
Effects of Thyroid Hormones on Lipid Metabolism Pathologies in Non-Alcoholic Fatty Liver Disease

 ,
,
Abstract
:1. Thyroid Hormones and Action
1.1. Thyroid Hormones
1.2. TH Actions
2. TH and Lipid Metabolism in the Liver
2.1. Lipid Metabolism in the Liver
2.2. THs and Lipid Metabolism in the Liver
2.2.1. Regulation of Fatty Acid Uptake by THs
2.2.2. De Novel Lipogenesis (DNL)
2.2.3. β-Oxidation
2.2.4. Cholesterol Homeostasis
3. Pathological Roles of TH Associated with Lipid Metabolism in the Liver
3.1. Lipid Metabolism and Liver Diseases: Non-Alcoholic Fatty Liver Disease (NAFLD)
3.2. Correlations between THs and NAFLD
4. Drugs Targeting THs and Their Clinical Applications in NAFLD
4.1. Management of NAFLD
4.2. Therapeutic Potential of THs, TH Mimetics, and TH Metabolites in NAFLD
4.2.1. GC-1
4.2.2. GC-24
4.2.3. KB141
4.2.4. KB2115
4.2.5. MB07811
4.2.6. MGL-3196
4.2.7. 3,5,-L-diiodothyronine (T2)
4.2.8. 3-iodothyronamine (T1AM)
4.3. Clinical Application for Thyroid Hormones Status and NAFLD
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ortiga-Carvalho, T.M.; Sidhaye, A.R.; Wondisford, F.E. Thyroid hormone receptors and resistance to thyroid hormone disorders. Nat. Rev. Endocrinol. 2014, 10, 582–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, A.; Martinez, M.E.; Ng, L.; Forrest, D. Thyroid Hormone Deiodinases: Dynamic Switches in Developmental Transitions. Endocrinology 2021, 162, bqab091. [Google Scholar] [CrossRef] [PubMed]
- Lazar, M.A.; Berrodin, T.J.; Harding, H.P. Differential DNA binding by monomeric, homodimeric, and potentially heteromeric forms of the thyroid hormone receptor. Mol. Cell. Biol. 1991, 11, 5005–5015. [Google Scholar] [CrossRef] [PubMed]
- Berrodin, T.J.; Marks, M.S.; Ozato, K.; Linney, E.; Lazar, M.A. Heterodimerization among thyroid hormone receptor, retinoic acid receptor, retinoid X receptor, chicken ovalbumin upstream promoter transcription factor, and an endogenous liver protein. Mol. Endocrinol. 1992, 6, 1468–1478. [Google Scholar] [CrossRef] [PubMed]
- Segal, J.; Ingbar, S.H. In vivo stimulation of sugar uptake in rat thymocytes. An extranuclear action of 3,5,3′-triiodothyronine. J. Clin. Investig. 1985, 76, 1575–1580. [Google Scholar] [CrossRef] [Green Version]
- Flamant, F.; Cheng, S.Y.; Hollenberg, A.N.; Moeller, L.C.; Samarut, J.; Wondisford, F.E.; Yen, P.M.; Refetoff, S. Thyroid Hormone Signaling Pathways: Time for a More Precise Nomenclature. Endocrinology 2017, 158, 2052–2057. [Google Scholar] [CrossRef]
- Perez, P.; Schönthal, A.; Aranda, A. Repression of c-fos gene expression by thyroid hormone and retinoic acid receptors. J. Biol. Chem. 1993, 268, 23538–23543. [Google Scholar] [CrossRef]
- Méndez-Pertuz, M.; Sánchez-Pacheco, A.; Aranda, A. The thyroid hormone receptor antagonizes CREB-mediated transcription. EMBO J. 2003, 22, 3102–3112. [Google Scholar] [CrossRef] [Green Version]
- Lasa, M.; Gil-Araujo, B.; Palafox, M.; Aranda, A. Thyroid hormone antagonizes tumor necrosis factor-alpha signaling in pituitary cells through the induction of dual specificity phosphatase 1. Mol. Endocrinol. 2010, 24, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Davis, P.J.; Davis, F.B.; Cody, V. Membrane receptors mediating thyroid hormone action. Trends Endocrinol. Metab. 2005, 16, 429–435. [Google Scholar] [CrossRef]
- Hiroi, Y.; Kim, H.H.; Ying, H.; Furuya, F.; Huang, Z.; Simoncini, T.; Noma, K.; Ueki, K.; Nguyen, N.H.; Scanlan, T.S.; et al. Rapid nongenomic actions of thyroid hormone. Proc. Natl. Acad. Sci. USA 2006, 103, 14104–14109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergh, J.J.; Lin, H.Y.; Lansing, L.; Mohamed, S.N.; Davis, F.B.; Mousa, S.; Davis, P.J. Integrin alphaVbeta3 contains a cell surface receptor site for thyroid hormone that is linked to activation of mitogen-activated protein kinase and induction of angiogenesis. Endocrinology 2005, 146, 2864–2871. [Google Scholar] [CrossRef]
- Davis, P.J.; Davis, F.B.; Mousa, S.A.; Luidens, M.K.; Lin, H.Y. Membrane receptor for thyroid hormone: Physiologic and pharmacologic implications. Annu. Rev. Pharm. Toxicol. 2011, 51, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.C.; Chen, C.Y.; Tsai, M.M.; Tsai, C.Y.; Lin, K.H. Molecular functions of thyroid hormones and their clinical significance in liver-related diseases. Biomed. Res. Int. 2013, 2013, 601361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, R.L.; Hunt, H.M.; Hurxthal, L. Blood Cholesterol Values in Hyperthyroidism and Hypothyroidism—Their Significance. N. Engl. J. Med. 1930, 203, 1273–1278. [Google Scholar] [CrossRef]
