
Eosinophilic inflammation: An Appealing Target for Pharmacologic Treatments in Severe Asthma

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
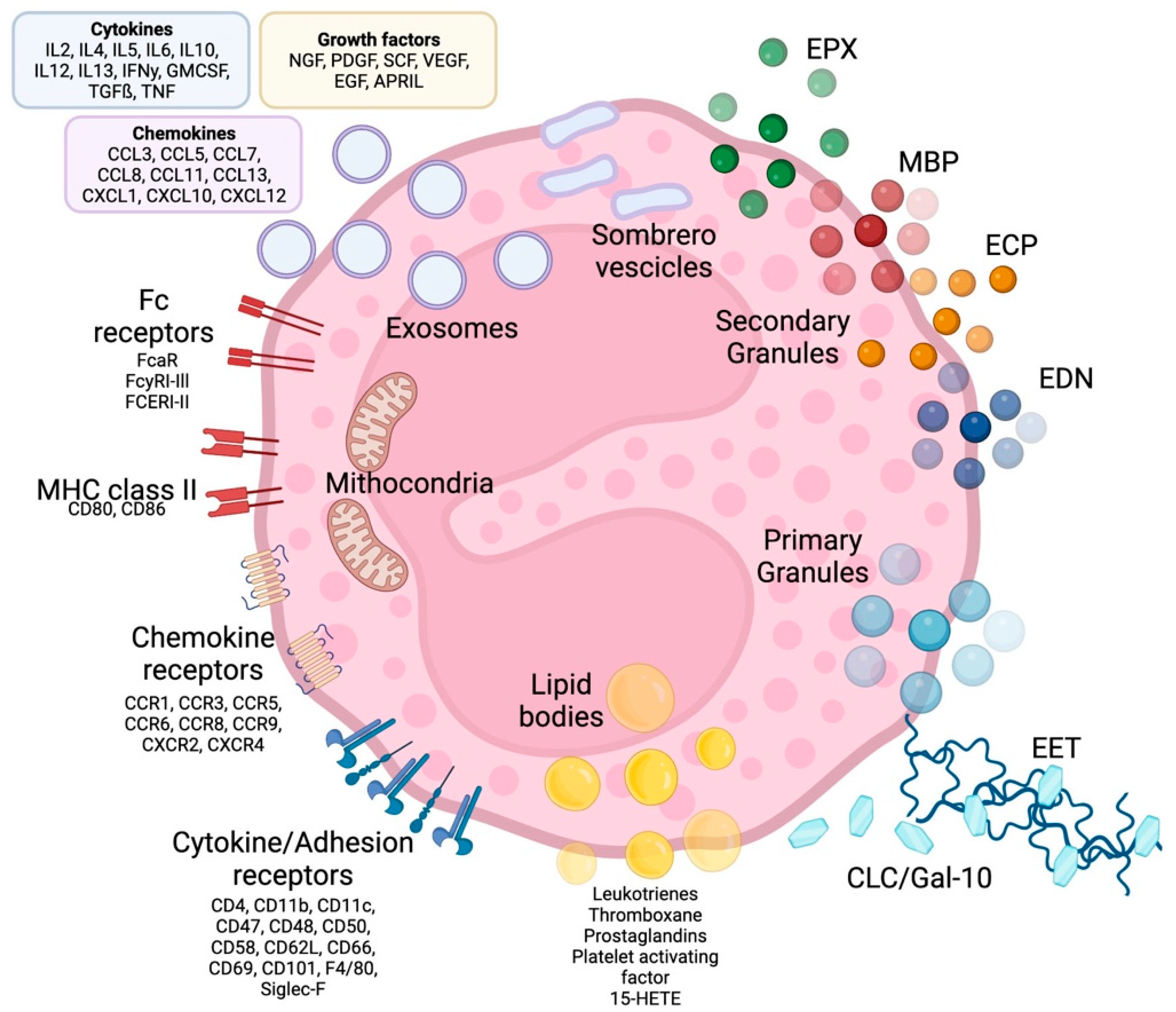
2. Eosinophil Structure and Biology
2.1. Eosinophil Structure
2.1.1. Eosinophil Granules
2.1.2. Eosinophil Surface Receptors
- Cytokine Receptors
- Adhesion Receptors
- Chemoattractant Receptors
- Fc Receptors
- Major Histocompatibility Complex-II
- Pattern Recognition Receptors (PRRs)
- Lipid Mediator Receptors
- Siglec-8
- Inhibitory Receptors
2.1.3. Intracellular Receptors
2.2. Eosinophil Biology
2.2.1. Eosinophil Differentiation and Maturation
2.2.2. Eosinophil Migration and Activation
2.2.3. Eosinophil Degranulation
2.3. Eosinophil Heterogeneity
2.4. Overview of Eosinophil-Driven Pathological Conditions
3. Eosinophils in Pathobiology of Severe Asthma
4. Targeting Eosinophils in Severe Asthma
4.1. Corticosteroids
4.2. Anti-Eosinophil Biological Therapies
4.2.1. IL-5 Inhibition
4.2.2. IL-5 Receptor Antagonism
4.2.3. Currently Available Biological Therapies Indirectly Targeting Eosinophils
4.3. Experimental Anti-Eosinophil Therapies
4.3.1. Chemokine Receptor Antagonists
4.3.2. Antagonists of Prostaglandin D2 Receptor
4.3.3. Inducers of Eosinophil Apoptosis
4.3.4. Inhibitors of Kinases and Transcription Factors
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Papi, A.; Brightling, C.; Pedersen, S.E.; Reddel, H.K. Asthma. Lancet 2018, 391, 783–800. [Google Scholar] [CrossRef]
- Wenzel, S.E. Asthma phenotypes: The evolution from clinical to molecular approaches. Nat. Med. 2012, 18, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Vatrella, A.; Maglio, A.; Pellegrino, S.; Pelaia, C.; Stellato, C.; Pelaia, G.; Vitale, C. Phenotyping severe asthma: A rationale for biologic therapy. Expert Rev. Precis. Med. Drug Dev. 2020, 5, 265–274. [Google Scholar] [CrossRef]
- Kuruvilla, M.E.; Lee, F.E.-H.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef] [PubMed]
- McDowell, P.J.; Heaney, L.G. Different endotypes and phenotypes drive the heterogeneity in severe asthma. Allergy 2020, 75, 302–310. [Google Scholar] [CrossRef]
- Chung, K.F.; Wenzel, S.E.; Brozek, J.L.; Bush, A.; Castro, M.; Sterk, P.J.; Adcock, I.M.; Bateman, E.D.; Bel, E.H.; Bleecker, E.R.; et al. International ERS/ATS guidelines on definition, evaluation and treatment of severe asthma. Eur. Respir. J. 2014, 43, 343–373. [Google Scholar] [CrossRef]
- Israel, E.; Reddel, H.K. Severe and Difficult-to-Treat Asthma in Adults. N. Engl. J. Med. 2017, 377, 965–976. [Google Scholar] [CrossRef]
- Moore, W.C.; Bleecker, E.R.; Curran-Everett, D.; Erzurum, S.C.; Ameredes, B.T.; Bacharier, L.; Calhoun, W.J.; Castro, M.; Chung, K.F.; Clark, M.P.; et al. Characterization of the severe asthma phenotype by the National Heart, Lung, and Blood Institute’s Severe Asthma Research Program. J. Allergy Clin. Immunol. 2007, 119, 405–413. [Google Scholar] [CrossRef]
- Shaw, D.E.; Sousa, A.R.; Fowler, S.J.; Fleming, L.J.; Roberts, G.; Corfield, J.; Pandis, I.; Bansal, A.T.; Bel, E.H.; Auffray, C.; et al. Clinical and inflammatory characteristics of the European U-BIOPRED adult severe asthma cohort. Eur. Respir. J. 2015, 46, 1308–1321. [Google Scholar] [CrossRef]
- Hoda, U.; Pavlidis, S.; Bansal, A.T.; Takahashi, K.; Hu, S.; Ng Kee Kwong, F.; Rossios, C.; Sun, K.; Bhavsar, P.; Loza, M.; et al. Clinical and transcriptomic features of persistent exacerbation-prone severe asthma in U-BIOPRED cohort. Clin. Transl. Med. 2022, 12, e816. [Google Scholar] [CrossRef]
- de Groot, J.C.; Storm, H.; Amelink, M.; de Nijs, S.B.; Eichhorn, E.; Reitsma, B.H.; Bel, E.H.D.; ten Brinke, A. Clinical profile of patients with adult-onset eosinophilic asthma. ERJ Open Res. 2016, 2, 00100–02015. [Google Scholar] [CrossRef] [PubMed]
- Buhl, R.; Humbert, M.; Bjermer, L.; Chanez, P.; Heaney, L.G.; Pavord, I.; Quirce, S.; Virchow, J.C.; Holgate, S. Severe eosinophilic asthma: A roadmap to consensus. Eur. Respir. J. 2017, 49, 1700634. [Google Scholar] [CrossRef] [PubMed]
- Kurukulaaratchy, R.J.; Mistry, H. New Real-World Insights Into Severe Asthma: All About the Eosinophil? Chest 2021, 160, 789–790. [Google Scholar] [CrossRef] [PubMed]
- Heaney, L.G.; Perez de Llano, L.; Al-Ahmad, M.; Backer, V.; Busby, J.; Canonica, G.W.; Christoff, G.C.; Cosio, B.G.; FitzGerald, J.M.; Heffler, E.; et al. Eosinophilic and Noneosinophilic Asthma. Chest 2021, 160, 814–830. [Google Scholar] [CrossRef]
- Pelaia, C.; Pelaia, G.; Crimi, C.; Maglio, A.; Stanziola, A.A.; Calabrese, C.; Terracciano, R.; Longhini, F.; Vatrella, A. Novel Biological Therapies for Severe Asthma Endotypes. Biomedicines 2022, 10, 1064. [Google Scholar] [CrossRef]
- Rodrigo-Muñoz, J.M.; Gil-Martínez, M.; Sastre, B.; del Pozo, V. Emerging Evidence for Pleiotropism of Eosinophils. Int. J. Mol. Sci. 2021, 22, 7075. [Google Scholar] [CrossRef]
- Sastre, B.; Rodrigo-Muñoz, J.M.; Garcia-Sanchez, D.A.; Cañas, J.A.; Del Pozo, V. Eosinophils: Old Players in a New Game. J. Investig. Allergol. Clin. Immunol. 2018, 28, 289–304. [Google Scholar] [CrossRef]
- McBrien, C.N.; Menzies-Gow, A. The Biology of Eosinophils and Their Role in Asthma. Front. Med. 2017, 4, 93. [Google Scholar] [CrossRef]
- Gleich, G.J. Mechanisms of eosinophil-associated inflammation. J. Allergy Clin. Immunol. 2000, 105, 651–663. [Google Scholar] [CrossRef]
- Michail, S.; Mezoff, E.; Abernathy, F. Role of Selectins in the Intestinal Epithelial Migration of Eosinophils. Pediatr. Res. 2005, 58, 644–647. [Google Scholar] [CrossRef] [Green Version]
