
Biological Activities of Secondary Metabolites from the Edible-Medicinal Macrofungi

1
State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China
2
University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
J. Fungi 2024, 10(2), 144; https://doi.org/10.3390/jof10020144
Submission received: 31 December 2023
/
Revised: 26 January 2024
/
Accepted: 8 February 2024
/
Published: 11 February 2024
(This article belongs to the Special Issue Edible and Medicinal Macrofungi, 3rd Edition)
Abstract
:Macrofungi are well-known as edible-medicinal mushrooms, which belong mostly to Basidiomycota, with a few from Ascomycota. In recent years, macrofungi have been recognized as a rich resource of structurally unique secondary metabolites, demonstrating a wide range of bioactivities, including anti-tumor, antioxidant, anti-inflammatory, antimicrobial, antimalarial, neuro-protective, hypoglycemic, and hypolipidemic activities. This review highlights over 270 natural products produced by 17 families of macrofungi covering 2017 to 2023, including their structures, bioactivities, and related molecular mechanisms.
1. Introduction
Macrofungi, a kind of large and visible fungi, can form fleshy or gelatinous macroscopic fruiting bodies or sclerotia, which are usually known as mushrooms [1]. Most mushrooms belong to the Basidiomycete (such as Lentinus edodes), and a small portion belong to the Ascomycete (such as morels and truffles) [2]. These fruiting bodies contain a variety of nutrients, including protein, essential fatty acids, vitamins, and minerals [1]. Therefore, mushrooms are deeply loved by people due to their nutritional value. In order to protect the fruiting body from harmful organisms, including viruses, bacteria, and insects, mushrooms can secrete various secondary metabolites (such as terpenoids and phenolics) with antiviral, antioxidant, and other biological activities as chemical weapons. Mushrooms contain abundant active proteins, such as lectins with an antiviral effect [3], deoxyribonuclease with an anti-tumor function [4], and ribotoxin with an anti-proliferative effect [5]. Similarly, mushrooms are also one of the main sources of antioxidants ergothioneine and glutathione [6]. Modern pharmacology found that the secondary metabolites produced by mushrooms have certain medicinal value, including anti-Alzheimer [7], antidiabetic [8], and antitumor [9]. Lentinan is a type of glucan isolated from the fruiting body of shiitake mushrooms. It has detoxification activity and can significantly alleviate side effects such as leukocyte suppression, nausea, and vomiting caused by cancer chemotherapy. Currently, lentinan is used as injections for adjuvant treatment of cancer in clinical practices [10]. Additionally, lentinan can be used in combination with rabies vaccines. It not only resolves vaccine-induced side effects, but it also enhances therapeutic effects [11]. The mushroom resources that can be developed and utilized are very abundant. At the present time, approximately 2000 species of edible mushrooms and 650 species of medicinal mushrooms have been found in forests around the world [12], which have attracted the interest of food and pharmacy scholars due to their rich nutritional and medicinal values, as well as the abundant resources.
A comprehensive literature search of studies published from 2017 to 2023 was conducted by using the keywords “edible-medicinal mushroom”, “edible-medicinal macrofungi”, “edible mushroom”, “edible macrofungi”, “medicinal mushroom”, “medicinal macrofungi”, “mushroom”, “macrofungi” and “secondary metabolite”, and “secondary product” on PubMed, Web of Science, ACS, RSC, Reaxys, EMBASE, Springer Link, Elsevier Science Direct and Wiley. In this review, we focus on the latest progress of the biological activities of 274 secondary metabolites (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9) derived from macrofungi (Table 1). The aim of this review is to provide the theoretical basis for the research of secondary metabolites from large edible and medicinal fungi.
2. Biological Activities of Secondary Metabolites
2.1. Anti-Tumor Activity
The cancer incidence rate and cancer-related mortality are rising in the whole world. Based on cancer prevalence data, it is estimated that 28.4-million cancer cases will be diagnosed by 2040 [75]. At present, there are various methods of cancer treatment (such as chemotherapy, radiotherapy, immunotherapy, and others), among which chemotherapy is the most widely used [76]. Although chemotherapy drugs exhibit significant therapeutic effects, they have the disadvantages of low bioavailability, prominent side effects, and high susceptibility to drug resistance. In the last few years, it has been reported in the literature that natural active ingredients exhibit significant activity and few side effects [77], which have the potential as therapeutic drugs for inhibiting and treating cancer [13].
The following is a systematic review of the direct cytotoxic effect of secondary metabolites obtained from macrofungi fruiting bodies or mycelium against cancer cells. These studies mainly validated the inhibitory activity of secondary metabolites on malignant tumors such as lung cancer, liver cancer, and cervical cancer through 3-(4,5-dimethylthiazol-2-yl) -2,5-diphenyltetrazolium bromide (MTT) experiments.
The first example is a research evaluating the effect of secondary metabolites on human lung cancer cell lines using cell proliferation and cytotoxicity assays. The results demonstrated the suppressive activity of 2, 3, and 72 against A549, H1264, H1299, and Calu-6 cells. Among them, 3 exhibited the best suppressive activity on Calu-6 cells, with the half maximal inhibitory concentration (IC50) of 133.1 ± 4.6 μM [13]. Compound 4 (IC50 = 37.5–150 μM) displayed obvious antiproliferative and cytotoxic activity to MCF-7, HepG2, HeLa, HCT-116, and Caco-2 cells [14]. The results of MTT displayed that 11–16 had inhibitory activity against HL-60, A-549, SMMC-7721, MCF-7, and SW480 cell lines. Compounds 11 (IC50 = 3.52 ± 0.79−12.95 ± 0.73 μM), 12 (IC50 = 2.88 ± 0.08−8.27 ± 0.12 μM), and 16 (IC50 = 3.09 ± 0.03−12.48 ± 0.5 μM) exhibited more significant cytotoxicities, while 13 (IC50 = 10.77 ± 0.85−25.89 ± 0.87 μM), 14 (IC50 = 3.52 ± 0.79−12.95 ± 0.73 μM), and 15 (IC50 = 14.09 ± 0.86−23.23 ± 0.88 μM) exhibited moderate cytotoxicities (cisplatin as positive drug, IC50 = 2.95 ± 0.13−23.64 ± 1.6 μM) [15]. Compound 20 showed a cytotoxic effect against PC-3 with an IC50 value of 27.43 ± 0.86 µg/mL (doxorubicin as a positive drug, IC50 = 1.38 ± 0.16 μg/mL). Compounds 18, 23, and 24 were cytotoxic against MCF-7 with IC50 values of 28.74 ± 0.36, 8.90 ± 0.27, and 17.05 ± 0.54 µg/mL, respectively (cycloheximide as a positive drug, IC50 = 0.073 ± 0.12 μg/mL). Among them, 23 had the strongest antiproliferative activity against the MCF-7 cell line [16]. Compounds 27 and 28 showed obvious cytotoxicity against HL-60, SMMC-7721, and SW480 cell lines, especially on SW480 cells with IC50 values of 0.7 and 1.1 μM, respectively [17]. Compounds 36, 37, 39, and 41 showed cytotoxicity against K562 cells, with IC50 values of 68.2 ± 2.2, 45.3 ± 1.2, 33.1 ± 0.5, and 25.7 ± 1.7 μM, respectively (cisplatin as a positive drug, IC50 = 3.8 ± 0.2 μM) [18]. Compound 47 showed obvious cytotoxic activity against HL-60, A-549, SMMC-7721, MCF-7, and SW480 cells, with the best suppressive activity on SW480 of the IC50 value of 18.1 μM [19]. Compounds 48–52 showed cytotoxicity against MCF-7 at a concentration of 100 μM [20]. Compound 53 exhibited a significant cytotoxic effect on HepG2 (IC50 = 9.1 μM) compared to the positive control sorafenib (IC50 = 5.5 μM). Moreover, using cisplatin with the IC50 value of 1.6 μM as the standard, 53 was evidently cytotoxic to the sorafenib-resistant Huh7 (IC50 = 6.2 μM) [21]. Compound 54 (IC50 = 6.82 ± 0.77 μM) had a cytotoxic effect on HGC-27 cells. Compounds 55 (IC50 = 13.67 ± 1.04 μM) and 56 (IC50 = 37.93 ± 3.22 μM) could inhibit the proliferation of A549 cells [22]. Compounds 60–63 had an inhibitory effect on HT29 and MCF7 cells, and 63 (IC50 = 38.8 ± 0.9 μg/mL) had a stronger effect on the HT29 cell line than the MCF7 cells [25]. The results of cell proliferation assay showed that 64 (IC50 = 1.5 ± 0.1−10.3 ± 0.25 μM) and 65 (IC50 = 0.3 ± 0.05−4.5 ± 0.15 μM) had cytotoxicity against HL-60, A-549, HepG2, Caki-1, and MCF-7 cells, which illustrated that 65 had stronger inhibitory activity (cisplatin as positive drug, IC50 = 1.2 ± 0.3−17.6 ± 0.4 μM) [26]. Compound 66 showed cytotoxicity against the SMMC-7721 cell line (IC50 = 15.8 μM) [27]. Compound 70 (IC50 = 17.1 μM) exhibited cytotoxicity against HL-60, while compound 1 had a moderate inhibitory effect on HL-60, SMMC-7721, A549, MCF-7, and SW-480 cells [28].
According to the above analysis, it is found that secondary metabolites derived from mushrooms have a significant anticancer effect, especially in inhibiting the growth of lung cancer cells (Calu-6), colorectal cancer cells (SW480), and hepatoma cells (SMMC-7721). Furthermore, it is worth mentioning that 27, 28, and 65 exhibit remarkable inhibitory activity against various cancer cells.
2.2. Antioxidant Activity
The pathogenesis of many human diseases (cancer, atherosclerosis, Alzheimer’s disease, and others) is related to oxidative stress. The occurrence of oxidative stress is caused by the imbalance between oxidation and antioxidation in the body [78]. Therefore, maintaining oxidative balance in the body helps to suppress the progression of diseases, and it is necessary to explore new drugs with antioxidant activity. The following is a general overview of studies on secondary metabolites with antioxidant activity of large edible and medicinal fungi in recent years. These studies mainly validated the antioxidant activity through 1,1-dipheny-1-2-picrylhydrazyl (DPPH) and 2,2-azinobis (3-ethylbenzothiazoline-6-sulphonic acid) (ABTS), the free-radical scavenging assay, the cupric-reducing antioxidant capacity (CUPRAC), and the oxygen radical-absorbance capacity [79].
The results of the ABTS assay showed that aromatic meroterpenoid compounds 74 (EC50 (concentration for 50% of maximal effect) = 0.59 ± 0.15 mM) and 75 (EC50 = 0.27 ± 0.05 mM) had stronger radical scavenging effects than the positive drug (trolox, EC50 = 0.42 ± 0.03 mM). Moreover, ORAC experimental results indicated that 74 (5.42 ± 0.2 µmol TE/µmol) and 75 (7.24 ± 0.15 µmol TE/µmol) had strong antioxidant activity. Among them, the antioxidant effect of 75 was similar to the positive control (quercetin 7.78 ± 0.27 µmol TE/µmol) [29]. ABTS, DPPH, and β-carotene linoleic acid experiments results showed that phenolic compounds 25 (IC50 = 1.06 ± 0.1−3.14 ± 0.11 μg/mL) and 26 (IC50 = 1.06 ± 0.46−10.26 ± 0.56 μg/mL) exhibited antioxidant activity [16]. In the DPPH radical-scavenging experiment, Vitamin C was used as the positive control. Compound 78 had strong scavenging ability at the concentration of 0.25 mg/mL, while 76 and 77 had moderate scavenging ability against DPPH radical [30]. Compound 79 had a scavenging ability against DPPH radicals, while 80 and 81 had less scavenging ability against DPPH radicals [31]. The DPPH radical-scavenging activity of 57 (IC50 = 21.7 μg/mL) was better than that of Vitamin C (IC50 = 50.0 μg/mL) [23]. Compound 68 showed antioxidant activity at the concentration value of 50 μM against the DPPH radical [27]. Moreover, it was found that phenolic compounds (84–87) had good antioxidant activity in a dose-dependent manner. And the antioxidant activity of 86 was the best, with the cellular antioxidant activity value of 5.31 μM [32]. Compounds 88 (IC50 = 10.39 ± 2.26−18.58 ± 2.33 μg/mL) and 89 (IC50 = 16.57 ± 2.48−20.43 ± 3.74 μg/mL) showed significant radical-scavenging activities [33]. An analysis of the results of antioxidant experiments revealed that 96 (IC50 = 11.5 ± 0.51 μM) and 98–101 (IC50 = 6.7 ± 0.05−15.5 ± 0.50 μM) had stronger activity against the DPPH radical than the positive drug (tert-butylhydroquinone, IC50 = 17.3 ± 1.32 μM). Moreover, Compounds 94–101 (IC50 = 44.6 ± 5.12−186.7 ± 8.14 μM) had the ability to scavenge superoxide anion radicals, and compounds 96–101 (IC50 = 99.5 ± 11.27−292.7 ± 12.5 μM) had ability to scavenge hydroxyl radicals than the positive drug (tert-butylhydroquinone, IC50 = 186.7 ± 8.14 μM). In the meanwhile, compounds 94 (IC50 = 783.4 ± 20.35 μM), 95 (IC50 = 897.9 ± 26.39 μM), 97 (IC50 = 667.2 ± 12.17 μM), and 101 (IC50 = 292.7 ± 12.5 μM) showed a weak ability to scavenge hydroxyl radicals (tert-butylhydroquinone as positive drug, IC50 = 271.5 ± 19.22 μM) [34]. Compound 103 (EC50 = 573 μM) showed moderate activity against the DPPH radical [35]. Phenolic compounds (104–110) showed a DPPH radical scavenging ability, and the IC50 value of these compounds were 1.79, 4.10, 4.28, 2.45, 4.40, 1.73, and 6.00 mM, respectively [36]. In the ABTS radical-scavenging activity test, 111 (EC50 = 0.001 mg/mL) and 112 (EC50 = 0.145 mg/mL) showed strong antioxidant activity compared with Vitamin C (EC50 < 0.025 mg/mL) [37].