- Muls, E.; Rosseneu, M.; Blaton, V.; Lesaffre, E.; Lamberigts, G.; de Moor, P. Serum lipids and apolipoproteins A-I, A-II and B in primary hypothyroidism before and during treatment. Eur. J. Clin. Investig. 1984, 14, 12–15. [Google Scholar] [CrossRef]
- Friis, T.; Pedersen, L.R. Serum lipids in hyper- and hypothyroidism before and after treatment. Clin. Chim. Acta 1987, 162, 155–163. [Google Scholar] [CrossRef]
- Packard, C.J.; Shepherd, J.; Lindsay, G.M.; Gaw, A.; Taskinen, M.R. Thyroid replacement therapy and its influence on postheparin plasma lipases and apolipoprotein-B metabolism in hypothyroidism. J. Clin. Endocrinol. Metab. 1993, 76, 1209–1216. [Google Scholar] [CrossRef]
- Wiseman, S.A.; Powell, J.T.; Humphries, S.E.; Press, M. The magnitude of the hypercholesterolemia of hypothyroidism is associated with variation in the low density lipoprotein receptor gene. J. Clin. Endocrinol. Metab. 1993, 77, 108–112. [Google Scholar] [CrossRef]
- Pazos, F.; Alvarez, J.J.; Rubiés-Prat, J.; Varela, C.; Lasunción, M.A. Long-term thyroid replacement therapy and levels of lipoprotein(a) and other lipoproteins. J. Clin. Endocrinol. Metab. 1995, 80, 562–566. [Google Scholar] [CrossRef]
- Araki, O.; Ying, H.; Zhu, X.G.; Willingham, M.C.; Cheng, S.Y. Distinct dysregulation of lipid metabolism by unliganded thyroid hormone receptor isoforms. Mol. Endocrinol. 2009, 23, 308–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grefhorst, A.; van de Peppel, I.P.; Larsen, L.E.; Jonker, J.W.; Holleboom, A.G. The Role of Lipophagy in the Development and Treatment of Non-Alcoholic Fatty Liver Disease. Front. Endocrinol. 2020, 11, 601627. [Google Scholar] [CrossRef] [PubMed]
- Hajri, T.; Abumrad, N.A. Fatty acid transport across membranes: Relevance to nutrition and metabolic pathology. Annu. Rev. Nutr. 2002, 22, 383–415. [Google Scholar] [CrossRef] [PubMed]
- Klieverik, L.P.; Coomans, C.P.; Endert, E.; Sauerwein, H.P.; Havekes, L.M.; Voshol, P.J.; Rensen, P.C.; Romijn, J.A.; Kalsbeek, A.; Fliers, E. Thyroid hormone effects on whole-body energy homeostasis and tissue-specific fatty acid uptake in vivo. Endocrinology 2009, 150, 5639–5648. [Google Scholar] [CrossRef] [Green Version]
- Santana-Farré, R.; Mirecki-Garrido, M.; Bocos, C.; Henríquez-Hernández, L.A.; Kahlon, N.; Herrera, E.; Norstedt, G.; Parini, P.; Flores-Morales, A.; Fernández-Pérez, L. Influence of neonatal hypothyroidism on hepatic gene expression and lipid metabolism in adulthood. PLoS ONE 2012, 7, e37386. [Google Scholar] [CrossRef] [Green Version]
- Perra, A.; Simbula, G.; Simbula, M.; Pibiri, M.; Kowalik, M.A.; Sulas, P.; Cocco, M.T.; Ledda-Columbano, G.M.; Columbano, A. Thyroid hormone (T3) and TRbeta agonist GC-1 inhibit/reverse nonalcoholic fatty liver in rats. FASEB J. 2008, 22, 2981–2989. [Google Scholar] [CrossRef]
- Shu, L.; Hoo, R.L.C.; Wu, X.; Pan, Y.; Lee, I.P.C.; Cheong, L.Y.; Bornstein, S.R.; Rong, X.; Guo, J.; Xu, A. A-FABP mediates adaptive thermogenesis by promoting intracellular activation of thyroid hormones in brown adipocytes. Nat. Commun. 2017, 8, 14147. [Google Scholar] [CrossRef] [Green Version]
- Giudetti, A.M.; Leo, M.; Siculella, L.; Gnoni, G.V. Hypothyroidism down-regulates mitochondrial citrate carrier activity and expression in rat liver. Biochim. Biophys. Acta 2006, 1761, 484–491. [Google Scholar] [CrossRef]
- Siculella, L.; Sabetta, S.; Giudetti, A.M.; Gnoni, G.V. Hypothyroidism reduces tricarboxylate carrier activity and expression in rat liver mitochondria by reducing nuclear transcription rate and splicing efficiency. J. Biol. Chem. 2006, 281, 19072–19080. [Google Scholar] [CrossRef] [Green Version]
- Paradies, G.; Ruggiero, F.M. Effect of hyperthyroidism on the transport of pyruvate in rat-heart mitochondria. Biochim. Biophys. Acta 1988, 935, 79–86. [Google Scholar] [CrossRef]
- Paradies, G.; Ruggiero, F.M. Enhanced activity of the tricarboxylate carrier and modification of lipids in hepatic mitochondria from hyperthyroid rats. Arch. Biochem. Biophys. 1990, 278, 425–430. [Google Scholar] [CrossRef]
- Spence, J.T.; Pitot, H.C.; Zalitis, G. Regulation of ATP-citrate lyase in primary cultures of adult rat hepatocytes. J. Biol. Chem. 1979, 254, 12169–12173. [Google Scholar] [CrossRef]
- Gnoni, A.; Siculella, L.; Paglialonga, G.; Damiano, F.; Giudetti, A.M. 3,5-diiodo-L-thyronine increases de novo lipogenesis in liver from hypothyroid rats by SREBP-1 and ChREBP-mediated transcriptional mechanisms. IUBMB Life 2019, 71, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.-A.; Lee, J.-J.; Park, S.-W.; Ahn, Y.-H.; Kim, K.-S. The Roles of Sterol Regulatory Element-binding Proteins in the Transactivation of the Rat ATP Citrate-Lyase Promoter. J. Biol. Chem. 2000, 275, 30280–30286. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Yamada, M.; Matsumoto, S.; Monden, T.; Satoh, T.; Mori, M. Mouse Sterol Response Element Binding Protein-1c Gene Expression Is Negatively Regulated by Thyroid Hormone. Endocrinology 2006, 147, 4292–4302. [Google Scholar] [CrossRef]
