Immune Response of Galleria mellonella against Human Fungal Pathogens


1
Albert Einstein College of Medicine, Bronx 10461, NY, USA
2
Mycology Reference Laboratory, National Centre for Microbiology, Instituto de Salud Carlos III, Majadahonda, 28220 Madrid, Spain
*
Author to whom correspondence should be addressed.
J. Fungi 2019, 5(1), 3; https://doi.org/10.3390/jof5010003
Submission received: 12 November 2018
/
Revised: 14 December 2018
/
Accepted: 20 December 2018
/
Published: 26 December 2018
(This article belongs to the Special Issue Alternative Models for the Study of Fungal Host-Pathogen Interactions)
{kind=link}
Abstract
:In many aspects, the immune response against pathogens in insects is similar to the innate immunity in mammals. This has caused a strong interest in the scientific community for the use of this model in research of host–pathogen interactions. In recent years, the use of Galleria mellonella larvae, an insect belonging to the Lepidoptera order, has emerged as an excellent model to study the virulence of human pathogens. It is a model that offers many advantages; for example, it is easy to handle and establish in every laboratory, the larvae have a low cost, and they tolerate a wide range of temperatures, including human temperature 37 °C. The immune response of G. mellonella is innate and is divided into a cellular component (hemocytes) and humoral component (antimicrobial peptides, lytic enzymes, and peptides and melanin) that work together against different intruders. It has been shown that the immune response of this insect has a great specificity and has the ability to distinguish between different classes of microorganisms. In this review, we delve into the different components of the innate immune response of Galleria mellonella, and how these components manifest in the infection of fungal pathogens including Candida albicans, Aspergillus fumigatus, Cryptococcus neoformans, and Histoplasma capsulatum.
1. Introduction
The investigation of infectious diseases has been one of the priority fields in medicine in recent years. The use of animal models is crucial in this area, as it allows us to define the causative agents of these diseases, examine mechanisms of interaction between host/pathogen, and evaluate the efficacy of drugs among other uses. One of the most commonly used models for studying microbial infections involves the experimentation with mice. This model is very versatile because it allows the use of genetically modified animals and the characterization of the role of the host immune response during infection. Despite its importance, the use of mouse models has several ethical and logistical issues associated. The facilities and maintenance of these animals is expensive. In addition, many models of infection can induce acute sepsis and physiological alterations in animals, which can create great discomfort and pain. For this reason, animal experimentation is nowadays regulated by authorities and bioethical committees. To reduce these bioethical problems, there is a strong trend to apply the “3 Rs” rule in experiments that involve animal use, which are: reduce the numbers of animals; refine the protocols to increase animal comfort and reduce pain; replace animals for other models that do not have bioethical problems associated [1]. In addition, the use of mice for the study of some infectious diseases might present other limitations. For example, in the case of fungal pathogens, the development of murine infection models is not straightforward. Except a few cases (such as Candida albicans or Cryptococcus neoformans), establishment of an infection and disease in mice requires the use of immunosuppressants or may take a considerable amount of time. Therefore, virulence is often assessed by measuring the fungal burden in the animals rather than death. For these reasons, particularly in the field of fungal pathogens, microbiologists have sought “non-conventional” hosts to investigate microbial pathogenesis and for testing antimicrobial drugs [1]. Among them, nematodes (mainly Caenorhabditis elegans) and insects are being more widely used to study infectious diseases.
The Insecta class (insects) are a large and very diverse group of invertebrate animals that contains more than one million described species [2]. Among the main orders of insects are the Diptera, Hemiptera, Coleoptera, Hymenoptera, and Lepidoptera, to which Galleria mellonella belongs. In nature, they are exposed to a wide variety of pathogens, which has led to the development of protective systems that include resistant physical barriers, such as the cuticle. In addition, they have developed complex and effective immune defense systems.
During its life cycle, G. mellonella can be found in the larval, pupal, or moth form and it is known as honeycomb moth. The life cycle of this organism comprises a larva stage (size around 1–3 cm) that transforms into pupae and finally into moth (3–4 cm) [3,4]. The size of the larvae makes their manipulation easy. Galleria mellonella has been used prominently as a model to investigate microbial pathogenesis. The application of this model to study the role of virulence factors in disease and efficacy of antimicrobial compounds has been demonstrated in more than 1000 publications on PubMed [5]. For this purpose, mainly adult larvae are used, before they transform into the pupa stage of the life cycle. The structure of this insect is very simple, it consists of a digestive tube, a neural tube that runs from the frontal side through the dorsal part, and a fat body. All of these organs are surrounded by hemolymph.
Given that some aspects of the G. mellonella innate immunity are similar to the immunity of mammals, it makes this model a feasible option to study human infections [6]. Recently, the use of this model in the mycology field has been applied to Aspergillus fumigatus [7], Candida albicans [8,9,10], and Cryptococcus neoformans [11].
2. Advantages and Disadvantages of the G. mellonella Model
This model offers several advantages which can be highlighted: it is an easy model to implement in the laboratory without the need to invest a large amount of money; due to their size and life cycle, the larvae are easy to handle and there is no need for personnel with specialized expertise; it is easy to determine the viability of the larvae; they are affordable for all laboratories; it is possible to administer exact doses of drugs and pathogens; they can be incubated and survive in a very wide range of temperatures (including 37 °C, which is the human body temperature) even though the optimum temperature of growth is 25 °C [12].
Although it is a model widely used in research, there are also some limitations. Although its genome has been recently sequenced [13], it is not possible to perform genetic manipulations. In addition, it can be difficult to contact a reliable company that provides the larvae in optimal conditions. In many cases, these companies do not control the absence of infection prior to shipment. There are also some distributors who grow the larvae specifically for research, but the price to purchase is higher. Although working with G. mellonella does not require special equipment, it is recommended to keep the larvae in a confined space to prevent the release of moths. Finally, it requires proper prior planning to have an appropriate supply of larvae of the same size and age for experimentation.
3. Immune Response of G. mellonella
The components that comprise the G. mellonella immune system are: fatty body, lymph nodes, and the hemocytes that originate from the mesoderm. These insects have an open circulatory system referred to as the hemolymph, which occupies the general cavity of the body (hemocele). The immune response of G. mellonella is innate, and it is composed of three parts: physical as well as chemical barriers that protect the insect from the external environment, the humoral response, and the cellular response [17]. The cellular response is mediated by hemocytes (blood cells of G. mellonella). The functions of these hemocytes include storage of various types of substances, as well as providing defense of the organism, based on coagulation mechanisms, phagocytosis, nodulation, and melanization. The humoral response is based on different types of soluble molecules secreted against pathogens. This response mainly comprises lytic enzymes (lysozyme), antimicrobial peptides, and melanin.
4. Cellular Immune Response
The main component of the cellular immune response is composed of hemocytes, which are cells with phagocytic capacity that have the ability to produce antimicrobial compounds. Hemocytes are involved in all the mechanisms of insect defense. These cells are not responsible for the transport of gases, since there is a complex network of tubes (tracheal trunks, tracheas, air sacs) that fulfill this function [18]. The main processes of cellular response include coagulation, nodulation, phagocytosis, encapsulation, and melanization [19]. Up to six classes of hemocytes have been described in this insect. These include prohemocytes, plasmatocytes, granular cells, coagulocytes, spherulocytes, and enocytoids [5].
