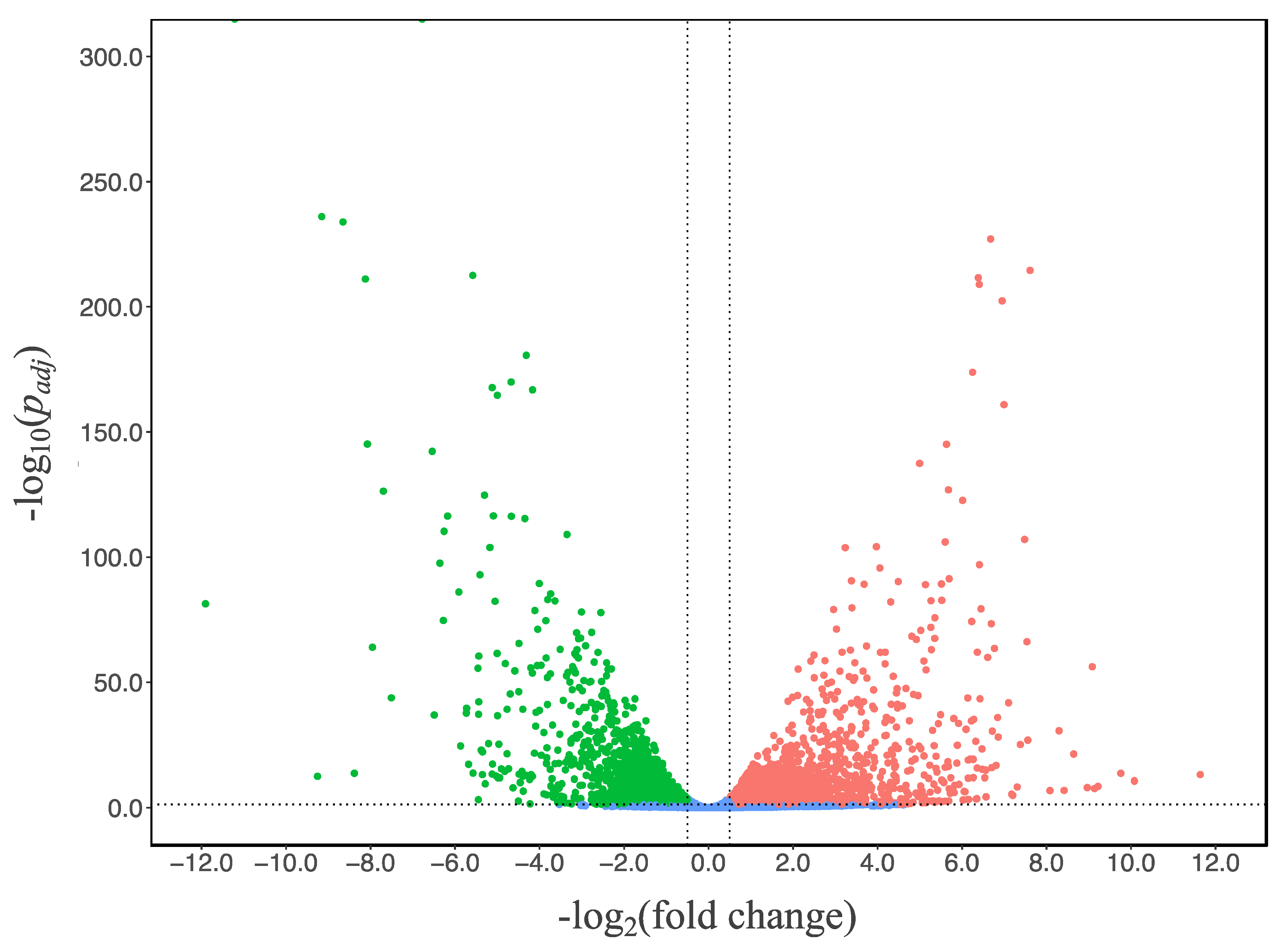
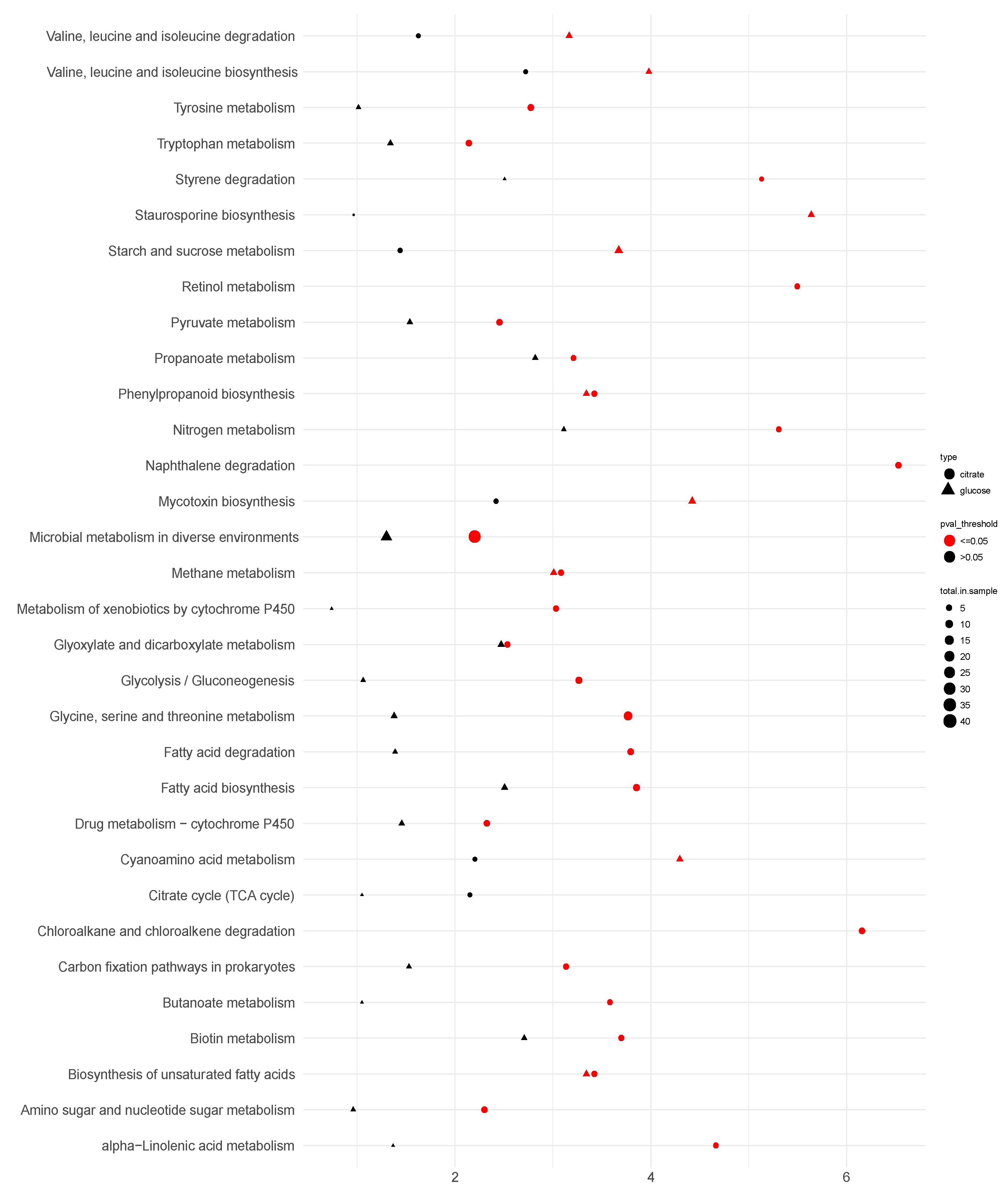
3.5. Transcriptome and Pathway Enrichment Analysis
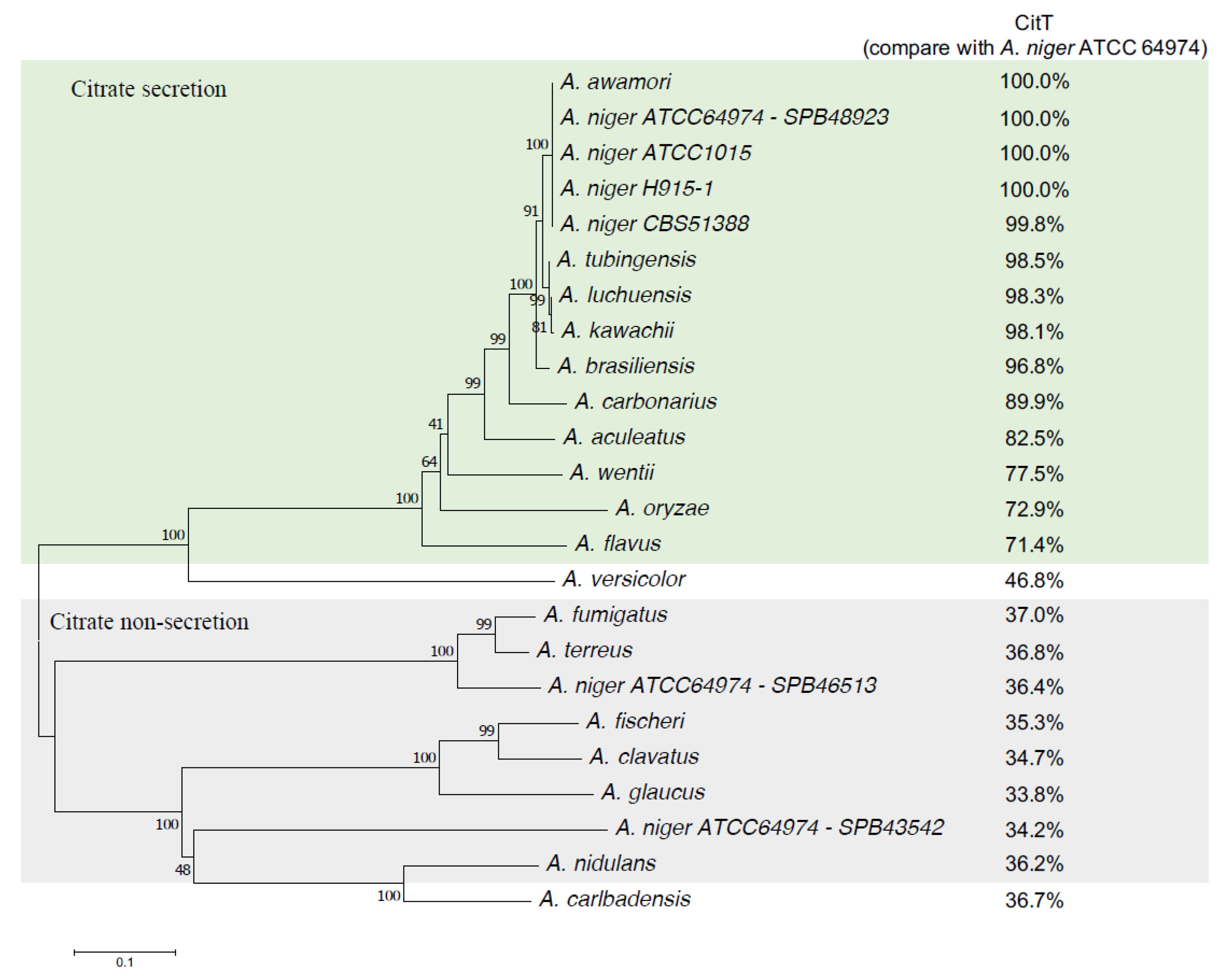
3.6. Distribution of the Citrate Exporter Gene among Aspergillus Species
4. Discussion
4.1. citT Is Responsible for Active A. niger Citrate Export, but Not for Citrate Uptake
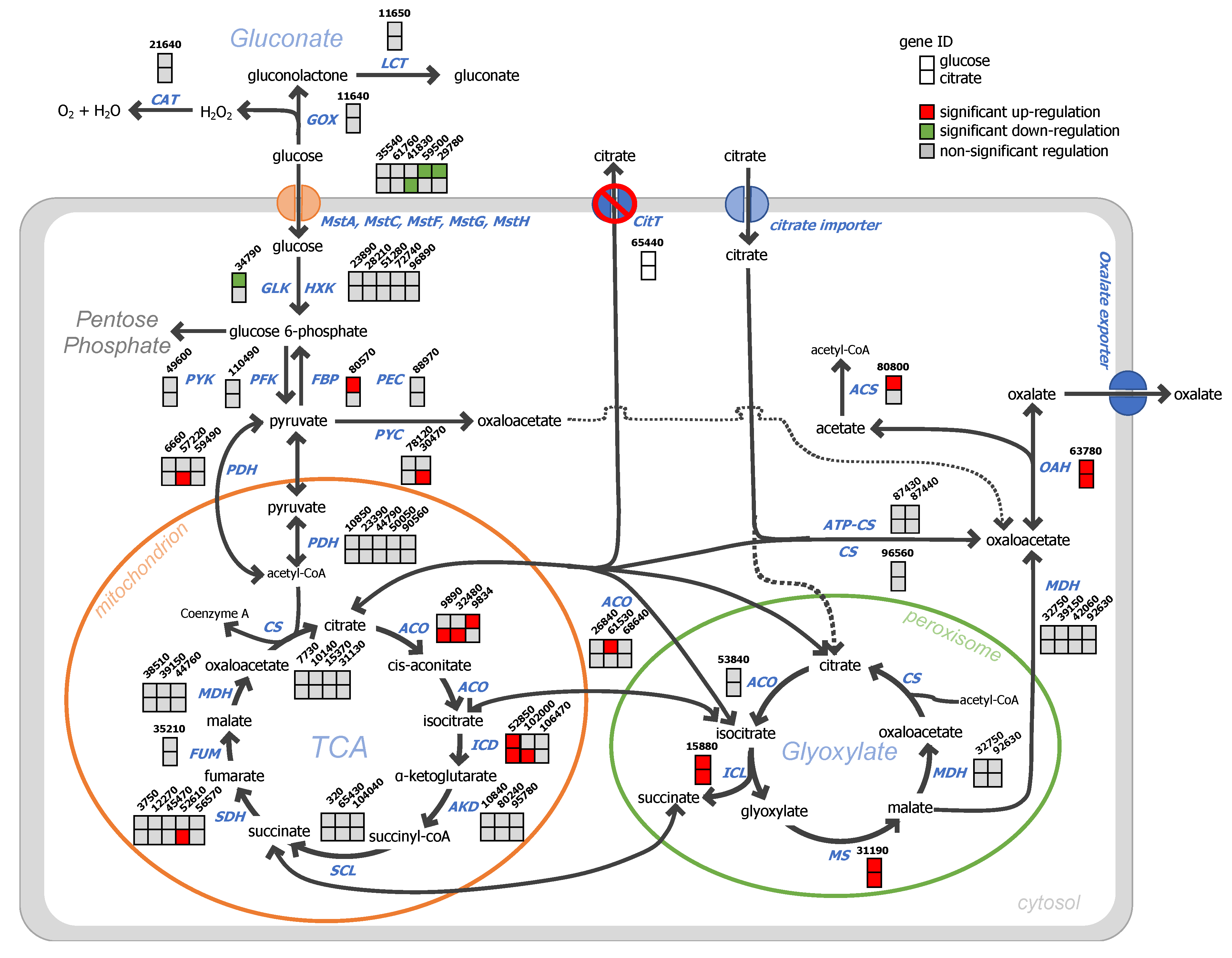
4.2. Blocking Active Citrate Export under Citrate Producing Conditions Reduces the Uptake of Glucose, Induces the Secretion of Oxalic Acid, and Activates the Glyoxylate Shunt
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fiedler, M.M.; Nitsche, B.; Wanka, F.; Meyer, V. Aspergillus: A cell factory with unlimited prospects. In Applications of Microbial Engineering, 1st ed.; Gupta, V.K.S., Monika Maki, M., Tuohy, M., Mazutti, M.A., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 1–51. [Google Scholar]
- Show, P.L.; Oladele, K.O.; Siew, Q.Y.; Aziz Zakry, F.A.; Lan, J.C.-W.; Ling, T.C. Overview of citric acid production from Aspergillus niger. Front. Life Sci. 2015, 8, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Cairns, T.C.; Nai, C.; Meyer, V. How a fungus shapes biotechnology: 100 years of Aspergillus niger research. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Singh Dhillon, G.; Kaur Brar, S.; Verma, M.; Tyagi, R.D. Recent Advances in Citric Acid Bio-production and Recovery. Food Bioprocess Technol. 2011, 4, 505–529. [Google Scholar] [CrossRef]
- Sweis, I.E.; Cressey, B.C. Potential role of the common food additive manufactured citric acid in eliciting significant inflammatory reactions contributing to serious disease states: A series of four case reports. Toxicol. Rep. 2018, 5, 808–812. [Google Scholar] [CrossRef]
- Ruijter, G.J.G.; Van de Vondervoort, P.J.I.; Visser, J. Oxalic acid production by Aspergillus niger: An oxalate-non-producing mutant produces citric acid at pH 5 and in the presence of manganese. Microbiology 1999, 145, 2569–2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaffa, L.; Kubicek, C.P. Aspergillus niger citric acid accumulation: Do we understand this well working black box? Appl. Microbiol. Biotechnol. 2003, 61, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, J.K.; Lasure, L.L. Organic acid production by filamentous fungi. In Advances in Fungal Biotechnology for Industry, Agriculture, and Medicine; Tkacz, J.S., Lange, L., Eds.; Springer US: Boston, MA, USA, 2004; pp. 307–340. [Google Scholar]