- Gnoni, G.V.; Rochira, A.; Leone, A.; Damiano, F.; Marsigliante, S.; Siculella, L. 3,5,3′triiodo-L-thyronine induces SREBP-1 expression by non-genomic actions in human HEP G2 cells. J. Cell. Physiol. 2012, 227, 2388–2397. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, L.; Hillgartner, F.B. SREBP-1 integrates the actions of thyroid hormone, insulin, cAMP, and medium-chain fatty acids on ACCalpha transcription in hepatocytes. J. Lipid Res. 2003, 44, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Radenne, A.; Akpa, M.; Martel, C.; Sawadogo, S.; Mauvoisin, D.; Mounier, C. Hepatic regulation of fatty acid synthase by insulin and T3: Evidence for T3 genomic and nongenomic actions. Am. J. Physiol.-Endocrinol. Metab. 2008, 295, E884–E894. [Google Scholar] [CrossRef] [Green Version]
- Jump, D.B.; Thelen, A.P.; Mater, M.K. Functional Interaction between Sterol Regulatory Element-binding Protein-1c, Nuclear Factor Y, and 3,5,3′-Triiodothyronine Nuclear Receptors. J. Biol. Chem. 2001, 276, 34419–34427. [Google Scholar] [CrossRef] [Green Version]
- Brenta, G.; Berg, G.; Miksztowicz, V.; Lopez, G.; Lucero, D.; Faingold, C.; Murakami, M.; Machima, T.; Nakajima, K.; Schreier, L. Atherogenic Lipoproteins in Subclinical Hypothyroidism and Their Relationship with Hepatic Lipase Activity: Response to Replacement Treatment with Levothyroxine. Thyroid 2016, 26, 365–372. [Google Scholar] [CrossRef]
- Grasselli, E.; Voci, A.; Demori, I.; Vecchione, G.; Compalati, A.D.; Gallo, G.; Goglia, F.; De Matteis, R.; Silvestri, E.; Vergani, L. Triglyceride Mobilization from Lipid Droplets Sustains the Anti-Steatotic Action of Iodothyronines in Cultured Rat Hepatocytes. Front. Physiol. 2016, 6, 418. [Google Scholar] [CrossRef] [PubMed]
- Saggerson, E.D.; Carpenter, C.A. Carnitine palmitoyltransferase in liver and five extrahepatic tissues in the rat. Inhibition by dl-2-bromopalmitoyl-CoA and effect of hypothyroidism. Biochem. J. 1986, 236, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Mynatt, R.L.; Park, E.A.; Thorngate, F.E.; Das, H.K.; Cook, G.A. Changes in carnitine palmitoyltransferase-I mRNA abundance produced by hyperthyroidism and hypothyroidism parallel changes in activity. Biochem. Biophys. Res. Commun. 1994, 201, 932–937. [Google Scholar] [CrossRef] [PubMed]
- Jackson-Hayes, L.; Song, S.; Lavrentyev, E.N.; Jansen, M.S.; Hillgartner, F.B.; Tian, L.; Wood, P.A.; Cook, G.A.; Park, E.A. A thyroid hormone response unit formed between the promoter and first intron of the carnitine palmitoyltransferase-Ialpha gene mediates the liver-specific induction by thyroid hormone. J. Biol. Chem. 2003, 278, 7964–7972. [Google Scholar] [CrossRef] [Green Version]
- Potthoff Matthew, J.; Inagaki, T.; Satapati, S.; Ding, X.; He, T.; Goetz, R.; Mohammadi, M.; Finck Brian, N.; Mangelsdorf David, J.; Kliewer Steven, A.; et al. FGF21 induces PGC-1α and regulates carbohydrate and fatty acid metabolism during the adaptive starvation response. Proc. Natl. Acad. Sci. USA 2009, 106, 10853–10858. [Google Scholar] [CrossRef] [Green Version]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a novel metabolic regulator. J. Clin. Investig. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.C.; Chen, W.; Cooper, A.D. LDL receptor-related protein mediates cell-surface clustering and hepatic sequestration of chylomicron remnants in LDLR-deficient mice. J. Clin. Investig. 2001, 107, 1387–1394. [Google Scholar] [CrossRef] [Green Version]
- Lopez, D.; Abisambra Socarrás, J.F.; Bedi, M.; Ness, G.C. Activation of the hepatic LDL receptor promoter by thyroid hormone. Biochim. Biophys. Acta 2007, 1771, 1216–1225. [Google Scholar] [CrossRef]
- Moon, J.H.; Kim, H.J.; Kim, H.M.; Choi, S.H.; Lim, S.; Park, Y.J.; Jang, H.C.; Cha, B.S. Decreased expression of hepatic low-density lipoprotein receptor-related protein 1 in hypothyroidism: A novel mechanism of atherogenic dyslipidemia in hypothyroidism. Thyroid 2013, 23, 1057–1065. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.J.; Osborne, T.F. Thyroid hormone regulation and cholesterol metabolism are connected through Sterol Regulatory Element-Binding Protein-2 (SREBP-2). J. Biol. Chem. 2003, 278, 34114–34118. [Google Scholar] [CrossRef] [Green Version]