- Ying, S.; Meng, Q.; Zeibecoglou, K.; Robinson, D.S.; Macfarlane, A.; Humbert, M.; Kay, A.B. Eosinophil chemotactic chemokines (eotaxin, eotaxin-2, RANTES, monocyte chemoattractant protein-3 (MCP-3), and MCP-4), and C-C chemokine receptor 3 expression in bronchial biopsies from atopic and nonatopic (Intrinsic) asthmatics. J. Immunol. 1999, 163, 6321–6329. [Google Scholar] [PubMed]
- Stone, K.D.; Prussin, C.; Metcalfe, D.D. IgE, Mast Cells, Basophils, and Eosinophils. J. Allergy Clin. Immunol. 2010, 125, S73–S80. [Google Scholar] [CrossRef]
- Muraki, M.; Gleich, G.J.; Kita, H. Antigen-Specific IgG and IgA, but Not IgE, Activate the Effector Functions of Eosinophils in the Presence of Antigen. Int. Arch. Allergy Immunol. 2011, 154, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Sedgwick, J.B.; Calhoun, W.J.; Vrtis, R.F.; Bates, M.E.; McAllister, P.K.; Busse, W.W. Comparison of airway and blood eosinophil function after in vivo antigen challenge. J. Immunol. 1992, 149, 3710–3718. [Google Scholar] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Dembic, Z. The Function of Toll-like Receptors; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Kerr, S.C.; Gonzalez, J.R.; Schanin, J.; Peters, M.C.; Lambrecht, B.N.; Brock, E.C.; Charbit, A.; Ansel, K.M.; Youngblood, B.A.; Fahy, J.V. An anti-siglec-8 antibody depletes sputum eosinophils from asthmatic subjects and inhibits lung mast cells. Clin. Exp. Allergy 2020, 50, 904–914. [Google Scholar] [CrossRef]
- Carr, T.F.; Berdnikovs, S.; Simon, H.-U.; Bochner, B.S.; Rosenwasser, L.J. Eosinophilic bioactivities in severe asthma. World Allergy Organ. J. 2016, 9, 21. [Google Scholar] [CrossRef]
- Kiwamoto, T.; Kawasaki, N.; Paulson, J.C.; Bochner, B.S. Siglec-8 as a drugable target to treat eosinophil and mast cell-associated conditions. Pharmacol. Ther. 2012, 135, 327–336. [Google Scholar] [CrossRef]
- Mori, Y.; Iwasaki, H.; Kohno, K.; Yoshimoto, G.; Kikushige, Y.; Okeda, A.; Uike, N.; Niiro, H.; Takenaka, K.; Nagafuji, K.; et al. Identification of the human eosinophil lineage-committed progenitor: Revision of phenotypic definition of the human common myeloid progenitor. J. Exp. Med. 2009, 206, 183–193. [Google Scholar] [CrossRef]
- Klion, A. Recent advances in understanding eosinophil biology. F1000Research 2017, 6, 1084. [Google Scholar] [CrossRef] [Green Version]
- Dorman, S.C.; Efthimiadis, A.; Babirad, I.; Watson, R.M.; Denburg, J.A.; Hargreave, F.E.; O’Byrne, P.M.; Sehmi, R. Sputum CD34+IL-5Ralpha+ cells increase after allergen: Evidence for in situ eosinophilopoiesis. Am. J. Respir. Crit. Care Med. 2004, 169, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Rådinger, M.; Bossios, A.; Sjöstrand, M.; Lu, Y.; Malmhäll, C.; Dahlborn, A.-K.; Lee, J.J.; Lötvall, J. Local proliferation and mobilization of CCR3(+) CD34(+) eosinophil-lineage-committed cells in the lung. Immunology 2011, 132, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Sehmi, R.; Howie, K.; Sutherland, D.R.; Schragge, W.; O’Byrne, P.M.; Denburg, J.A. Increased levels of CD34+ hemopoietic progenitor cells in atopic subjects. Am. J. Respir. Cell Mol. Biol. 1996, 15, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Sehmi, R.; Smith, S.G.; Kjarsgaard, M.; Radford, K.; Boulet, L.-P.; Lemiere, C.; Prazma, C.M.; Ortega, H.; Martin, J.G.; Nair, P. Role of local eosinophilopoietic processes in the development of airway eosinophilia in prednisone-dependent severe asthma. Clin. Exp. Allergy 2016, 46, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Stankiewicz, M.J.; Liu, Y.; Xi, Q.; Schmitz, J.E.; Lekstrom-Himes, J.A.; Ackerman, S.J. Novel Combinatorial Interactions of GATA-1, PU.1, and C/EBPε Isoforms Regulate Transcription of the Gene Encoding Eosinophil Granule Major Basic Protein. J. Biol. Chem. 2002, 277, 43481–43494. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Cantor, A.B.; Yang, H.; Browne, C.; Wells, R.A.; Fujiwara, Y.; Orkin, S.H. Targeted Deletion of a High-Affinity GATA-binding Site in the GATA-1 Promoter Leads to Selective Loss of the Eosinophil Lineage In Vivo. J. Exp. Med. 2002, 195, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Dougan, M.; Dranoff, G.; Dougan, S.K. GM-CSF, IL-3, and IL-5 Family of Cytokines: Regulators of Inflammation. Immunity 2019, 50, 796–811. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, M.E.; Munitz, A.; Ackerman, S.J.; Drake, M.G.; Jackson, D.J.; Wardlaw, A.J.; Dougan, S.K.; Berdnikovs, S.; Schleich, F.; Matucci, A.; et al. Eosinophils in Health and Disease: A State-of-the-Art Review. Mayo Clin. Proc. 2021, 96, 2694–2707. [Google Scholar] [CrossRef]
- Eng, S.S.; DeFelice, M.L. The Role and Immunobiology of Eosinophils in the Respiratory System: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2016, 50, 140–158. [Google Scholar] [CrossRef]
- Bochner, B.S. The eosinophil. Ann. Allergy. Asthma. Immunol. 2018, 121, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, G.A.; Yacoub, M.-R.; Ripa, M.; Mannina, D.; Cariddi, A.; Saporiti, N.; Ciceri, F.; Castagna, A.; Colombo, G.; Dagna, L. Eosinophils from Physiology to Disease: A Comprehensive Review. BioMed Res. Int. 2018, 2018, 9095275. [Google Scholar] [CrossRef] [PubMed]
- Katsoulis, K.; Kipourou, M.; Loukides, S. Reduction/elimination of blood eosinophils in severe asthma: Should there be a safety consideration? Expert Opin. Biol. Ther. 2021, 22, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Phipps, S.; Foster, P.S. Eosinophil trafficking in allergy and asthma. J. Allergy Clin. Immunol. 2007, 119, 1303–1310; quiz 1311–1312. [Google Scholar] [CrossRef] [PubMed]
- Woltmann, G.; McNulty, C.A.; Dewson, G.; Symon, F.A.; Wardlaw, A.J. Interleukin-13 induces PSGL-1/P-selectin-dependent adhesion of eosinophils, but not neutrophils, to human umbilical vein endothelial cells under flow. Blood 2000, 95, 3146–3152. [Google Scholar] [CrossRef]