- Andersen, M.R.; Lehmann, L.; Nielsen, J. Systemic analysis of the response of Aspergillus niger to ambient pH. Genome Biol. 2009, 10, R47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubicek, C.P.; Zehentgruber, O.; El-Kalak, H.; Röhr, M. Regulation of citric acid production by oxygen: Effect of dissolved oxygen tension on adenylate levels and respiration in Aspergillus niger. Eur. J. Appl. Microbiol. Biotechnol. 1980, 9, 101–115. [Google Scholar] [CrossRef]
- Hang, Y.D.; Woodams, E.E. Apple pomace: A potential substrate for citric acid production by Aspergillus niger. Biotechnol. Lett. 1984, 6, 763–764. [Google Scholar] [CrossRef]
- Kareem, S.O.; Rahman, R.A. Utilization of banana peels for citric acid production by Aspergillus niger. Agric. Biol. J. N. Am. 2011, 4, 384–387. [Google Scholar] [CrossRef]
- Papagianni, M. Advances in citric acid fermentation by Aspergillus niger: Biochemical aspects, membrane transport and modeling. Biotechnol. Adv. 2007, 25, 244–263. [Google Scholar] [CrossRef]
- Bateman, A.; Coin, L.; Durbin, R.; Finn, R.D.; Hollich, V.; Griffiths-Jones, S.; Khanna, A.; Marshall, M.; Moxon, S.; Sonnhammer, E.L.L.; et al. The Pfam protein families database. Nucleic Acids Res. 2004, 32, D138–D141. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.R.; Salazar, M.P.; Schaap, P.J.; Van De Vondervoort, P.J.I.; Culley, D.; Thykaer, J.; Frisvad, J.C.; Nielsen, K.F.; Albang, R.; Albermann, K.; et al. Comparative genomics of citric-acid-producing Aspergillus niger ATCC 1015 versus enzyme-producing CBS 513.88. Genome Res. 2011, 21, 885–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sloothaak, J.; Odoni, D.I.; de Graaff, L.H.; Martins dos Santos, V.A.P.; Schaap, P.J.; Tamayo-Ramos, J.A. Aspergillus niger membrane-associated proteome analysis for the identification of glucose transporters. Biotechnol. Biofuels 2015, 8, 150. [Google Scholar] [CrossRef] [PubMed]
