Immunomodulation as a Therapy for Aspergillus Infection: Current Status and Future Perspectives


{kind=link}
{kind=link}
Abstract
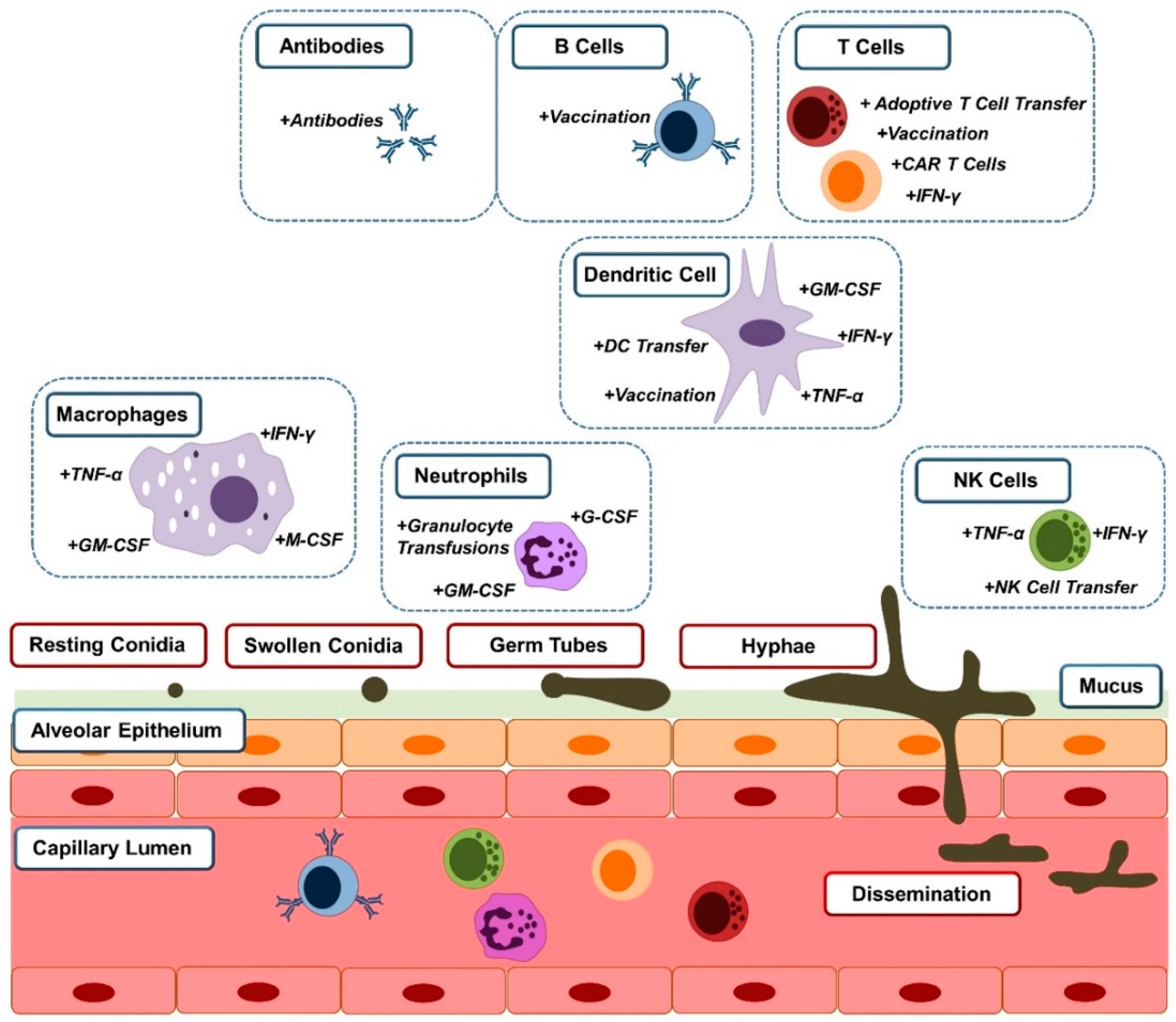
:1. Introduction
2. Cell Therapy
2.1. Granulocyte Transfusion
2.2. Dendritic Cells (DCs)
2.3. Natural Killer Cell Therapy
2.4. Adoptive T Cell Transfer
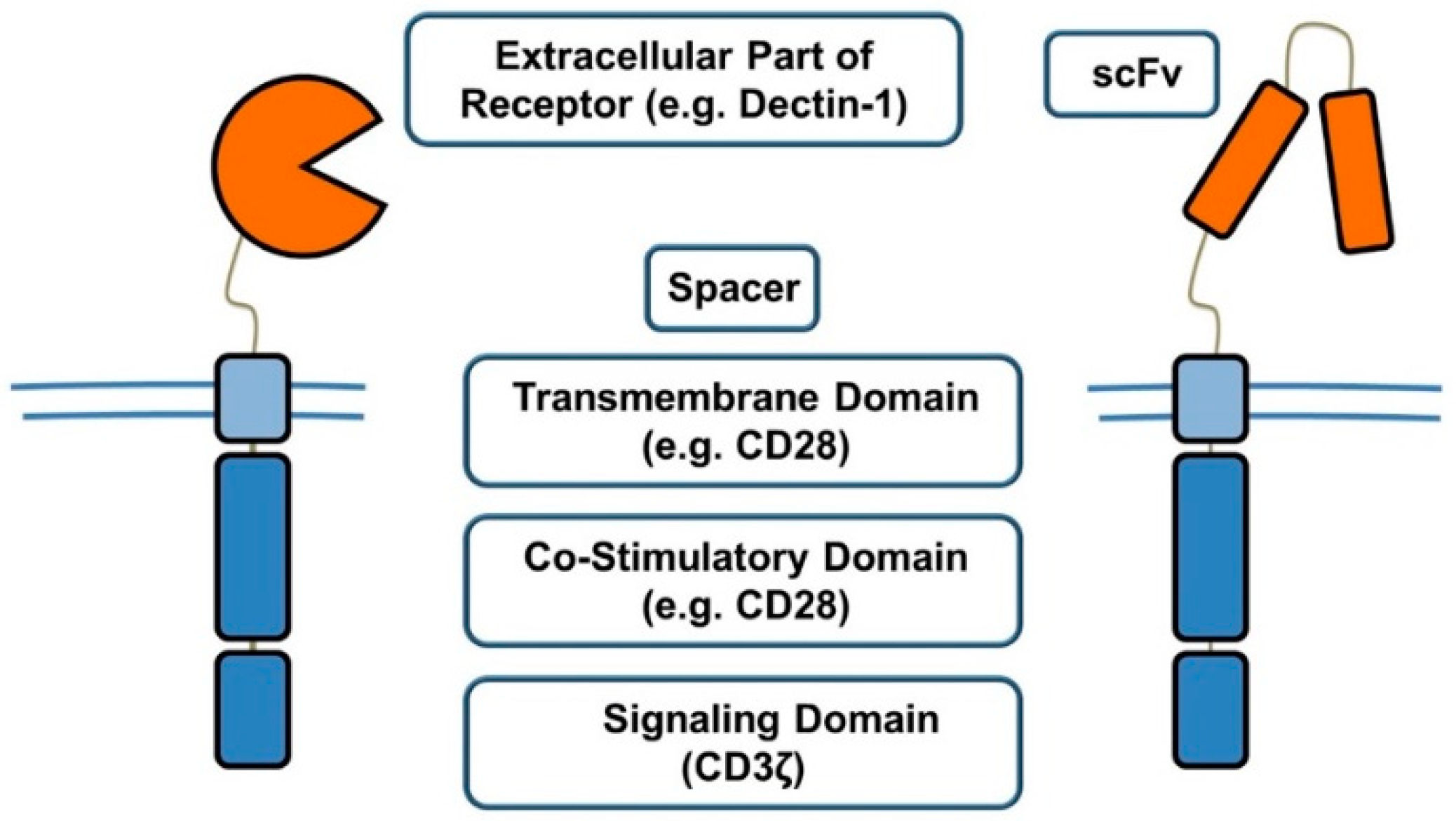
2.5. Chimeric Antigen Receptors (CARs)
3. Cytokine Therapy
3.1. Colony Stimulating Factors
3.1.1. G-CSF
3.1.2. M-CSF
3.1.3. GM-CSF
3.2. IFN-γ
3.3. TNF-α
4. Other Immunotherapeutic Approaches
4.1. Vaccination
4.2. Antibodies
5. Summary and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taccone, F.S.; Van den Abeele, A.M.; Bulpa, P.; Misset, B.; Meersseman, W.; Cardoso, T.; Paiva, J.A.; Blasco-Navalpotro, M.; De Laere, E.; Dimopoulos, G.; et al. Epidemiology of invasive aspergillosis in critically ill patients: Clinical presentation, underlying conditions, and outcomes. Crit. Care 2015, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Filler, S.G.; Sheppard, D.C. Fungal invasion of normally non-phagocytic host cells. PLoS Pathog. 2006, 2, e129. [Google Scholar] [CrossRef] [PubMed]
- White, P.L.; Posso, R.B.; Barnes, R.A. Analytical and Clinical Evaluation of the PathoNostics AsperGenius Assay for Detection of Invasive Aspergillosis and Resistance to Azole Antifungal Drugs Directly from Plasma Samples. J. Clin. Microbiol. 2017, 55, 2356–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohl, T.M. Immune responses to invasive aspergillosis: New understanding and therapeutic opportunities. Curr. Opin. Infect. Dis. 2017, 30, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Aimanianda, V.; Bayry, J.; Bozza, S.; Kniemeyer, O.; Perruccio, K.; Elluru, S.R.; Clavaud, C.; Paris, S.; Brakhage, A.A.; Kaveri, S.V.; et al. Surface hydrophobin prevents immune recognition of airborne fungal spores. Nature 2009, 460, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Bidula, S.; Schelenz, S. A Sweet Response to a Sour Situation: The Role of Soluble Pattern Recognition Receptors in the Innate Immune Response to Invasive Aspergillus fumigatus Infections. PLoS Pathog. 2016, 12, e1005637. [Google Scholar] [CrossRef]
- Garth, J.M.; Reeder, K.M.; Godwin, M.S.; Mackel, J.J.; Dunaway, C.W.; Blackburn, J.P.; Steele, C. IL-33 Signaling Regulates Innate IL-17A and IL-22 Production via Suppression of Prostaglandin E2 during Lung Fungal Infection. J. Immunol. 2017. [Google Scholar] [CrossRef]
- Espinosa, V.; Rivera, A. First line of defense: Innate cell-mediated control of pulmonary aspergillosis. Front. Microbiol. 2016, 7, 272. [Google Scholar] [CrossRef]
- Khanna, N.; Stuehler, C.; Lünemann, A.; Wójtowicz, A.; Bochud, P.Y.; Leibundgut-Landmann, S. Host response to fungal infections—How immunology and host genetics could help to identify and treat patients at risk. Swiss Med. Wkly. 2016, 146. [Google Scholar] [CrossRef]
- Heinz, W.J.; Vehreschild, J.J.; Buchheidt, D. Diagnostic workout to assess early response indicators in invasive pulmonary aspergillosis in adult patients with hematologic malignancies. Mycoses 2018. [Google Scholar] [CrossRef]
- Leroux, S.; Ullmann, A.J. Management and diagnostic guidelines for fungal diseases in infectious diseases and clinical microbiology: Critical appraisal. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2013, 19, 1115–1121. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-Resistant Aspergillosis: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S436–S444. [Google Scholar] [CrossRef] [PubMed]
- Hickey, M.J.; Kubes, P. Intravascular immunity: The host-pathogen encounter in blood vessels. Nat. Rev. Immunol. 2009, 9, 364–375. [Google Scholar] [CrossRef]
- Shoham, S.; Levitz, S.M. The immune response to fungal infections. Br. J. Haematol. 2005, 129, 569–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubel, K.; Carter, R.A.; Liles, W.C.; Dale, D.C.; Price, T.H.; Bowden, R.A.; Rowley, S.D.; Chauncey, T.R.; Bensinger, W.I.; Boeckh, M. Granulocyte transfusion therapy for infections in candidates and recipients of HPC transplantation: A comparative analysis of feasibility and outcome for community donors versus related donors. Transfusion 2002, 42, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Price, T.H.; Bowden, R.A.; Boeckh, M.; Bux, J.; Nelson, K.; Liles, W.C.; Dale, D.C. Phase I/II trial of neutrophil transfusions from donors stimulated with G-CSF and dexamethasone for treatment of patients with infections in hematopoietic stem cell transplantation. Blood 2000, 95, 3302–3309. [Google Scholar] [PubMed]