- Janson, C.; Bjermer, L.; Lehtimäki, L.; Kankaanranta, H.; Karjalainen, J.; Altraja, A.; Yasinska, V.; Aarli, B.; Rådinger, M.; Hellgren, J.; et al. Eosinophilic airway diseases: Basic science, clinical manifestations and future challenges. Eur. Clin. Respir. J. 2022, 9, 2040707. [Google Scholar] [CrossRef]
- Mjösberg, J.; Spits, H. Human innate lymphoid cells. J. Allergy Clin. Immunol. 2016, 138, 1265–1276. [Google Scholar] [CrossRef] [PubMed]
- Boberg, E.; Johansson, K.; Malmhäll, C.; Calvén, J.; Weidner, J.; Rådinger, M. Interplay Between the IL-33/ST2 Axis and Bone Marrow ILC2s in Protease Allergen-Induced IL-5-Dependent Eosinophilia. Front. Immunol. 2020, 11, 1058. [Google Scholar] [CrossRef] [PubMed]
- Johansson, K.; Malmhäll, C.; Ramos-Ramírez, P.; Rådinger, M. Bone marrow type 2 innate lymphoid cells: A local source of interleukin-5 in interleukin-33-driven eosinophilia. Immunology 2018, 153, 268–278. [Google Scholar] [CrossRef]
- Wardlaw, A.J. Molecular basis for selective eosinophil trafficking in asthma: A multistep paradigm. J. Allergy Clin. Immunol. 1999, 104, 917–926. [Google Scholar] [CrossRef]
- Venge, J.; Lampinen, M.; Håkansson, L.; Rak, S.; Venge, P. Identification of IL-5 and RANTES as the major eosinophil chemoattractants in the asthmatic lung. J. Allergy Clin. Immunol. 1996, 97, 1110–1115. [Google Scholar] [CrossRef]
- Sjöberg, L.C.; Nilsson, A.Z.; Lei, Y.; Gregory, J.A.; Adner, M.; Nilsson, G.P. Interleukin 33 exacerbates antigen driven airway hyperresponsiveness, inflammation and remodeling in a mouse model of asthma. Sci. Rep. 2017, 7, 4219. [Google Scholar] [CrossRef] [PubMed]
- Watson, B.; Gauvreau, G.M. Thymic stromal lymphopoietin: A central regulator of allergic asthma. Expert Opin. Ther. Targets 2014, 18, 771–785. [Google Scholar] [CrossRef]
- Yao, X.; Sun, Y.; Wang, W.; Sun, Y. Interleukin (IL)-25: Pleiotropic roles in asthma. Respirology 2016, 21, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Smith, S.G.; Salter, B.; El-Gammal, A.; Oliveria, J.P.; Obminski, C.; Watson, R.; O’Byrne, P.M.; Gauvreau, G.M.; Sehmi, R. Allergen-induced Increases in Sputum Levels of Group 2 Innate Lymphoid Cells in Subjects with Asthma. Am. J. Respir. Crit. Care Med. 2017, 196, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Johnston, L.K.; Hsu, C.-L.; Krier-Burris, R.A.; Chhiba, K.D.; Chien, K.B.; McKenzie, A.; Berdnikovs, S.; Bryce, P.J. IL-33 Precedes IL-5 in Regulating Eosinophil Commitment and Is Required for Eosinophil Homeostasis. J. Immunol. 2016, 197, 3445–3453. [Google Scholar] [CrossRef]
- Palframan, R.T.; Collins, P.D.; Williams, T.J.; Rankin, S.M. Eotaxin Induces a Rapid Release of Eosinophils and Their Progenitors From the Bone Marrow. Blood 1998, 91, 2240–2248. [Google Scholar] [CrossRef]
- Esnault, S.; Kelly, E.A. Essential Mechanisms of Differential Activation of Eosinophils by IL-3 Compared to GM-CSF and IL-5. Crit. Rev. Immunol. 2016, 36, 429–444. [Google Scholar] [CrossRef]
- Schwartz, C.; Willebrand, R.; Huber, S.; Rupec, R.A.; Wu, D.; Locksley, R.; Voehringer, D. Eosinophil-specific deletion of IκBα in mice reveals a critical role of NF-κB-induced Bcl-xL for inhibition of apoptosis. Blood 2015, 125, 3896–3904. [Google Scholar] [CrossRef]
- Fettrelet, T.; Gigon, L.; Karaulov, A.; Yousefi, S.; Simon, H.-U. The Enigma of Eosinophil Degranulation. Int. J. Mol. Sci. 2021, 22, 7091. [Google Scholar] [CrossRef]
- Ueki, S.; Tokunaga, T.; Melo, R.C.N.; Saito, H.; Honda, K.; Fukuchi, M.; Konno, Y.; Takeda, M.; Yamamoto, Y.; Hirokawa, M.; et al. Charcot-Leyden crystal formation is closely associated with eosinophil extracellular trap cell death. Blood 2018, 132, 2183–2187. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, S.; Gold, J.A.; Andina, N.; Lee, J.J.; Kelly, A.M.; Kozlowski, E.; Schmid, I.; Straumann, A.; Reichenbach, J.; Gleich, G.J.; et al. Catapult-like release of mitochondrial DNA by eosinophils contributes to antibacterial defense. Nat. Med. 2008, 14, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Mazzeo, C.; Cañas, J.A.; Zafra, M.P.; Rojas Marco, A.; Fernández-Nieto, M.; Sanz, V.; Mittelbrunn, M.; Izquierdo, M.; Baixaulli, F.; Sastre, J.; et al. Exosome secretion by eosinophils: A possible role in asthma pathogenesis. J. Allergy Clin. Immunol. 2015, 135, 1603–1613. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Raposo, G.; Stahl, P.D. Extracellular vesicles: A new communication paradigm? Nat. Rev. Mol. Cell Biol. 2019, 20, 509–510. [Google Scholar] [CrossRef]
- Choi, Y.; Sim, S.; Park, H.-S. Distinct functions of eosinophils in severe asthma with type 2 phenotype: Clinical implications. Korean J. Intern. Med. 2020, 35, 823–833. [Google Scholar] [CrossRef]
- Sastre, B.; Cañas, J.A.; Rodrigo-Muñoz, J.M.; Del Pozo, V. Novel Modulators of Asthma and Allergy: Exosomes and MicroRNAs. Front. Immunol. 2017, 8, 826. [Google Scholar] [CrossRef]
- Van Hulst, G.; Batugedara, H.M.; Jorssen, J.; Louis, R.; Bureau, F.; Desmet, C.J. Eosinophil diversity in asthma. Biochem. Pharmacol. 2020, 179, 113963. [Google Scholar] [CrossRef]
- Mesnil, C.; Raulier, S.; Paulissen, G.; Xiao, X.; Birrell, M.A.; Pirottin, D.; Janss, T.; Starkl, P.; Ramery, E.; Henket, M.; et al. Lung-resident eosinophils represent a distinct regulatory eosinophil subset. J. Clin. Investig. 2016, 126, 3279–3295. [Google Scholar] [CrossRef]
- Davoine, F.; Lacy, P. Eosinophil cytokines, chemokines, and growth factors: Emerging roles in immunity. Front. Immunol. 2014, 5, 570. [Google Scholar] [CrossRef] [Green Version]
- Yukawa, T.; Kroegel, C.; Evans, P.; Fukuda, T.; Chung, K.F.; Barnes, P.J. Density heterogeneity of eosinophil leucocytes: Induction of hypodense eosinophils by platelet-activating factor. Immunology 1989, 68, 140–143. [Google Scholar] [PubMed]