- Alborn, W.E.; Cao, G.; Careskey, H.E.; Qian, Y.-W.; Subramaniam, D.R.; Davies, J.; Conner, E.M.; Konrad, R.J. Serum Proprotein Convertase Subtilisin Kexin Type 9 Is Correlated Directly with Serum LDL Cholesterol. Clin. Chem. 2007, 53, 1814–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazaeli, M.; Khoshdel, A.; Shafiepour, M.; Rohban, M. The influence of subclinical hypothyroidism on serum lipid profile, PCSK9 levels and CD36 expression on monocytes. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, A.M.; Koca, A.O.; Beyan, E.; Dogan, O.; Karakaya, S.; Aksoz, Z.; Ertuğrul, D.T. Association of serum proprotein convertase Subtilisin/Kexin Type 9 (PCSK9) level with thyroid function disorders. Eur. Rev. Med. Pharm. Sci. 2021, 25, 5511–5517. [Google Scholar] [CrossRef]
- Lin, J.Z.; Martagón, A.J.; Hsueh, W.A.; Baxter, J.D.; Gustafsson, J.-Å.; Webb, P.; Phillips, K.J. Thyroid Hormone Receptor Agonists Reduce Serum Cholesterol Independent of the LDL Receptor. Endocrinology 2012, 153, 6136–6144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lammel Lindemann, J.A.; Angajala, A.; Engler, D.A.; Webb, P.; Ayers, S.D. Thyroid hormone induction of human cholesterol 7 alpha-hydroxylase (Cyp7a1) in vitro. Mol. Cell. Endocrinol. 2014, 388, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Gälman, C.; Bonde, Y.; Matasconi, M.; Angelin, B.; Rudling, M. Dramatically increased intestinal absorption of cholesterol following hypophysectomy is normalized by thyroid hormone. Gastroenterology 2008, 134, 1127–1136. [Google Scholar] [CrossRef]
- Graf, G.A.; Yu, L.; Li, W.P.; Gerard, R.; Tuma, P.L.; Cohen, J.C.; Hobbs, H.H. ABCG5 and ABCG8 are obligate heterodimers for protein trafficking and biliary cholesterol excretion. J. Biol. Chem. 2003, 278, 48275–48282. [Google Scholar] [CrossRef] [Green Version]
- Bonde, Y.; Plösch, T.; Kuipers, F.; Angelin, B.; Rudling, M. Stimulation of murine biliary cholesterol secretion by thyroid hormone is dependent on a functional ABCG5/G8 complex. Hepatology 2012, 56, 1828–1837. [Google Scholar] [CrossRef] [Green Version]
- Tao, R.; Xiong, X.; DePinho, R.A.; Deng, C.-X.; Dong, X.C. Hepatic SREBP-2 and cholesterol biosynthesis are regulated by FoxO3 and Sirt6. J. Lipid Res. 2013, 54, 2745–2753. [Google Scholar] [CrossRef] [Green Version]
- Markovic, M.; Ben-Shabat, S.; Aponick, A.; Zimmermann, E.M.; Dahan, A. Lipids and Lipid-Processing Pathways in Drug Delivery and Therapeutics. Int. J. Mol. Sci. 2020, 21, 3248. [Google Scholar] [CrossRef]
- Tang, Q.-Q. Lipid metabolism and diseases. Sci. Bull. 2016, 61, 1471–1472. [Google Scholar] [CrossRef]
- Powell, E.E.; Wong, V.W.; Rinella, M. Non-alcoholic fatty liver disease. Lancet 2021, 397, 2212–2224. [Google Scholar] [CrossRef]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef] [PubMed]
- Estes, C.; Razavi, H.; Loomba, R.; Younossi, Z.; Sanyal, A.J. Modeling the epidemic of nonalcoholic fatty liver disease demonstrates an exponential increase in burden of disease. Hepatology 2018, 67, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Paik, J.M.; Golabi, P.; Younossi, Y.; Mishra, A.; Younossi, Z.M. Changes in the Global Burden of Chronic Liver Diseases from 2012 to 2017: The Growing Impact of NAFLD. Hepatology 2020, 72, 1605–1616. [Google Scholar] [CrossRef]
- Goldberg, D.; Ditah, I.C.; Saeian, K.; Lalehzari, M.; Aronsohn, A.; Gorospe, E.C.; Charlton, M. Changes in the Prevalence of Hepatitis C Virus Infection, Nonalcoholic Steatohepatitis, and Alcoholic Liver Disease Among Patients with Cirrhosis or Liver Failure on the Waitlist for Liver Transplantation. Gastroenterology 2017, 152, 1090–1099.e1. [Google Scholar] [CrossRef] [Green Version]
- Lonardo, A.; Nascimbeni, F.; Mantovani, A.; Targher, G. Hypertension, diabetes, atherosclerosis and NASH: Cause or consequence? J. Hepatol. 2018, 68, 335–352. [Google Scholar] [CrossRef]
- Al-Muzafar, H.M.; Alshehri, F.S.; Amin, K.A. The role of pioglitazone in antioxidant, anti-inflammatory, and insulin sensitivity in a high fat-carbohydrate diet-induced rat model of insulin resistance. Braz. J. Med. Biol. Res. 2021, 54, e10782. [Google Scholar] [CrossRef]
- Chung, G.E.; Kim, D.; Kim, W.; Yim, J.Y.; Park, M.J.; Kim, Y.J.; Yoon, J.H.; Lee, H.S. Non-alcoholic fatty liver disease across the spectrum of hypothyroidism. J. Hepatol. 2012, 57, 150–156. [Google Scholar] [CrossRef]
- Pagadala, M.R.; Zein, C.O.; Dasarathy, S.; Yerian, L.M.; Lopez, R.; McCullough, A.J. Prevalence of hypothyroidism in nonalcoholic fatty liver disease. Dig. Dis. Sci. 2012, 57, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Bano, A.; Chaker, L.; Plompen, E.P.; Hofman, A.; Dehghan, A.; Franco, O.H.; Janssen, H.L.; Darwish Murad, S.; Peeters, R.P. Thyroid Function and the Risk of Nonalcoholic Fatty Liver Disease: The Rotterdam Study. J. Clin. Endocrinol. Metab. 2016, 101, 3204–3211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manka, P.; Bechmann, L.; Best, J.; Sydor, S.; Claridge, L.C.; Coombes, J.D.; Canbay, A.; Moeller, L.; Gerken, G.; Wedemeyer, H.; et al. Low Free Triiodothyronine Is Associated with Advanced Fibrosis in Patients at High Risk for Nonalcoholic Steatohepatitis. Dig. Dis. Sci. 2019, 64, 2351–2358. [Google Scholar] [CrossRef] [PubMed]