- Seidel, M.G.; Peters, C.; Wacker, A.; Northoff, H.; Moog, R.; Boehme, A.; Silling, G.; Grimminger, W.; Einsele, H. Randomized phase III study of granulocyte transfusions in neutropenic patients. Bone Marrow Transplant. 2008, 42, 679–684. [Google Scholar] [CrossRef] [Green Version]
- Mousset, S.; Hermann, S.; Klein, S.A.; Bialleck, H.; Duchscherer, M.; Bomke, B.; Wassmann, B.; Bohme, A.; Hoelzer, D.; Martin, H. Prophylactic and interventional granulocyte transfusions in patients with haematological malignancies and life-threatening infections during neutropenia. Ann. Hematol. 2005, 84, 734–741. [Google Scholar] [CrossRef]
- Price, T.H.; Boeckh, M.; Harrison, R.W.; McCullough, J.; Ness, P.M.; Strauss, R.G.; Nichols, W.G.; Hamza, T.H.; Cushing, M.M.; King, K.E.; et al. Efficacy of transfusion with granulocytes from G-CSF/dexamethasone-treated donors in neutropenic patients with infection. Blood 2015, 126, 2153–2161. [Google Scholar] [CrossRef] [Green Version]
- BitMansour, A.; Burns, S.M.; Traver, D.; Akashi, K.; Contag, C.H.; Weissman, I.L.; Brown, J.M. Myeloid progenitors protect against invasive aspergillosis and Pseudomonas aeruginosa infection following hematopoietic stem cell transplantation. Blood 2002, 100, 4660–4667. [Google Scholar] [CrossRef] [Green Version]
- BitMansour, A.; Cao, T.M.; Chao, S.; Shashidhar, S.; Brown, J.M. Single infusion of myeloid progenitors reduces death from Aspergillus fumigatus following chemotherapy-induced neutropenia. Blood 2005, 105, 3535–3537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, T.F.; Thompson, G.R., 3rd; Denning, D.W.; Fishman, J.A.; Hadley, S.; Herbrecht, R.; Kontoyiannis, D.P.; Marr, K.A.; Morrison, V.A.; Nguyen, M.H.; et al. Practice Guidelines for the Diagnosis and Management of Aspergillosis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 63, e1–e60. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Ortiz, Z.G.; Means, T.K. The role of dendritic cells in the innate recognition of pathogenic fungi (A. fumigatus, C. neoformans and C. albicans). Virulence 2012, 3, 635–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, R.; Anand, R.; Tiwari, S.; Singh, A.P.; Tiwary, B.N.; Shankar, J. Cytokines induce effector T-helper cells during invasive aspergillosis; what we have learned about T-helper cells? Front. Microbiol. 2015, 6, 429. [Google Scholar] [CrossRef] [PubMed]
- Zelante, T.; Wong, A.Y.; Ping, T.J.; Chen, J.; Sumatoh, H.R.; Vigano, E.; Hong Bing, Y.; Lee, B.; Zolezzi, F.; Fric, J.; et al. CD103(+) Dendritic Cells Control Th17 Cell Function in the Lung. Cell Rep. 2015, 12, 1789–1801. [Google Scholar] [CrossRef] [PubMed]
- Bozza, S.; Gaziano, R.; Lipford, G.B.; Montagnoli, C.; Bacci, A.; Di Francesco, P.; Kurup, V.P.; Wagner, H.; Romani, L. Vaccination of mice against invasive aspergillosis with recombinant Aspergillus proteins and CpG oligodeoxynucleotides as adjuvants. Microbes Infect. 2002, 4, 1281–1290. [Google Scholar] [CrossRef]
- Bozza, S.; Perruccio, K.; Montagnoli, C.; Gaziano, R.; Bellocchio, S.; Burchielli, E.; Nkwanyuo, G.; Pitzurra, L.; Velardi, A.; Romani, L. A dendritic cell vaccine against invasive aspergillosis in allogeneic hematopoietic transplantation. Blood 2003, 102, 3807–3814. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.J.; Chen, L.; Su, Y.B. DNA fragment encoding human IL-1beta 163-171 peptide enhances the immune responses elicited in mice by DNA vaccine against foot-and-mouth disease. Vet. Res. Commun. 2005, 29, 35–46. [Google Scholar] [CrossRef]
- Zhu, F.; Ramadan, G.; Davies, B.; Margolis, D.A.; Keever-Taylor, C.A. Stimulation by means of dendritic cells followed by Epstein-Barr virus-transformed B cells as antigen-presenting cells is more efficient than dendritic cells alone in inducing Aspergillus f16-specific cytotoxic T cell responses. Clin. Exp. Immunol. 2008, 151, 284–296. [Google Scholar] [CrossRef]
- Roy, R.M.; Klein, B.S. Dendritic cells in antifungal immunity and vaccine design. Cell Host Microbe 2012, 11, 436–446. [Google Scholar] [CrossRef]