- Frick, W.E.; Sedgwick, J.B.; Busse, W.W. The appearance of hypodense eosinophils in antigen-dependent late phase asthma. Am. Rev. Respir. Dis. 1989, 139, 1401–1406. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Dunnette, S.L.; Reed, C.E.; Ackerman, S.J.; Peters, M.S.; Gleich, G.J. Increased numbers of hypodense eosinophils in the blood of patients with bronchial asthma. Am. Rev. Respir. Dis. 1985, 132, 981–985. [Google Scholar] [CrossRef]
- Fukuda, T.; Gleich, G.J. Heterogeneity of human eosinophils. J. Allergy Clin. Immunol. 1989, 83, 369–373. [Google Scholar] [CrossRef]
- Campbell, H.D.; Tucker, W.Q.; Hort, Y.; Martinson, M.E.; Mayo, G.; Clutterbuck, E.J.; Sanderson, C.J.; Young, I.G. Molecular cloning, nucleotide sequence, and expression of the gene encoding human eosinophil differentiation factor (interleukin 5). Proc. Natl. Acad. Sci. USA 1987, 84, 6629–6633. [Google Scholar] [CrossRef]
- Garcia-Romo, G.S.; Caielli, S.; Vega, B.; Connolly, J.; Allantaz, F.; Xu, Z.; Punaro, M.; Baisch, J.; Guiducci, C.; Coffman, R.L.; et al. Netting neutrophils are major inducers of type I IFN production in pediatric systemic lupus erythematosus. Sci. Transl. Med. 2011, 3, 73ra20. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, S.M.; Thieblemont, N.; Witko-Sarsat, V. Expanding Neutrophil Horizons: New Concepts in Inflammation. J. Innate Immun. 2018, 10, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Laviolette, M.; Gossage, D.L.; Gauvreau, G.; Leigh, R.; Olivenstein, R.; Katial, R.; Busse, W.W.; Wenzel, S.; Wu, Y.; Datta, V.; et al. Effects of benralizumab on airway eosinophils in asthmatic patients with sputum eosinophilia. J. Allergy Clin. Immunol. 2013, 132, 1086–1096.e5. [Google Scholar] [CrossRef]
- Kanda, A.; Yun, Y.; Bui, D.V.; Nguyen, L.M.; Kobayashi, Y.; Suzuki, K.; Mitani, A.; Sawada, S.; Hamada, S.; Asako, M.; et al. The multiple functions and subpopulations of eosinophils in tissues under steady-state and pathological conditions. Allergol. Int. Off. J. Jpn; thusc. Allergol. 2021, 70, 9–18. [Google Scholar] [CrossRef]
- Lombardi, C.; Passalacqua, G. Eosinophilia and diseases: Clinical revision of 1862 cases. Arch. Intern. Med. 2003, 163, 1371–1373. [Google Scholar] [CrossRef]
- Lombardi, C.; Berti, A.; Cottini, M. The emerging roles of eosinophils: Implications for the targeted treatment of eosinophilic-associated inflammatory conditions. Curr. Res. Immunol. 2022, 3, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The Cytokines of Asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef]
- Pelaia, C.; Paoletti, G.; Puggioni, F.; Racca, F.; Pelaia, G.; Canonica, G.W.; Heffler, E. Interleukin-5 in the Pathophysiology of Severe Asthma. Front. Physiol. 2019, 10, 1514. [Google Scholar] [CrossRef]
- Kouro, T.; Takatsu, K. IL-5- and eosinophil-mediated inflammation: From discovery to therapy. Int. Immunol. 2009, 21, 1303–1309. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, A.; Mukherjee, M.; Nair, P. Airway Eosinophilopoietic and Autoimmune Mechanisms of Eosinophilia in Severe Asthma. Immunol. Allergy Clin. N. Am. 2018, 38, 639–654. [Google Scholar] [CrossRef]
- Greenfeder, S.; Umland, S.P.; Cuss, F.M.; Chapman, R.W.; Egan, R.W. Th2 cytokines and asthma. The role of interleukin-5 in allergic eosinophilic disease. Respir. Res. 2001, 2, 71–79. [Google Scholar] [CrossRef]
- Xu, J.; Jiang, F.; Nayeri, F.; Zetterström, O. Apoptotic eosinophils in sputum from asthmatic patients correlate negatively with levels of IL-5 and eotaxin. Respir. Med. 2007, 101, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Fulkerson, P.C.; Rothenberg, M.E. Targeting eosinophils in allergy, inflammation and beyond. Nat. Rev. Drug Discov. 2013, 12, 117–129. [Google Scholar] [CrossRef]
- Johansson, M.W. Eosinophil Activation Status in Separate Compartments and Association with Asthma. Front. Med. 2017, 4, 75. [Google Scholar] [CrossRef]
- Mould, A.W.; Ramsay, A.J.; Matthaei, K.I.; Young, I.G.; Rothenberg, M.E.; Foster, P.S. The effect of IL-5 and eotaxin expression in the lung on eosinophil trafficking and degranulation and the induction of bronchial hyperreactivity. J. Immunol. 2000, 164, 2142–2150. [Google Scholar] [CrossRef] [Green Version]
- Vatrella, A.; Ponticiello, A.; Parrella, R.; Romano, I.; Zofra, S.; Leva, A.D.; Bariffi, F. Serum eosinophil cationic protein (ECP) as a marker of disease activity and treatment efficacy in seasonal asthmi. Allergy 1996, 51, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Pascual, R.M.; Peters, S.P. Airway remodeling contributes to the progressive loss of lung function in asthma: An overview. J. Allergy Clin. Immunol. 2005, 116, 477–486; quiz 487. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, S.E.; Schwartz, L.B.; Langmack, E.L.; Halliday, J.L.; Trudeau, J.B.; Gibbs, R.L.; Chu, H.W. Evidence that severe asthma can be divided pathologically into two inflammatory subtypes with distinct physiologic and clinical characteristics. Am. J. Respir. Crit. Care Med. 1999, 160, 1001–1008. [Google Scholar] [CrossRef]
- Redington, A.E.; Madden, J.; Frew, A.J.; Djukanovic, R.; Roche, W.R.; Holgate, S.T.; Howarth, P.H. Transforming growth factor-beta 1 in asthma. Measurement in bronchoalveolar lavage fluid. Am. J. Respir. Crit. Care Med. 1997, 156, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Ohno, I.; Nitta, Y.; Yamauchi, K.; Hoshi, H.; Honma, M.; Woolley, K.; O’Byrne, P.; Tamura, G.; Jordana, M.; Shirato, K. Transforming growth factor beta 1 (TGF beta 1) gene expression by eosinophils in asthmatic airway inflammation. Am. J. Respir. Cell Mol. Biol. 1996, 15, 404–409. [Google Scholar] [CrossRef]