- Pel, H.J.; De Winde, J.H.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; de Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Laothanachareon, T.; Tamayo-Ramos, J.A.; Nijsse, B.; Schaap, P.J. Forward Genetics by Genome Sequencing Uncovers the Central Role of the Aspergillus niger goxB Locus in Hydrogen Peroxide Induced Glucose Oxidase Expression. Front. Microbiol. 2018, 9, 2269. [Google Scholar] [CrossRef]
- Odoni, D.I. Organic Acid Production in Aspergillus niger and other Filamentous Fungi; Wageningen University & Research: Wageningen, The Netherlands, 2017. [Google Scholar]
- Odoni, D.I.; Vazquez-Vilar, M.; Van Gaal, M.P.; Schonewille, T.; Martins Dos Santos, V.A.; Tamayo-Ramos, J.A.; Suarez-Diez, M.; Schaap, P.J. Aspergillus niger citrate exporter revealed by comparison of two alternative citrate producing conditions. FEMS Microbiol. Lett. 2019, 366, fnz071. [Google Scholar] [CrossRef] [PubMed]
- Steiger, M.G.; Rassinger, A.; Mattanovich, D.; Sauer, M. Engineering of the citrate exporter protein enables high citric acid production in Aspergillus niger. Metab. Eng. 2019, 52, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Bos, C.J.; Debets, A.J.M.; Swart, K.; Huybers, A.; Kobus, G.; Slakhorst, S.M. Genetic analysis and the construction of master strains for assignment of genes to six linkage groups in Aspergillus niger. Curr. Genet. 1988, 14, 437–443. [Google Scholar] [CrossRef]
- Carvalho, N.D.S.P.; Arentshorst, M.; Jin Kwon, M.; Meyer, V.; Ram, A.F.J. Expanding the ku70 toolbox for filamentous fungi: Establishment of complementation vectors and recipient strains for advanced gene analyses. Appl. Microbiol. Biotechnol. 2010, 87, 1463–1473. [Google Scholar] [CrossRef] [Green Version]
- Vishniac, W.; Santer, M. The thiobacilli. Bacteriol. Rev. 1957, 21, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Arentshorst, M.; Ram, A.F.J.; Meyer, V. Using Non-homologous End-Joining-Deficient strains for functional gene analyses in filamentous fungi. In Plant Fungal Pathogens: Methods and Protocols; Bolton, M.D., Thomma, B.P.H.J., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 133–150. [Google Scholar]
- Kirimura, K.; Kobayashi, K.; Ueda, Y.; Hattori, T. Phenotypes of gene disruptants in relation to a putative mitochondrial malate–citrate shuttle protein in citric acid-producing Aspergillus niger. Biosci. Biotechnol. Biochem. 2016, 80, 1737–1746. [Google Scholar] [CrossRef] [Green Version]