Prohemocytes are a small cell type that have a large nucleus and the ability to differentiate into different cell types [20]. Plasmocytes are larger than prohemocytes with an oval shape and are the most common type of hemocytes. Granular cells have a small nucleus and many granules in the cytoplasm. Spherocytes can be found with different shapes and have inclusions of rounded shape. These hemocytes can transport cuticular components. Coagulocytes are hemocytes whose main function is hemolymph clotting, which is an important process because it is the first defense induced by these insects after microbial exposure. Oenocytoids, which represent 5–10% of the total hemocytes, contain cytoplasmic phenoloxidase and participate in the melanization of the hemolymph [21]. Oenocytoids can also secrete nucleic acids that have been described as “a new alarm signal” in the defense of these insects. Although many studies mention that only plasmatocytes are involved in phagocytosis, it has been shown that both plasmatocytes and granular cells participate in this process as well as in encapsulation [5,19,22].
The process of nodulation occurs when pathogens are recognized and killed by trapping them in hemocyte aggregates (nodules), whereas melanization occurs inside the nodules [16,22]. Encapsulation is performed by granular cells and plasmatocytes. Granular cells bind to the microorganism and trigger the union of plasmatocytes that surround the pathogen, forming a multicellular capsule where the pathogen is killed by the release of molecules by both types of cells.
The process of phagocytosis occurs in two main steps: first, the granular cells make contact with the pathogen, which causes the release of their granular content. This promotes the second step, which is adhesion of plasmatocytes to the pathogen to complete the process [23]. Once internalized, several killing mechanisms are induced, such as the production of superoxide, which leads to the destruction of the pathogen [24,25].
5. Humoral Immune Response
The humoral response comprises of many components, such as antimicrobial peptides (AMPs), lytic enzymes, opsonins, as well as molecules with direct activity against pathogens, such as phenoloxidase, which protects the host by catalyzing the formation of melanin.
5.1. Antimicrobials Peptides (AMPs)
The humoral immune response of insects is mainly based on the synthesis of antimicrobial peptides (AMPs). The contact with microorganisms induces the transcription of AMP-encoding genes. These molecules play a fundamental role in host defense and are produced mainly in the fat body, in both the digestive and reproductive tract, and subsequently are released into the hemolymph [26]. In some models, the induction of AMPs is transient, being very strong within the first 6 hours of infection but diminished after 3 days. AMPs are divided into cationic and anionic antimicrobial peptides. Cationic AMPs are basically classified into three groups based on their structure: (a) linear α-helical peptides without cysteine residues (among them, cepropins and moricins are active against bacteria and filamentous fungi [27,28,29]); (b) peptides with a structure stabilized with disulfide bridges, such as the cysteine-rich peptides (for example, gallerimycin and galiomycin, which are defensive peptides against fungi) [5,29,30]; (c) peptides with proline and/or glycine residues, such as Gm proline-rich peptide 1, with the ability to inhibit growth against yeast [31] and glycin-rich AMPs, such as gloverin, which inhibits the synthesis of membrane proteins in bacteria [5,31,32,33]. The anionic defense AMPs are activated against pathogens resistant to cationic peptides. In G. mellonella, two of these peptides, purified from the hemolymph, AP1 and AP2, have been described [31,34].
5.2. Lytic Enzymes (Lysozymes)
Lysozymes are an important element within the humoral response against pathogens found in the hemolymph of insects belonging to the orders Lepidoptera, Diptera, Coleoptera, and Hymenoptera [35,36,37,38]). Lysozyme is an antimicrobial enzyme that damages the bacterial cells by hydrolyzing β-1,4 linkages. Lysozyme also presents antifungal activity through enzymatic activity of the fungal cell walls, which results in growth inhibition [34,39,40,41].
5.3. Melanization
Melanin is a compound that is synthesized in response to foreign particles and plays a crucial role in sclerotization, wound healing, and in defense reactions. Melanin produces the accumulation of nodules, which can be visible in histological sections whose main function is to contain the replication of microorganisms [42,43]. The enzyme that catalyzes the synthesis of melanin is a phenoloxidase (PO) that oxidizes phenols to quinones, which subsequently polymerize non-enzymatically to melanin. Phenoloxidase is found in the hemolymph and hemocytes in its inactive form prophenoloxidase (PPO). PPO is activated by cell wall components of fungi and bacteria (lipopolysaccharides, peptidoglycans, and β-1,3-glucans) [44,45,46].
5.4. Opsonins
Galleria mellonella produces hemolymph proteins that act as opsonins. These proteins recognize components in the cell wall of different microorganisms, such as bacteria and fungi. Most of these molecules recognize and bind to lipopolysaccharides (LPS), peptidoglycan, and β-1,3-glucan [47,48,49,50]. apolipophorin-III (apoLp-III) is a protein that facilities lipid transport [51] and it has been associated with pathogen recognition and apoptosis [52]. Peptidoglycan recognition proteins (PGRPs) bind to bacterial peptidoglycan of the cell wall, causing the hydrolysis of the pathogens [5]. Hemolin is a member of the immunoglobulin superfamily. There are studies that have observed an increase in the production of hemolin after infection with bacteria and viruses [53,54].
6. Galleria mellonella as a Model to Study Fungal Pathogens
Invasive fungal diseases have become a major life threat to a large population of patients, mainly those that are immunosuppressed. Besides a few human fungal pathogens (such as C. albicans or C. neoformans), most of the fungal species present low virulence in mice. Therefore, the use of alternative models, and in particular, G. mellonella, has offered an alternative means in which to investigate the pathogenic mechanisms of most of the pathogenic fungal species. Recent studies have revealed that the innate immune response has great specificity, in addition to having the ability to distinguish between different classes of microorganisms [55]. Galleria mellonella was first described as a model for studying human fungal pathogen in the yeast C. albicans [9]. It is currently a model used to assess virulence of fungi such as A. fumigatus, Paracoccidioides lutzii, Histoplasma capsulatum, and C. neoformans [6,11,56,57,58]. In this last case, it has been even used to assess the virulence of a large collection of mutants [59]. Galleria mellonella model can also be used to assess toxicity and efficacy of antifungal agents during infection. The results obtained with this model have been shown to have a strong correlation with mammal models [60,61].
7. Response of G. mellonella to Different Fungi
7.1. Candida spp.
Candida spp. are the most common opportunistic fungal pathogen of humans, which can transform from superficial mucosal infections to systemic infections [62,63,64]. Bloodstream infections (BSIs) caused by Candida spp. remain a frequent cause of morbidity and mortality, particularly within the immunocompromised population [65,66,67]. The virulence of several Candida spp. (C. albicans, C. glabrata, C. tropicalis C. krusei, C. haemulonii, and C. auris) has been evaluated in G. mellonella [68,69,70,71,72,73].
The most common pathogen is C. albicans, and multiple studies have shown that this yeast can effectively kill G. mellonella. Both the cellular and humoral responses are important to control Candida infection. Different conditions that activate these responses protect against the infection by C. albicans. For example, physical stress, pre-incubation of the larvae at high (37 °C) or low (4 °C) temperature, or preexposure to non-lethal doses of C. albicans are conditions that increase the concentration of hemocytes as well as the expression of AMPs encoding genes. This has been shown to protect against later challenge with C. albicans [74,75,76]. Interestingly, some antifungal compounds that protect during Candida infection (such as caspofungin and micafungin) also have immunomodulatory effects and “boost” the immune response of G. mellonella during infection [14,74].