- Wong, D.T.; Elovic, A.; Matossian, K.; Nagura, N.; McBride, J.; Chou, M.Y.; Gordon, J.R.; Rand, T.H.; Galli, S.J.; Weller, P.F. Eosinophils from patients with blood eosinophilia express transforming growth factor beta 1. Blood 1991, 78, 2702–2707. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, H.H.; Robinson, D.S. The role of eosinophils in airway tissue remodelling in asthma. Curr. Opin. Immunol. 2007, 19, 681–686. [Google Scholar] [CrossRef]
- Van Hulst, G.; Bureau, F.; Desmet, C.J. Eosinophils as Drivers of Severe Eosinophilic Asthma: Endotypes or Plasticity? Int. J. Mol. Sci. 2021, 22, 10150. [Google Scholar] [CrossRef]
- Amison, R.T.; Page, C.P. Novel pharmacological therapies for the treatment of bronchial asthma. Minerva Med. 2022, 113, 31–50. [Google Scholar] [CrossRef]
- Cusack, R.P.; Whetstone, C.E.; Xie, Y.; Ranjbar, M.; Gauvreau, G.M. Regulation of Eosinophilia in Asthma-New Therapeutic Approaches for Asthma Treatment. Cells 2021, 10, 817. [Google Scholar] [CrossRef]
- Pelaia, G.; Vatrella, A.; Cuda, G.; Maselli, R.; Marsico, S.A. Molecular mechanisms of corticosteroid actions in chronic inflammatory airway diseases. Life Sci. 2003, 72, 1549–1561. [Google Scholar] [CrossRef]
- Barnes, P.J. New therapies for asthma. Trends Mol. Med. 2006, 12, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Rhen, T.; Cidlowski, J.A. Antiinflammatory action of glucocorticoids—New mechanisms for old drugs. N. Engl. J. Med. 2005, 353, 1711–1723. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Glucocorticosteroids. Handb. Exp. Pharmacol. 2017, 237, 93–115. [Google Scholar] [CrossRef]
- Hebbar, P.B.; Archer, T.K. Chromatin remodeling by nuclear receptors. Chromosoma 2003, 111, 495–504. [Google Scholar] [CrossRef]
- Barnes, P.J. Corticosteroid effects on cell signalling. Eur. Respir. J. 2006, 27, 413–426. [Google Scholar] [CrossRef]
- Pelaia, C.; Vatrella, A.; Gallelli, L.; Lombardo, N.; Sciacqua, A.; Savino, R.; Pelaia, G. Role of p38 Mitogen-Activated Protein Kinase in Asthma and COPD: Pathogenic Aspects and Potential Targeted Therapies. Drug Des. Dev. Ther. 2021, 15, 1275–1284. [Google Scholar] [CrossRef]
- Barnes, P.J.; Adcock, I.M. How Do Corticosteroids Work in Asthma? Ann. Intern. Med. 2003, 139, 359–370. [Google Scholar] [CrossRef]
- Barnes, P.J. Biochemical Basis of Asthma Therapy. J. Biol. Chem. 2011, 286, 32899–32905. [Google Scholar] [CrossRef]
- Smoak, K.; Cidlowski, J.A. Glucocorticoids Regulate Tristetraprolin Synthesis and Posttranscriptionally Regulate Tumor Necrosis Factor Alpha Inflammatory Signaling. Mol. Cell. Biol. 2006, 26, 9126–9135. [Google Scholar] [CrossRef] [Green Version]
- Giembycz, M.A.; Lindsay, M.A. Pharmacology of the eosinophil. Pharmacol. Rev. 1999, 51, 213–340. [Google Scholar] [PubMed]
- Rogliani, P.; Ritondo, B.L.; Puxeddu, E.; Pane, G.; Cazzola, M.; Calzetta, L. Experimental Glucocorticoid Receptor Agonists for the Treatment of Asthma: A Systematic Review. J. Exp. Pharmacol. 2020, 12, 233–254. [Google Scholar] [CrossRef] [PubMed]
- Ingawale, D.; Mandlik, S. New insights into the novel anti-inflammatory mode of action of glucocorticoids. Immunopharmacol. Immunotoxicol. 2020, 42, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Shalit, M.; Sekhsaria, S.; Malech, H.L. Modulation of growth and differentiation of eosinophils from human peripheral blood CD34+ cells by IL5 and other growth factors. Cell. Immunol. 1995, 160, 50–57. [Google Scholar] [CrossRef]
- Pavord, I.D. Oral corticosteroid-dependent asthma: Current knowledge and future needs. Curr. Opin. Pulm. Med. 2019, 25, 51–58. [Google Scholar] [CrossRef]
- Uller, L.; Lloyd, C.M.; Rydell-Törmänen, K.; Persson, C.G.A.; Erjefält, J.S. Effects of steroid treatment on lung CC chemokines, apoptosis and transepithelial cell clearance during development and resolution of allergic airway inflammation. Clin. Exp. Allergy 2006, 36, 111–121. [Google Scholar] [CrossRef]
- Vatrella, A.; Perna, F.; Pelaia, G.; Parrella, R.; Maselli, R.; Marsico, S.A.; Calabrese, C. T cell activation state in the induced sputum of asthmatics treated with budesonide. Int. J. Immunopathol. Pharmacol. 2010, 23, 745–753. [Google Scholar] [CrossRef]
- Atsuta, J.; Plitt, J.; Bochner, B.S.; Schleimer, R.P. Inhibition of VCAM-1 expression in human bronchial epithelial cells by glucocorticoids. Am. J. Respir. Cell Mol. Biol. 1999, 20, 643–650. [Google Scholar] [CrossRef]
- Adelroth, E.; Rosenhall, L.; Johansson, S.A.; Linden, M.; Venge, P. Inflammatory cells and eosinophilic activity in asthmatics investigated by bronchoalveolar lavage. The effects of antiasthmatic treatment with budesonide or terbutaline. Am. Rev. Respir. Dis. 1990, 142, 91–99. [Google Scholar] [CrossRef]
- Vitale, C.; Maglio, A.; Pelaia, C.; Vatrella, A. Long-term treatment in pediatric asthma: An update on chemical pharmacotherapy. Expert Opin. Pharmacother. 2017, 18, 667–676. [Google Scholar] [CrossRef]
- Mukherjee, M.; Svenningsen, S.; Nair, P. Glucocortiosteroid subsensitivity and asthma severity. Curr. Opin. Pulm. Med. 2017, 23, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Dunican, E.M.; Fahy, J.V. Asthma and corticosteroids: Time for a more precise approach to treatment. Eur. Respir. J. 2017, 49, 1701167. [Google Scholar] [CrossRef]
- Barnes, P.J. Corticosteroid resistance in patients with asthma and chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2013, 131, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Pelaia, C.; Vatrella, A.; Crimi, C.; Gallelli, L.; Terracciano, R.; Pelaia, G. Clinical relevance of understanding mitogen-activated protein kinases involved in asthma. Expert Rev. Respir. Med. 2020, 14, 501–510. [Google Scholar] [CrossRef] [PubMed]
- 2022 GINA Main Report. Available online: https://ginasthma.org/gina-reports/ (accessed on 9 August 2022).