- Usami, S. Production of citric acid by submerged culture. In Mem School Sci. Engg; Waseda University: Tokio, Japan, 1978; Volume 42, pp. 17–26. [Google Scholar]
- Odoni, D.I.; Van Gaal, M.P.; Schonewille, T.; Tamayo-Ramos, J.A.; Martins Dos Santos, V.A.P.; Suarez-Diez, M.; Schaap, P.J. Aspergillus niger secretes citrate to increase iron bioavailability. Front. Microbiol. 2017, 8, 1424. [Google Scholar] [CrossRef] [PubMed]
- Arentshorst, M.; Niu, J.; Ram, A.F.J. Efficient generation of Aspergillus niger knock out strains by combining NHEJ mutants and a split marker approach. In Genetic Transformation Systems in Fungi, Volume 1; Van Den Berg, M.A., Maruthachalam, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 263–272. [Google Scholar]
- Arentshorst, M.; Lagendijk, E.L.; Ram, A.F. A new vector for efficient gene targeting to the pyrG locus in Aspergillus niger. Fungal Biol. Biotechnol. 2015, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minkin, I.; Patel, A.; Kolmogorov, M.; Vyahhi, N.; Pham, S. Sibelia: A scalable and comprehensive synteny block generation tool for closely related microbial genomes. In Algorithms in Bioinformatics; WABI 2013; Darling, A., Stoye, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 8126, pp. 215–229. [Google Scholar]
- Sloothaak, J.; Tamayo-Ramos, J.A.; Odoni, D.I.; Laothanachareon, T.; Derntl, C.; Mach-Aigner, A.R.; Martins Dos Santos, V.A.P.; Schaap, P.J. Identification and functional characterization of novel xylose transporters from the cell factories Aspergillus niger and Trichoderma reesei. Biotechnol. Biofuels 2016, 9, 148. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [Green Version]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A Cytoscape plugin to assess overrepresentation of Gene Ontology categories in Biological Networks. Bioinformatics 2005, 21, 3448–3449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blom, R.H.; Pfeifer, V.F.; Moyer, A.J.; TraufRer, D.H.; Conway, H.F.; Crocker, C.K.; Farison, R.E.; Hannibal, D.V. Sodium gluconate production. Fermentation with Aspergillus niger. Ind. Eng. Chem. 1952, 44, 435–440. [Google Scholar] [CrossRef]
- Yin, X.; Shin, H.-D.; Li, J.; Du, G.; Liu, L.; Chen, J. Comparative genomics and transcriptome analysis of Aspergillus niger and metabolic engineering for citrate production. Sci. Rep. 2017, 7, 41040. [Google Scholar] [CrossRef] [Green Version]