Inhibition of hemocyte function with cytochalasin and nocodazole enhances the susceptibility of the larvae to the infection [75]. After challenge with C. albicans, G. mellonella elicits activation of both the cellular and humoral responses. It has been shown that the number of circulating hemocytes fluctuate during the first hours of infection [76] but tend to decrease with time, in association with the severity of the disease [73]. Hemocytes induce nodulation at the site of C. albicans infection to avoid fungal replication. This nodulation is accompanied by the early accumulation of melanin at these sites [77].
Recognition of C. albicans by G. mellonella cells depends on several receptors and proteins. Base on current knowledge, one of the main receptors involved in this recognition is GmCD8 [77], which can also recognize other microbial pathogens. This receptor is not required for the formation of nodules by hemocytes and seems to act as an opsonin that directly induces phagocytosis of the fungal cells. Another protein that binds to the C. albicans surface is apolipophorin III [78], which is an apolipoprotein involved in lipid trafficking during insect flight [79]. Apolipophorin can exist in lipid-free or lipid-bound forms, being active when bound to lipophorin particles. In the case of C. albicans, apolipophorin III can bind to the surface of this pathogen and trigger the transition from yeast to hyphae, suggesting that this C. albicans can use some elements from the G. mellonella immune system to activate protective and adaptive responses. Furthermore, infection with C. albicans can alter the proportion of apoLp-III-free and apoLp-III-bound in the hemolymph [80], suggesting that they also participate in the recognition of fungal pathogens.
The main epitopes that are recognized by immune cells are located at the cell wall. In particular, β-glucans are the polysaccharides that elicit some of the strongest responses in mammalian cells [81]. In the case of G. mellonella, β-glucans can also activate some of the immune responses. Administration of β-glucan augments the density of hemocytes in the hemolymph and also increases the expression of some AMP-encoding genes [82]. This effect is in agreement with the finding that administration of β-glucan or laminarin can protect the larvae against lethal doses of C. albicans cells [83].
Elements of the humoral response are also involved in the defense against C. albicans. In particular, lysozymes can kill C. albicans [34]. Interestingly, this fungicidal effect is not directly related to degradation of the cell wall, but to induction of apoptosis in the yeast cells [84]. Another peptide that has been involved in the immune response against this pathogen is the anionic peptide 2 (AP2). However, this peptide does not kill the yeast cells, but has a fungistatic effect and, furthermore, enhances the killing activity of lysozyme [34].
7.2. Aspergillus fumigatus
Aspergillus fumigatus is the most common mold involved in human infections and the most isolated of the Aspergillus [56]. A. fumigatus produces allergic, chronic, and invasive disease depending upon the host immune system [85]. The G. mellonella model has been widely used in this pathogen for studies of virulence, survival, susceptibility, and antifungal resistance [56,57]. Killing of larvae by A. fumigatus is associated with massive replication of the fungus and filamentation. Furthermore, some toxins produced by A. fumigatus, such as fumagillin, can decrease the phagocytic and antifungal activity of hemocytes, which highlights another mechanism by which this mold can interfere with the larva immune response [86].
Infection of G. mellonella with high A. fumigatus doses results in dissemination of the fungus through the larva body after 24 h. However, G. mellonella induces early immune responses. For example, after 2 h of inoculation, there is an increase in the hemocyte density in the hemolymph [85]. Furthermore, fungal cells are contained in melanized nodules, as demonstrated with Candida spp. Proteomic analysis also indicate that A. fumigatus induces the accumulation of AMPs and immune receptors [85].
Interestingly, infection with non-lethal doses is enough to prime the larva immune response by inducing the expression of AMPs and the accumulation of hemocytes. This response is sufficient to protect against infection with higher and lethal doses of the fungus [87]. However, it has also been indicated that an exaggerated immune response might have negative effects for G. mellonella. In contrast to the situation in mice, A. fumigatus mutants that do not produce melanin have enhanced virulence in G. mellonella [88]. This phenotype is not due to an increased germination of the conidia. In contrast, A. fumigatus pigmentation mutants induce stronger darkening of the larvae after inoculation, suggesting that they overactivate the immune response of the insect. Furthermore, coinfection with wild type and pigmentation mutants results in a trend to increased virulence. Jackson et al. suggested that absence of melanin at the surface unmasks some fungal PAMPs (such as β-1,3-glucan), which triggers an excessive immune response that has deleterious effects on the larvae [88].
7.3. Cryptococcus neoformans
Cryptococcus neoformans is a basidiomycetes yeast widely distributed in the environment that can behave as a pathogen in susceptible patients [89,90]. Cryptococcus neoformans infects many people, yet few develop disease, most commonly cryptococcal meningoencephalitis. The infection produced by this pathogen occurs mainly in HIV patients. Cryptococcus has the capacity to adapt to several environmental hosts and evade the immune response through multiple mechanisms [91,92,93,94].
This pathogen has a polysaccharide capsule that plays multiple roles during infection. It protects the fungus against stress factors, but it can also act as a virulence factor and alter the host immune response [95]. Another interesting factor of this yeast is its ability to evade killing by phagocytic cells after phagocytosis, so it is considered a facultative intracellular pathogen [90,96,97,98]. In addition, Cryptococcus can also develop some typical morphological transitions that contribute to immune evasion. The most characteristic consist basically of the increase in the size of the capsule [95,99,100] and the formation of titan cells [101,102,103,104,105].
Cryptococcus neoformans can infect and kill larvae [11], and this insect has been frequently used to investigate virulence of clinical and environmental strains. The responses of C. neoformans in G. mellonella are similar to those observed in mice. For example, capsule growth and titan cells have been observed during G. mellonella infection [106]. The immune response of G. mellonella against this pathogen has been also investigated [6]. Interestingly, infection with C. neoformans does not result in early melanization at the infection sites, as it happens with Candida. This is most probably due to the presence of the capsule and to the different cell wall composition between these two fungal species. Galleria mellonella induces the accumulation of AMPs after cryptococcal challenge, which is activated in part by the presence of the capsule. Furthermore, C. neoformans cells are avidly phagocytosed by G. mellonella hemocytes and, interestingly, this fungal pathogen can survive and replicate after internalization, indicating that it can also behave as an intracellular fungal pathogen in G. mellonella hemocytes [6].
7.4. Other Fungal Pathogens
The virulence of other fungal pathogens in G. mellonella has been also studied. For example, Fusarium spp. (in particular, F. oxysporum and F. solani) are filamentous fungi that can also cause different diseases in immunocompromised patients. These fungi are widely found in the environment, where they can also behave as frequent plant pathogens. Fusarium oxysporum can cause disease in G. mellonella, and its virulence depends on the inoculum size and conidia germination and hypha proliferation [107]. Virulence in G. mellonella correlates with the pathogenicity in mice [108]. Furthermore, Fusarium macroconidia are more virulent than microconidia in G. mellonella, despite both forms being effectively phagocytosed by the hemocytes [108]. An elegant proteomic analysis using iTRAQ labeling revealed the changes that F. oxysporum induces in protein profile in G. mellonella. Munoz-Gomez et al. found that G. mellonella response to F. oxysporum is highly dynamic and dependent on the inoculum size (104 or 106 conidia per larva) and on the incubation temperature (25 or 37 °C ) [109].