- Schmidt, S.; Tramsen, L.; Perkhofer, S.; Lass-Florl, C.; Hanisch, M.; Roger, F.; Klingebiel, T.; Koehl, U.; Lehrnbecher, T. Rhizopus oryzae hyphae are damaged by human natural killer (NK) cells, but suppress NK cell mediated immunity. Immunobiology 2013, 218, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Tramsen, L.; Lehrnbecher, T. Natural Killer Cells in Antifungal Immunity. Front. Immunol. 2017, 8, 1623. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, S.; Weiss, E.; Schmitt, A.L.; Schlegel, J.; Burgert, A.; Terpitz, U.; Sauer, M.; Moretta, L.; Sivori, S.; Leonhardt, I.; et al. CD56 Is a Pathogen Recognition Receptor on Human Natural Killer Cells. Sci. Rep. 2017, 7, 6138. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Tramsen, L.; Hanisch, M.; Latge, J.P.; Huenecke, S.; Koehl, U.; Lehrnbecher, T. Human natural killer cells exhibit direct activity against Aspergillus fumigatus hyphae, but not against resting conidia. J. Infect. Dis. 2011, 203, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Stuehler, C.; Kuenzli, E.; Jaeger, V.K.; Baettig, V.; Ferracin, F.; Rajacic, Z.; Kaiser, D.; Bernardini, C.; Forrer, P.; Weisser, M.; et al. Immune Reconstitution After Allogeneic Hematopoietic Stem Cell Transplantation and Association With Occurrence and Outcome of Invasive Aspergillosis. J. Infect. Dis. 2015, 212, 959–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Hughes, M.A.; Burdick, M.; Strieter, R.M.; Mehrad, B. Early NK cell-derived IFN-{gamma} is essential to host defense in neutropenic invasive aspergillosis. J. Immunol. 2009, 182, 4306–4312. [Google Scholar] [CrossRef] [PubMed]
- Benedetto, N.; Sabatini, P.; Sellitto, C.; Romano Carratelli, C. Interleukin-2 and increased natural killer activity in mice experimentally infected with Aspergillus niger. Microbiologica 1988, 11, 339–345. [Google Scholar]
- Tonn, T.; Schwabe, D.; Klingemann, H.G.; Becker, S.; Esser, R.; Koehl, U.; Suttorp, M.; Seifried, E.; Ottmann, O.G.; Bug, G. Treatment of patients with advanced cancer with the natural killer cell line NK-92. Cytotherapy 2013, 15, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Meagher, R.; Swearingen, M.; Myint, H.; Rich, E.; Martinson, J.; Klingemann, H. Infusion of the allogeneic cell line NK-92 in patients with advanced renal cell cancer or melanoma: A phase I trial. Cytotherapy 2008, 10, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Boyiadzis, M.; Agha, M.; Redner, R.L.; Sehgal, A.; Im, A.; Hou, J.Z.; Farah, R.; Dorritie, K.A.; Raptis, A.; Lim, S.H.; et al. Phase 1 clinical trial of adoptive immunotherapy using “off-the-shelf” activated natural killer cells in patients with refractory and relapsed acute myeloid leukemia. Cytotherapy 2017, 19, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Klingemann, H.; Grodman, C.; Cutler, E.; Duque, M.; Kadidlo, D.; Klein, A.K.; Sprague, K.A.; Miller, K.B.; Comenzo, R.L.; Kewalramani, T.; et al. Autologous stem cell transplant recipients tolerate haploidentical related-donor natural killer cell-enriched infusions. Transfusion 2013, 53, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Li, L.; McCarty, J.; Kaur, I.; Yvon, E.; Shaim, H.; Muftuoglu, M.; Liu, E.; Orlowski, R.Z.; Cooper, L.; et al. Phase I study of cord blood-derived natural killer cells combined with autologous stem cell transplantation in multiple myeloma. Br. J. Haematol. 2017, 177, 457–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewi, I.; van de Veerdonk, F.; Gresnigt, M. The Multifaceted Role of T-Helper Responses in Host Defense against Aspergillus fumigatus. J. Fungi 2017. [Google Scholar] [CrossRef] [PubMed]
- Perruccio, K.; Tosti, A.; Burchielli, E.; Topini, F.; Ruggeri, L.; Carotti, A.; Capanni, M.; Urbani, E.; Mancusi, A.; Aversa, F.; et al. Transferring functional immune responses to pathogens after haploidentical hematopoietic transplantation. Blood 2005, 106, 4397–4406. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulou, A.; Kaloyannidis, P.; Yannaki, E.; Cruz, C.R. Adoptive transfer of Aspergillus-specific T cells as a novel anti-fungal therapy for hematopoietic stem cell transplant recipients: Progress and challenges. Crit. Rev. Oncol. Hematol. 2016, 98, 62–72. [Google Scholar] [CrossRef]