- Emiliani, E.; Riera, B. Enzymatic oxalate decarboxylation in Aspergillus niger: II. Hydrogen peroxide formation and other characteristics of the oxalate decarboxylase. Biochim. Biophys. Acta (BBA) Enzymol. 1968, 167, 414–421. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Schreferl-Kunar, G.; Wöhrer, W.; Röhr, M. Evidence for a cytoplasmic pathway of oxalate biosynthesis in Aspergillus niger. Appl. Environ. Microbiol. 1988, 54, 633–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joosten, H.-J.; Han, Y.; Niu, W.; Vervoort, J.; Dunaway-Mariano, D.; Schaap, P.J. Identification of fungal oxaloacetate hydrolyase within the isocitrate lyase/PEP mutase enzyme superfamily using a sequence marker-based method. Proteins Struct. Funct. Bioinform. 2008, 70, 157–166. [Google Scholar] [CrossRef]
- Han, Y.; Joosten, H.-J.; Niu, W.; Zhao, Z.; Mariano, P.S.; McCalman, M.; van Kan, J.; Schaap, P.J.; Dunaway-Mariano, D. Oxaloacetate hydrolase, the C–C bond lyase of oxalate secreting fungi. J. Biol. Chem. 2007, 282, 9581–9590. [Google Scholar] [CrossRef] [Green Version]
- Guebel, D.V.; Torres Darias, N.V. Optimization of the citric acid production by Aspergillus niger through a metabolic flux balance model. Electron. J. Biotechnol. 2001, 4. [Google Scholar] [CrossRef]
- Netik, A.; Torres, N.V.; Riol, J.-M.; Kubicek, C.P. Uptake and export of citric acid by Aspergillus niger is reciprocally regulated by manganese ions. Biochim. Biophys. Acta (BBA) Biomembr. 1997, 1326, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, E.; Kadooka, C.; Okutsu, K.; Yoshizaki, Y.; Takamine, K.; Goto, M.; Tamaki, H.; Futagami, T. Citrate exporter enhances both extracellular and intracellular citric acid accumulation in the koji fungi Aspergillus luchuensis mut. kawachii and Aspergillus oryzae. J. Biosci. Bioeng. 2020, 131, 68–76. [Google Scholar] [CrossRef]
- Jernejc, K.; Vendramin, M.; Cimerman, A. Lipid composition of Aspergillus niger in citric acid accumulating and nonaccumulating conditions. Enzym. Microb. Technol. 1989, 11, 452–456. [Google Scholar] [CrossRef]