Dimorphic fungi are clinically considered as those that present a filamentous morphology in the environment but behave as yeasts at 37 °C in the host. These fungi can cause disease in both immunocompromised and immunocompetent individuals. Main dimorphic fungi are Histoplasma capsulatum, Coccidioides immitis, Blastomyces dermatitidis, Paracoccidioides brasiliensis, and P. lutzii. Infection models of these fungi in mice can be very long (even months, see [110]), which poses a great limitation to assessing their virulence traits. For this reason, the G. mellonella model has been used to investigate the virulence of some of these fungi, such as H. capsulatum and P. lutzii [58,111,112]. Interestingly, the virulence of these two fungi in G. mellonella does not correlate with the inoculum size [58]. In contrast, both fungi induce early melanization of the larva in a dose-dependent manner. These authors also compared the virulence of two H. capsulatum strains that differ in their cell wall composition. In particular, the strain that lacks a layer of α-glucan on the surface that unmasks the β-glucan layer, has significantly reduced virulence, suggesting that activation of the G. mellonella immunity by the β-glucan component elicits a protective response against this pathogen (Figure 1).
8. Future Perspectives
Animal experimentation is nowadays subjected to strict laws and regulations. Furthermore, society has increasingly become concerned about the use of animals in research due to the pain derived from the experimental procedures. For these reasons, in the last decades, there has been a great interest to implement and expand the “3Rs” rule. The application of G. mellonella in biomedical research is becoming a useful model for investigating microbial pathogenesis. However, it is important to understand the limitations of this model too. For example, since this lepidopteran does not have adaptive immunity, it is not feasible to propose that G. mellonella can fully replace the use of other complex models, such as mice. However, the simplicity of the G. mellonella model to analyze microbial virulence and the efficacy of antibiotics and antifungals provides an excellent alternative to reduce the number of experimentation animals. The lack of genetic tools in G. mellonella poses another limitation to investigate the role of specific elements of the immune system. However, the recent elucidation of its genome will open new perspectives in the experimental approaches carried out with this insect.
Author Contributions
N.T.-C. and O.Z. have participated in experimental planning and writing of the manuscript.
Funding
O.Z. is funded by grants SAF2014-54336-R and SAF2017-86192-R1 from the former Spanish Ministry for Economics, Industry and Competitivity.
Acknowledgements
We want to thank Rachelle Babb for reading and helping us to edit this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Trevijano-Contador, N.; Zaragoza, O. Expanding the use of alternative models to investigate novel aspects of immunity to microbial pathogens. Virulence 2014, 5, 454–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilmos, P.; Kurucz, E. Insect immunity: Evolutionary roots of the mammalian innate immune system. Immunol. Lett. 1998, 62, 59–66. [Google Scholar] [CrossRef]
- Junqueira, J.C. Galleria mellonella as a model host for human pathogens: Recent studies and new perspectives. Virulence 2012, 3, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.B.; O’Brien, E.; Khoury, J.B.; Mylonakis, E. Methods for using Galleria mellonella as a model host to study fungal pathogenesis. Virulence 2010, 1, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.J.; Loh, J.M.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef] [PubMed]
- Trevijano-Contador, N.; Herrero-Fernandez, I.; Garcia-Barbazan, I.; Scorzoni, L.; Rueda, C.; Rossi, S.A.; Garcia-Rodas, R.; Zaragoza, O. Cryptococcus neoformans induces antimicrobial responses and behaves as a facultative intracellular pathogen in the non mammalian model Galleria mellonella. Virulence 2015, 6, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Reeves, E.P.; Messina, C.G.; Doyle, S.; Kavanagh, K. Correlation between gliotoxin production and virulence of Aspergillus fumigatus in Galleria mellonella. Mycopathologia 2004, 158, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.; Thomas, D.Y.; Whiteway, M.; Kavanagh, K. Correlation between virulence of Candida albicans mutants in mice and Galleria mellonella larvae. FEMS Immunol. Med. Microbiol. 2002, 34, 153–157. [Google Scholar] [CrossRef]
- Cotter, G.; Doyle, S.; Kavanagh, K. Development of an insect model for the in vivo pathogenicity testing of yeasts. FEMS Immunol. Med. Microbiol. 2000, 27, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Dunphy, G.B.; Oberholzer, U.; Whiteway, M.; Zakarian, R.J.; Boomer, I. Virulence of Candida albicans mutants toward larval Galleria mellonella (insecta, lepidoptera, galleridae). Can. J. Microbiol. 2003, 49, 514–524. [Google Scholar] [CrossRef]
- Mylonakis, E.; Moreno, R.; El Khoury, J.B.; Idnurm, A.; Heitman, J.; Calderwood, S.B.; Ausubel, F.M.; Diener, A. Galleria mellonella as a model system to study Cryptococcus neoformans pathogenesis. Infect. Immunity 2005, 73, 3842–3850. [Google Scholar] [CrossRef]
- Aperis, G.; Fuchs, B.B.; Anderson, C.A.; Warner, J.E.; Calderwood, S.B.; Mylonakis, E. Galleria mellonella as a model host to study infection by the francisella tularensis live vaccine strain. Microbes Infect. 2007, 9, 729–734. [Google Scholar] [CrossRef]
- Lange, A.; Beier, S.; Huson, D.H.; Parusel, R.; Iglauer, F.; Frick, J.S. Genome sequence of Galleria mellonella (greater wax moth). Genome Announc. 2018, 6, e01220-17. [Google Scholar] [CrossRef]
- Kelly, J.; Kavanagh, K. Caspofungin primes the immune response of the larvae of Galleria mellonella and induces a non-specific antimicrobial response. J. Med. Microbiol. 2011, 60, 189–196. [Google Scholar] [CrossRef]