- Pelaia, C.; Vatrella, A.; Busceti, M.T.; Gallelli, L.; Terracciano, R.; Savino, R.; Pelaia, G. Severe eosinophilic asthma: From the pathogenic role of interleukin-5 to the therapeutic action of mepolizumab. Drug Des. Dev. Ther. 2017, 11, 3137–3144. [Google Scholar] [CrossRef] [PubMed]
- Pavord, I.D.; Bel, E.H.; Bourdin, A.; Chan, R.; Han, J.K.; Keene, O.N.; Liu, M.C.; Martin, N.; Papi, A.; Roufosse, F.; et al. From DREAM to REALITI-A and beyond: Mepolizumab for the treatment of eosinophil-driven diseases. Allergy 2022, 77, 778–797. [Google Scholar] [CrossRef]
- Haldar, P.; Brightling, C.E.; Hargadon, B. Mepolizumab and exacerbations of refractory eosinophilic asthma. N. Engl. J. Med. 2009, 360, 973–984. [Google Scholar] [CrossRef]
- Nair, P.; Pizzichini, M.M.M.; Kjarsgaard, M.; Inman, M.D.; Efthimiadis, A.; Pizzichini, E.; Hargreave, F.E.; O’Byrne, P.M. Mepolizumab for prednisone-dependent asthma with sputum eosinophilia. N. Engl. J. Med. 2009, 360, 985–993. [Google Scholar] [CrossRef]
- Pavord, I.D.; Korn, S.; Howarth, P.; Bleecker, E.R.; Buhl, R.; Keene, O.N.; Ortega, H.; Chanez, P. Mepolizumab for severe eosinophilic asthma (DREAM): A multicentre, double-blind, placebo-controlled trial. Lancet 2012, 380, 651–659. [Google Scholar] [CrossRef]
- Ortega, H.G.; Liu, M.C.; Pavord, I.D.; Brusselle, G.G.; FitzGerald, J.M.; Chetta, A.; Humbert, M.; Katz, L.E.; Keene, O.N.; Yancey, S.W.; et al. Mepolizumab Treatment in Patients with Severe Eosinophilic Asthma. N. Engl. J. Med. 2014, 371, 1198–1207. [Google Scholar] [CrossRef] [Green Version]
- Bel, E.H.; Wenzel, S.E.; Thompson, P.J.; Prazma, C.M.; Keene, O.N.; Yancey, S.W.; Ortega, H.G.; Pavord, I.D. SIRIUS Investigators Oral glucocorticoid-sparing effect of mepolizumab in eosinophilic asthma. N. Engl. J. Med. 2014, 371, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Chupp, G.L.; Bradford, E.S.; Albers, F.C.; Bratton, D.J.; Wang-Jairaj, J.; Nelsen, L.M.; Trevor, J.L.; Magnan, A.; Ten Brinke, A. Efficacy of mepolizumab add-on therapy on health-related quality of life and markers of asthma control in severe eosinophilic asthma (MUSCA): A randomised, double-blind, placebo-controlled, parallel-group, multicentre, phase 3b trial. Lancet Respir. Med. 2017, 5, 390–400. [Google Scholar] [CrossRef]
- Albers, F.C.; Bratton, D.J.; Gunsoy, N.B.; Cockle, S.M.; Alfonso-Cristancho, R.; Braunstahl, G.-J. Mepolizumab improves work productivity, activity limitation, symptoms, and rescue medication use in severe eosinophilic asthma. Clin. Respir. J. 2022, 16, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Lugogo, N.; Domingo, C.; Chanez, P.; Leigh, R.; Gilson, M.J.; Price, R.G.; Yancey, S.W.; Ortega, H.G. Long-term Efficacy and Safety of Mepolizumab in Patients With Severe Eosinophilic Asthma: A Multi-center, Open-label, Phase IIIb Study. Clin. Ther. 2016, 38, 2058–2070.e1. [Google Scholar] [CrossRef]
- Harrison, T.; Canonica, G.W.; Chupp, G.; Lee, J.; Schleich, F.; Welte, T.; Valero, A.; Gemzoe, K.; Maxwell, A.; Joksaite, S.; et al. Real-world mepolizumab in the prospective severe asthma REALITI-A study: Initial analysis. Eur. Respir. J. 2020, 56, 2000151. [Google Scholar] [CrossRef]
- Bagnasco, D.; Milanese, M.; Rolla, G.; Lombardi, C.; Bucca, C.; Heffler, E.; Canonica, G.W.; Passalacqua, G. The North-Western Italian experience with anti IL-5 therapy amd comparison with regulatory trials. World Allergy Organ. J. 2018, 11, 34. [Google Scholar] [CrossRef]
- Maglio, A.; Vitale, C.; Pellegrino, S.; Calabrese, C.; D’amato, M.; Molino, A.; Pelaia, C.; Triggiani, M.; Pelaia, G.; Stellato, C.; et al. Real-life effectiveness of mepolizumab on forced expiratory flow between 25% and 75% of forced vital capacity in patients with severe eosinophilic asthma. Biomedicines 2021, 9, 1550. [Google Scholar] [CrossRef]
- Harvey, E.S.; Langton, D.; Katelaris, C.; Stevens, S.; Farah, C.S.; Gillman, A.; Harrington, J.; Hew, M.; Kritikos, V.; Radhakrishna, N.; et al. Mepolizumab effectiveness and identification of super-responders in severe asthma. Eur. Respir. J. 2020, 55, 1902420. [Google Scholar] [CrossRef]
- Pelaia, C.; Crimi, C.; Pelaia, G.; Nolasco, S.; Campisi, R.; Heffler, E.; Valenti, G.; Crimi, N. Real-life evaluation of mepolizumab efficacy in patients with severe eosinophilic asthma, according to atopic trait and allergic phenotype. Clin. Exp. Allergy 2020, 50, 780–788. [Google Scholar] [CrossRef]
- Chapman, K.R.; Albers, F.C.; Chipps, B.; Muñoz, X.; Devouassoux, G.; Bergna, M.; Galkin, D.; Azmi, J.; Mouneimne, D.; Price, R.G.; et al. The clinical benefit of mepolizumab replacing omalizumab in uncontrolled severe eosinophilic asthma. Allergy 2019, 74, 1716–1726. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.C.; Chipps, B.; Munoz, X.; Devouassoux, G.; Bergna, M.; Smith, S.G.; Price, R.G.; Galkin, D.V.; Azmi, J.; Mouneimne, D.; et al. Benefit of switching to mepolizumab from omalizumab in severe eosinophilic asthma based on patient characteristics. Respir. Res. 2021, 22, 144. [Google Scholar] [CrossRef] [PubMed]
- Pelaia, G.; Vatrella, A.; Busceti, M.T.; Gallelli, L.; Preianò, M.; Lombardo, N.; Terracciano, R.; Maselli, R. Role of biologics in severe eosinophilic asthma—Focus on reslizumab. Ther. Clin. Risk Manag. 2016, 12, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Senna, G.; Loffredo, S.; Bagnasco, D.; Ferrando, M.; Canonica, G.W. Reslizumab and Eosinophilic Asthma: One Step Closer to Precision Medicine? Front. Immunol. 2017, 8, 242. [Google Scholar] [CrossRef]
- Kips, J.C.; O’Connor, B.J.; Langley, S.J.; Woodcock, A.; Kerstjens, H.A.M.; Postma, D.S.; Danzig, M.; Cuss, F.; Pauwels, R.A. Effect of SCH55700, a humanized anti-human interleukin-5 antibody, in severe persistent asthma: A pilot study. Am. J. Respir. Crit. Care Med. 2003, 167, 1655–1659. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.; Mathur, S.; Hargreave, F.; Boulet, L.-P.; Xie, F.; Young, J.; Wilkins, H.J.; Henkel, T.; Nair, P. Res-5-0010 Study Group Reslizumab for poorly controlled, eosinophilic asthma: A randomized, placebo-controlled study. Am. J. Respir. Crit. Care Med. 2011, 184, 1125–1132. [Google Scholar] [CrossRef]
- Castro, M.; Zangrilli, J.; Wechsler, M.E.; Bateman, E.D.; Brusselle, G.G.; Bardin, P.; Murphy, K.; Maspero, J.F.; O’Brien, C.; Korn, S. Reslizumab for inadequately controlled asthma with elevated blood eosinophil counts: Results from two multicentre, parallel, double-blind, randomised, placebo-controlled, phase 3 trials. Lancet Respir. Med. 2015, 3, 355–366. [Google Scholar] [CrossRef]
- Brusselle, G.; Germinaro, M.; Weiss, S.; Zangrilli, J. Reslizumab in patients with inadequately controlled late-onset asthma and elevated blood eosinophils. Pulm. Pharmacol. Ther. 2017, 43, 39–45. [Google Scholar] [CrossRef]
- Bjermer, L.; Lemiere, C.; Maspero, J.; Weiss, S.; Zangrilli, J.; Germinaro, M. Reslizumab for Inadequately Controlled Asthma With Elevated Blood Eosinophil Levels: A Randomized Phase 3 Study. Chest 2016, 150, 789–798. [Google Scholar] [CrossRef]