- Eshraghian, A.; Dabbaghmanesh, M.H.; Eshraghian, H.; Fattahi, M.R.; Omrani, G.R. Nonalcoholic fatty liver disease in a cluster of Iranian population: Thyroid status and metabolic risk factors. Arch. Iran. Med. 2013, 16, 584–589. [Google Scholar] [PubMed]
- Posadas-Romero, C.; Jorge-Galarza, E.; Posadas-Sánchez, R.; Acuña-Valerio, J.; Juárez-Rojas, J.G.; Kimura-Hayama, E.; Medina-Urrutia, A.; Cardoso-Saldaña, G.C. Fatty liver largely explains associations of subclinical hypothyroidism with insulin resistance, metabolic syndrome, and subclinical coronary atherosclerosis. Eur. J. Endocrinol. 2014, 171, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Jaruvongvanich, V.; Sanguankeo, A.; Upala, S. Nonalcoholic Fatty Liver Disease Is Not Associated with Thyroid Hormone Levels and Hypothyroidism: A Systematic Review and Meta-Analysis. Eur. Thyroid. J. 2017, 6, 208–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Nascimbeni, F.; Lonardo, A.; Zoppini, G.; Bonora, E.; Mantzoros, C.S.; Targher, G. Association Between Primary Hypothyroidism and Nonalcoholic Fatty Liver Disease: A Systematic Review and Meta-Analysis. Thyroid 2018, 28, 1270–1284. [Google Scholar] [CrossRef]
- Lonardo, A.; Ballestri, S.; Mantovani, A.; Nascimbeni, F.; Lugari, S.; Targher, G. Pathogenesis of hypothyroidism-induced NAFLD: Evidence for a distinct disease entity? Dig. Liver. Dis. 2019, 51, 462–470. [Google Scholar] [CrossRef]
- Romero-Gómez, M.; Zelber-Sagi, S.; Trenell, M. Treatment of NAFLD with diet, physical activity and exercise. J. Hepatol. 2017, 67, 829–846. [Google Scholar] [CrossRef] [Green Version]
- Głuszyńska, P.; Lemancewicz, D.; Dzięcioł, J.B.; Razak Hady, H. Non-Alcoholic Fatty Liver Disease (NAFLD) and Bariatric/Metabolic Surgery as Its Treatment Option: A Review. J. Clin. Med. 2021, 10, 5721. [Google Scholar] [CrossRef]
- Bril, F.; Biernacki, D.M.; Kalavalapalli, S.; Lomonaco, R.; Subbarayan, S.K.; Lai, J.; Tio, F.; Suman, A.; Orsak, B.K.; Hecht, J.; et al. Role of Vitamin E for Nonalcoholic Steatohepatitis in Patients with Type 2 Diabetes: A Randomized Controlled Trial. Diabetes Care 2019, 42, 1481–1488. [Google Scholar] [CrossRef] [PubMed]
- Cable, E.E.; Finn, P.D.; Stebbins, J.W.; Hou, J.; Ito, B.R.; van Poelje, P.D.; Linemeyer, D.L.; Erion, M.D. Reduction of hepatic steatosis in rats and mice after treatment with a liver-targeted thyroid hormone receptor agonist. Hepatology 2009, 49, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Iannucci, L.F.; Cioffi, F.; Senese, R.; Goglia, F.; Lanni, A.; Yen, P.M.; Sinha, R.A. Metabolomic analysis shows differential hepatic effects of T2 and T3 in rats after short-term feeding with high fat diet. Sci. Rep. 2017, 7, 2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalik, M.A.; Columbano, A.; Perra, A. Thyroid Hormones, Thyromimetics and Their Metabolites in the Treatment of Liver Disease. Front. Endocrinol. 2018, 9, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, M.J.; Amano, I.; Hollenberg, A.N. Thyroid Hormone Signaling and the Liver. Hepatology 2020, 72, 742–752. [Google Scholar] [CrossRef]
- Chiellini, G.; Apriletti, J.W.; Yoshihara, H.A.; Baxter, J.D.; Ribeiro, R.C.J.; Scanlan, T.S. A high-affinity subtype-selective agonist ligand for the thyroid hormone receptor. Chem. Biol. 1998, 5, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Trost, S.U.; Swanson, E.; Gloss, B.; Wang-Iverson, D.B.; Zhang, H.; Volodarsky, T.; Grover, G.J.; Baxter, J.D.; Chiellini, G.; Scanlan, T.S.; et al. The Thyroid Hormone Receptor-β-Selective Agonist GC-1 Differentially Affects Plasma Lipids and Cardiac Activity. Endocrinology 2000, 141, 3057–3064. [Google Scholar] [CrossRef]
- Martagón, A.J.; Lin, J.Z.; Cimini, S.L.; Webb, P.; Phillips, K.J. The amelioration of hepatic steatosis by thyroid hormone receptor agonists is insufficient to restore insulin sensitivity in ob/ob mice. PLoS ONE 2015, 10, e0122987. [Google Scholar] [CrossRef]
- Vatner, D.F.; Weismann, D.; Beddow, S.A.; Kumashiro, N.; Erion, D.M.; Liao, X.H.; Grover, G.J.; Webb, P.; Phillips, K.J.; Weiss, R.E.; et al. Thyroid hormone receptor-β agonists prevent hepatic steatosis in fat-fed rats but impair insulin sensitivity via discrete pathways. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E89–E100. [Google Scholar] [CrossRef]