- Maguire, R.; Duggan, O.; Kavanagh, K. Evaluation of Galleria mellonella larvae as an in vivo model for assessing the relative toxicity of food preservative agents. Cell Biol. Toxicol. 2016, 32, 209–216. [Google Scholar] [CrossRef]
- Kavanagh, K.; Reeves, E.P. Exploiting the potential of insects for in vivo pathogenicity testing of microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 101–112. [Google Scholar] [CrossRef] [Green Version]
- James, R.R.; Xu, J. Mechanisms by which pesticides affect insect immunity. J. Invertebr. Pathol. 2012, 109, 175–182. [Google Scholar] [CrossRef]
- Boman, H.G.; Hultmark, D. Cell-free immunity in insects. Annu. Rev. Microbiol. 1987, 41, 103–126. [Google Scholar] [CrossRef]
- Tojo, S.; Naganuma, F.; Arakawa, K.; Yokoo, S. Involvement of both granular cells and plasmatocytes in phagocytic reactions in the greater wax moth, Galleria mellonella. J. Insect. Physiol. 2000, 46, 1129–1135. [Google Scholar] [CrossRef]
- Browne, N.; Heelan, M.; Kavanagh, K. An analysis of the structural and functional similarities of insect hemocytes and mammalian phagocytes. Virulence 2013, 4, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Altincicek, B.; Stotzel, S.; Wygrecka, M.; Preissner, K.T.; Vilcinskas, A. Host-derived extracellular nucleic acids enhance innate immune responses, induce coagulation, and prolong survival upon infection in insects. J. Immunol. 2008, 181, 2705–2712. [Google Scholar] [CrossRef]
- Gillespie, J.P.; Kanost, M.R.; Trenczek, T. Biological mediators of insect immunity. Annu. Rev. Entomol. 1997, 42, 611–643. [Google Scholar] [CrossRef]
- Schmit, A.R.; Ratcliffe, N.A. The encapsulation of foreign tissue implants in Galleria mellonella larvae. J. Insect. Physiol. 1977, 23, 175–184. [Google Scholar] [CrossRef]
- Bergin, D.; Reeves, E.P.; Renwick, J.; Wientjes, F.B.; Kavanagh, K. Superoxide production in Galleria mellonella hemocytes: Identification of proteins homologous to the nadph oxidase complex of human neutrophils. Infect. Immunity 2005, 73, 4161–4170. [Google Scholar] [CrossRef]
- Renwick, J.; Reeves, E.P.; Wientjes, F.B.; Kavanagh, K. Translocation of proteins homologous to human neutrophil p47phox and p67phox to the cell membrane in activated hemocytes of Galleria mellonella. Dev. Comp. Immunol. 2007, 31, 347–359. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. The discovery and analysis of a diverged family of novel antifungal moricin-like peptides in the wax moth Galleria mellonella. Insect Biochem. Mol. Biol. 2008, 38, 201–212. [Google Scholar] [CrossRef]
- Kim, C.H.; Lee, J.H.; Kim, I.; Seo, S.J.; Son, S.M.; Lee, K.Y.; Lee, I.H. Purification and cdna cloning of a cecropin-like peptide from the great wax moth, Galleria mellonella. Mol. Cells 2004, 17, 262–266. [Google Scholar]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. A peptidomics study reveals the impressive antimicrobial peptide arsenal of the wax moth Galleria mellonella. Insect Biochem. Mol. Biol. 2009, 39, 792–800. [Google Scholar] [CrossRef]
- Langen, G.; Imani, J.; Altincicek, B.; Kieseritzky, G.; Kogel, K.H.; Vilcinskas, A. Transgenic expression of gallerimycin, a novel antifungal insect defensin from the greater wax moth Galleria mellonella, confers resistance to pathogenic fungi in tobacco. Biol. Chem. 2006, 387, 549–557. [Google Scholar] [CrossRef]
- Cytrynska, M.; Mak, P.; Zdybicka-Barabas, A.; Suder, P.; Jakubowicz, T. Purification and characterization of eight peptides from Galleria mellonella immune hemolymph. Peptides 2007, 28, 533–546. [Google Scholar] [CrossRef]
- Kawaoka, S.; Katsuma, S.; Daimon, T.; Isono, R.; Omuro, N.; Mita, K.; Shimada, T. Functional analysis of four gloverin-like genes in the silkworm, Bombyx mori. Arch. Insect Biochem. Physiol. 2008, 67, 87–96. [Google Scholar] [CrossRef]
- Bulet, P.; Hetru, C.; Dimarcq, J.L.; Hoffmann, D. Antimicrobial peptides in insects; structure and function. Dev. Comp. Immunol. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Sowa-Jasilek, A.; Zdybicka-Barabas, A.; Staczek, S.; Wydrych, J.; Mak, P.; Jakubowicz, T.; Cytrynska, M. Studies on the role of insect hemolymph polypeptides: Galleria mellonella anionic peptide 2 and lysozyme. Peptides 2014, 53, 194–201. [Google Scholar] [CrossRef]
- Lockey, T.D.; Ourth, D.D. Purification and characterization of lysozyme from hemolymph of heliothis virescens larvae. Biochem. Biophys. Res. Commun. 1996, 220, 502–508. [Google Scholar] [CrossRef]
- Mohrig, W.; Messner, B. Lysozyme as antibacterial agent in honey and bees venom. Acta Biol. Med. Ger. 1968, 21, 85–95. [Google Scholar]
- Vogel, H.; Altincicek, B.; Glockner, G.; Vilcinskas, A. A comprehensive transcriptome and immune-gene repertoire of the lepidopteran model host Galleria mellonella. BMC Genom. 2011, 12, 308. [Google Scholar] [CrossRef]
- Yu, K.H.; Kim, K.N.; Lee, J.H.; Lee, H.S.; Kim, S.H.; Cho, K.Y.; Nam, M.H.; Lee, I.H. Comparative study on characteristics of lysozymes from the hemolymph of three lepidopteran larvae, Galleria mellonella, Bombyx mori, Agrius convolvuli. Dev. Comp. Immunol. 2002, 26, 707–713. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, Y.Y.; Chang, J.Y.; Park, M.S.; Kho, H.S. The effects of peroxidase on the enzymatic and candidacidal activities of lysozyme. Arch. Oral Biol. 2010, 55, 607–612. [Google Scholar] [CrossRef]
- Nishiyama, Y.; Nakaoka, C.; Hiratani, T.; Abe, S.; Uchida, K.; Yamaguchi, H. Synergy of lysozyme and lanoconazole on the morphology of Candida albicans. J. Electron Microsc. 2001, 50, 41–49. [Google Scholar] [CrossRef]
- Wu, T.; Samaranayake, L.P.; Leung, W.K.; Sullivan, P.A. Inhibition of growth and secreted aspartyl proteinase production in Candida albicans by lysozyme. J. Med. Microbiol. 1999, 48, 721–730. [Google Scholar] [CrossRef]