- Wechsler, M.E.; Peters, S.P.; Hill, T.D.; Ariely, R.; DePietro, M.R.; Driessen, M.T.; Terasawa, E.L.; Thomason, D.R.; Panettieri, R.A. Clinical Outcomes and Health-Care Resource Use Associated With Reslizumab Treatment in Adults With Severe Eosinophilic Asthma in Real-World Practice. Chest 2021, 159, 1734–1746. [Google Scholar] [CrossRef]
- Virchow, J.C.; Katial, R.; Brusselle, G.G.; Shalit, Y.; Garin, M.; McDonald, M.; Castro, M. Safety of Reslizumab in Uncontrolled Asthma with Eosinophilia: A Pooled Analysis from 6 Trials. J. Allergy Clin. Immunol. Pract. 2020, 8, 540–548.e1. [Google Scholar] [CrossRef] [Green Version]
- Pelaia, C.; Calabrese, C.; Vatrella, A.; Busceti, M.T.; Garofalo, E.; Lombardo, N.; Terracciano, R.; Pelaia, G. Benralizumab: From the basic mechanism of action to the potential use in the biological therapy of severe eosinophilic asthma. BioMed Res. Int. 2018, 2018, 4839230. [Google Scholar] [CrossRef] [PubMed]
- Bleecker, E.R.; FitzGerald, J.M.; Chanez, P.; Papi, A.; Weinstein, S.F.; Barker, P.; Sproule, S.; Gilmartin, G.; Aurivillius, M.; Werkström, V.; et al. Efficacy and safety of benralizumab for patients with severe asthma uncontrolled with high-dosage inhaled corticosteroids and long-acting β2-agonists (SIROCCO): A randomised, multicentre, placebo-controlled phase 3 trial. Lancet 2016, 388, 2115–2127. [Google Scholar] [CrossRef]
- FitzGerald, J.M.; Bleecker, E.R.; Nair, P.; Korn, S.; Ohta, K.; Lommatzsch, M.; Ferguson, G.T.; Busse, W.W.; Barker, P.; Sproule, S.; et al. Benralizumab, an anti-interleukin-5 receptor α monoclonal antibody, as add-on treatment for patients with severe, uncontrolled, eosinophilic asthma (CALIMA): A randomised, double-blind, placebo-controlled phase 3 trial. Lancet 2016, 388, 2128–2141. [Google Scholar] [CrossRef]
- Ferguson, G.T.; FitzGerald, J.M.; Bleecker, E.R.; Laviolette, M.; Bernstein, D.; LaForce, C.; Mansfield, L.; Barker, P.; Wu, Y.; Jison, M.; et al. Benralizumab for patients with mild to moderate, persistent asthma (BISE): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Respir. Med. 2017, 5, 568–576. [Google Scholar] [CrossRef]
- Harrison, T.W.; Chanez, P.; Menzella, F.; Canonica, G.W.; Louis, R.; Cosio, B.G.; Lugogo, N.L.; Mohan, A.; Burden, A.; McDermott, L.; et al. Onset of effect and impact on health-related quality of life, exacerbation rate, lung function, and nasal polyposis symptoms for patients with severe eosinophilic asthma treated with benralizumab (ANDHI): A randomised, controlled, phase 3b trial. Lancet Respir. Med. 2021, 9, 260–274. [Google Scholar] [CrossRef]
- Nair, P.; Wenzel, S.; Rabe, K.F.; Bourdin, A.; Lugogo, N.L.; Kuna, P.; Barker, P.; Sproule, S.; Ponnarambil, S.; Goldman, M. Oral Glucocorticoid–Sparing Effect of Benralizumab in Severe Asthma. N. Engl. J. Med. 2017, 376, 2448–2458. [Google Scholar] [CrossRef]
- Menzies-Gow, A.; Gurnell, M.; Heaney, L.G.; Corren, J.; Bel, E.H.; Maspero, J.; Harrison, T.; Jackson, D.J.; Price, D.; Lugogo, N.; et al. Oral corticosteroid elimination via a personalised reduction algorithm in adults with severe, eosinophilic asthma treated with benralizumab (PONENTE): A multicentre, open-label, single-arm study. Lancet Respir. Med. 2022, 10, 47–58. [Google Scholar] [CrossRef]
- Busse, W.W.; Bleecker, E.R.; FitzGerald, J.M.; Ferguson, G.T.; Barker, P.; Sproule, S.; Olsson, R.F.; Martin, U.J.; Goldman, M. BORA study investigators Long-term safety and efficacy of benralizumab in patients with severe, uncontrolled asthma: 1-year results from the BORA phase 3 extension trial. Lancet Respir. Med. 2019, 7, 46–59. [Google Scholar] [CrossRef]
- Chipps, B.E.; Newbold, P.; Hirsch, I.; Trudo, F.; Goldman, M. Benralizumab efficacy by atopy status and serum immunoglobulin E for patients with severe, uncontrolled asthma. Ann. Allergy Asthma Immunol. Off. Publ. Am. Coll. Allergy Asthma Immunol. 2018, 120, 504–511.e4. [Google Scholar] [CrossRef]
- Pelaia, C.; Crimi, C.; Benfante, A.; Caiaffa, M.F.; Calabrese, C.; Carpagnano, G.E.; Ciotta, D.; D’Amato, M.; Macchia, L.; Nolasco, S.; et al. Therapeutic Effects of Benralizumab Assessed in Patients with Severe Eosinophilic Asthma: Real-Life Evaluation Correlated with Allergic and Non-Allergic Phenotype Expression. J. Asthma Allergy 2021, 14, 163–173. [Google Scholar] [CrossRef]
- Menzella, F.; Bargagli, E.; Aliani, M.; Bracciale, P.; Brussino, L.; Caiaffa, M.F.; Caruso, C.; Centanni, S.; D’Amato, M.; Del Giacco, S.; et al. ChAracterization of ItaliaN severe uncontrolled Asthmatic patieNts Key features when receiving Benralizumab in a real-life setting: The observational rEtrospective ANANKE study. Respir. Res. 2022, 23, 36. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, J.E.; Hearn, A.P.; Dhariwal, J.; d’Ancona, G.; Douiri, A.; Roxas, C.; Fernandes, M.; Green, L.; Thomson, L.; Nanzer, A.M.; et al. Real-World Effectiveness of Benralizumab in Severe Eosinophilic Asthma. Chest 2021, 159, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Di Bona, D.; Crimi, C.; D’Uggento, A.M.; Benfante, A.; Caiaffa, M.F.; Calabrese, C.; Campisi, R.; Carpagnano, G.E.; Ciotta, D.; D’Amato, M.; et al. Effectiveness of benralizumab in severe eosinophilic asthma: Distinct sub-phenotypes of response identified by cluster analysis. Clin. Exp. Allergy 2022, 52, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Harb, H.; Chatila, T.A. Mechanisms of Dupilumab. Clin. Exp. Allergy 2020, 50, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.; Corren, J.; Pavord, I.D.; Maspero, J.; Wenzel, S.; Rabe, K.F.; Busse, W.W.; Ford, L.; Sher, L.; FitzGerald, J.M.; et al. Dupilumab Efficacy and Safety in Moderate-to-Severe Uncontrolled Asthma. N. Engl. J. Med. 2018, 378, 2486–2496. [Google Scholar] [CrossRef]
- Menzies-Gow, A.; Mansur, A.H.; Brightling, C.E. Clinical utility of fractional exhaled nitric oxide in severe asthma management. Eur. Respir. J. 2020, 55, 1901633. [Google Scholar] [CrossRef]
- Dupin, C.; Belhadi, D.; Guilleminault, L.; Gamez, A.-S.; Berger, P.; De Blay, F.; Bonniaud, P.; Leroyer, C.; Mahay, G.; Girodet, P.-O.; et al. Effectiveness and safety of dupilumab for the treatment of severe asthma in a real-life French multi-centre adult cohort. Clin. Exp. Allergy 2020, 50, 789–798. [Google Scholar] [CrossRef]
- Menzies-Gow, A.; Corren, J.; Bourdin, A.; Chupp, G.; Israel, E.; Wechsler, M.E.; Brightling, C.E.; Griffiths, J.M.; Hellqvist, Å.; Bowen, K.; et al. Tezepelumab in Adults and Adolescents with Severe, Uncontrolled Asthma. N. Engl. J. Med. 2021, 384, 1800–1809. [Google Scholar] [CrossRef]
- Corren, J.; Parnes, J.R.; Wang, L.; Mo, M.; Roseti, S.L.; Griffiths, J.M.; van der Merwe, R. Tezepelumab in Adults with Uncontrolled Asthma. N. Engl. J. Med. 2017, 377, 936–946. [Google Scholar] [CrossRef]
- Sverrild, A.; Hansen, S.; Hvidtfeldt, M.; Clausson, C.-M.; Cozzolino, O.; Cerps, S.; Uller, L.; Backer, V.; Erjefält, J.; Porsbjerg, C. The effect of tezepelumab on airway hyperresponsiveness to mannitol in asthma (UPSTREAM). Eur. Respir. J. 2022, 59, 2101296. [Google Scholar] [CrossRef]