- Borngraeber, S.; Budny, M.J.; Chiellini, G.; Cunha-Lima, S.T.; Togashi, M.; Webb, P.; Baxter, J.D.; Scanlan, T.S.; Fletterick, R.J. Ligand selectivity by seeking hydrophobicity in thyroid hormone receptor. Proc. Natl. Acad. Sci. USA 2003, 100, 15358–15363. [Google Scholar] [CrossRef] [Green Version]
- Amorim, B.S.; Ueta, C.B.; Freitas, B.C.; Nassif, R.J.; Gouveia, C.H.; Christoffolete, M.A.; Moriscot, A.S.; Lancelloti, C.L.; Llimona, F.; Barbeiro, H.V.; et al. A TRbeta-selective agonist confers resistance to diet-induced obesity. J. Endocrinol. 2009, 203, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grijota-Martínez, C.; Samarut, E.; Scanlan, T.S.; Morte, B.; Bernal, J. In Vivo Activity of the Thyroid Hormone Receptor β- and α-Selective Agonists GC-24 and CO23 on Rat Liver, Heart, and Brain. Endocrinology 2011, 152, 1136–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grover, G.J.; Mellström, K.; Ye, L.; Malm, J.; Li, Y.L.; Bladh, L.G.; Sleph, P.G.; Smith, M.A.; George, R.; Vennström, B.; et al. Selective thyroid hormone receptor-beta activation: A strategy for reduction of weight, cholesterol, and lipoprotein (a) with reduced cardiovascular liability. Proc. Natl. Acad. Sci. USA 2003, 100, 10067–10072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grover, G.J.; Mellstrom, K.; Malm, J. Development of the thyroid hormone receptor beta-subtype agonist KB-141: A strategy for body weight reduction and lipid lowering with minimal cardiac side effects. Cardiovasc. Drug Rev. 2005, 23, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Berkenstam, A.; Kristensen, J.; Mellström, K.; Carlsson, B.; Malm, J.; Rehnmark, S.; Garg, N.; Andersson Carl, M.; Rudling, M.; Sjöberg, F.; et al. The thyroid hormone mimetic compound KB2115 lowers plasma LDL cholesterol and stimulates bile acid synthesis without cardiac effects in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 663–667. [Google Scholar] [CrossRef] [Green Version]
- Sjouke, B.; Langslet, G.; Ceska, R.; Nicholls, S.J.; Nissen, S.E.; Öhlander, M.; Ladenson, P.W.; Olsson, A.G.; Hovingh, G.K.; Kastelein, J.J. Eprotirome in patients with familial hypercholesterolaemia (the AKKA trial): A randomised, double-blind, placebo-controlled phase 3 study. Lancet Diabetes Endocrinol. 2014, 2, 455–463. [Google Scholar] [CrossRef]
- Ladenson, P.W.; Kristensen, J.D.; Ridgway, E.C.; Olsson, A.G.; Carlsson, B.; Klein, I.; Baxter, J.D.; Angelin, B. Use of the Thyroid Hormone Analogue Eprotirome in Statin-Treated Dyslipidemia. N. Engl. J. Med. 2010, 362, 906–916. [Google Scholar] [CrossRef] [Green Version]
- Erion, M.D.; Cable, E.E.; Ito, B.R.; Jiang, H.; Fujitaki, J.M.; Finn, P.D.; Zhang, B.H.; Hou, J.; Boyer, S.H.; van Poelje, P.D.; et al. Targeting thyroid hormone receptor-beta agonists to the liver reduces cholesterol and triglycerides and improves the therapeutic index. Proc. Natl. Acad. Sci. USA 2007, 104, 15490–15495. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.J.; Pietranico-Cole, S.; Larigan, J.D.; Haynes, N.-E.; Reynolds, C.H.; Scott, N.; Vermeulen, J.; Dvorozniak, M.; Conde-Knape, K.; Huang, K.-S.; et al. Discovery of 2-[3,5-Dichloro-4-(5-isopropyl-6-oxo-1,6-dihydropyridazin-3-yloxy)phenyl]-3,5-dioxo-2,3,4,5-tetrahydro[1,2,4]triazine-6-carbonitrile (MGL-3196), a Highly Selective Thyroid Hormone Receptor β Agonist in Clinical Trials for the Treatment of Dyslipidemia. J. Med. Chem. 2014, 57, 3912–3923. [Google Scholar] [CrossRef]
- Harrison, S.A.; Bashir, M.R.; Guy, C.D.; Zhou, R.; Moylan, C.A.; Frias, J.P.; Alkhouri, N.; Bansal, M.B.; Baum, S.; Neuschwander-Tetri, B.A.; et al. Resmetirom (MGL-3196) for the treatment of non-alcoholic steatohepatitis: A multicentre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2019, 394, 2012–2024. [Google Scholar] [CrossRef]
- Harrison, S.A.; Bashir, M.; Moussa, S.E.; McCarty, K.; Pablo Frias, J.; Taub, R.; Alkhouri, N. Effects of Resmetirom on Noninvasive Endpoints in a 36-Week Phase 2 Active Treatment Extension Study in Patients with NASH. Hepatol. Commun. 2021, 5, 573–588. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.G.; Sokolov, J.; Chin, W.W. 3,5-Diiodo-L-thyronine (T2) has selective thyromimetic effects in vivo and in vitro. J. Mol. Endocrinol. 1997, 19, 137–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollica, M.P.; Lionetti, L.; Moreno, M.; Lombardi, A.; De Lange, P.; Antonelli, A.; Lanni, A.; Cavaliere, G.; Barletta, A.; Goglia, F. 3,5-diiodo-l-thyronine, by modulating mitochondrial functions, reverses hepatic fat accumulation in rats fed a high-fat diet. J. Hepatol. 2009, 51, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Lanni, A.; Moreno, M.; Lombardi, A.; de Lange, P.; Silvestri, E.; Ragni, M.; Farina, P.; Baccari, G.C.; Fallahi, P.; Antonelli, A.; et al. 3,5-diiodo-L-thyronine powerfully reduces adiposity in rats by increasing the burning of fats. FASEB J. 2005, 19, 1552–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lange, P.; Cioffi, F.; Senese, R.; Moreno, M.; Lombardi, A.; Silvestri, E.; De Matteis, R.; Lionetti, L.; Mollica, M.P.; Goglia, F.; et al. Nonthyrotoxic prevention of diet-induced insulin resistance by 3,5-diiodo-L-thyronine in rats. Diabetes 2011, 60, 2730–2739. [Google Scholar] [CrossRef] [Green Version]