- Šimkovič, M.; Kaliňák, M.; Burgstaller, W.; Varečka, L.U. Characterization of an inducible citrate uptake system in Penicillium simplicissimum. FEMS Microbiol. Lett. 2002, 213, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Liu, P.; Madzak, C.; Du, G.; Zhou, J.; Chen, J. Identification and application of keto acids transporters in Yarrowia lipolytica. Sci. Rep. 2015, 5, 8138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sá-Pessoa, J.; Amillis, S.; Casal, M.; Diallinas, G. Expression and specificity profile of the major acetate transporter AcpA in Aspergillus nidulans. Fungal Genet. Biol. 2015, 76, 93–103. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Locus Tag. | Protein ID | log2(Fold Change) | TMHs |
|---|---|---|---|
| ATCC64974_65440 | SPB48923.1 | −6.7765 | 12 |
| ATCC64974_85340 | SPB50921.1 | −5.5734 | 12 |
| ATCC64974_44020 | SPB46772.1 | −5.0525 | 10 |
| ATCC64974_28550 | SPB45214.1 | −4.9393 | 12 |
| ATCC64974_43890 | SPB46759.1 | −4.8217 | 8 |
| ATCC64974_4840 | SPB42837.1 | −4.7665 | 12 |
| ATCC64974_25910 | SPB44949.1 | −4.4900 | 12 |
| ATCC64974_29710 | SPB45330.1 | −4.1990 | 11 |
| ATCC64974_26160 | SPB44974.1 | −3.8357 | 12 |
| ATCC64974_85360 | SPB50923.1 | −3.8232 | 11 |
| ATCC64974_950 | SPB42446.1 | −3.7484 | 14 |
| ATCC64974_26910 | SPB45050.1 | −3.5111 | 9 |
| ATCC64974_93260 | SPB51716.1 | −3.2956 | 12 |
| ATCC64974_72710 | SPB49652.1 | −3.2174 | 12 |
| ATCC64974_21780 | SPB44533.1 | −3.1779 | 9 |
| ATCC64974_105810 | SPB52979.1 | −3.1237 | 10 |
| ATCC64974_72040 | SPB49585.1 | −3.0387 | 9 |
| ATCC64974_10650 | SPB43419.1 | −3.0383 | 13 |
| Protein ID | Description | log2(Fold Change) | ||
|---|---|---|---|---|
| Glucose | Citrate | |||
| ATCC64974_103440 | SPB52738.1 | 2OG-Fe(II) oxygenase family oxidoreductase | 4.7227 | 2.2847 |
| ATCC64974_30940 | SPB45454.1 | carboxymuconolactone decarboxylase family protein | 2.3894 | 1.6078 |
| ATCC64974_6060 | SPB42959.1 | HMGL-like family protein | 2.1506 | 1.1072 |
| ATCC64974_91920 | SPB51582.1 | uncharacterized protein | 2.1075 | 0.7707 |
| ATCC64974_63780 | SPB48756.1 | oxaloacetate acetylhydrolase | 2.0182 | 1.9265 |
| ATCC64974_26450 | SPB45003.1 | zinc-binding dehydrogenase family protein | 1.8572 | 1.4575 |
| ATCC64974_15880 | SPB43942.1 | isocitrate lyase | 1.5948 | 0.8524 |
| ATCC64974_67790 | SPB49158.1 | acetyl-CoA hydrolase | 1.5526 | 0.9054 |
| ATCC64974_4720 | SPB42825.1 | O-methyltransferase family protein | 1.4041 | 0.8684 |