- Tramsen, L.; Koehl, U.; Tonn, T.; Latge, J.P.; Schuster, F.R.; Borkhardt, A.; Uharek, L.; Quaritsch, R.; Beck, O.; Seifried, E.; et al. Clinical-scale generation of human anti-Aspergillus T cells for adoptive immunotherapy. Bone Marrow Transplant. 2009, 43, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Tramsen, L.; Schmidt, S.; Boenig, H.; Latge, J.P.; Lass-Florl, C.; Roeger, F.; Seifried, E.; Klingebiel, T.; Lehrnbecher, T. Clinical-scale generation of multi-specific anti-fungal T cells targeting Candida, Aspergillus and mucormycetes. Cytotherapy 2013, 15, 344–351. [Google Scholar] [CrossRef]
- Khanna, N.; Stuehler, C.; Conrad, B.; Lurati, S.; Krappmann, S.; Einsele, H.; Berges, C.; Topp, M.S. Generation of a multipathogen-specific T-cell product for adoptive immunotherapy based on activation-dependent expression of CD154. Blood 2011, 118, 1121–1131. [Google Scholar] [CrossRef] [Green Version]
- Stuehler, C.; Nowakowska, J.; Bernardini, C.; Topp, M.S.; Battegay, M.; Passweg, J.; Khanna, N. Multispecific Aspergillus T cells selected by CD137 or CD154 induce protective immune responses against the most relevant mold infections. J. Infect. Dis. 2015, 211, 1251–1261. [Google Scholar] [CrossRef]
- Bacher, P.; Jochheim-Richter, A.; Mockel-Tenbrink, N.; Kniemeyer, O.; Wingenfeld, E.; Alex, R.; Ortigao, A.; Karpova, D.; Lehrnbecher, T.; Ullmann, A.J.; et al. Clinical-scale isolation of the total Aspergillus fumigatus-reactive T-helper cell repertoire for adoptive transfer. Cytotherapy 2015, 17, 1396–1405. [Google Scholar] [CrossRef]
- Tramsen, L.; Schmidt, S.; Roeger, F.; Schubert, R.; Salzmann-Manrique, E.; Latge, J.P.; Klingebiel, T.; Lehrnbecher, T. Immunosuppressive compounds exhibit particular effects on functional properties of human anti-Aspergillus Th1 cells. Infect. Immun. 2014, 82, 2649–2656. [Google Scholar] [CrossRef] [PubMed]
- Tzannou, I.; Papadopoulou, A.; Naik, S.; Leung, K.; Martinez, C.A.; Ramos, C.A.; Carrum, G.; Sasa, G.; Lulla, P.; Watanabe, A.; et al. Off-the-Shelf Virus-Specific T Cells to Treat BK Virus, Human Herpesvirus 6, Cytomegalovirus, Epstein-Barr Virus, and Adenovirus Infections After Allogeneic Hematopoietic Stem-Cell Transplantation. J. Clin. Oncol. 2017, 35, 3547–3557. [Google Scholar] [CrossRef] [PubMed]
- Iannitti, R.G.; Carvalho, A.; Romani, L. From memory to antifungal vaccine design. Trends Immunol. 2012, 33, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Cutler, J.E.; Deepe, G.S., Jr.; Klein, B.S. Advances in combating fungal diseases: Vaccines on the threshold. Nat. Rev. Microbiol. 2007, 5, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhu, P.; Li, L.; Wan, Z.; Zhao, Z.; Li, R. Adoptive immunity mediated by HLA-A*0201 restricted Asp f16 peptides-specific CD8+ T cells against Aspergillus fumigatus infection. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2012, 31, 3089–3096. [Google Scholar] [CrossRef] [PubMed]
- Maldini, C.R.; Ellis, G.I.; Riley, J.L. CAR T cells for infection, autoimmunity and allotransplantation. Nat. Rev. Immunol. 2018, 18, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.-P.; Kros, J.M.; Li, J. Approved CAR T cell therapies: Ice bucket challenges on glaring safety risks and long-term impacts. Drug Discov. Today 2018, 23, 1175–1182. [Google Scholar] [CrossRef]
- Johnson, L.A.; June, C.H. Driving gene-engineered T cell immunotherapy of cancer. Cell Res. 2017, 27, 38–58. [Google Scholar] [CrossRef]
- Dotti, G.; Gottschalk, S.; Savoldo, B.; Brenner, M.K. Design and development of therapies using chimeric antigen receptor-expressing T cells. Immunol. Rev. 2014, 257, 107–126. [Google Scholar] [CrossRef]
- Kumaresan, P.R.; Silva, T.; Kontoyiannis, D.P. Methods of Controlling Invasive Fungal Infections Using CD8+ T Cells. Front. Immunol. 2018, 8, 1939. [Google Scholar] [CrossRef]
- Bridgeman, J.S.; Hawkins, R.E.; Bagley, S.; Blaylock, M.; Holland, M.; Gilham, D.E. The optimal antigen response of chimeric antigen receptors harboring the CD3zeta transmembrane domain is dependent upon incorporation of the receptor into the endogenous TCR/CD3 complex. J. Immunol. 2010, 184, 6938–6949. [Google Scholar] [CrossRef] [PubMed]