- Michan, S.; Sinclair, D. Sirtuins in mammals: Insights into their biological function. Biochem. J. 2007, 404, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Thakran, S.; Sharma, P.; Attia, R.R.; Hori, R.T.; Deng, X.; Elam, M.B.; Park, E.A. Role of sirtuin 1 in the regulation of hepatic gene expression by thyroid hormone. J. Biol. Chem. 2013, 288, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Suh, J.H.; Sieglaff, D.H.; Zhang, A.; Xia, X.; Cvoro, A.; Winnier, G.E.; Webb, P. SIRT1 is a direct coactivator of thyroid hormone receptor β1 with gene-specific actions. PLoS ONE 2013, 8, e70097. [Google Scholar] [CrossRef]
- Senese, R.; Cioffi, F.; de Lange, P.; Leanza, C.; Iannucci, L.F.; Silvestri, E.; Moreno, M.; Lombardi, A.; Goglia, F.; Lanni, A. Both 3,5-Diiodo-L-Thyronine and 3,5,3’-Triiodo-L-Thyronine Prevent Short-term Hepatic Lipid Accumulation via Distinct Mechanisms in Rats Being Fed a High-Fat Diet. Front. Physiol. 2017, 8, 706. [Google Scholar] [CrossRef]
- Jonas, W.; Lietzow, J.; Wohlgemuth, F.; Hoefig, C.S.; Wiedmer, P.; Schweizer, U.; Köhrle, J.; Schürmann, A. 3,5-Diiodo-L-Thyronine (3,5-T2) Exerts Thyromimetic Effects on Hypothalamus-Pituitary-Thyroid Axis, Body Composition, and Energy Metabolism in Male Diet-Induced Obese Mice. Endocrinology 2015, 156, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Scanlan, T.S. Minireview: 3-Iodothyronamine (T1AM): A new player on the thyroid endocrine team? Endocrinology 2009, 150, 1108–1111. [Google Scholar] [CrossRef] [Green Version]
- Zucchi, R.; Chiellini, G.; Scanlan, T.S.; Grandy, D.K. Trace amine-associated receptors and their ligands. Br. J. Pharmacol. 2006, 149, 967–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanlan, T.S.; Suchland, K.L.; Hart, M.E.; Chiellini, G.; Huang, Y.; Kruzich, P.J.; Frascarelli, S.; Crossley, D.A.; Bunzow, J.R.; Ronca-Testoni, S.; et al. 3-Iodothyronamine is an endogenous and rapid-acting derivative of thyroid hormone. Nat. Med. 2004, 10, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Selen Alpergin, E.S.; Bolandnazar, Z.; Sabatini, M.; Rogowski, M.; Chiellini, G.; Zucchi, R.; Assadi-Porter, F.M. Metabolic profiling reveals reprogramming of lipid metabolic pathways in treatment of polycystic ovary syndrome with 3-iodothyronamine. Physiol. Rep. 2017, 5, e13097. [Google Scholar] [CrossRef] [Green Version]
- Haviland, J.A.; Reiland, H.; Butz, D.E.; Tonelli, M.; Porter, W.P.; Zucchi, R.; Scanlan, T.S.; Chiellini, G.; Assadi-Porter, F.M. NMR-based metabolomics and breath studies show lipid and protein catabolism during low dose chronic T(1)AM treatment. Obesity 2013, 21, 2538–2544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Xu, L.; Yu, C.; Miao, M.; Li, Y. Association between thyroid function and nonalcoholic fatty liver disease in euthyroid elderly Chinese. Clin. Endocrinol. 2011, 75, 240–246. [Google Scholar] [CrossRef]
- Wang, B.; Wang, B.; Yang, Y.; Xu, J.; Hong, M.; Xia, M.; Li, X.; Gao, X. Thyroid function and non-alcoholic fatty liver disease in hyperthyroidism patients. BMC Endocr. Disord. 2021, 21, 27. [Google Scholar] [CrossRef] [PubMed]
- Liangpunsakul, S.; Chalasani, N. Is hypothyroidism a risk factor for non-alcoholic steatohepatitis? J. Clin. Gastroenterol. 2003, 37, 340–343. [Google Scholar] [CrossRef]
- Mazo, D.F.; Lima, V.M.; Stefano, J.T.; Rabelo, F.; Faintuch, J.; Oliveira, C.P. Gluco-lipidic indices in treated hypothyroidism associated with nonalcoholic fatty liver disease. Arq. Gastroenterol. 2011, 48, 186–189. [Google Scholar] [CrossRef] [Green Version]
- Krause, C.; Grohs, M.; El Gammal, A.T.; Wolter, S.; Lehnert, H.; Mann, O.; Mittag, J.; Kirchner, H. Reduced expression of thyroid hormone receptor β in human nonalcoholic steatohepatitis. Endocr. Connect. 2018, 7, 1448–1456. [Google Scholar] [CrossRef] [Green Version]