Based on the above analysis, it is found that aromatic meroterpenoid and phenolic compounds derived from mushrooms have obvious antioxidant activities, especially phenolic compounds. To sum up, 25, 26, 57, 78, 86, 96, 98–101, 109, 111, and 112 exhibit outstanding a radical scavenging activity.
2.3. Anti-Inflammatory Activity
Inflammation is a protective response against injury and infection by secreting nitric oxide (NO) and pro-inflammatory cytokines [80]. A moderate inflammatory response helps to resist external harmful stimuli. However, excessive inflammation can easily lead to acute diseases such as enteritis and arthritis. And prolonged inflammation can easily cause chronic diseases [81]. Drugs including steroids and nonsteroidals are commonly used in clinical to alleviate inflammation even though their side effects (hypertension, hepatotoxicity, and others) are non-negligible [82]. Therefore, it is imperative to search for potential low-toxic and effective anti-inflammatory drugs. The following is a summary of studies on secondary metabolites with the anti-inflammatory activity of large edible and medicinal fungi in recent years. These studies mainly validated the anti-inflammatory activity through constructing inflammation models using lipopolysaccharide (LPS).
Compounds 5–10 (IC50 = 17.9–34.9 μM) could prominently inhibit the production of NO by LPS-induced RAW264.7 macrophages in mice and were superior to L-NG monomethylarginine (IC50 = 47.1 μM) [15]. Evaluated by the same experiment, Compounds 69–72 had anti-inflammatory properties at lower concentrations (6.25–25 μM) [28]. Compounds 113–117 (IC50 = 49.43–82.32 μM) exhibited potential anti-inflammatory activity and could strongly inhibit the production of NO in RAW264.7 macrophages; compound 117 (IC50 = 49.43 μM) had the strongest activity. Further study on the mechanism showed that 117 inhibited the expression of nuclear factor-kappa B (NF-κB), nitric oxide synthase (iNOS), and cyclooxygenase-2 (COX-2) to exert anti-inflammatory effect [38]. In the LPS-induced inflammatory response of RAW264.7 macrophages, 118 (IC50 = 38.6 ± 1.0 μM) considerably inhibited the release of NO, as well as pro-inflammatory cytokines (interleukin-1β (IL-1β) and interleukin-6 (IL-6)) together with the phosphorylation of inhibitor kappa B kinase β (IKKβ) and inhibitor kappa B alpha (IκBα). These results indicated that 118 inhibited the NF-κB-signaling pathway to exert anti-inflammatory activity [39]. Compounds 119–126 could inhibit the NO released from the LPS-induced RAW264.7 macrophages, with IC50 values in the range of 21.4–27.2 μM. Among them, 126 (IC50 = 21.4 ± 0.04 μM) exhibited better anti-inflammatory activity (hydrocortisone as a positive drug, IC50 = 22.4 ± 1.56 μM) [40]. Compounds 128 (IC50 = 20.77 μM) and 129 (IC50 = 18.52 μM) had strong anti-inflammatory activity and could suppress the NO released from the LPS-induced BV-2 microglial cells [41]. Compounds 130, 131, 134, and 136 could notably inhibit the NO released from the LPS-induced BV-2 microglial cells with IC50 values ranging from 2.32 to 23.83 μM. Among them, 130 (IC50 = 3.985 ± 0.01 μM), 131 (IC50 = 4.074 ± 0.03 μM), and 134 (IC50 = 2.32 ± 0.02 μM) had better anti-inflammatory activity and were superior to positive drugs (quercetin, IC50 = 4.01 ± 0.7 μM) [42]. Compounds 137 (IC50 = 56.33 ± 6.81–87.31 ± 8.77 μM) and 138 (IC50 = 48.50 ± 6.54−76.16 ± 9.11 μM) inhibited the release of pro-inflammatory mediators (tumor necrosis factor-α [TNF-α], IL-6, and NO) in LPS-induced RAW 264.7 macrophage cells in a dose-dependent manner (aspirin as positive drug, IC50 = 27.08 ± 1.86−51.82 ± 8.62 μM) [43]. Compounds 139–141 (IC50 = 14.3–42.3 μM) could significantly inhibit the production of NO by the LPS-induced RAW264.7 cell line and were superior to minocycline (IC50 = 73.0 μM) [44]. The anti-inflammatory experimental results showed that 142 (IC50 = 0.9 ± 0.1 μM) and 143 (IC50 = 0.6 ± 0.1 μM) prominently reduced the secretion of NO at extremely low doses and completely suppressed the release of TNF-α and IL-6 at 10 μM (silymarin as a positive drug, IC50 = 1.8 ± 0.4−86.5 ± 2.5 μM) [45].
As to the analysis above, it is found that these compounds exert an anti-inflammatory effect by inhibiting the secretion of NO and the expression of IL-6 and TNF-α and the NF-κB-signaling pathway. Meanwhile, Compounds 69–72, 126, 129–131, 134, and 143 have remarkable anti-inflammatory effects. Therefore, it is speculated that these secondary metabolites have the potential to become highly effective anti-inflammatory agents.
2.4. Antimicrobial Activity
Invasive fungal and bacterial infections are key causes of the incidence rate and mortality in immune-compromised populations [83,84]. Antibiotics are often used for the treatment of microbe infections. However, the emergence of antibiotic resistance has greatly reduced the effectiveness of antimicrobial drugs and will increase the severity of infection, morbidity, and treatment costs [85]. Therefore, newer and more effective antibiotics are urgently needed. Similarly, the treatment cycle for fungal infection is long, which means there is an urgent need to explore more effective antifungal drugs. The following is a systematic review of studies on secondary metabolites with the antimicrobial activity of mushrooms in recent years. These studies mostly evaluated the inhibitory activity of secondary metabolites on various fungi (Candida albicans and Cryptococcus neoformans) and bacteria (Mycobacterium tuberculosis and Staphylococcus aureus) through the minimum inhibitory concentration (MIC) index. Moreover, in order to facilitate the comparison of the strength of activity of the compounds, the MIC values were categorized into three classes according to Kuete’s work [86]: significant (MIC < 10 μg/mL), moderate (10 < MIC < 100 μg/mL), and low or negligible (MIC > 100 μg/mL).
Compared with the positive drug (isoniazid, MIC = 0.047 μg/mL), 13 (MIC = 50 μg/mL) was moderately active in inhibiting the growth of M. tuberculosis H37Ra [15]. The results of the antimicrobial activity experiment showed that 58 (MIC = 66 μg/mL) had moderate activity against the sensitive non-pathogenic zygomycete Mucor hiemalis [24]. Using ciprofloxacin (MIC = 0.9 μM) as the positive drug, 102 (MIC = 90.3 μM) exhibited moderate inhibitory activity against S. aureus [35]. Among the secondary metabolites obtained from Ganoderma species, 7α-acetoxy derivatives 144–146 (MIC = 12.5−25 μg/mL) exhibited moderate anti-tuberculosis activity, while 147 (MIC = 6.25 μg/mL) and 148 (MIC = 1.56 μg/mL) could significantly inhibit M. tuberculosis H37Ra growth (isoniazid as positive drug, MIC = 0.094 μg/mL) [46]. The results of the anti-tuberculosis activity experiment showed that compound 163 (MIC = 12.5 μg/mL) had moderate activity [47]. Furthermore, both 166 (MIC = 16.7–100 μg/mL) and 167 (MIC = 8.3–66.7 μg/mL) had shown a moderate inhibitory effect on fungi (M. hiemalis, C. tenuis) and Gram-positive bacteria (Micrococcus luteus, S. aureus, and Bacillus subtilis). Compounds 164 (MIC = 33.3–66.7 μg/mL) and 165 (MIC = 33.3–66.7 μg/mL) had a moderate inhibitory effect on some yeasts (Pichia anomala and Rhodotorula glutinis) [48]. Compounds 168 (MIC = 25 μg/mL) and 170 (MIC = 100 μg/mL) could inhibit the proliferation of M. luteus [49]. Compounds 171–174 (MIC = 75–100 μg/mL) exhibited moderate antibacterial activity (S. aureus), while 174 exhibited an inhibitory effect on both fungi (M. hiemalis) and bacteria (S. aureus and B. subtilis) [50]. Compound 175 (MIC = 0.9–3.1 μg/mL) could significantly inhibit the reproduction of human pathogenic fungi (C. albicans and C. neoformans), and its activity was equivalent to the amphotericin B (MIC = 0.4–0.8 μg/mL) [51].
Based on the above analysis, compounds 147 and 148 are the potential drugs for the treatment of tuberculosis.
2.5. Antimalarial Activity
Malaria is a disease caused by the infection of protozoan parasites in the genus Plasmodium, which is prevalent in tropical regions (especially in Africa, Southeast Asia, and South America) and has a significant mortality rate [87]. It is necessary to explore effective antimalarial drugs. The following is a brief review of studies on secondary metabolites with antimalarial activity of mushrooms in recent years. Compounds 149–162 (IC50 = 5.1−19 μM) from the fruiting body of the wood-rot Tomophagus sp. showed antimalarial activity, of which 150 (IC50 = 5.1 μM) had the strongest antimalarial activity (dihydroartemisinin as positive drug, IC50 = 0.0028 μM) [47]. Compound 176 (IC50 = 257.8 nM) had an inhibitory effect on chloroquine-sensitive strain P. falciparum (chloroquine as positive drug, IC50 = 22.66 nM) [52]. The antimalaria test results showed that 177 (IC50 = 0.05 μM) and 178 (IC50 = 0.45 μM) had significant inhibitory effects on P. falciparum, while chloroquine (IC50 = 0.50 μM) was used as positive control [53]. Overall, compounds 150, 177, and 178 exhibit potential antimalarial activity effects. It is speculated that these compounds have the potential to become more effective antimalarial drugs.
2.6. Neuro-Protective Activity
In recent years, the incidence rate of nervous system diseases (Alzheimer’s disease, stroke, and others) has increased continuously and has gradually become the main cause of global disability [88]. Neuroglial cells are the most important cells in the central nervous system, which maintain homeostasis and the operation of the central nervous system through interactions with neurons, immune cells, and other factors [89]. The following is a systematic review of studies on secondary metabolites with the neuroprotective effect of large edible and medicinal fungi in recent years. These studies mainly validated the neuroprotective effect by constructing neural system injury models using the PC12 cell line and BV-2 microglia.