- Ratcliffe, N.A.; Rowley, A.F.; Fitzgerald, S.W.; Rhodes, C.P. Invertebrate immunity: Basic concepts and recent advances. Int. Rev. Cytol. 1985, 97, 183–350. [Google Scholar]
- Götz, P. Encapsulation in arthropods. Immunity Invertebr. 1986, 153–170. [Google Scholar]
- Masaaki Ashida, H.Y. Limited proteolysis of prophenoloxidase during activation by microbial products in insect plasma and effect of phenoloxidase on electrophoretic mobilities of plasma proteins. J. Insect Biochem. 1988, 18, 11–19. [Google Scholar] [CrossRef]
- Soderhall, K. Prophenoloxidase-activating cascade as a recognition and defense system in arthropods. Immunity Invertebr. 1986, 208–223. [Google Scholar]
- Soderhall, K.S.; Smith, V.J. The prophenoloxidase activating system. The biochemistry of its activation and role in arthropod cellular immunity, with special reference to crustacenad. Immunity Arthropods 1986, 251–285. [Google Scholar]
- Yu, X.Q.; Kanost, M.R. Immulectin-2, a lipopolysaccharide-specific lectin from an insect, manduca sexta, is induced in response to gram-negative bacteria. J. Biol. Chem. 2000, 275, 37373–37381. [Google Scholar] [CrossRef]
- Yu, X.Q.; Kanost, M.R. Binding of hemolin to bacterial lipopolysaccharide and lipoteichoic acid. An immunoglobulin superfamily member from insects as a pattern-recognition receptor. Eur. J. Biochem. 2002, 269, 1827–1834. [Google Scholar] [CrossRef]
- Yoshida, H.; Kinoshita, K.; Ashida, M. Purification of a peptidoglycan recognition protein from hemolymph of the silkworm, Bombyx mori. J. Biol. Chem. 1996, 271, 13854–13860. [Google Scholar] [CrossRef]
- Wang, X.; Rocheleau, T.A.; Fuchs, J.F.; Christensen, B.M. Beta 1, 3-glucan recognition protein from the mosquito, armigeres subalbatus, is involved in the recognition of distinct types of bacteria in innate immune responses. Cell. Microbiol. 2006, 8, 1581–1590. [Google Scholar] [CrossRef]
- Blacklock, B.J.; Ryan, R.O. Hemolymph lipid transport. Insect Biochem. Mol. Biol. 1994, 24, 855–873. [Google Scholar] [CrossRef]
- Sun, D.; Ziegler, R.; Milligan, C.E.; Fahrbach, S.; Schwartz, L.M. Apolipophorin iii is dramatically up-regulated during the programmed death of insect skeletal muscle and neurons. J. Neurobiol. 1995, 26, 119–129. [Google Scholar] [CrossRef]
- Ladendorff, N.E.; Kanost, M.R. Isolation and characterization of bacteria-induced protein p4 from hemolymph of manduca sexta. Arch. Insect Biochem. Physiol. 1990, 15, 33–41. [Google Scholar] [CrossRef]
- Terenius, O. Hemolin-a lepidopteran anti-viral defense factor? Dev. Comp. Immunol. 2008, 32, 311–316. [Google Scholar] [CrossRef]
- Hoffmann, J.A. The immune response of drosophila. Nature 2003, 426, 33–38. [Google Scholar] [CrossRef]
- Alcazar-Fuoli, L.; Buitrago, M.; Gomez-Lopez, A.; Mellado, E. An alternative host model of a mixed fungal infection by azole susceptible and resistant aspergillus spp strains. Virulence 2015, 6, 376–384. [Google Scholar] [CrossRef]
- Gomez-Lopez, A.; Forastiero, A.; Cendejas-Bueno, E.; Gregson, L.; Mellado, E.; Howard, S.J.; Livermore, J.L.; Hope, W.W.; Cuenca-Estrella, M. An invertebrate model to evaluate virulence in Aspergillus fumigatus: The role of azole resistance. Med. Mycol. 2014, 52, 311–319. [Google Scholar] [CrossRef]
- Thomaz, L.; Garcia-Rodas, R.; Guimaraes, A.J.; Taborda, C.P.; Zaragoza, O.; Nosanchuk, J.D. Galleria mellonella as a model host to study Paracoccidioides lutzii and histoplasma capsulatum. Virulence 2013, 4, 139–146. [Google Scholar] [CrossRef]
- Desalermos, A.; Tan, X.; Rajamuthiah, R.; Arvanitis, M.; Wang, Y.; Li, D.; Kourkoumpetis, T.K.; Fuchs, B.B.; Mylonakis, E. A multi-host approach for the systematic analysis of virulence factors in Cryptococcus neoformans. J. Infect. Dis. 2015, 211, 298–305. [Google Scholar] [CrossRef]
- Slater, J.L.; Gregson, L.; Denning, D.W.; Warn, P.A. Pathogenicity of Aspergillus fumigatus mutants assessed in Galleria mellonella matches that in mice. Med. Mycol. 2011, 49 (Suppl. 1), S107–S113. [Google Scholar] [CrossRef]
- Halldorsdottir, A.M.; Carayannopoulos, M.O.; Scrivner, M.; Gronowski, A.M. Method evaluation for total beta-human chorionic gonadotropin using urine and the advia centaur. Clin. Chem. 2003, 49, 1421–1422. [Google Scholar] [CrossRef]
- Fidel, P.L., Jr. Candida-host interactions in hiv disease: Implications for oropharyngeal candidiasis. Adv. Dent. Res. 2011, 23, 45–49. [Google Scholar] [CrossRef]
- Benjamin, D.K., Jr.; Garges, H.; Steinbach, W.J. Candida bloodstream infection in neonates. Semin. Perinatol. 2003, 27, 375–383. [Google Scholar] [CrossRef]
- Li, D.D.; Deng, L.; Hu, G.H.; Zhao, L.X.; Hu, D.D.; Jiang, Y.Y.; Wang, Y. Using Galleria mellonella—Candida albicans infection model to evaluate antifungal agents. Biol. Pharm. Bull. 2013, 36, 1482–1487. [Google Scholar] [CrossRef]
- Sherry, L.; Rajendran, R.; Lappin, D.F.; Borghi, E.; Perdoni, F.; Falleni, M.; Tosi, D.; Smith, K.; Williams, C.; Jones, B.; et al. Biofilms formed by Candida albicans bloodstream isolates display phenotypic and transcriptional heterogeneity that are associated with resistance and pathogenicity. BMC Microbiol. 2014, 14, 182. [Google Scholar] [CrossRef]
- Mensa, J.; Pitart, C.; Marco, F. Treatment of critically ill patients with candidemia. Int. J. Antimicrob. Agents 2008, 32 (Suppl. 2), S93–S97. [Google Scholar] [CrossRef]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Nosocomial bloodstream infections in us hospitals: Analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef]
- Santos, R.B.; Scorzoni, L.; Namba, A.M.; Rossoni, R.D.; Jorge, A.O.C.; Junqueira, J.C. Lactobacillus species increase the survival of Galleria mellonella infected with Candida albicans and non-albicans Candida clinical isolates. Med. Mycol. 2018. [Google Scholar] [CrossRef]
- Silva, L.N.; Campos-Silva, R.; Ramos, L.S.; Trentin, D.S.; Macedo, A.J.; Branquinha, M.H.; Santos, A.L.S. Virulence of Candida haemulonii complex in Galleria mellonella and efficacy of classical antifungal drugs: A comparative study with other clinically relevant non-albicans Candida species. FEMS Yeast Res. 2018, 18, foy082. [Google Scholar] [CrossRef]