| ATCC64974_31190 | SPB45479.1 | malate synthase | 1.3546 | 1.2960 |
| ATCC64974_103400 | SPB52734.1 | uncharacterized protein | 1.3405 | 1.3934 |
| ATCC64974_18700 | SPB44224.1 | alpha/beta hydrolase family protein | 1.2769 | 0.9495 |
| ATCC64974_71600 | SPB49541.1 | uncharacterized protein | 1.2265 | 0.8148 |
| ATCC64974_4600 | SPB42813.1 | AMP-binding enzyme family protein | 1.0504 | 0.6859 |
| ATCC64974_58540 | SPB48229.1 | endo-polygalacturonase B | 0.9209 | 0.7973 |
| ATCC64974_63790 | SPB48757.1 | DUF1275 domain protein | 0.9072 | 1.2557 |
| ATCC64974_47060 | SPB47077.1 | methyltransferase domain family protein | 0.7848 | 1.0456 |
| ATCC64974_102280 | SPB52622.1 | glycosyl hydrolase family 61 family protein | 0.7174 | 0.8594 |
| ATCC64974_82680 | SPB50654.1 | WSC domain family protein | 0.6728 | 0.8253 |
| ATCC64974_4610 | SPB42814.1 | fungal Zn(2)-Cys(6) binuclear cluster domain family protein | 0.6693 | 1.3609 |
| ATCC64974_34830 | SPB45846.1 | TAP-like protein family protein | 0.6533 | 0.7306 |
| ATCC64974_104990 | SPB52896.1 | peptidase inhibitor I78 family protein | 0.6393 | 0.5458 |
| ATCC64974_64070 | SPB48785.1 | aldehyde dehydrogenase family protein | 0.6174 | 0.7522 |
| ATCC64974_63760 | SPB48754.1 | purine nucleoside permease | 0.5466 | 0.9801 |
| ATCC64974_34150 | SPB45775.1 | short chain dehydrogenase family protein | 0.5283 | 0.6748 |
| ATCC64974_52850 | SPB47657.1 | mitochondrial NADP- and NAD-isocitrate dehydrogenases | 0.5169 | 0.6755 |
| ATCC64974_34010 | SPB45761.1 | uncharacterized protein | 0.5107 | 0.7421 |
| Gene Locus Tag | Protein ID | Description | log2(Fold Change) | |
|---|---|---|---|---|
| Glucose | Citrate | |||
| ATCC64974_89450 | SPB51334.1 | uncharacterized protein | −6.2145 | −4.0918 |
| ATCC64974_92610 | SPB51651.1 | uncharacterized protein | −3.0166 | −1.3542 |
| ATCC64974_1740 | SPB42525.1 | alcohol dehydrogenase | −2.4351 | −1.2566 |
| ATCC64974_12500 | SPB43604.1 | laccase | −2.3820 | −0.7627 |
| ATCC64974_93830 | SPB51773.1 | uncharacterized protein | −2.1959 | −2.0151 |
| ATCC64974_89550 | SPB51344.1 | acid-stable alpha-amylase | −1.8176 | −0.7229 |
| ATCC64974_88150 | SPB51204.1 | fungal Zn(2)-Cys(6) binuclear cluster domain family protein | −1.7703 | −2.6046 |