- Abken, H. Costimulation Engages the Gear in Driving CARs. Immunity 2016, 44, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Kumaresan, P.R.; Manuri, P.R.; Albert, N.D.; Maiti, S.; Singh, H.; Mi, T.; Roszik, J.; Rabinovich, B.; Olivares, S.; Krishnamurthy, J.; et al. Bioengineering T cells to target carbohydrate to treat opportunistic fungal infection. Proc. Natl. Acad. Sci. USA 2014, 111, 10660–10665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, H.; Manuri, P.R.; Olivares, S.; Dara, N.; Dawson, M.J.; Huls, H.; Hackett, P.B.; Kohn, D.B.; Shpall, E.J.; Champlin, R.E.; et al. Redirecting specificity of T-cell populations for CD19 using the Sleeping Beauty system. Cancer Res. 2008, 68, 2961–2971. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.R.; Ward, A.C.; Russell, A.P. Granulocyte Colony-Stimulating Factor and Its Potential Application for Skeletal Muscle Repair and Regeneration. Mediat. Inflamm. 2017, 2017, 7517350. [Google Scholar] [CrossRef] [PubMed]
- Clark, O.A.; Lyman, G.H.; Castro, A.A.; Clark, L.G.; Djulbegovic, B. Colony-stimulating factors for chemotherapy-induced febrile neutropenia: A meta-analysis of randomized controlled trials. J. Clin. Oncol. 2005, 23, 4198–4214. [Google Scholar] [CrossRef] [PubMed]
- Sionov, E.; Mendlovic, S.; Segal, E. Experimental systemic murine aspergillosis: Treatment with polyene and caspofungin combination and G-CSF. J. Antimicrob. Chemother. 2005, 56, 594–597. [Google Scholar] [CrossRef]
- Liles, W.C.; Huang, J.E.; van Burik, J.A.; Bowden, R.A.; Dale, D.C. Granulocyte colony-stimulating factor administered in vivo augments neutrophil-mediated activity against opportunistic fungal pathogens. J. Infect. Dis. 1997, 175, 1012–1015. [Google Scholar] [CrossRef]
- Smith, T.J.; Khatcheressian, J.; Lyman, G.H.; Ozer, H.; Armitage, J.O.; Balducci, L.; Bennett, C.L.; Cantor, S.B.; Crawford, J.; Cross, S.J.; et al. 2006 update of recommendations for the use of white blood cell growth factors: An evidence-based clinical practice guideline. J. Clin. Oncol. 2006, 24, 3187–3205. [Google Scholar] [CrossRef]
- Ringden, O.; Labopin, M.; Gorin, N.C.; Le Blanc, K.; Rocha, V.; Gluckman, E.; Reiffers, J.; Arcese, W.; Vossen, J.M.; Jouet, J.P.; et al. Treatment with granulocyte colony-stimulating factor after allogeneic bone marrow transplantation for acute leukemia increases the risk of graft-versus-host disease and death: A study from the Acute Leukemia Working Party of the European Group for Blood and Marrow Transplantation. J. Clin. Oncol. 2004, 22, 416–423. [Google Scholar] [CrossRef]
- Hume, D.A.; MacDonald, K.P. Therapeutic applications of macrophage colony-stimulating factor-1 (CSF-1) and antagonists of CSF-1 receptor (CSF-1R) signaling. Blood 2012, 119, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Nemunaitis, J.; Shannon-Dorcy, K.; Appelbaum, F.R.; Meyers, J.; Owens, A.; Day, R.; Ando, D.; O’Neill, C.; Buckner, D.; Singer, J. Long-term follow-up of patients with invasive fungal disease who received adjunctive therapy with recombinant human macrophage colony-stimulating factor. Blood 1993, 82, 1422–1427. [Google Scholar] [PubMed]
- Kandalla, P.K.; Sarrazin, S.; Molawi, K.; Berruyer, C.; Redelberger, D.; Favel, A.; Bordi, C.; de Bentzmann, S.; Sieweke, M.H. M-CSF improves protection against bacterial and fungal infections after hematopoietic stem/progenitor cell transplantation. J. Exp. Med. 2016, 213, 2269–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, C.E.; Lyman, C.A.; Lee, S.; Del Guercio, C.; Roilides, E.; Bacher, J.; Gehrt, A.; Feuerstein, E.; Tsokos, M.; Walsh, T.J. Recombinant human macrophage colony-stimulating factor augments pulmonary host defences against Aspergillus fumigatus. Cytokine 2001, 15, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, A.; Usui, T. Pivotal roles of GM-CSF in autoimmunity and inflammation. Mediat. Inflamm. 2015, 2015, 568543. [Google Scholar] [CrossRef] [PubMed]
- Scriven, J.E.; Tenforde, M.W.; Levitz, S.M.; Jarvis, J.N. Modulating host immune responses to fight invasive fungal infections. Curr. Opin. Microbiol. 2017, 40, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Ruef, C.; Coleman, D.L. Granulocyte-macrophage colony-stimulating factor: Pleiotropic cytokine with potential clinical usefulness. Rev. Infect. Dis. 1990, 12, 41–62. [Google Scholar] [CrossRef]
- Wan, L.; Zhang, Y.; Lai, Y.; Jiang, M.; Song, Y.; Zhou, J.; Zhang, Z.; Duan, X.; Fu, Y.; Liao, L.; et al. Effect of Granulocyte-Macrophage Colony-Stimulating Factor on Prevention and Treatment of Invasive Fungal Disease in Recipients of Allogeneic Stem-Cell Transplantation: A Prospective Multicenter Randomized Phase IV Trial. J. Clin. Oncol. 2015, 33, 3999–4006. [Google Scholar] [CrossRef]