Compounds 73–75 could alleviate H2O2, and amyloid β-protein-induced SH-SY5Y cells damaged through the reduction of the production of reactive oxygen species (ROS) or free radicals [29]. Compounds 128 and 129 could exert neuroprotective effects by reducing the production of NO in BV-2 microglia [41]. Compounds 130–136 exerted neuroprotective effects by reducing the production of NO in BV-2 microglia, with 130, 131, and 134 exhibiting superior neuroprotective activity compared to positive drugs (quercetin) [42]. Compounds 179 and 180 could enhance nerve growth factor-induced neurite outgrowth in PC12 cells through the tyrosine kinase A (TrkA) and kinase1/2 (ERK1/2) pathway to exert neuroprotective effect [54]. Compounds 181 and 182 could alleviate hydrogenperoxide (H2O2)-induced PC12 cell damage by stimulating neurite activity. Through electron microscopy observation, it was found that 181 and 182 showed neurotrophic effects on undifferentiated PC12 cells [55]. Compounds 183–186 exerted neuroprotective effects by promoting the release of neurotrophic factors in astrocytic cells [56]. Compounds 187–192 exhibited neurotrophic activity and could promote axonal growth [57]. Compounds 193–199 could exert neuroprotective effects by reducing the production of NO in BV-2 microglia, and 193 had the most significant effect. Further research found that the neuroprotective effect of 193 could be related to the reduction of the levels of IL-1β, IL-6, and TNF-α and the mitigation of abnormal changes in mitochondrial membrane potential and reduction in ROS generation. Mechanism studies showed that the neuroprotective effect of 193 was related to its inhibition of TLR-4/NF-κB and MAPK-signaling pathways and activation of the Akt/GSK-3β/Nrf2-signaling pathway [58]. Compound 200 reduced the production of ROS and alleviated mitochondrial damage to protect SH-SY5Y cells from H2O2-induced damage, which could be related to the Nrf2- and BDNF/TrkB/ERK/CREB-signaling pathways [59]. Compounds 201–203 could considerably enhance neurite outgrowth in PC-12 cells [60]. Compounds 204–206 exerted a neuroprotective effect by reducing the levels of inflammatory cytokines. Compound 201 could markedly reduce the level of TNF-α in BV-2 microglia, and 205 significantly reduced the level of IL-6 in BV-2 microglia. Compound 206 noticeably reduced the levels of NO and IL-1β in BV-2 microglia, as well as the expression of phosphorylated nuclear factor-kappa B inhibitor-α and the activity of iNOS [61].
According to the above analysis, it is found that these compounds exert neuroprotective effects by mitigating oxidative stress, expressing pro-inflammatory factors, activating TrkA/ERK1/2-, Nrf2-, or BDNF/TrkB/ERK/CREB-signaling pathways, or inhibiting TLR-4/NF-κB- or MAPK-signaling pathways.
2.7. Hypoglycemic Activity
The elevation of blood glucose levels mainly causes diabetes, which also causes metabolic disorders in the organism. Moreover, high blood glucose can easily induce cardiovascular diseases, obesity, kidney diseases, and other diseases [90]. Therefore, maintaining blood glucose balance in the body is of great importance. The α-glucosidase, aldose reductase (AR), maltase, sucrase, and protein tyrosine phosphatase 1b (PTP1B) play important roles in lowering blood glucose levels [91,92,93,94]. The α-glucosidase and AR can mainly act on hyperglycemia to lower blood glucose levels [95,96]. PTP1B can reduce blood glucose levels by negatively regulating the insulin metabolism pathway [97]. This section reviewed promising drugs derived from mushrooms that can suppress α-glucosidase, AR, maltase, sucrase, and PTP1B enzyme activities.
Compounds 29–36 and 38–45 exhibited insulin-sensitization effects, and compound 46 significantly increased glucose uptake in 3T3-L1 adipocytes [18]. Compounds 82–87 could effectively alleviate insulin-induced decreases in glucose uptake in HepG2 cells and 82 had the minimum glucose uptake rate of 36.6% [32]. Compounds 121–125 and 127 had significant inhibitory effects on PTP1B, with IC50 values in the range from 20.5 to 56.4 μM. Compounds 123, 124, and 127 could effectively improve the decrease of glucose uptake in HepG2 cells induced by insulin [40]. Compounds 207, 208, 210–212, 215, 216, and 222–229 had prominent suppressed effects against AR. Compound 224 (IC50 = 19.1 μM) had similar suppressed activity on AR as positive control epalrestat (IC50 = 17.5 μM). Compounds 207 (IC50 = 60.1 ± 10.1 μM), 209 (IC50 = 91.2 ± 13.6 μM), 210 (IC50 = 18.1 ± 2.3 μM), 218 (IC50 = 46.5 ± 5.1 μM), 220 (IC50 = 32.5 ± 3.1 μM), 223 (IC50 = 5.4 ± 0.4 μM), 224 (IC50 = 0.6 ± 0.12 μM), 228 (IC50 = 21.5 ± 2.2 μM), and 230 (IC50 = 8.1 ± 2.9 μM) had a suppressed effect against α-glucosidase, while acarbose (IC50 = 38.1 ± 6.0 μM) was used as positive control. Moreover, Compounds 207 (IC50 = 51 ± 3.5 μM), 210 (IC50 = 11.1 ± 0.3 μM), 218 (IC50 = 52.1 ± 9.1 μM), 220 (IC50 = 16.9 ± 1.2 μM), 223 (IC50 = 17.1 ± 4.2 μM), 224 (IC50 = 3.9 ± 0.7 μM), and 230 (IC50 = 10.1 ± 1.9 μM) had inhibitory effects against maltase, while acarbose (IC50 = 16.1 ± 4.1 μM) was used as a positive control. Compounds 211−215 (IC50 = 18.1 ± 2.5−41.9 ± 4.9 μM), 219 (IC50 = 7.6 ± 2.8 μM), and 220 (IC50 = 25.8 ± 3.1 μM) showed suppressed activity against PTP1B, while sodium vanadate (IC50 = 1.3 ± 0.2 μM) was used as a positive control [62]. Compounds 231–233 (IC50 = 36.2−40.8 μM) displayed good α-glucosidase inhibitory activity [63]. Compounds 234–238 (IC50 = 3.9−19.6 μM) exhibited significant inhibition of α-glucosidase, and 237 (IC50 = 3.9 μM) had better inhibitory activity, while acarbose (IC50 = 71.2 μM) was used as a positive control [64]. Similarly, the acarbose (IC50 = 59.48 ± 2.13 μM) was used as the positive drug, 239–242 exhibited significant inhibitory against α-glucosidase, and 241 had the best inhibitory activity (IC50 = 14.65 ± 1.68 μM) [65].
Based on the above analysis, it is found that 210, 223, and 224 have a significant suppression of α-glucosidase, AR, maltase, and sucrase. Additionally, compounds 35, 38, 40, 82, 123, 124, and 127 effectively inhibit cell absorption of glucose. Therefore, they have great potential in the treatment of diabetes.
2.8. Hypolipidemic Activity
The global prevalence of obesity is on the rise, and obesity is prone to induce a variety of diseases (diabetes, cardiovascular disease, and others) [98]. Lipases, especially pancreatic lipase, are essential enzymes for lipid absorption, and 3-hydroxy-3-methyl glutaryl Coenzyme A reductase (HMG-CoA) is a rate-limiting enzyme in cholesterol biosynthesis [99,100]. Therefore, HMG-CoA and lipase are important targets for treating obesity symptoms. This section reviewed secondary metabolites derived from mushrooms with inhibitory effects against HMG-CoA reductase and lipase enzyme. Using atorvastatin (IC50 = 32.1 μM) as the standard, 217 (IC50 = 29.8 μM), 221 (IC50 = 16.5 μM), 223 (IC50 = 30.3 μM), and 224 (IC50 = 14.3 μM) dramatically inhibited HMG-CoA [62]. Analyzing the experimental results of HMG-CoA activity, it was speculated that 243–245 were HMG-CoA reductase inhibitors and 245 (IC50 = 8.68 μM) had the best inhibitory activity against HMG-CoA, while atorvastatin (IC50 = 32.1 ± 7.7 μM) was used as a positive control [66]. Compared with the positive drug (orlistat), 246 exhibited inhibitory activity against pancreatic lipase [67]. Compounds 247–251 had prominent inhibitory activity on lipase [68]. In general, 224, 246, and 251 have great potential in the treatment of obesity.
2.9. Other Bioactivities
Edible and medicinal mushrooms can effectively produce useful bioactive metabolites. Due to their anti-tumor, anti-inflammatory, and antioxidant activities, these secondary metabolites have protective effects on organs such as the liver and kidneys. In addition, these secondary metabolites also have immunomodulatory activity and inhibitory effects on some cholinesterase and tyrosinase.
Compounds 17, 18, 20, and 22–24 exhibited inhibitory effects on acetylcholinesterase (AChE) at the concentration of 100 μg/mL. Moreover, Compounds 20 and 23 exhibited inhibitory effects on butrylcholinesterase (BchE) at 100 μg/mL. Compounds 18–20, 21, 23, 25, and 26 exhibited a suppressed effect on tyrosinase at 100 μg/mL [16]. Compounds 66 and 67 exerted immunosuppressive activity by inhibiting an LPS-induced proliferation of T cells at 20 μM [27]. Compounds 88–93 (MIC = 25.66 ± 2.84−55.28 ± 3.34 μg/mL) exhibited a moderate inhibitory effect on tyrosinase [33]. The results of lymphocyte proliferation test and the Concanavalin A-induced T lymphocyte mitogenic activity test showed that 252 had immune activity at 0.1 μM [69]. Compounds 253 (IC50 = 6.61 μM) and 254 (IC50 = 10.67 μM) significantly suppressed B lymphocyte cell proliferation induced by LPS [70]. Compounds 255–264 could reduce the activities of alanine aminotransferase and aspartate aminotransferase at 5−20 μM in HepG2 cells to exert liver-protective activity [71]. Compounds 265–268 (10 μM) alleviated cisplatin-induced nephrotoxicity by inhibiting the JNK-caspase-3 pathway [72]. Compound 269 exhibited inhibitory effect against AchE at 50 μM [73]. Compounds 270–274 (IC50 = 60.47 ± 2.63−148.38 ± 23.67 μM) could inhibit tyrosinase activity, and 274 (IC50 = 60.47 ± 2.63 μM) had the strongest activity (arbutin, MIC = 58.17 ± 6.09 μM, was used as the standard) [74]. Therefore, 66 and 253 show the best immunosuppressive effect. Compounds 255–268 have a protective effect on the liver or kidney, while Compounds 20, 269, and 223 exhibit potential activities to be used as AchE or BchE inhibitors. Similarly, it is speculated that 26 and 274 can serve as tyrosinase inhibitory agents.
3. Discussion
As early as prehistoric times, there were records of mushrooms as medicines [101]. Due to the rich nutritional content of mushrooms, they have always attracted scholars to deeply research the mechanisms behind their medicinal properties [102]. At present, there is considerable research on mushroom polysaccharides, and it has been reported that the immune regulation and anti-tumor effect of mushrooms mainly stem from mushroom polysaccharides [103]. With the continuous deepening of research on the composition of mushrooms, the secondary metabolites produced from mushrooms have also attracted scholars’ attention.
According to further data analysis, 61 secondary metabolites show hypoglycemic activity, 42 metabolites exhibit anti-tumor function, 40 metabolites have neuroprotective effects, 19 metabolites are able to inhibit microorganisms, 13 metabolites have hypolipidemic activity, 18 metabolites display inhibitory tyrosinase activity, 11 metabolites exhibit enzyme-inhibitory activity, 3 metabolites possess insecticidal activity, and 6 metabolites exhibit organ-protective functions. Furthermore, it is worth mentioning that oxidative stress and inflammatory responses are often key pathological processes in some acute and chronic diseases (Alzheimer’s disease, heart failure, and others) [104,105,106]. There are 35 and 37 secondary metabolites with antioxidant and anti-inflammatory effects, respectively. And their mechanisms may relate to TrkA/ERK1/2, Nrf2, BDNF/TrkB/ERK/CREB, TLR-4/NF-κB, and MAPK pathways. Summarizing the above, it can be hypothesized that mushrooms are important drug resources used to treat diabetes, cancer, neurological diseases, fungal and bacterial infectious diseases, and hyperlipidemia. However, it is undeniable that although these secondary metabolites reviewed in this paper have certain medicinal potential, further exploration is still needed to determine whether they produce side effects and have sufficient yield.