- Tanaka, J.; Watanabe, N.; Kido, M.; Saga, K.; Akamatsu, T.; Nishio, A.; Chiba, T. Human TSLP and TLR3 ligands promote differentiation of Th17 cells with a central memory phenotype under Th2-polarizing conditions. Clin. Exp. Allergy 2009, 39, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Adatia, A.; Wahab, M.; Satia, I. Is tezepelumab more than just an anti-eosinophil drug? Eur. Respir. J. 2022, 59, 2101700. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Xu, F.; Zhang, G.; Wang, S.; Xu, W. CCR3 monoclonal antibody inhibits airway eosinophilic inflammation and mucus overproduction in a mouse model of asthma. Acta Pharmacol. Sin. 2006, 27, 1594–1599. [Google Scholar] [CrossRef] [PubMed]
- Gauvreau, G.M.; FitzGerald, J.M.; Boulet, L.P.; Watson, R.M.; Hui, L.; Villineuve, H.; Scime, T.X.; Schlatman, A.R.; Obminski, C.; Kum, J.; et al. The effects of a CCR3 inhibitor, AXP1275, on allergen-induced airway responses in adults with mild-to-moderate atopic asthma. Clin. Exp. Allergy 2018, 48, 445–451. [Google Scholar] [CrossRef]
- Neighbour, H.; Boulet, L.-P.; Lemiere, C.; Sehmi, R.; Leigh, R.; Sousa, A.R.; Martin, J.; Dallow, N.; Gilbert, J.; Allen, A.; et al. Safety and efficacy of an oral CCR3 antagonist in patients with asthma and eosinophilic bronchitis: A randomized, placebo-controlled clinical trial. Clin. Exp. Allergy 2014, 44, 508–516. [Google Scholar] [CrossRef]
- Gauvreau, G.M.; Boulet, L.P.; Cockcroft, D.W.; Baatjes, A.; Cote, J.; Deschesnes, F.; Davis, B.; Strinich, T.; Howie, K.; Duong, M.; et al. Antisense therapy against CCR3 and the common beta chain attenuates allergen-induced eosinophilic responses. Am. J. Respir. Crit. Care Med. 2008, 177, 952–958. [Google Scholar] [CrossRef]
- Gauvreau, G.M.; Pageau, R.; Séguin, R.; Carballo, D.; Gauthier, J.; D’Anjou, H.; Campbell, H.; Watson, R.; Mistry, M.; Parry-Billings, M.; et al. Dose-response effects of TPI ASM8 in asthmatics after allergen. Allergy 2011, 66, 1242–1248. [Google Scholar] [CrossRef]
- Pelaia, C.; Crimi, C.; Vatrella, A.; Busceti, M.T.; Gaudio, A.; Garofalo, E.; Bruni, A.; Terracciano, R.; Pelaia, G. New treatments for asthma: From the pathogenic role of prostaglandin D2 to the therapeutic effects of fevipiprant. Pharmacol. Res. 2020, 155, 104490. [Google Scholar] [CrossRef]
- Barnes, N.; Pavord, I.; Chuchalin, A.; Bell, J.; Hunter, M.; Lewis, T.; Parker, D.; Payton, M.; Collins, L.P.; Pettipher, R.; et al. A randomized, double-blind, placebo-controlled study of the CRTH2 antagonist OC000459 in moderate persistent asthma. Clin. Exp. Allergy 2012, 42, 38–48. [Google Scholar] [CrossRef]
- Singh, D.; Cadden, P.; Hunter, M.; Collins, L.P.; Perkins, M.; Pettipher, R.; Townsend, E.; Vinall, S.; O’Connor, B. Inhibition of the asthmatic allergen challenge response by the CRTH2 antagonist OC000459. Eur. Respir. J. 2013, 41, 46–52. [Google Scholar] [CrossRef]
- Pettipher, R.; Hunter, M.G.; Perkins, C.M.; Collins, L.P.; Lewis, T.; Baillet, M.; Steiner, J.; Bell, J.; Payton, M.A. Heightened response of eosinophilic asthmatic patients to the CRTH2 antagonist OC000459. Allergy 2014, 69, 1223–1232. [Google Scholar] [CrossRef] [PubMed]
- Horak, F.; Zieglmayer, P.; Zieglmayer, R.; Lemell, P.; Collins, L.P.; Hunter, M.G.; Steiner, J.; Lewis, T.; Payton, M.A.; Perkins, C.M.; et al. The CRTH2 antagonist OC000459 reduces nasal and ocular symptoms in allergic subjects exposed to grass pollen, a randomised, placebo-controlled, double-blind trial. Allergy 2012, 67, 1572–1579. [Google Scholar] [CrossRef] [PubMed]
- Gonem, S.; Berair, R.; Singapuri, A.; Hartley, R.; Laurencin, M.F.M.; Bacher, G.; Holzhauer, B.; Bourne, M.; Mistry, V.; Pavord, I.D.; et al. Fevipiprant, a prostaglandin D2 receptor 2 antagonist, in patients with persistent eosinophilic asthma: A single-centre, randomised, double-blind, parallel-group, placebo-controlled trial. Lancet Respir. Med. 2016, 4, 699–707. [Google Scholar] [CrossRef]
- Brightling, C.E.; Gaga, M.; Inoue, H.; Li, J.; Maspero, J.; Wenzel, S.; Maitra, S.; Lawrence, D.; Brockhaus, F.; Lehmann, T.; et al. Effectiveness of fevipiprant in reducing exacerbations in patients with severe asthma (LUSTER-1 and LUSTER-2): Two phase 3 randomised controlled trials. Lancet Respir. Med. 2021, 9, 43–56. [Google Scholar] [CrossRef]
- O’Sullivan, J.A.; Chang, A.T.; Youngblood, B.A.; Bochner, B.S. Eosinophil and mast cell Siglecs: From biology to drug target. J. Leukoc. Biol. 2020, 108, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.; Lim, J.C.W.; Soo, H.L.; Stanslas, J. Molecularly targeted therapies for asthma: Current development, challenges and potential clinical translation. Pulm. Pharmacol. Ther. 2016, 40, 52–68. [Google Scholar] [CrossRef]
- Pelaia, C.; Heffler, E.; Crimi, C.; Maglio, A.; Vatrella, A.; Pelaia, G.; Canonica, G.W. Interleukins 4 and 13 in Asthma: Key Pathophysiologic Cytokines and Druggable Molecular Targets. Front. Pharmacol. 2022, 13, 851940. [Google Scholar] [CrossRef]
- Younis, U.S.; Vallorz, E.; Addison, K.J.; Ledford, J.G.; Myrdal, P.B. Preformulation and Evaluation of Tofacitinib as a Therapeutic Treatment for Asthma. AAPS PharmSciTech 2019, 20, 167. [Google Scholar] [CrossRef]
- Kudlacz, E.; Conklyn, M.; Andresen, C.; Whitney-Pickett, C.; Changelian, P. The JAK-3 inhibitor CP-690550 is a potent anti-inflammatory agent in a murine model of pulmonary eosinophilia. Eur. J. Pharmacol. 2008, 582, 154–161. [Google Scholar] [CrossRef]
- Nakamura, Y.; Ghaffar, O.; Olivenstein, R.; Taha, R.A.; Soussi-Gounni, A.; Zhang, D.H.; Ray, A.; Hamid, Q. Gene expression of the GATA-3 transcription factor is increased in atopic asthma. J. Allergy Clin. Immunol. 1999, 103, 215–222. [Google Scholar] [CrossRef]
- Klein-Hessling, S.; Jha, M.K.; Santner-Nanan, B.; Berberich-Siebelt, F.; Baumruker, T.; Schimpl, A.; Serfling, E. Protein kinase A regulates GATA-3-dependent activation of IL-5 gene expression in Th2 cells. J. Immunol. 2003, 170, 2956–2961. [Google Scholar] [CrossRef] [PubMed]
- Sel, S.; Wegmann, M.; Dicke, T.; Sel, S.; Henke, W.; Yildirim, A.O.; Renz, H.; Garn, H. Effective prevention and therapy of experimental allergic asthma using a GATA-3-specific DNAzyme. J. Allergy Clin. Immunol. 2008, 121, 910–916.e5. [Google Scholar] [CrossRef] [PubMed]
- Krug, N.; Hohlfeld, J.M.; Kirsten, A.-M.; Kornmann, O.; Beeh, K.M.; Kappeler, D.; Korn, S.; Ignatenko, S.; Timmer, W.; Rogon, C.; et al. Allergen-Induced Asthmatic Responses Modified by a GATA3-Specific DNAzyme. N. Engl. J. Med. 2015, 372, 1987–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vatrella, A.; Maglio, A.; Pelaia, C.; Ciampo, L.; Pelaia, G.; Vitale, C. Eosinophilic inflammation: An Appealing Target for Pharmacologic Treatments in Severe Asthma. Biomedicines 2022, 10, 2181. https://doi.org/10.3390/biomedicines10092181
Vatrella A, Maglio A, Pelaia C, Ciampo L, Pelaia G, Vitale C. Eosinophilic inflammation: An Appealing Target for Pharmacologic Treatments in Severe Asthma. Biomedicines. 2022; 10(9):2181. https://doi.org/10.3390/biomedicines10092181
Chicago/Turabian StyleVatrella, Alessandro, Angelantonio Maglio, Corrado Pelaia, Luigi Ciampo, Giulia Pelaia, and Carolina Vitale. 2022. "Eosinophilic inflammation: An Appealing Target for Pharmacologic Treatments in Severe Asthma" Biomedicines 10, no. 9: 2181. https://doi.org/10.3390/biomedicines10092181