- Safdar, A.; Rodriguez, G.; Zuniga, J.; Al Akhrass, F.; Georgescu, G.; Pande, A. Granulocyte macrophage colony-stimulating factor in 66 patients with myeloid or lymphoid neoplasms and recipients of hematopoietic stem cell transplantation with invasive fungal disease. Acta Haematol. 2013, 129, 26–34. [Google Scholar] [CrossRef]
- Brummer, E.; Maqbool, A.; Stevens, D.A. In vivo GM-CSF prevents dexamethasone suppression of killing of Aspergillus fumigatus conidia by bronchoalveolar macrophages. J. Leukoc. Biol. 2001, 70, 868–872. [Google Scholar]
- Quezada, G.; Koshkina, N.V.; Zweidler-McKay, P.; Zhou, Z.; Kontoyiannis, D.P.; Kleinerman, E.S. Intranasal granulocyte-macrophage colony-stimulating factor reduces the Aspergillus burden in an immunosuppressed murine model of pulmonary aspergillosis. Antimicrob. Agents Chemother. 2008, 52, 716–718. [Google Scholar] [CrossRef] [PubMed]
- Brummer, E.; Kamberi, M.; Stevens, D.A. Regulation by granulocyte-macrophage colony-stimulating factor and/or steroids given in vivo of proinflammatory cytokine and chemokine production by bronchoalveolar macrophages in response to Aspergillus conidia. J. Infect. Dis. 2003, 187, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Cenci, E.; Mencacci, A.; Fe d’Ostiani, C.; Del Sero, G.; Mosci, P.; Montagnoli, C.; Bacci, A.; Romani, L. Cytokine- and T helper-dependent lung mucosal immunity in mice with invasive pulmonary aspergillosis. J. Infect. Dis. 1998, 178, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Brieland, J.K.; Jackson, C.; Menzel, F.; Loebenberg, D.; Cacciapuoti, A.; Halpern, J.; Hurst, S.; Muchamuel, T.; Debets, R.; Kastelein, R.; et al. Cytokine networking in lungs of immunocompetent mice in response to inhaled Aspergillus fumigatus. Infect. Immun. 2001, 69, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Chai, L.Y.; van de Veerdonk, F.; Marijnissen, R.J.; Cheng, S.C.; Khoo, A.L.; Hectors, M.; Lagrou, K.; Vonk, A.G.; Maertens, J.; Joosten, L.A.; et al. Anti-Aspergillus human host defence relies on type 1 T helper (Th1), rather than type 17 T helper (Th17), cellular immunity. Immunology 2010, 130, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Estrada, C.; Desai, A.G.; Chirch, L.M.; Suh, H.; Seidman, R.; Darras, F.; Nord, E.P. Invasive aspergillosis in a renal transplant recipient successfully treated with interferon-gamma. Case Rep. Transplant. 2012, 2012, 493758. [Google Scholar] [CrossRef] [PubMed]
- Mezidi, M.; Belafia, F.; Nougaret, S.; Pageaux, G.P.; Conseil, M.; Panaro, F.; Boniface, G.; Morquin, D.; Jaber, S.; Jung, B. Interferon gamma in association with immunosuppressive drugs withdrawal and antifungal combination as a rescue therapy for cerebral invasive Aspergillosis in a liver transplant recipient. Minerva Anestesiol. 2014, 80, 1359–1360. [Google Scholar] [PubMed]
- Ellis, M.; Watson, R.; McNabb, A.; Lukic, M.L.; Nork, M. Massive intracerebral aspergillosis responding to combination high dose liposomal amphotericin B and cytokine therapy without surgery. J. Med. Microbiol. 2002, 51, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Kelleher, P.; Goodsall, A.; Mulgirigama, A.; Kunst, H.; Henderson, D.C.; Wilson, R.; Newman-Taylor, A.; Levin, M. Interferon-gamma therapy in two patients with progressive chronic pulmonary aspergillosis. Eur. Respir. J. 2006, 27, 1307–1310. [Google Scholar] [CrossRef]
- The International Chronic Granulomatous Disease Cooperative Study Group. A controlled trial of interferon gamma to prevent infection in chronic granulomatous disease. N. Engl. J. Med. 1991, 324, 509–516. [Google Scholar] [CrossRef]
- Armstrong-James, D.; Teo, I.A.; Shrivastava, S.; Petrou, M.A.; Taube, D.; Dorling, A.; Shaunak, S. Exogenous interferon-gamma immunotherapy for invasive fungal infections in kidney transplant patients. Am. J. Transplant. 2010, 10, 1796–1803. [Google Scholar] [CrossRef] [PubMed]
- Delsing, C.E.; Gresnigt, M.S.; Leentjens, J.; Preijers, F.; Frager, F.A.; Kox, M.; Monneret, G.; Venet, F.; Bleeker-Rovers, C.P.; van de Veerdonk, F.L.; et al. Interferon-gamma as adjunctive immunotherapy for invasive fungal infections: A case series. BMC Infect. Dis. 2014, 14, 166. [Google Scholar] [CrossRef] [PubMed]