In addition, most of the edible and medicinal mushrooms included in this article come from Ganodermataceae, Hymenochaetaceae, Polyporaceae, and Hericiaceae, indicating that mushrooms from these families produce a wide range of pharmacologically active secondary metabolites. Overall, macrofungi is a rich source of bioactive secondary metabolites with a high potential for exploitation.
4. Conclusions
In recent years, relevant scholars have elaborated on the types of medicinal and edible mushrooms, the extraction techniques of bioactive components from mushrooms, and the pharmacological effects of active metabolites. The research on active ingredients focuses on primary metabolites (polysaccharides, proteins, polyunsaturated fatty acids) and nutritional components of mushrooms themselves, along with their development in food, medicine, and other fields [107,108,109]. This review mainly aims at the pharmacological activity and action mechanism of secondary metabolites derived from mushrooms. 274 secondary metabolites (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9) from 17 families of mushrooms, including 116 terpenoids, 71 sterols, 22 phenols, 14 polyketides, 9 alkaloids, and some aromatics and pyridines, as well as their biological activities, are reviewed. Among them, terpenoids and sterol-active metabolites accounted for the highest percentage. For example, Terpenoids 27, 28, and 65 possess significant anti-tumour activity; 69–72, 126, 129–131, and 134 show primary anti-inflammatory activity; compounds 35, 38, 40, 82, 123, 124, and 127 possess a significant hypoglycaemic effect. Sterols 147 and 148 exhibit anti-microbial activity, while 176–178 exhibit better anti-malarial activity.
In summary, the purpose of the current review is to provide a valuable theoretical reference for researchers to reasonably develop and utilize edible and medicinal mushrooms.
Author Contributions
Conceptualization, L.L.; methodology, X.S.; writing—original draft, X.S., Y.S. and D.S.; investigation, X.S., Y.S. and D.S.; formal analysis, X.S., Y.S., Y.T. and D.S.; project administration, L.L.; supervision, L.L.; funding acquisition, L.L; writing—review and editing, L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by grants from the National Key Research and Development Program of China (2022YFC2303100) and the National Natural Science Foundation of China (32022002 and 21977113).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Himanshi, R.; Shalinee, P.; Satyawati, S. Mushroom nutraceuticals for improved nutrition and better human health: A review. PharmaNutrition 2017, 5, 35–46. [Google Scholar]
- Sergey, G.; Ramin, R.; Josef, D.; Aristidis, T. Poisoning associated with the use of mushrooms: A review of the global pattern and main characteristics. Food Chem. Toxicol. 2019, 128, 267–279. [Google Scholar]
- El-Maradny, Y.A.; El-Fakharany, E.M.; Abu-Serie, M.M.; Hashish, M.H.; Selim, H.S. Lectins purified from medicinal and edible mushrooms: Insights into their antiviral activity against pathogenic viruses. Int. J. Biol. Macromol. 2021, 179, 239–258. [Google Scholar] [CrossRef]
- Chen, Y.J.; Jiang, S.; Jin, Y.X.; Yin, Y.L.; Yu, G.J.; Lan, X.Q.; Cui, M.Y.; Liang, Y.; Wong, B.H.C.; Guo, L.; et al. Purification and characterization of an antitumor protein with deoxyribonuclease activity from edible mushroom Agrocybe aegerita. Mol. Nutr. Food Res. 2012, 56, 1729–1738. [Google Scholar] [CrossRef]
- Citores, L.; Ragucci, S.; Ferreras, J.M.; Di Maro, A.; Iglesias, R. Ageritin, a Ribotoxin from poplar mushroom (Agrocybe aegerita) with defensive and antiproliferative activities. ACS Chem. Biol. 2019, 14, 1319–1327. [Google Scholar] [CrossRef]
- Kalaras, M.D.; Richie, J.P.; Calcagnotto, A.; Beelman, R.B. Mushrooms: A rich source of the antioxidants ergothioneine and glutathione. Food Chem. 2017, 233, 429–433. [Google Scholar] [CrossRef]
- Tong, Z.J.; Chu, G.D.; Wan, C.M.; Wang, Q.Y.; Yang, J.L.; Meng, Z.L.; Du, L.N.; Yang, J.; Ma, H.X. Multiple metabolites derived from mushrooms and their beneficial effect on Alzheimer’s diseases. Nutrients 2023, 15, 2758. [Google Scholar] [CrossRef]
- Martel, J.; Ojcius, D.M.; Chang, C.J.; Lin, C.S.; Lu, C.C.; Ko, Y.F.; Tseng, S.F.; Lai, H.C.; Young, J.D. Anti-obesogenic and antidiabetic effects of plants and mushrooms. Nat. Rev. Endocrinol. 2016, 13, 149–160. [Google Scholar] [CrossRef]
- Qi, J.S.; Duan, Y.; Li, Z.C.; Gao, J.M.; Qi, J.; Liu, C. The alkynyl-containing compounds from mushrooms and their biological activities. Nat. Prod. Bioprospect. 2023, 13, 50. [Google Scholar] [CrossRef]
- Wang, Y.X.; Wang, H.J.; Chai, K.Y.; Guo, S.Y.; Zhai, Y.Y.; Shi, R.; Zhang, F.Q.; Huang, J.Q.; Jin, Z.S.; Gao, Y.F.; et al. Systematic review and meta-analysis on the efficacy and safety of injectable lentinan combined with chemotherapy in the treatment of gastric cancer. Phytomedicine 2024, 123, 155242. [Google Scholar] [CrossRef]
- Zhou, X.Y.; Wang, H.Z.; Zhang, J.C.; Guan, Y.; Zhang, Y.J. Single-injection subunit vaccine for rabies prevention using lentinan as adjuvant. Int. J. Biol. Macromol. 2024, 254, 128118. [Google Scholar] [CrossRef]
- Meenu, M.; Xu, B. Application of vibrational spectroscopy for classification, authentication and quality analysis of mushroom: A concise review. Food Chem. 2019, 289, 545–557. [Google Scholar] [CrossRef]
- Lee, S.R.; Roh, H.S.; Lee, S.; Park, H.B.; Jang, T.S.; Ko, Y.J.; Baek, K.H.; Kim, K.H. Bioactivity-guided isolation and chemical characterization of antiproliferative constituents from morel mushroom (Morchella esculenta) in human lung adenocarcinoma cells. J. Funct. Foods 2018, 40, 249–260. [Google Scholar] [CrossRef]
- Satria, D.; Amen, Y.; Niwa, Y.; Ashour, A.; Allam, A.E.; Shimizu, K. Lucidumol D, a new lanostane-type triterpene from fruiting bodies of reishi (Ganoderma lingzhi). Nat. Prod. Rep. 2018, 33, 189–195. [Google Scholar] [CrossRef]
- Chen, H.P.; Zhao, Z.Z.; Li, Z.H.; Huang, Y.; Zhang, S.B.; Tang, Y.; Yao, J.N.; Chen, L.; Isaka, M.; Feng, T.; et al. Anti-Proliferative and anti-inflammatory lanostane triterpenoids from the polish edible mushroom Macrolepiota procera. J. Agric. Food Chem. 2018, 66, 3146–3154. [Google Scholar] [CrossRef] [PubMed]
- Deveci, E.; Tel-Çayan, G.; Duru, M.E.; Öztürk, M. Isolation, characterization, and bioactivities of compounds from Fuscoporia torulosa mushroom. J. Food Biochem. 2019, 43, e13074. [Google Scholar] [CrossRef]
- Wu, P.F.; Ding, R.; Tan, R.; Liu, J.; Hu, E.M.; Li, C.Y.; Liang, G.Y.; Yi, P. Sesquiterpenes from cultures of the fungus Phellinus igniarius and their cytotoxicities. Fitoterapia 2020, 140, 104415. [Google Scholar] [CrossRef]
- Chen, B.S.; Zhang, J.J.; Han, J.J.; Zhao, R.L.; Bao, L.; Huang, Y.; Liu, H.W. Lanostane triterpenoids with glucose-uptake-stimulatory activity from peels of the cultivated edible mushroom Wolfiporia cocos. J. Agric. Food Chem. 2019, 67, 7348–7364. [Google Scholar] [CrossRef]
- Zhao, Z.Z.; Liang, X.B.; Feng, W.S.; Wu, Y.; Zhi, Y.L.; Xue, G.M.; Chen, H.P.; Liu, J.K. Unusual constituents from the medicinal mushroom Ganoderma lingzhi. RSC Adv. 2019, 9, 36931–36939. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.; Ryoo, R.; Kim, J.C.; Park, H.B.; Kang, K.S.; Kim, K.H. Calvatianone, a sterol possessing a 6/5/6/5-fused ring system with a contracted tetrahydrofuran B-ring, from the fruiting bodies of Calvatia nipponica. J. Nat. Prod. 2020, 83, 2737–2742. [Google Scholar] [CrossRef]
- Su, L.H.; Geng, C.A.; Li, T.Z.; Huang, X.Y.; Ma, Y.B.; Zhang, X.M.; Wu, G.; Yang, Z.L.; Chen, J.J. Spiroseoflosterol, a rearranged ergostane-steroid from the fruiting bodies of Butyriboletus roseoflavus. J. Nat. Prod. 2020, 83, 1706–1710. [Google Scholar] [CrossRef]
- Li, X.C.; Liu, F.; Su, H.G.; Peng, C.; Zhou, Q.M.; Liu, J.; Huang, Y.J.; Guo, L.; Xiong, L. Twelve undescribed derivatives of ganoderic acid isolated from Ganoderma luteomarginatum and their cytotoxicity against three human cancer cell lines. Phytochemistry 2021, 183, 112617. [Google Scholar] [CrossRef]
- Menaga, D.; Rahman, P.K.S.M.; Rajakumar, S.; Ayyasamy, P.M. Antioxidant and cytotoxic activities of a novel isomeric molecule (PF5) obtained from methanolic extract of Pleurotus Florida mushroom. J. Biosci. Bioeng. 2021, 6, 338–349. [Google Scholar] [CrossRef]
- Hassan, K.; Matio, K.B.; Stadler, M. Two new triterpenes from basidiomata of the medicinal and edible mushroom, Laetiporus sulphureus. Molecules 2021, 26, 7090. [Google Scholar] [CrossRef]
- Ullah, Z.; Öztürk, M.; Ertaş, A.; Wahab, A.; Ben, M.R.; Iqbal Choudhary, M. Insight into isolation and elucidation of cytotoxic ergostanoids from the mushroom Sarcosphaera crassa (Santi) Pouzar: An edible mushroom. Steroids 2022, 181, 108990. [Google Scholar] [CrossRef] [PubMed]
- Gilardoni, G.; Negri, F.; Vita, F.P.; Hussain, F.H.S.; Vidari, G. New tricholidic acid triterpenoids from the mushroom Tricholoma ustaloides collected in an Italian beech wood. Molecules 2023, 28, 3864. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Z.; Zhang, F.; Ji, B.Y.; Zhou, N.; Chen, H.; Sun, Y.J.; Feng, W.S.; Zheng, X.K. Pyrrole alkaloids from the fruiting bodies of edible mushroom Lentinula edodes. RSC Adv. 2023, 13, 18223–18228. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Z.; Ji, B.Y.; Wang, Z.Z.; Si, Y.Y.; Sun, Y.J.; Chen, H.; Feng, W.S.; Zheng, X.K.; Liu, J.K. Lanostane triterpenoids with anti-proliferative and anti-inflammatory activities from medicinal mushroom Ganoderma lingzhi. Phytochemistry 2023, 213, 113791. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, X.; Lian, C.; Ke, J.; Liu, J. Triterpenes and aromatic meroterpenoids with antioxidant activity and neuroprotective effects from Ganoderma lucidum. Molecules 2019, 24, 4353. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhou, X.; Meng, Q.; Wang, M.; Zhang, Y.; Fu, S. Secondary metabolites and antiradical activity of liquid fermentation of Morchella sp. isolated from southwest China. Molecules 2019, 24, 1706. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, C.; Meng, Q.; Cui, Y.; Wang, Y.; Chen, X.; Fu, S. Investigation of chemical compounds and DPPH radical scavenging activity of Oudemansiella raphanipes (Agaricomycetes) based on fermentation. Int. J. Med. Mushrooms 2020, 22, 299–304. [Google Scholar] [CrossRef]