- Moralez, A.T.; Perini, H.F.; Furlaneto-Maia, L.; Almeida, R.S.; Panagio, L.A.; Furlaneto, M.C. Phenotypic switching of Candida tropicalis is associated with cell damage in epithelial cells and virulence in Galleria mellonella model. Virulence 2016, 7, 379–386. [Google Scholar] [CrossRef]
- Mesa-Arango, A.C.; Forastiero, A.; Bernal-Martinez, L.; Cuenca-Estrella, M.; Mellado, E.; Zaragoza, O. The non-mammalian host Galleria mellonella can be used to study the virulence of the fungal pathogen Candida tropicalis and the efficacy of antifungal drugs during infection by this pathogenic yeast. Med. Mycol. 2012, 51, 461–472. [Google Scholar] [CrossRef]
- Ames, L.; Duxbury, S.; Pawlowska, B.; Ho, H.L.; Haynes, K.; Bates, S. Galleria mellonella as a host model to study candida glabrata virulence and antifungal efficacy. Virulence 2017, 8, 1909–1917. [Google Scholar] [CrossRef]
- Scorzoni, L.; de Lucas, M.P.; Mesa-Arango, A.C.; Fusco-Almeida, A.M.; Lozano, E.; Cuenca-Estrella, M.; Mendes-Giannini, M.J.; Zaragoza, O. Antifungal efficacy during Candida krusei infection in non-conventional models correlates with the yeast in vitro susceptibility profile. PLoS ONE 2013, 8, e60047. [Google Scholar] [CrossRef]
- Fuchs, B.B.; Li, Y.; Li, D.; Johnston, T.; Hendricks, G.; Li, G.; Rajamuthiah, R.; Mylonakis, E. Micafungin elicits an immunomodulatory effect in Galleria mellonella and mice. Mycopathologia 2016, 181, 17–25. [Google Scholar] [CrossRef]
- Banville, N.; Fallon, J.; McLoughlin, K.; Kavanagh, K. Disruption of haemocyte function by exposure to cytochalasin b or nocodazole increases the susceptibility of Galleria mellonella larvae to infection. Microbes Infect. 2011, 13, 1191–1198. [Google Scholar] [CrossRef]
- Sheehan, G.; Kavanagh, K. Analysis of the early cellular and humoral responses of Galleria mellonella larvae to infection by Candida albicans. Virulence 2018, 9, 163–172. [Google Scholar] [CrossRef]
- Kim, C.H.; Shin, Y.P.; Noh, M.Y.; Jo, Y.H.; Han, Y.S.; Seong, Y.S.; Lee, I.H. An insect multiligand recognition protein functions as an opsonin for the phagocytosis of microorganisms. J. Biol. Chem. 2010, 285, 25243–25250. [Google Scholar] [CrossRef]
- Zdybicka-Barabas, A.; Staczek, S.; Mak, P.; Piersiak, T.; Skrzypiec, K.; Cytrynska, M. The effect of Galleria mellonella apolipophorin iii on yeasts and filamentous fungi. J. Insect. Physiol. 2012, 58, 164–177. [Google Scholar] [CrossRef]
- Weers, P.M.; Ryan, R.O. Apolipophorin iii: A lipid-triggered molecular switch. Insect Biochem. Mol. Biol. 2003, 33, 1249–1260. [Google Scholar] [CrossRef]
- Staczek, S.; Zdybicka-Barabas, A.; Mak, P.; Sowa-Jasilek, A.; Kedracka-Krok, S.; Jankowska, U.; Suder, P.; Wydrych, J.; Grygorczuk, K.; Jakubowicz, T.; et al. Studies on localization and protein ligands of Galleria mellonella apolipophorin iii during immune response against different pathogens. J. Insect. Physiol. 2018, 105, 18–27. [Google Scholar] [CrossRef]
- Camilli, G.; Tabouret, G.; Quintin, J. The complexity of fungal beta-glucan in health and disease: Effects on the mononuclear phagocyte system. Front. Immunol. 2018, 9, 673. [Google Scholar] [CrossRef]
- Mowlds, P.; Coates, C.; Renwick, J.; Kavanagh, K. Dose-dependent cellular and humoral responses in Galleria mellonella larvae following beta-glucan inoculation. Microbes Infect. 2010, 12, 146–153. [Google Scholar] [CrossRef]
- Bergin, D.; Murphy, L.; Keenan, J.; Clynes, M.; Kavanagh, K. Pre-exposure to yeast protects larvae of Galleria mellonella from a subsequent lethal infection by Candida albicans and is mediated by the increased expression of antimicrobial peptides. Microbes Infect. 2006, 8, 2105–2112. [Google Scholar] [CrossRef]
- Sowa-Jasilek, A.; Zdybicka-Barabas, A.; Staczek, S.; Wydrych, J.; Skrzypiec, K.; Mak, P.; Derylo, K.; Tchorzewski, M.; Cytrynska, M. Galleria mellonella lysozyme induces apoptotic changes in Candida albicans cells. Microbiol. Res. 2016, 193, 121–131. [Google Scholar] [CrossRef]
- Sheehan, G.; Clarke, G.; Kavanagh, K. Characterisation of the cellular and proteomic response of Galleria mellonella larvae to the development of invasive aspergillosis. BMC Microbiol. 2018, 18, 63. [Google Scholar] [CrossRef]
- Fallon, J.P.; Reeves, E.P.; Kavanagh, K. The Aspergillus fumigatus toxin fumagillin suppresses the immune response of Galleria mellonella larvae by inhibiting the action of haemocytes. Microbiology 2011, 157, 1481–1488. [Google Scholar] [CrossRef]
- Fallon, J.P.; Troy, N.; Kavanagh, K. Pre-exposure of Galleria mellonella larvae to different doses of Aspergillus fumigatus conidia causes differential activation of cellular and humoral immune responses. Virulence 2011, 2, 413–421. [Google Scholar] [CrossRef]
- Jackson, J.C.; Higgins, L.A.; Lin, X. Conidiation color mutants of Aspergillus fumigatus are highly pathogenic to the heterologous insect host Galleria mellonella. PLoS ONE 2009, 4, e4224. [Google Scholar] [CrossRef]
- Lazera, M.S.; Salmito Cavalcanti, M.A.; Londero, A.T.; Trilles, L.; Nishikawa, M.M.; Wanke, B. Possible primary ecological niche of Cryptococcus neoformans. Med. Mycol. 2000, 38, 379–383. [Google Scholar] [CrossRef]
- Casadevall, A.; Perfect, J. Cryptococcus Neoformans; ASM: Washington, DC, USA, 1998; p. 541. [Google Scholar]
- Venn-Watson, S.; Daniels, R.; Smith, C. Thirty year retrospective evaluation of pneumonia in a bottlenose dolphin Tursiops truncatus population. Dis. Aquat. Organ. 2012, 99, 237–242. [Google Scholar] [CrossRef]
- Warpeha, K.M.; Park, Y.D.; Williamson, P.R. Susceptibility of intact germinating arabidopsis thaliana to human fungal pathogens Cryptococcus neoformans and C. gattii. Appl. Environ. Microbiol. 2013, 79, 2979–2988. [Google Scholar] [CrossRef]