- Bandera, A.; Trabattoni, D.; Ferrario, G.; Cesari, M.; Franzetti, F.; Clerici, M.; Gori, A. Interferon-gamma and granulocyte-macrophage colony stimulating factor therapy in three patients with pulmonary aspergillosis. Infection 2008, 36, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Gil-Lamaignere, C.; Winn, R.M.; Simitsopoulou, M.; Maloukou, A.; Walsh, T.J.; Roilides, E. Inteferon gamma and granulocyte-macrophage colony-stimulating factor augment the antifungal activity of human polymorphonuclear leukocytes against Scedosporium spp.: Comparison with Aspergillus spp. Med. Mycol. 2005, 43, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Gaviria, J.M.; van Burik, J.A.; Dale, D.C.; Root, R.K.; Liles, W.C. Comparison of interferon-gamma, granulocyte colony-stimulating factor, and granulocyte-macrophage colony-stimulating factor for priming leukocyte-mediated hyphal damage of opportunistic fungal pathogens. J. Infect. Dis. 1999, 179, 1038–1041. [Google Scholar] [CrossRef] [PubMed]
- Safdar, A.; Rodriguez, G.; Ohmagari, N.; Kontoyiannis, D.P.; Rolston, K.V.; Raad, I.I.; Champlin, R.E. The safety of interferon-gamma-1b therapy for invasive fungal infections after hematopoietic stem cell transplantation. Cancer 2005, 103, 731–739. [Google Scholar] [CrossRef]
- Sainz, J.; Perez, E.; Hassan, L.; Moratalla, A.; Romero, A.; Collado, M.D.; Jurado, M. Variable number of tandem repeats of TNF receptor type 2 promoter as genetic biomarker of susceptibility to develop invasive pulmonary aspergillosis. Hum. Immunol. 2007, 68, 41–50. [Google Scholar] [CrossRef]
- Roilides, E.; Dimitriadou-Georgiadou, A.; Sein, T.; Kadiltsoglou, I.; Walsh, T.J. Tumor necrosis factor alpha enhances antifungal activities of polymorphonuclear and mononuclear phagocytes against Aspergillus fumigatus. Infect. Immun. 1998, 66, 5999–6003. [Google Scholar]
- Nagai, H.; Guo, J.; Choi, H.; Kurup, V. Interferon-gamma and tumor necrosis factor-alpha protect mice from invasive aspergillosis. J. Infect. Dis. 1995, 172, 1554–1560. [Google Scholar] [CrossRef]
- Mehrad, B.; Strieter, R.M.; Standiford, T.J. Role of TNF-alpha in pulmonary host defense in murine invasive aspergillosis. J. Immunol. 1999, 162, 1633–1640. [Google Scholar]
- Roberts, N.J.; Zhou, S.; Diaz, L.A., Jr.; Holdhoff, M. Systemic use of tumor necrosis factor alpha as an anticancer agent. Oncotarget 2011, 2, 739–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pikman, R.; Ben-Ami, R. Immune modulators as adjuncts for the prevention and treatment of invasive fungal infections. Immunotherapy 2012, 4, 1869–1882. [Google Scholar] [CrossRef] [PubMed]
- Cassone, A.; Casadevall, A. Recent progress in vaccines against fungal diseases. Curr. Opin. Microbiol. 2012, 15, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, J.I.; Lyons, J.M. Vaccination of corticosteroid immunosuppressed mice against invasive pulmonary aspergillosis. J. Infect. Dis. 2002, 186, 869–871. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Capilla, J.; Johansen, M.E.; Alvarado, D.; Martinez, M.; Chen, V.; Clemons, K.V.; Stevens, D.A. Saccharomyces as a vaccine against systemic aspergillosis: ‘the friend of man’ a friend again? J. Med. Microbiol. 2011, 60, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Cenci, E.; Mencacci, A.; Bacci, A.; Bistoni, F.; Kurup, V.P.; Romani, L. T cell vaccination in mice with invasive pulmonary aspergillosis. J. Immunol. 2000, 165, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.I.; Lyons, J.M.; Hong, T.B.; Tamae, D.; Liu, Y.-K.; Wilczynski, S.P.; Kalkum, M. Vaccinations with recombinant variants of Aspergillus fumigatus allergen Asp f 3 protect mice against invasive aspergillosis. Infect. Immun. 2006, 74, 5075–5084. [Google Scholar] [CrossRef]
- Bozza, S.; Clavaud, C.; Giovannini, G.; Fontaine, T.; Beauvais, A.; Sarfati, J.; D’Angelo, C.; Perruccio, K.; Bonifazi, P.; Zagarella, S.; et al. Immune sensing of Aspergillus fumigatus proteins, glycolipids, and polysaccharides and the impact on Th immunity and vaccination. J. Immunol. 2009, 183, 2407–2414. [Google Scholar] [CrossRef]
- Liu, M.; Machova, E.; Nescakova, Z.; Medovarska, I.; Clemons, K.V.; Martinez, M.; Chen, V.; Bystricky, S.; Stevens, D.A. Vaccination with mannan protects mice against systemic aspergillosis. Med. Mycol. 2012, 50, 818–828. [Google Scholar] [CrossRef] [Green Version]