- Zhang, J.J.; Chen, B.S.; Dai, H.Q.; Ren, J.W.; Zhou, L.W.; Wu, S.H.; Liu, H.W. Sesquiterpenes and polyphenols with glucose-uptake stimulatory and antioxidant activities from the medicinal mushroom Sanghuangporus sanghuang. Chin. J. Nat. Med. 2021, 19, 693–699. [Google Scholar] [CrossRef]
- Lv, J.H.; Yao, L.; Duan, C.; Li, Z.; Zhang, J.; Li, C.T.; Li, Y. New bioactive α-pyrone from wild mushroom Paxillus involutus. Nat. Prod. Rep. 2021, 36, 2707–2712. [Google Scholar] [CrossRef]
- Lv, J.H.; Yao, L.; Zhang, J.X.; Wang, L.A.; Zhang, J.; Wang, Y.P.; Xiao, S.Y.; Li, C.T.; Li, Y. Novel 2,5-Diarylcyclopentenone derivatives from the wild edible mushroom Paxillus involutus and their antioxidant activities. J. Agric. Food Chem. 2021, 69, 5040–5048. [Google Scholar] [CrossRef]
- Ge, X.X.; Wang, Y.; Sun, C.X.; Zhang, Z.P.; Song, L.; Tan, L.L.; Li, D.H.; Yang, S.; Yu, G.H. Secondary metabolites produced by coculture of Pleurotus ostreatus SY10 and Pleurotus eryngii SY302. Chem. Biodivers. 2022, 19, e202100832. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.F.; Tan, J.; Jiang, R.X.; Li, F.F.; Zheng, R.Q.; Yu, L.J.; Luo, L.Z.; Zheng, Y.B. DPPH radical scavenging activity of new phenolics from the fermentation broth of mushroom Morehella importuna. Molecules 2023, 28, 4760. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.T.; Li, Y.C.; He, Y.D.; Meng, Q.F.; Ju, J.H.; Fu, S.B. Chemical constituents from mycelia of Lepista sordida (Agaricomycetes) and their ABTS radical scavenging activity. Int. J. Med. Mushrooms 2023, 25, 31–39. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.; Lee, S.O.; Ryu, J.Y.; Choi, S.Z.; Kang, K.S.; Kim, K.H. Anti-inflammatory activity of the sclerotia of edible fungus, Poria Cocos wolf and their active ianostane triterpenoids. J. Funct. Foods 2017, 32, 27–36. [Google Scholar] [CrossRef]
- Wu, Y.L.; Han, F.; Luan, S.S.; Ai, R.; Zhang, P.; Li, H.; Chen, L.X. Triterpenoids from Ganoderma lucidum and their potential anti-inflammatory effects. J. Agric. Food Chem. 2019, 67, 5147–5158. [Google Scholar] [CrossRef]
- Zhang, J.J.; Chen, B.S.; Liang, J.; Han, J.J.; Zhou, L.W.; Zhao, R.L.; Liu, H.W.; Dai, H.Q. Lanostane triterpenoids with PTP1B inhibitory and glucose-uptake stimulatory activities from mushroom Fomitopsis pinicola collected in north America. J. Agric. Food Chem. 2020, 68, 10036–10049. [Google Scholar] [CrossRef]
- Yin, X.; Tuong, T.M.L.; Tian, J.M.; Pescitelli, G.; Gao, J.M. Ganorbifates A and B from Ganoderma orbiforme, determined by DFT calculations of NMR data and ECD spectra. Chem. Commun. 2020, 56, 10195–10198. [Google Scholar] [CrossRef] [PubMed]
- Kou, R.W.; Han, R.; Gao, Y.Q.; Li, D.; Yin, X.; Gao, J.M. Anti-neuroinflammatory polyoxygenated lanostanoids from Chaga mushroom Inonotus obliquus. Phytochemistry 2021, 184, 112647. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.B.; Tang, L.; Xie, Y.; Xie, L.Y. Secondary metabolites from Hericium erinaceus and their anti-inflammatory activities. Molecules 2022, 27, 2157. [Google Scholar] [CrossRef] [PubMed]
- Khalilov, Q.; Numonov, S.; Sukhrobov, P.; Bobakulov, K.; Sharopov, F.; Habasi, M.; Zhao, J.; Yuan, T.; Aisa, H.A. New triterpenoids from the fruiting bodies of Laetiporus sulphureus and their anti-inflammatory activity. ACS Omega 2022, 7, 27272–27277. [Google Scholar] [CrossRef] [PubMed]
- Teng, L.; Wang, C.; Cui, B.; Zhang, J.; Zhou, S.; Pan, X.; Pan, F.; Dai, Y.; Feng, N. Lanostane triterpenoids from mycelia-associated Ganoderma sinense and their anti-inflammatory activity. Phytochemistry 2023, 215, 113870. [Google Scholar] [CrossRef]
- Isaka, M.; Chinthanom, P.; Sappan, M.; Supothina, S.; Vichai, V.; Danwisetkanjana, K.; Boonpratuang, T.; Hyde, K.D.; Choeyklin, R. Antitubercular activity of mycelium-associated Ganoderma lanostanoids. J. Nat. Prod. 2017, 80, 1361–1369. [Google Scholar] [CrossRef]
- Isaka, M.; Chinthanom, P.; Thummarukcharoen, T.; Boonpratuang, T.; Choowong, W. Highly modified lanostane triterpenes from fruiting bodies of the basidiomycete Tomophagus sp. J. Nat. Prod. 2019, 82, 1165–1176. [Google Scholar] [CrossRef]
- Sandargo, B.; Thongbai, B.; Stadler, M.; Surup, F. Cysteine-derived pleurotin congeners from the nematode-trapping basidiomycete Hohenbuehelia grisea. J. Nat. Prod. 2018, 81, 286–291. [Google Scholar] [CrossRef]
- Chepkirui, C.; Cheng, T.; Matasyoh, J.; Decock, C.; Stadler, M. An unprecedented spiro [furan-2,1′-indene]-3-one derivative and other nematicidal and antimicrobial metabolites from Sanghuangporus sp. (Hymenochaetaceae, Basidiomycota) collected in Kenya. Phytochem. Lett. 2018, 25, 141–146. [Google Scholar] [CrossRef]
- Cheng, T.; Chepkirui, C.; Decock, C.; Matasyoh, J.C.; Stadler, M. Sesquiterpenes from an eastern African medicinal mushroom belonging to the genus Sanghuangporus. J. Nat. Prod. 2019, 82, 1283–1291. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.H.; Ge, X.X.; Wang, Y.; Mo, X.H.; Yu, H.; Tan, L.L.; Yang, S. Discovery of novel terpenoids from the basidiomycete Pleurotus ostreatus through genome mining and coculture optimization. J. Agric. Food Chem. 2023, 71, 11110–11123. [Google Scholar] [CrossRef]
- Wahba, A.E.; El, S.A.; El, A.A.; Soliman, E.M. New antimalarial lanostane triterpenes from a new isolate of Egyptian Ganoderma species. Med. Chem. Res. 2019, 28, 2246–2251. [Google Scholar] [CrossRef]
- Isaka, M.; Chinthanom, P.; Vichai, V.; Sommai, S.; Choeyklin, R. Ganoweberianones A and B, antimalarial lanostane dimers from cultivated fruiting bodies of the basidiomycete Ganoderma weberianum. J. Nat. Prod. 2020, 83, 3404–3412. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.C.; Cao, C.Y.; Kubo, M.; Harada, K.; Yan, X.T.; Fukuyama, Y.; Gao, J.M. Chemical constituents from Hericium erinaceus promote neuronal survival and potentiate neurite outgrowth via the TrkA/Erk1/2 pathway. Int. J. Mol. Sci. 2017, 18, 1659. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Zhang, J.J.; Ren, J.W.; Wang, W.Z.; Xiong, W.P.; Zhang, Y.D.; Bao, L.; Liu, H.W. Triterpenes and meroterpenes with neuroprotective effects from Ganoderma leucocontextum. Chem. Biodivers. 2018, 15, e1700567. [Google Scholar] [CrossRef] [PubMed]
- Rupcic, Z.; Rascher, M.; Kanaki, S.; Köster, R.; Stadler, M.; Wittstein, K. Two new cyathane diterpenoids from mycelial cultures of the medicinal mushroom Hericium erinaceus and the rare species, Hericium flagellum. Int. J. Mol. Sci. 2018, 19, 740. [Google Scholar] [CrossRef] [PubMed]
- Kou, R.W.; Du, S.T.; Xia, B.; Zhang, Q.; Yin, X.; Gao, J.M. Phenolic and steroidal metabolites from the cultivated edible Inonotus hispidus mushroom and their bioactivities. J. Agric. Food Chem. 2021, 69, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Kou, R.W.; Xia, B.; Wang, Z.J.; Li, J.N.; Yang, J.R.; Gao, Y.Q.; Yin, X.; Gao, J.M. Triterpenoids and meroterpenoids from the edible Ganoderma resinaceum and their potential anti-inflammatory, antioxidant and anti-apoptosis activities. Bioorg. Chem. 2022, 121, 105689. [Google Scholar] [CrossRef] [PubMed]
- Kou, R.W.; Xia, B.; Han, R.; Li, Z.Q.; Yang, J.R.; Yin, X.; Gao, Y.Q.; Gao, J.M. Neuroprotective effects of a new triterpenoid from edible mushroom on oxidative stress and apoptosis through the BDNF/TrkB/ERK/CREB and Nrf2 signaling pathway in vitro and in vivo. Food Funct. 2022, 13, 12121–12134. [Google Scholar] [CrossRef] [PubMed]
- Sum, W.C.; Ebada, S.S.; Kirchenwitz, M.; Kellner, H.; Ibrahim, M.A.A.; Stradal, T.E.B.; Matasyoh, J.C.; Stadler, M. Hericioic acids A–G and hericiofuranoic acid; neurotrophic agents from cultures of the european mushroom Hericium flagellum. J. Agric. Food Chem. 2023, 71, 11094–11103. [Google Scholar] [CrossRef]
- Zhang, Y.; Xun, H.; Gao, Q.; Qi, F.; Sun, J.; Tang, F. Chemical constituents of the mushroom Dictyophora indusiata and their anti-inflammatory activities. Molecules 2023, 28, 2760. [Google Scholar] [CrossRef]
- Chen, B.S.; Tian, J.; Zhang, J.J.; Wang, K.; Liu, L.; Yang, B.; Bao, L.; Liu, H.W. Triterpenes and meroterpenes from Ganoderma lucidum with inhibitory activity against HMGs reductase, aldose reductase and α-glucosidase. Fitoterapia 2017, 120, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, Z.H.; Yao, J.N.; Peng, Y.L.; Huang, R.; Feng, T.; Liu, J.K. Isoindolinone-containing meroterpenoids with α-glucosidase inhibitory activity from mushroom Hericium caput-medusae. Fitoterapia 2017, 122, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Ryu, S.H.; Turk, A.; Yeon, S.W.; Jo, Y.H.; Han, Y.K.; Hwang, B.Y.; Lee, K.Y.; Lee, M.K. Characterization of α-glucosidase inhibitory constituents of the fruiting body of lion’s mane mushroom (Hericium erinaceus). J. Ethnopharmacol. 2020, 262, 113197. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.H.; Yao, L.; Li, D.; Jia, C.W.; Zhang, J.X.; Wang, L.A.; Li, C.T.; Li, Y. Novel hypoglycemic compounds from wild mushroom Paxillus involutus. Bioorg. Chem. 2021, 112, 104984. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Ma, K.; Han, J.J.; Wang, K.; Chen, H.Y.; Bao, L.; Liu, L.; Xiong, W.P.; Zhang, Y.D.; Huang, Y.; et al. Eight new triterpenoids with inhibitory activity against HMG-CoA reductase from the medical mushroom Ganoderma leucocontextum collected in Tibetan plateau. Fitoterapia 2018, 130, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.L.; Meng, Q.F.; Zhang, Y.; Hu, Y.; Xiao, S.J.; Zhang, Y.Q.; Ju, J.H.; Fu, S.B. Morelsins A–F, six sesquiterpenoids from the liquid culture of Morchella importuna. Tetrahedron 2020, 76, 31–32. [Google Scholar] [CrossRef]