- Lai, S.; Li, J.; Wang, Z.; Wang, W.; Guan, H. Sensitivity to Thyroid Hormone Indices Are Closely Associated with NAFLD. Front. Endocrinol. 2021, 12, 766419. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yu, Y.; Zhao, M.; Zheng, D.; Zhang, X.; Guan, Q.; Xu, C.; Gao, L.; Zhao, J.; Zhang, H. Benefits of Levothyroxine Replacement Therapy on Nonalcoholic Fatty Liver Disease in Subclinical Hypothyroidism Patients. Int. J. Endocrinol. 2017, 2017, 5753039. [Google Scholar] [CrossRef] [PubMed]
- Bruinstroop, E.; Dalan, R.; Cao, Y.; Bee, Y.M.; Chandran, K.; Cho, L.W.; Soh, S.B.; Teo, E.K.; Toh, S.-A.; Leow, M.K.S.; et al. Low-Dose Levothyroxine Reduces Intrahepatic Lipid Content in Patients with Type 2 Diabetes Mellitus and NAFLD. J. Clin. Endocrinol. Metab. 2018, 103, 2698–2706. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Goossens, N.; Jornayvaz, F.R. Translational Aspects of Diet and Non-Alcoholic Fatty Liver Disease. Nutrients 2017, 9, 1077. [Google Scholar] [CrossRef] [Green Version]
- Schattenberg, J.M.; Ekstedt, M. Assessing the disease burden of non-alcoholic fatty liver disease in the real world—Big data and big numbers. BMC Med. 2019, 17, 123. [Google Scholar] [CrossRef] [Green Version]
- Al Taii, H.; Yaqoob, Z.; Al-Kindi, S.G. Underutilization of Statin Therapy Among Patients with NAFLD in the USA: Validation with Big Data. Dig. Dis. Sci. 2016, 61, 2760. [Google Scholar] [CrossRef] [Green Version]
- Roldan-Valadez, E.; Favila, R.; Martínez-López, M.; Uribe, M.; Ríos, C.; Méndez-Sánchez, N. In vivo 3T spectroscopic quantification of liver fat content in nonalcoholic fatty liver disease: Correlation with biochemical method and morphometry. J Hepatol. 2010, 53, 732–737. [Google Scholar] [CrossRef]
- Roldan-Valadez, E.; Favila, R.; Martínez-López, M.; Uribe, M.; Méndez-Sánchez, N. Imaging techniques for assessing hepatic fat content in nonalcoholic fatty liver disease. Ann. Hepatol. 2008, 7, 212–220. [Google Scholar] [CrossRef]
- Wong, V.W.; Adams, L.A.; de Lédinghen, V.; Wong, G.L.; Sookoian, S. Noninvasive biomarkers in NAFLD and NASH—Current progress and future promise. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 461–478. [Google Scholar] [CrossRef]


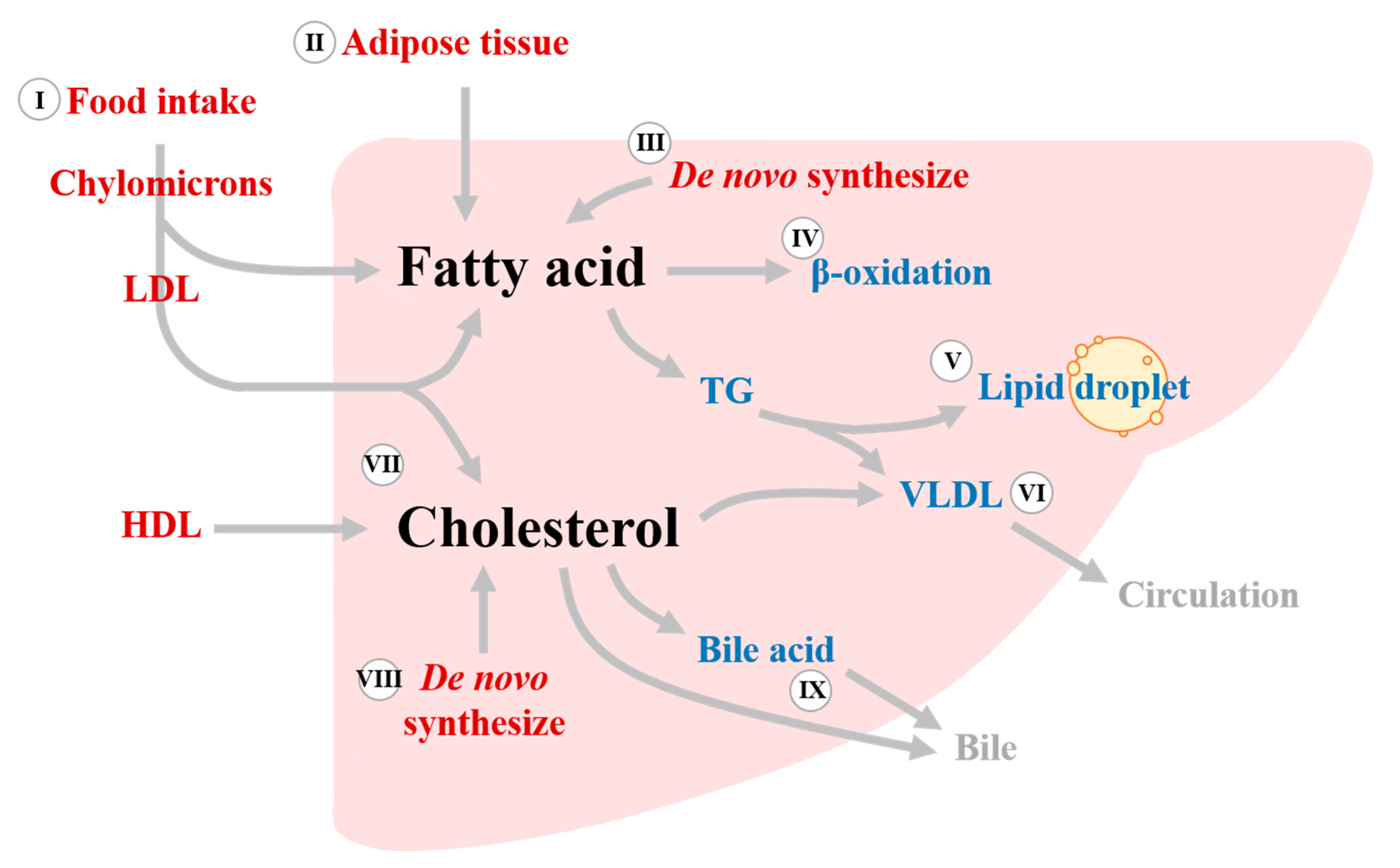{kind=link}
| Compound | Type | Favorable Effects | Unfavorable Effects | Clinical Trials |
|---|---|---|---|---|
| GC-1 (Sobetirome) | TRβ- and liver- selective thyromimetic | 10-fold lower TRα affinity [86] ↓ triglyceride [87,88] ↓ cholesterol [87] Prevent hepatic steatosis in rodent model [26,88,89] ↑ β-oxidation [26] | Fasting hyperglycemia and hyper- insulinemia [89] | |
| GC-24 | TRβ-selective thyromimetic | 40-fold higher TRβ affinity than TRα [90] ↑ energy expenditure [91] Eliminate the increase in adiposity [91] ↑ insulin sensitivity [91] ↓ triglyceride [91] | No improve in hepatic cholesterol [91] Less potent on liver target [92] | |
| KB141 | TRβ-selective thyromimetics | 14-fold higher affinity to TRβ than TRα [93] ↑ metabolic rate [93] ↓ cholesterol [93] ↓ lipoprotein [93] ↓ body weight [93] | ↑ heart rate [93] No liver- selective [93,94] | |
| KB-2115 (Eprotirome) | TRβ- and liver- selective thyromimetic | ↓ triglyceride [88] Ameliorate hepatic steatosis [88,89] ↓ LDL cholesterol [95] ↑ bile acid synthesis [95] | Fasting hyperinsulinemia [89] Liver injury and cartilage damage [96] | NCT00593047 (phase 2) [97] NCT00776321 (phase 2) NCT00677248 (phase 2) NCT01410383 (phase 3) [96] |
| MB07811 (VK2809) | TRβ- and liver-selective prodrug | ↓ cholesterol [98] ↓ triglyceride [82,98] ↓ hepatic steatosis [82] ↓ free fatty acid [82] ↑ β-oxidation [82] | NCT02927184 (phase 2) NCT04173065 (phase 2) | |
| MGL-3196 (Resmetirom) | TRβ- and liver- selective thyromimetic | 28-fold higher affinity to TRβ than TRα [99] ↓ cholesterol [99,100] ↓ triglyceride [99,100] ↓ lipoprotein [100] ↓ fibrosis [100] | Mild diarrhea and nausea [100] | NCT03900429 (phase 3) [101] NCT04643795 (phase 1) NCT04951219 (phase 3) NCT04197479 (phase 3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, C.-J.; Huang, P.-S.; Chien, H.-T.; Lin, T.-K.; Yeh, C.-T.; Lin, K.-H. Effects of Thyroid Hormones on Lipid Metabolism Pathologies in Non-Alcoholic Fatty Liver Disease. Biomedicines 2022, 10, 1232. https://doi.org/10.3390/biomedicines10061232
Liao C-J, Huang P-S, Chien H-T, Lin T-K, Yeh C-T, Lin K-H. Effects of Thyroid Hormones on Lipid Metabolism Pathologies in Non-Alcoholic Fatty Liver Disease. Biomedicines. 2022; 10(6):1232. https://doi.org/10.3390/biomedicines10061232
Chicago/Turabian StyleLiao, Chia-Jung, Po-Shuan Huang, Hui-Tzu Chien, Tzu-Kang Lin, Chau-Ting Yeh, and Kwang-Huei Lin. 2022. "Effects of Thyroid Hormones on Lipid Metabolism Pathologies in Non-Alcoholic Fatty Liver Disease" Biomedicines 10, no. 6: 1232. https://doi.org/10.3390/biomedicines10061232