- Tomblyn, M.; Chiller, T.; Einsele, H.; Gress, R.; Sepkowitz, K.; Storek, J.; Wingard, J.R.; Young, J.A.; Boeckh, M.J. Guidelines for preventing infectious complications among hematopoietic cell transplantation recipients: A global perspective. Biol. Blood Marrow Transplant. 2009, 15, 1143–1238. [Google Scholar] [CrossRef]
- Edwards, J.E., Jr.; Schwartz, M.M.; Schmidt, C.S.; Sobel, J.D.; Nyirjesy, P.; Schodel, F.; Marchus, E.; Lizakowski, M.; DeMontigny, E.A.; Hoeg, J.; et al. A Fungal Immunotherapeutic Vaccine (NDV-3A) for Treatment of Recurrent Vulvovaginal Candidiasis-A Phase 2 Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Infect. Dis. 2018, 66, 1928–1936. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L.A. Immunoglobulins in defense, pathogenesis, and therapy of fungal diseases. Cell Host Microbe 2012, 11, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Malphettes, M.; Gerard, L.; Galicier, L.; Boutboul, D.; Asli, B.; Szalat, R.; Perlat, A.; Masseau, A.; Schleinitz, N.; Le Guenno, G.; et al. Good syndrome: An adult-onset immunodeficiency remarkable for its high incidence of invasive infections and autoimmune complications. Clin. Infect. Dis. 2015, 61, e13–e19. [Google Scholar] [CrossRef] [PubMed]
- Braem, S.G.; Rooijakkers, S.H.; van Kessel, K.P.; de Cock, H.; Wosten, H.A.; van Strijp, J.A.; Haas, P.J. Effective Neutrophil Phagocytosis of Aspergillus fumigatus Is Mediated by Classical Pathway Complement Activation. J. Innate Immun. 2015, 7, 364–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torosantucci, A.; Chiani, P.; Bromuro, C.; De Bernardis, F.; Palma, A.S.; Liu, Y.; Mignogna, G.; Maras, B.; Colone, M.; Stringaro, A.; et al. Protection by anti-beta-glucan antibodies is associated with restricted beta-1,3 glucan binding specificity and inhibition of fungal growth and adherence. PLoS ONE 2009, 4, e5392. [Google Scholar] [CrossRef] [PubMed]
- Cenci, E.; Mencacci, A.; Spreca, A.; Montagnoli, C.; Bacci, A.; Perruccio, K.; Velardi, A.; Magliani, W.; Conti, S.; Polonelli, L.; et al. Protection of killer antiidiotypic antibodies against early invasive aspergillosis in a murine model of allogeneic T-cell-depleted bone marrow transplantation. Infect. Immun. 2002, 70, 2375–2382. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Dadachova, E. Radioimmunotherapy of fungal diseases: The therapeutic potential of cytocidal radiation delivered by antibody targeting fungal cell surface antigens. Front. Microbiol. 2011, 2, 283. [Google Scholar] [CrossRef]
- Rezvani, K.; Rouce, R.; Liu, E.; Shpall, E. Engineering Natural Killer Cells for Cancer Immunotherapy. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 1769–1781. [Google Scholar] [CrossRef]
- Daver, N.; Kontoyiannis, D.P. Checkpoint inhibitors and aspergillosis in AML: The double hit hypothesis. Lancet Oncol. 2017, 18, 1571–1573. [Google Scholar] [CrossRef]
- Fites, J.S.; Gui, M.; Kernien, J.F.; Negoro, P.; Dagher, Z.; Sykes, D.B.; Nett, J.E.; Mansour, M.K.; Klein, B.S. An unappreciated role for neutrophil-DC hybrids in immunity to invasive fungal infections. PLoS Pathog. 2018, 14, e1007073. [Google Scholar] [CrossRef]
- Ngo, H.X.; Shrestha, S.K.; Garneau-Tsodikova, S. Identification of Ebsulfur Analogues with Broad-Spectrum Antifungal Activity. ChemMedChem 2016, 11, 1507–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiederhold, N.P.; Patterson, T.F.; Srinivasan, A.; Chaturvedi, A.K.; Fothergill, A.W.; Wormley, F.L.; Ramasubramanian, A.K.; Lopez-Ribot, J.L. Repurposing auranofin as an antifungal: In vitro activity against a variety of medically important fungi. Virulence 2017, 8, 138–142. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauruschkat, C.D.; Einsele, H.; Loeffler, J. Immunomodulation as a Therapy for Aspergillus Infection: Current Status and Future Perspectives. J. Fungi 2018, 4, 137. https://doi.org/10.3390/jof4040137
Lauruschkat CD, Einsele H, Loeffler J. Immunomodulation as a Therapy for Aspergillus Infection: Current Status and Future Perspectives. Journal of Fungi. 2018; 4(4):137. https://doi.org/10.3390/jof4040137
Chicago/Turabian StyleLauruschkat, Chris D., Hermann Einsele, and Juergen Loeffler. 2018. "Immunomodulation as a Therapy for Aspergillus Infection: Current Status and Future Perspectives" Journal of Fungi 4, no. 4: 137. https://doi.org/10.3390/jof4040137