- Meng, Q.F.; Hu, W.T.; Wang, H.; Wu, L.H.; Yang, C.L.; Zhou, X.; Tian, Y.; Fu, S. One new compound from fruiting body of cultivated caterpillar medicinal mushroom, Cordyceps militaris (ascomycetes), and its lipase inhibitory activity. Int. J. Med. Mushrooms 2021, 23, 9–16. [Google Scholar] [CrossRef]
- Pu, D.B.; Zheng, X.; Gao, J.B.; Zhang, X.J.; Qi, Y.; Li, X.S.; Wang, Y.M.; Li, X.N.; Li, X.L.; Wan, C.P.; et al. Highly oxygenated lanostane-type triterpenoids and their bioactivity from the fruiting body of Ganoderma gibbosum. Fitoterapia 2017, 119, 1–7. [Google Scholar] [CrossRef]
- Zhang, S.B.; Huang, Y.; He, S.J.; Chen, H.P.; Wu, B.; Li, S.Y.; Zhao, Z.Z.; Li, Z.H.; Wang, X.; Zuo, J.P.; et al. Heterocyclic compounds from the mushroom Albatrellus confluens and their inhibitions against lipopolysaccharides-induced B lymphocyte cell proliferation. J. Org. Chem. 2018, 83, 10158–10165. [Google Scholar] [CrossRef]
- Shi, Q.Q.; Huang, Y.J.; Su, H.G.; Gao, Y.; Lu, S.Y.; Peng, X.R.; Li, X.N.; Zhou, L.; Qiu, M.H. Structurally diverse lanostane triterpenoids from medicinal and edible mushroom Ganoderma resinaceum boud. Bioorg. Chem. 2020, 100, 103871. [Google Scholar] [CrossRef]
- Lee, S.R.; Lee, D.; Lee, H.J.; Noh, H.J.; Jung, K.; Kang, K.S.; Kim, K.H. Renoprotective chemical constituents from an edible mushroom, Pleurotus cornucopiae in cisplatin-induced nephrotoxicity. Bioorg. Chem. 2017, 71, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gao, J.; Chen, Q.B.; Liu, W.; Qi, Y.R.; Aisa, H.A.; Yuan, T. Applanaic acids A–C, three new highly oxygenated lanostane triterpenoids from the fruiting bodies of Ganoderma applanatum. Nat. Prod. Res. 2020, 35, 3918–3924. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Jia, J.H.; Wang, Q.; Wei, Y.L.; Yuan, H.S. Secondary metabolites from the cultures of medicinal mushroom Vanderbylia robiniophila and their tyrosinase inhibitory activities. J. Fungi 2023, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Nounou, M.I.; ElAmrawy, F.; Ahmed, N.; Abdelraouf, K.; Goda, S.; Syed-Sha-Qhattal, H. Breast cancer: Conventional diagnosis and treatment modalities and recent patents and technologies. Breast Cancer Res. 2015, 9 (Suppl. S2), 17–34. [Google Scholar] [CrossRef] [PubMed]
- Li, B.Q.; Shao, H.L.; Gao, L.; Li, H.; Sheng, H.G.; Zhu, L.Q. Nano-drug co-delivery system of natural active ingredients and chemotherapy drugs for cancer treatment: A review. Drug Deliv. 2022, 29, 2130–2161. [Google Scholar] [CrossRef]
- Teleanu, D.M.; Niculescu, A.G.; Lungu, I.I.; Radu, C.I.; Vladâcenco, O.; Roza, E.; Costăchescu, B.; Grumezescu, A.M.; Teleanu, R.I. An overview of oxidative stress, neuroinflammation, and neurodegenerative diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.; Maestre, A.B.; Hernández, R.J.; Arnao, M.B. ABTS/TAC methodology: Main milestones and recent applications. Processes 2023, 11, 185. [Google Scholar] [CrossRef]
- Hou, C.Y.; Chen, L.L.; Yang, L.Z.; Ji, X.L. An insight into anti-inflammatory effects of natural polysaccharides. Int. J. Biol. Macromol. 2020, 153, 248–255. [Google Scholar] [CrossRef]
- Alessandri, A.L.; Sousa, L.P.; Lucas, C.D.; Rossi, A.G.; Pinho, V.; Teixeira, M.M. Resolution of inflammation: Mechanisms and opportunity for drug development. Chin. J. Nat. Med. 2013, 139, 189–212. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liu, T.W.; Wallig, M.A.; Dobrucki, I.T.; Dobrucki, L.W.; Nelson, E.R.; Swanson, K.S.; Smith, A.M. Efficient targeting of adipose tissue macrophages in obesity with polysaccharide nanocarriers. ACS Nano 2016, 10, 6952–6962. [Google Scholar] [CrossRef] [PubMed]
- Suleyman, G.; Alangaden, G.J. Nosocomial fungal infections: Epidemiology, infection control, and prevention. Infect. Dis. Clin. N. Am. 2021, 35, 1027–1053. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Pang, X.; Zhang, H.; Ji, P. The cGAS-STING pathway in bacterial infection and bacterial immunity. Front. Immunol. 2021, 12, 814709. [Google Scholar] [CrossRef]
- McEwen, S.A.; Collignon, P.J. Antimicrobial resistance: A one health perspective. Microbiol. Spectr. 2018, 6, 521–547. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Efferth, T. Cameroonian medicinal plants: Pharmacology and derived natural products. Front. Pharmacol. 2010, 1, 123. [Google Scholar] [CrossRef]
- Alaithan, H.; Kumar, N.; Islam, M.Z.; Liappis, A.P.; Nava, V.E. Novel therapeutics for malaria. Pharmaceutics 2023, 15, 1800. [Google Scholar] [CrossRef]
- Racca, F.; Vianello, A.; Mongini, T.; Ruggeri, P.; Versaci, A.; Vita, G.L.; Vita, G. Practical approach to respiratory emergencies in neurological diseases. Neurol. Sci. 2020, 41, 497–508. [Google Scholar] [CrossRef]
- Yang, Q.Q.; Zhou, J.W. Neuroinflammation in the central nervous system: Symphony of glial cells. Glia 2019, 67, 1017–1035. [Google Scholar] [CrossRef]
- Kshirsagar, R.P.; Kulkarni, A.A.; Chouthe, R.S.; Pathan, S.K.; Une, H.D.; Reddy, G.B.; Diwan, P.V.; Ansari, S.A.; Sangshetti, J.N. SGLT inhibitors as antidiabetic agents: A comprehensive review. RSC Adv. 2020, 10, 1733–1756. [Google Scholar] [CrossRef]
- Thakur, S.; Gupta, S.K.; Ali, V.; Singh, P.; Verma, M. Aldose Reductase: A cause and a potential target for the treatment of diabetic complications. Arch. Pharm. Res. 2021, 44, 655–667. [Google Scholar] [CrossRef]
- Abduldileep, S.; Narayanasamy, R.; Usharani, D.; Singh, A.; Rajasekharan, R. A bioactive polypeptide from sugarcane selectively inhibits intestinal sucrase. Int. J. Biol. Macromol. 2020, 156, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Hossain, U.; Das, A.K.; Ghosh, S.; Sil, P.C. An overview on the role of bioactive α-glucosidase inhibitors in ameliorating diabetic complications. Food Chem. Toxicol. 2020, 145, 111738. [Google Scholar] [CrossRef]
- Sharma, B.; Xie, L.; Yang, F.; Wang, W.; Zhou, Q.; Xiang, M.; Zhou, S.; Lv, W.; Jia, Y.; Pokhrel, L.; et al. Recent advance on PTP1B inhibitors and their biomedical applications. Eur. J. Med. Chem. 2020, 199, 112376. [Google Scholar] [CrossRef]
- Ghani, U. Re-exploring promising α-glucosidase inhibitors for potential development into oral anti-diabetic drugs: Finding needle in the haystack. Eur. J. Med. Chem. 2015, 103, 133–162. [Google Scholar] [CrossRef] [PubMed]
- He, J.Y.; Hong, Q.; Chen, B.X.; Cui, S.Y.; Liu, R.; Cai, G.Y.; Guo, J.; Chen, X.M. Ginsenoside Rb1 alleviates diabetic kidney podocyte injury by inhibiting aldose reductase activity. Acta Pharmacol. Sin. 2022, 43, 342–353. [Google Scholar] [CrossRef]
- Kenner, K.A.; Anyanwu, E.; Olefsky, J.M.; Kusari, J. Protein-tyrosine phosphatase 1B is a negative regulator of insulin- and insulin-like growth factor-I-stimulated signaling. J. Biol. Chem. 1996, 271, 19810–19816. [Google Scholar] [CrossRef] [PubMed]
- Perdomo, C.M.; Cohen, R.V.; Sumithran, P.; Clément, K.; Frühbeck, G. Contemporary medical, device, and surgical therapies for obesity in adults. Lancet. 2023, 401, 1116–1130. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.S.; Li, L.X.; Liang, N.N.; Zhang, L.L.; Xu, X.D.; Chen, S.T.; Yin, H.Y. Acetaldehyde dehydrogenase 2 regulates HMG-CoA reductase stability and cholesterol synthesis in the liver. Redox Biol. 2021, 41, 101919. [Google Scholar] [CrossRef]
- Bialecka-Florjanczyk, E.; Fabiszewska, A.U.; Krzyczkowska, J.; Kurylowicz, A. Synthetic and natural lipase inhibitors. Mini. Rev. Med. Chem. 2018, 18, 672–683. [Google Scholar] [CrossRef]
- Pennerman, K.K.; Yin, G.; Bennett, J.W. Health effects of small volatile compounds from East Asian medicinal mushrooms. Mycobiology 2015, 43, 9–13. [Google Scholar] [CrossRef]
- Wang, X.M.; Zhang, J.; Wu, L.H.; Zhao, Y.L.; Li, T.; Li, J.Q.; Wang, Y.Z.; Liu, H.G. A mini-review of chemical composition and nutritional value of edible wild-grown mushroom from China. Food Chem. 2014, 151, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; de Mores, A.R.; Cohen, L.; Anwar, M.M.; Lazar, F.; Hicklen, R.; Lopez, G.; Yang, P.; Bruera, E. Medicinal mushroom supplements in cancer: A systematic review of clinical studies. Curr. Oncol. Rep. 2023, 25, 569–587. [Google Scholar] [CrossRef] [PubMed]
- van der Pol, A.; van Gilst, W.H.; Voors, A.A.; van der Meer, P. Treating oxidative stress in heart failure: Past, present and future. Eur. J. Heart. Fail. 2019, 21, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Marek-Jozefowicz, L.; Nedoszytko, B.; Grochocka, M.; Żmijewski, M.A.; Czajkowski, R.; Cubała, W.J.; Slominski, A.T. Molecular mechanisms of neurogenic inflammation of the skin. Int. J. Mol. Sci. 2023, 24, 5001. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Guo, J.; Ye, X.Y.; Xie, Y.; Xie, T. Oxidative stress: The core pathogenesis and mechanism of Alzheimer’s disease. Ageing Res. Rev. 2022, 77, 101619. [Google Scholar] [CrossRef] [PubMed]
- Bhambri, A.; Srivastava, M.; Mahale, V.G.; Mahale, S.; Karn, S.K. Mushrooms as potential sources of active metabolites and medicines. Front. Microbiol. 2022, 26, 837266. [Google Scholar] [CrossRef]
- Kumar, K.; Mehra, R.; Guiné, R.P.F.; Lima, M.J.; Kumar, N.; Kaushik, R.; Ahmed, N.; Yadav, A.N.; Kumar, H. Edible mushrooms: A comprehensive review on bioactive compounds with health benefits and processing aspects. Foods 2021, 10, 2996. [Google Scholar] [CrossRef]
- Estella, T.F.; Mbong, G.A.; Lem, E.A.; Manju, E.B.; Tchadji, M.V.; Dobgima, J.F.; Charles, F. Medicinal mushroom of potential pharmaceutical toxic importance: Contribution in phytotherapy. In Current Topics in Functional Food; IntechOpen Limited: London, UK, 2022. [Google Scholar]
Figure 1.
Structures of compounds 1–46.