- Steenbergen, J.N.; Shuman, H.A.; Casadevall, A. Cryptococcus neoformans interactions with amoebae suggest an explanation for its virulence and intracellular pathogenic strategy in macrophages. Proc. Natl. Acad. Sci. USA 2001, 98, 15245–15250. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.M.; Huang, M.; Botts, M.R.; Hull, C.M.; Huttenlocher, A. A zebrafish model of cryptococcal infection reveals roles for macrophages, endothelial cells, and neutrophils in the establishment and control of sustained fungemia. Infect. Immunity 2016, 84, 3047–3062. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza, O.; Rodrigues, M.L.; De Jesus, M.; Frases, S.; Dadachova, E.; Casadevall, A. The capsule of the fungal pathogen Cryptococcus neoformans. Adv. Appl. Microbiol. 2009, 68, 133–216. [Google Scholar] [PubMed]
- Heitman, J.; Kozel, T.R.; Kwon-Chung, K.J.; Perferct, J.R.; Casadevall, A. Cryptococcus. From Human Pathogen to Model Yeast; ASM Press: Washington, DC, USA, 2011. [Google Scholar]
- Smith, L.M.; Dixon, E.F.; May, R.C. The fungal pathogen Cryptococcus neoformans manipulates macrophage phagosome maturation. Cell. Microbiol. 2015, 17, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Tucker, S.C.; Casadevall, A. Replication of Cryptococcus neoformans in macrophages is accompanied by phagosomal permeabilization and accumulation of vesicles containing polysaccharide in the cytoplasm. Proc. Natl. Acad. Sci. USA 2002, 99, 3165–3170. [Google Scholar] [CrossRef] [PubMed]
- Doering, T.L. How sweet it is! Cell wall biogenesis and polysaccharide capsule formation in Cryptococcus neoformans. Annu. Rev. Microbiol. 2009, 63, 223–247. [Google Scholar] [CrossRef]
- O’Meara, T.R.; Alspaugh, J.A. The Cryptococcus neoformans capsule: A sword and a shield. Clin. Microbiol. Rev. 2012, 25, 387–408. [Google Scholar] [CrossRef]
- Feldmesser, M.; Kress, Y.; Casadevall, A. Dynamic changes in the morphology of Cryptococcus neoformans during murine pulmonary infection. Microbiology 2001, 147, 2355–2365. [Google Scholar] [CrossRef]
- Okagaki, L.H.; Strain, A.K.; Nielsen, J.N.; Charlier, C.; Baltes, N.J.; Chretien, F.; Heitman, J.; Dromer, F.; Nielsen, K. Cryptococcal cell morphology affects host cell interactions and pathogenicity. PLoS Pathog. 2010, 6, e1000953. [Google Scholar] [CrossRef]
- Zaragoza, O.; Garcia-Rodas, R.; Nosanchuk, J.D.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L.; Casadevall, A. Fungal cell gigantism during mammalian infection. PLoS Pathog. 2010, 6, e1000945. [Google Scholar] [CrossRef]
- Zaragoza, O.; Nielsen, K. Titan cells in Cryptococcus neoformans: Cells with a giant impact. Curr. Opin. Microbiol. 2013, 16, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Trevijano-Contador, N.; de Oliveira, H.C.; Garcia-Rodas, R.; Rossi, S.A.; Llorente, I.; Zaballos, A.; Janbon, G.; Arino, J.; Zaragoza, O. Cryptococcus neoformans can form titan-like cells in vitro in response to multiple signals. PLoS Pathog. 2018, 14, e1007007. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rodas, R.; Casadevall, A.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M.; Zaragoza, O. Cryptococcus neoformans capsular enlargement and cellular gigantism during Galleria mellonella infection. PLoS ONE 2011, 6, e24485. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Velasco, G.Y.; Prados-Rosales, R.C.; Ortiz-Urquiza, A.; Quesada-Moraga, E.; Di Pietro, A. Galleria mellonella as model host for the trans-kingdom pathogen fusarium oxysporum. Fungal Genet. Biol. 2011, 48, 1124–1129. [Google Scholar] [CrossRef]
- Coleman, J.J.; Muhammed, M.; Kasperkovitz, P.V.; Vyas, J.M.; Mylonakis, E. Fusarium pathogenesis investigated using Galleria mellonella as a heterologous host. Fungal Biol. 2011, 115, 1279–1289. [Google Scholar] [CrossRef]
- Munoz-Gomez, A.; Corredor, M.; Benitez-Paez, A.; Pelaez, C. Development of quantitative proteomics using itraq based on the immunological response of Galleria mellonella larvae challenged with fusarium oxysporum microconidia. PLoS ONE 2014, 9, e112179. [Google Scholar] [CrossRef]
- Kashino, S.S.; Singer-Vermes, L.M.; Calich, V.L.; Burger, E. Alterations in the pathogenicity of one Paracoccidioides brasiliensis isolate do not correlative with its in vitro growth. Mycopathologia 1990, 111, 173–180. [Google Scholar] [CrossRef]
- de Oliveira, H.C.; da Silva Jde, F.; Scorzoni, L.; Marcos, C.M.; Rossi, S.A.; de Paula, E.S.A.C.; Assato, P.A.; da Silva, R.A.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J. Importance of adhesins in virulence of paracoccidioides spp. Front Microbiol. 2015, 6, 303. [Google Scholar] [CrossRef]
- Scorzoni, L.; de Paula e Silva, A.C.; Singulani Jde, L.; Leite, F.S.; de Oliveira, H.C.; da Silva, R.A.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J. Comparison of virulence between Paracoccidioides brasiliensis and Paracoccidioides lutzii using Galleria mellonella as a host model. Virulence 2015, 6, 766–776. [Google Scholar] [CrossRef]
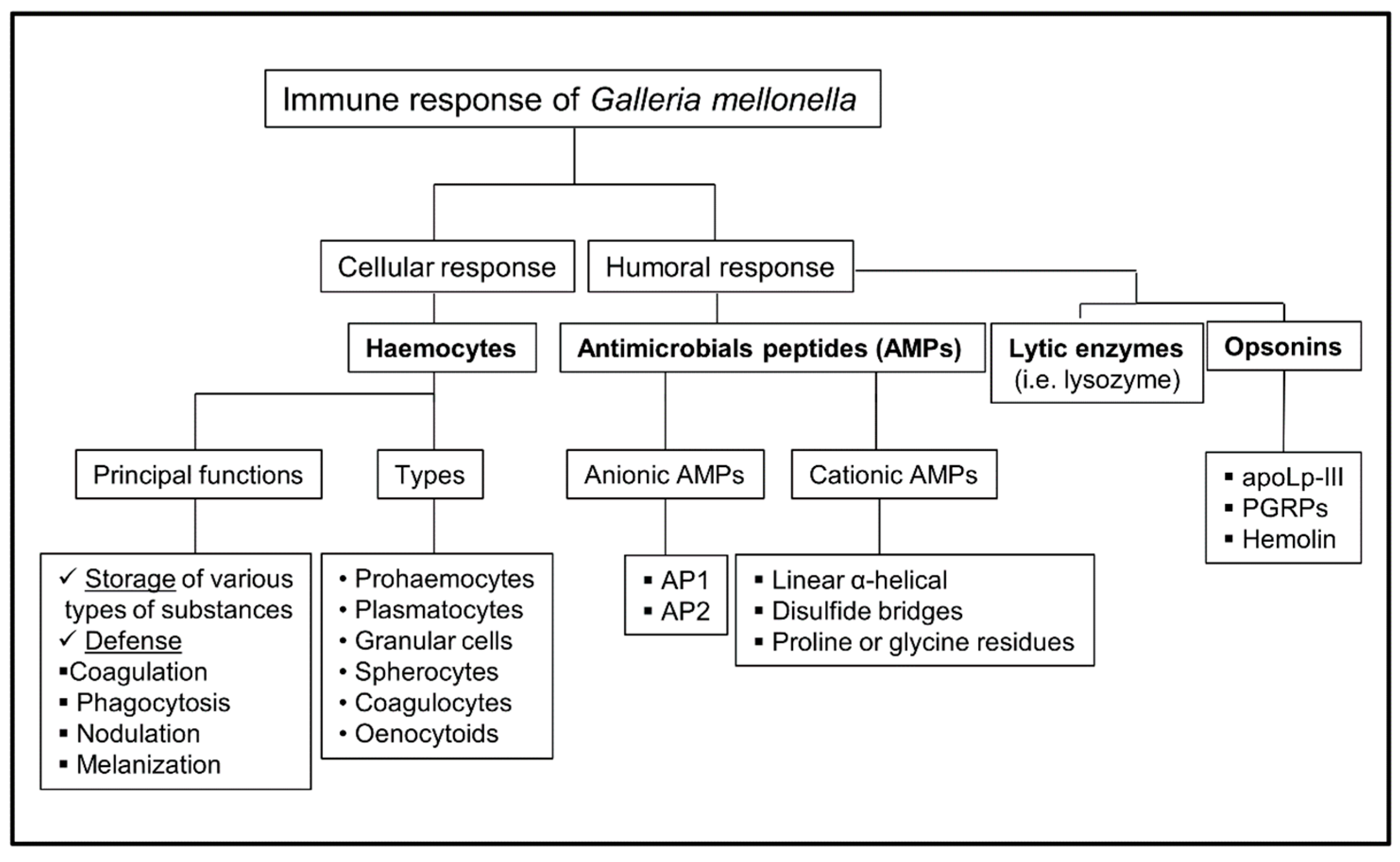
Figure 1.
Scheme of the immune response of Galleria mellonella. PGRPs: Peptidoglycan recognition proteins; AMPs: antimicrobial peptides.
Figure 1.
Scheme of the immune response of Galleria mellonella. PGRPs: Peptidoglycan recognition proteins; AMPs: antimicrobial peptides.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Trevijano-Contador, N.; Zaragoza, O. Immune Response of Galleria mellonella against Human Fungal Pathogens. J. Fungi 2019, 5, 3. https://doi.org/10.3390/jof5010003
AMA Style
Trevijano-Contador N, Zaragoza O. Immune Response of Galleria mellonella against Human Fungal Pathogens. Journal of Fungi. 2019; 5(1):3. https://doi.org/10.3390/jof5010003
Chicago/Turabian StyleTrevijano-Contador, Nuria, and Oscar Zaragoza. 2019. "Immune Response of Galleria mellonella against Human Fungal Pathogens" Journal of Fungi 5, no. 1: 3. https://doi.org/10.3390/jof5010003
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.