| ATCC64974_1920 | SPB42543.1 | uncharacterized protein | −1.7534 | −0.7516 |
| ATCC64974_96990 | SPB52093.1 | high-affinity xylose transporter (XltA) | −1.6515 | −1.4477 |
| ATCC64974_87010 | SPB51090.1 | uncharacterized protein | −1.5061 | −0.7842 |
| ATCC64974_23000 | SPB44655.1 | proteinase aspergillopepsin II | −1.3996 | −1.0743 |
| ATCC64974_81710 | SPB50557.1 | extracellular alpha-glucosidase aglU | −1.3178 | −0.9158 |
| ATCC64974_48920 | SPB47263.1 | carboxypeptidase | −1.0439 | −1.0691 |
| ATCC64974_88220 | SPB51211.1 | mago binding family protein | −0.8482 | −0.5610 |
| ATCC64974_26630 | SPB45021.1 | uncharacterized protein | −0.8254 | −1.2211 |
| ATCC64974_103090 | SPB52703.1 | glycine zipper 2TM domain family protein | −0.7409 | −0.9068 |
| ATCC64974_88240 | SPB51213.1 | ubiquitin-protein ligase E3 component | −0.7202 | −0.5844 |
| ATCC64974_14700 | SPB43824.1 | alpha-glucosidase | −0.7012 | −0.7846 |
| ATCC64974_88190 | SPB51208.1 | cytochrome P450 alkane hydroxylase | −0.6886 | −0.9308 |
| ATCC64974_88170 | SPB51206.1 | adenylate cyclase AcyA | −0.6885 | −0.6351 |
| ATCC64974_21300 | SPB44485.1 | MFS transporter | −0.6525 | −0.6241 |
| ATCC64974_88210 | SPB51210.1 | protoporphyrinogen oxidase | −0.6309 | −0.5470 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laothanachareon, T.; Bruinsma, L.; Nijsse, B.; Schonewille, T.; Suarez-Diez, M.; Tamayo-Ramos, J.A.; Martins dos Santos, V.A.P.; Schaap, P.J. Global Transcriptional Response of Aspergillus niger to Blocked Active Citrate Export through Deletion of the Exporter Gene. J. Fungi 2021, 7, 409. https://doi.org/10.3390/jof7060409
Laothanachareon T, Bruinsma L, Nijsse B, Schonewille T, Suarez-Diez M, Tamayo-Ramos JA, Martins dos Santos VAP, Schaap PJ. Global Transcriptional Response of Aspergillus niger to Blocked Active Citrate Export through Deletion of the Exporter Gene. Journal of Fungi. 2021; 7(6):409. https://doi.org/10.3390/jof7060409
Chicago/Turabian StyleLaothanachareon, Thanaporn, Lyon Bruinsma, Bart Nijsse, Tom Schonewille, Maria Suarez-Diez, Juan Antonio Tamayo-Ramos, Vitor A. P. Martins dos Santos, and Peter J. Schaap. 2021. "Global Transcriptional Response of Aspergillus niger to Blocked Active Citrate Export through Deletion of the Exporter Gene" Journal of Fungi 7, no. 6: 409. https://doi.org/10.3390/jof7060409