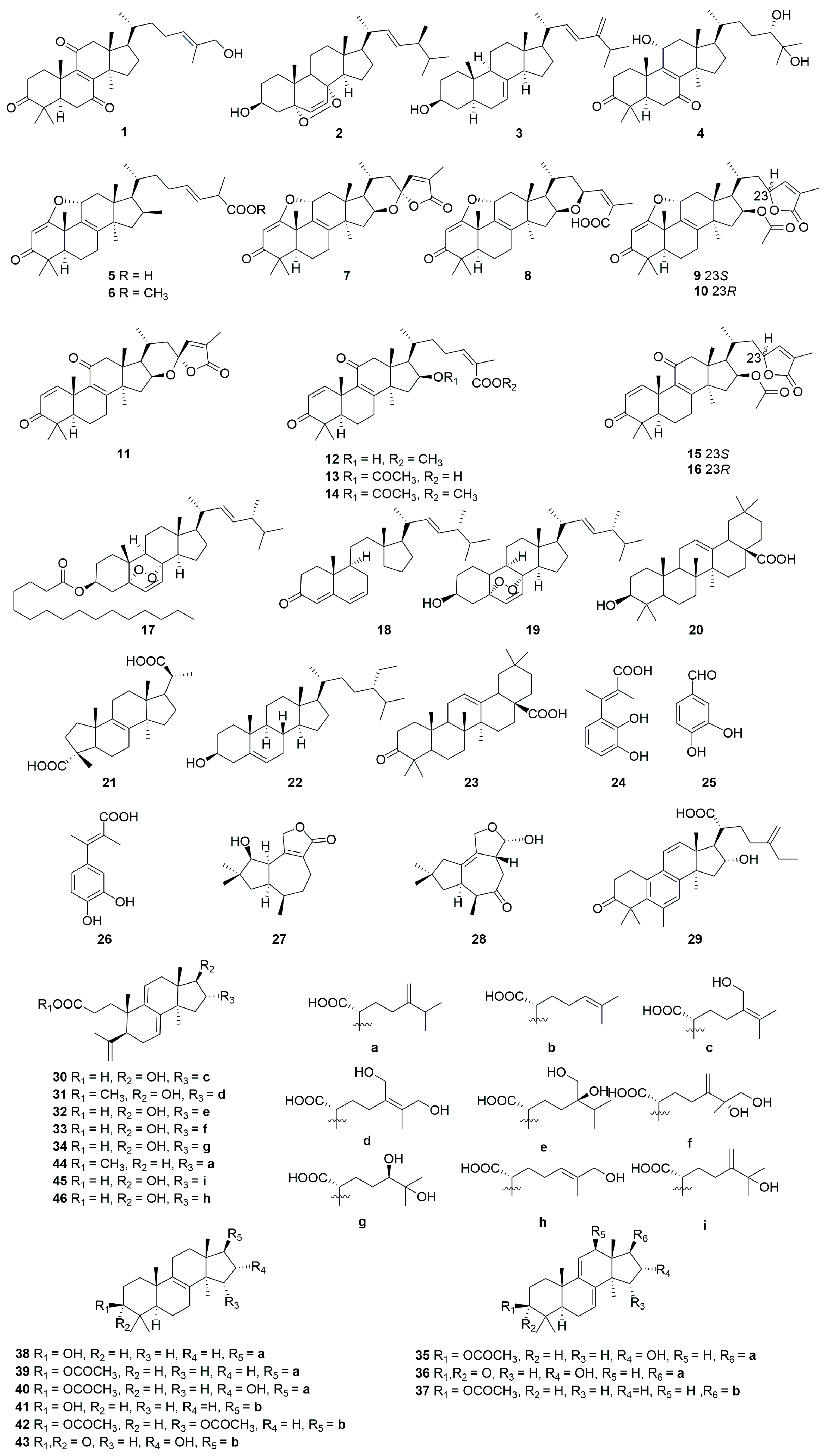
Figure 2.
Structures of compounds 47–82.

Figure 3.
Structures of compounds 83–118.
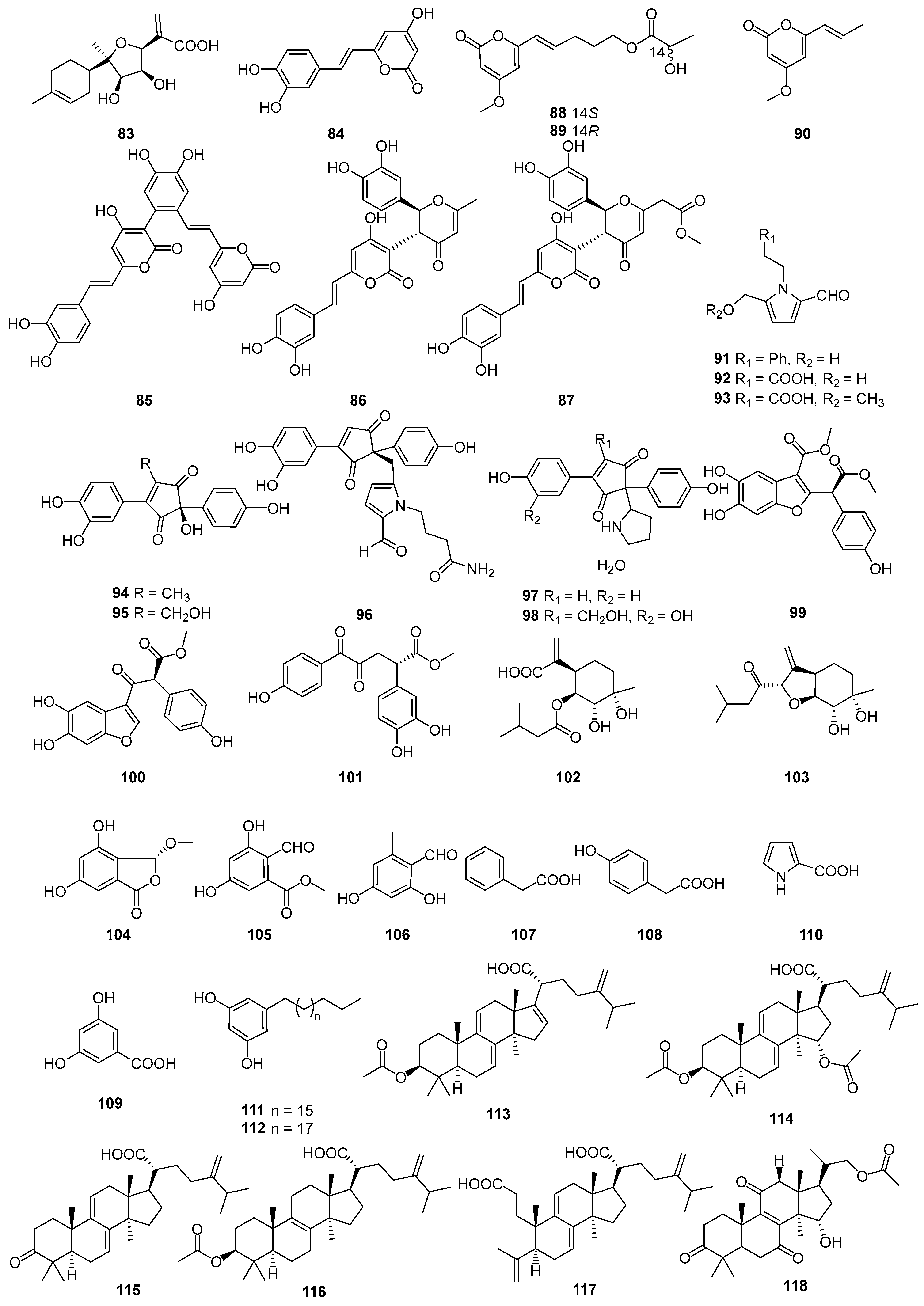
Figure 4.
Structures of compounds 119–148.

Figure 5.
Structures of compounds 149–170.
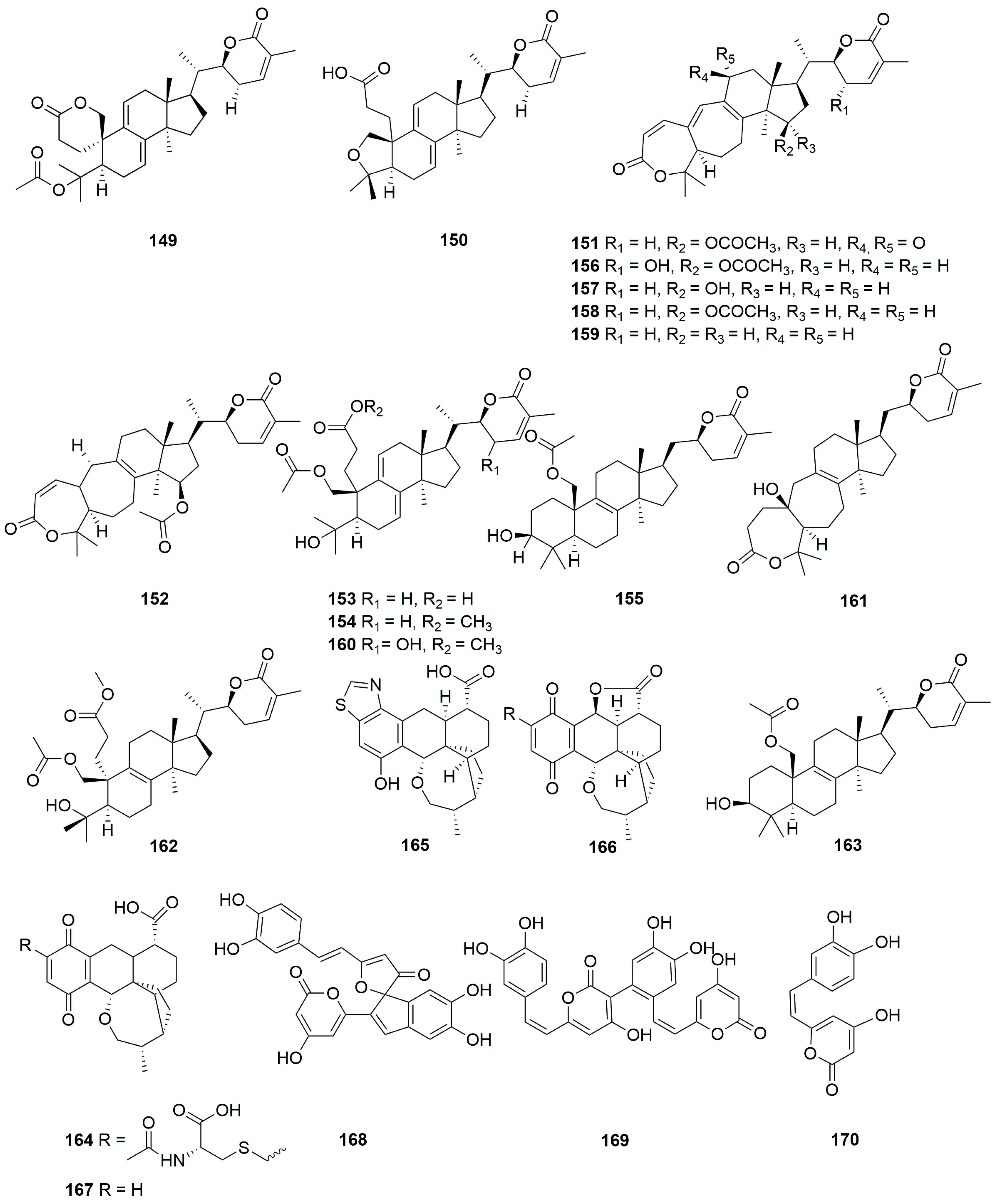
Figure 6.
Structures of compounds 171–197.
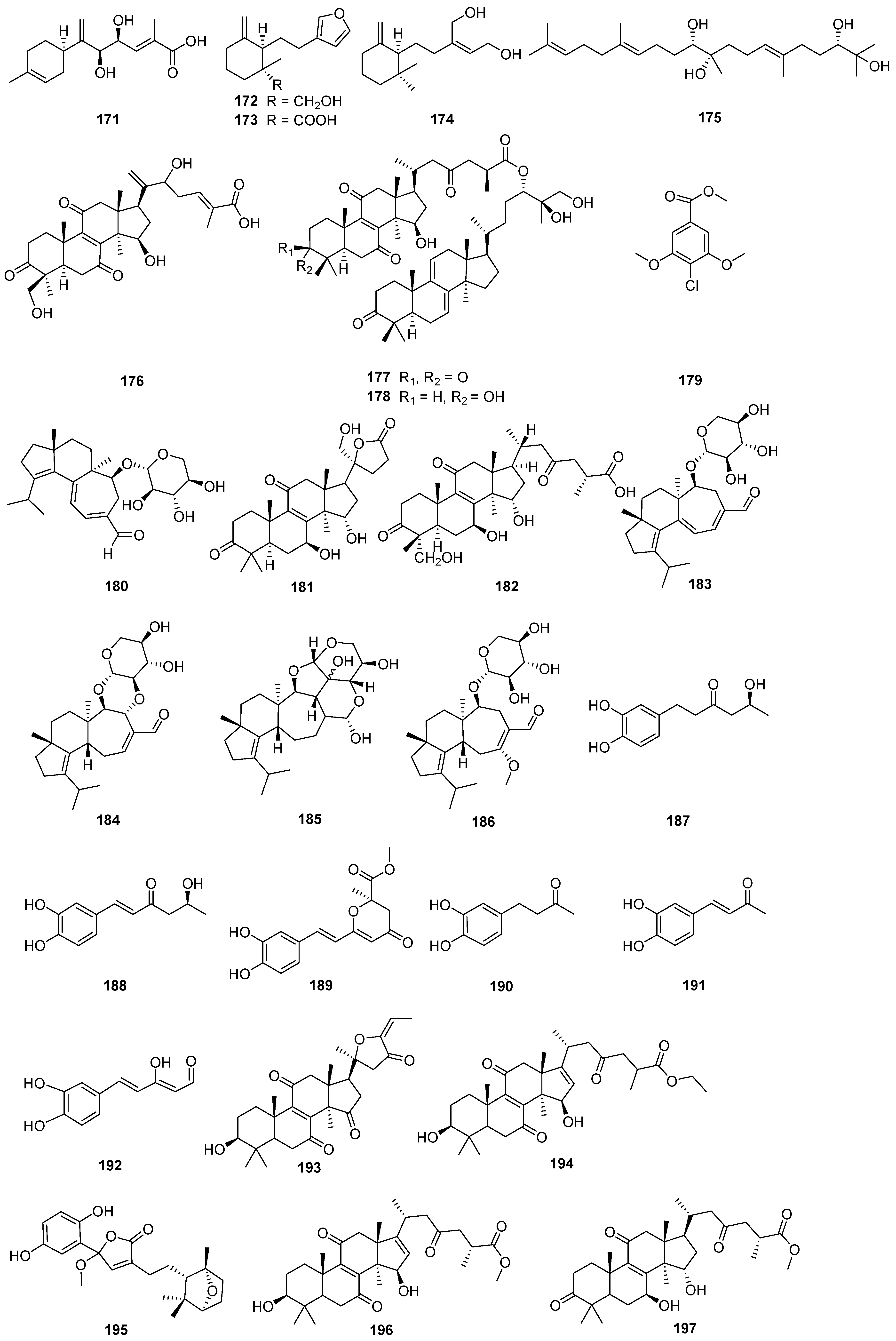
Figure 7.
Structures of compounds 198–224.
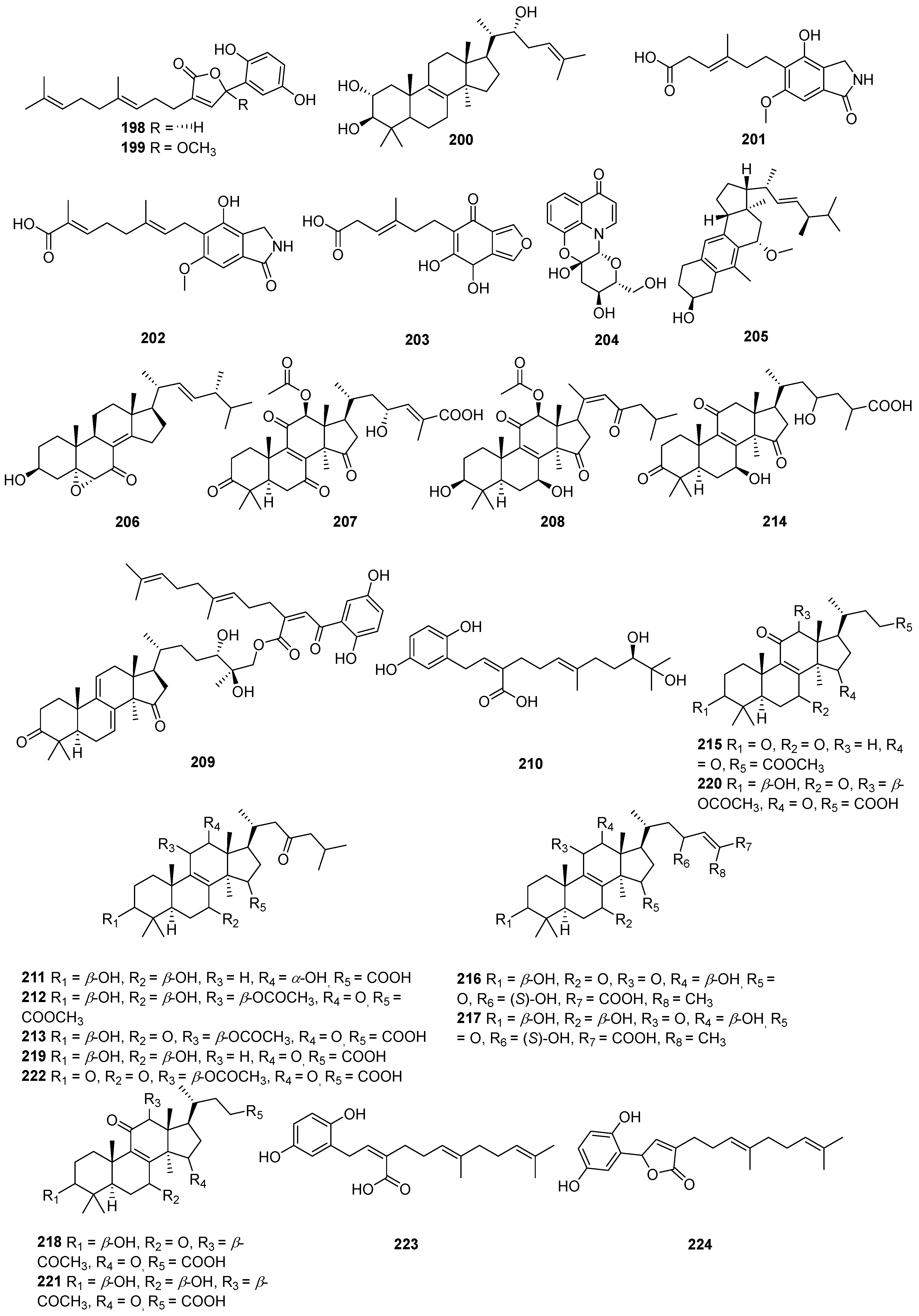
Figure 8.
Structures of compounds 225–247.

Figure 9.
Structures of compounds 248–274.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Different species of mushrooms and their secondary metabolites.
| Mushroom | Mushroom Family | Secondary Metabolites | Refs. |
|---|---|---|---|
| Morchella esculenta | Morchellaceae | 1–3 | [13] |
| Ganoderma lingzhi | Ganodermataceae | 4 | [14] |
| Macrolepiota procera | Agaricaceae | 5–16 | [15] |
| Fuscoporia torulosa | Hymenochaetaceae | 17–26 | [16] |
| Phellinus igniarius | Polyporaceae | 27–28 | [17] |
| Wolfiporia cocos | Polyporaceae | 29–46 | [18] |
| Ganoderma lingzhi | Ganodermataceae | 47 | [19] |
| Calvatia nipponica | Lycoperdaceae | 48–52 | [20] |
| Butyriboletus roseoflavus | Boletaceae | 53 | [21] |
| Ganoderma luteomarginatum | Ganodermataceae | 54–56 | [22] |
| Pleurotus florida | Pleurotaceae | 57 | [23] |
| Laetiporus sulphureus | Polyporaceae | 58–59 | [24] |
| Sarcosphaera crassa | Pezizaceae | 60–63 | [25] |
| Tricholoma ustaloides | Tricholomataceae | 64–65 | [26] |
| Lentinula edodes | Omphalotaceae | 66–68 | [27] |
| Ganoderma | Ganodermataceae | 69–72 | [28] |
| Ganoderma lucidum | Ganodermataceae | 73–75 | [29] |
| Morchella importuna | Morchellaceae | 76–78 | [30] |
| Oudemansiella raphanipes | Tricholomataceae | 79–81 | [31] |
| Sanghuangporus sanghuang | Hymenochaetaceae | 82–87 | [32] |
| Paxillus involutus | Paxillaceae | 88–93 | [33] |
| Paxillus involutus | Paxillaceae | 94–101 | [34] |
| Pleurotus ostreatus and Pleurotus eryngii | Pleurotaceae | 102–103 | [35] |
| Morehella importuna | Morchellaceae | 104–110 | [36] |
| Lepista sordida | Tricholomataceae | 111–112 | [37] |
| Poria cocos Wolf | Polyporaceae | 113–117 | [38] |
| Ganoderma lucidum | Ganodermataceae | 118 | [39] |
| Fomitopsis pinicola | Coriolaceae | 119–127 | [40] |
| Ganoderma orbiforme | Ganodermataceae | 128–129 | [41] |
| Inonotus obliquus | Hymenochaetaceae | 130–136 | [42] |
| Hericium erinaceus | Hericiaceae | 137–138 | [43] |
| Laetiporus sulphureus | Polyporaceae | 139–141 | [44] |
| Ganoderma sinense | Ganodermataceae | 142–143 | [45] |
| Ganoderma | Ganodermataceae | 144–148 | [46] |
| Tomophagus sp. | Ganodermataceae | 149–163 | [47] |
| Hohenbuehelia grisea | Pleurotaceae | 164–167 | [48] |
| Sanghuangporus | Hymenochaetaceae | 168–170 | [49] |
| Sanghuangporus | Hymenochaetaceae | 171–174 | [50] |
| Pleurotus ostreatus and Trametes robiniophila | Pleurotaceae and Polyporaceae | 175 | [51] |
| Ganoderma | Ganodermataceae | 176 | [52] |
| Ganoderma weberianum | Ganodermataceae | 177–178 | [53] |
| Hericium erinaceus | Hericiaceae | 179–180 | [54] |
| Ganoderma leucocontextum | Ganodermataceae | 181–182 | [55] |
| Hericium erinaceus and Hericium flagellum | Hericiaceae | 183–186 | [56] |
| Inonotus hispidus | Hymenochaetaceae | 187–192 | [57] |
| Ganoderma resinaceum | Ganodermataceae | 193–199 | [58] |
| Inonotus obliquus | Hymenochaetaceae | 200 | [59] |
| Hericium flagellum | Hericiaceae | 201–203 | [60] |
| Dictyophora indusiata | Phallaceae | 204–206 | [61] |
| Ganoderma lucidum | Ganodermataceae | 207–230 | [62] |
| Hericium caput−medusae | Hericiaceae | 231–233 | [63] |
| Hericium erinaceus | Hericiaceae | 234–238 | [64] |
| Paxillus involutus | Paxillaceae | 239–242 | [65] |
| Ganoderma leucocontextum | Ganodermataceae | 243–245 | [66] |
| Morchella importuna | Morchellaceae | 246 | [67] |
| Cordyceps militaris | Cordycepitaceae | 247–251 | [68] |
| Ganoderma gibbosum | Ganodermataceae | 252 | [69] |
| Albatrellus confluens | Crypsinus | 253–254 | [70] |
| Ganoderma resinaceum | Ganodermataceae | 255–264 | [71] |
| Pleurotus cornucopiae | Pleurotaceae | 265–268 | [72] |
| Ganoderma applanatum | Ganodermataceae | 269 | [73] |
| Vanderbylia robiniophila | Polyporaceae | 270–274 | [74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sun, X.; Shi, Y.; Shi, D.; Tu, Y.; Liu, L. Biological Activities of Secondary Metabolites from the Edible-Medicinal Macrofungi. J. Fungi 2024, 10, 144. https://doi.org/10.3390/jof10020144
AMA Style
Sun X, Shi Y, Shi D, Tu Y, Liu L. Biological Activities of Secondary Metabolites from the Edible-Medicinal Macrofungi. Journal of Fungi. 2024; 10(2):144. https://doi.org/10.3390/jof10020144
Chicago/Turabian StyleSun, Xiaoqi, Ying Shi, Dongxiao Shi, Yu Tu, and Ling Liu. 2024. "Biological Activities of Secondary Metabolites from the Edible-Medicinal Macrofungi" Journal of Fungi 10, no. 2: 144. https://doi.org/10.3390/jof10020144
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.