Towards Modelling Future Trends of Quebec’s Boreal Birds’ Species Distribution under Climate Change



Abstract
1. Introduction
2. Materials and Methods
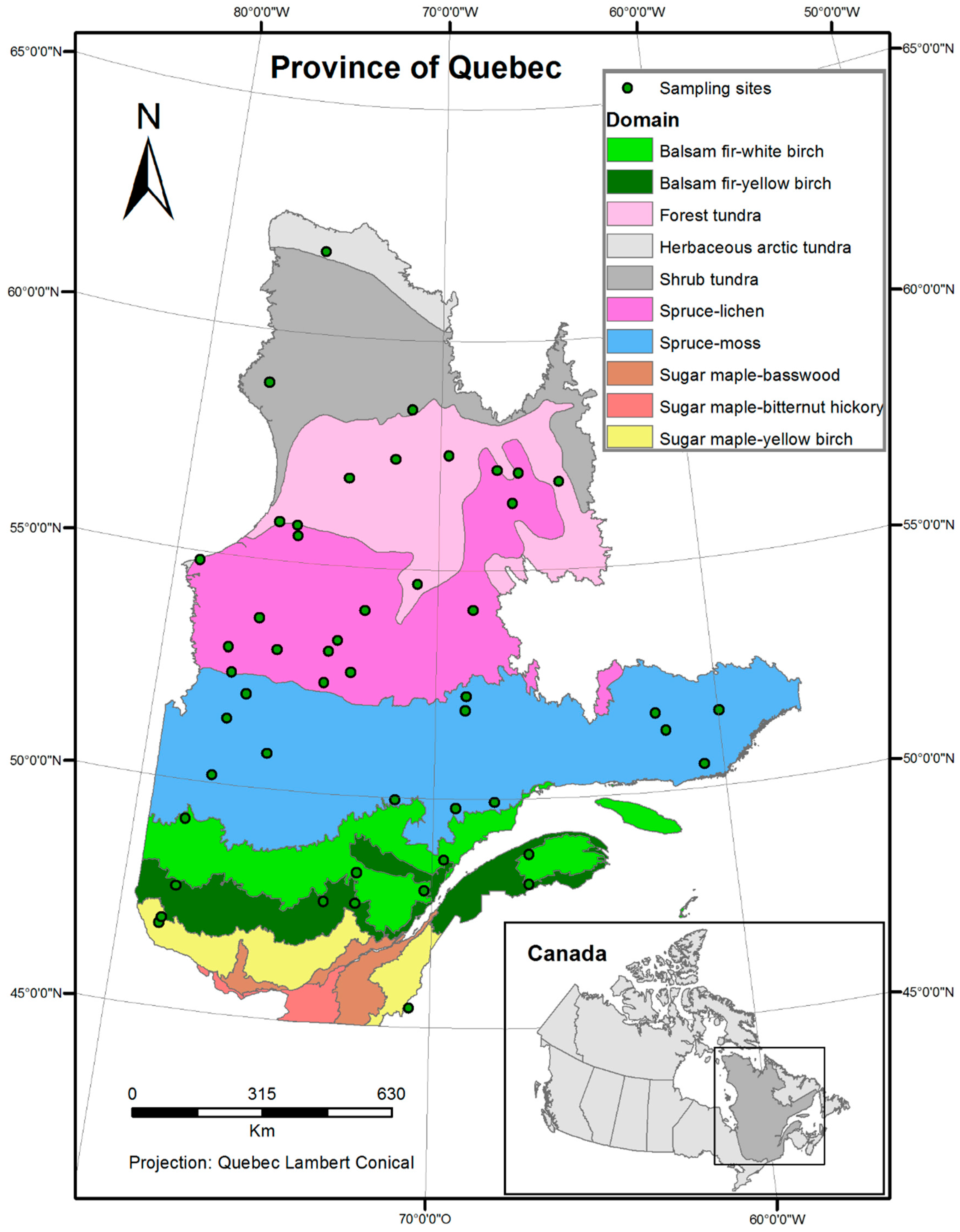
2.1. Study Area, Species Abundance, and Predictors Data
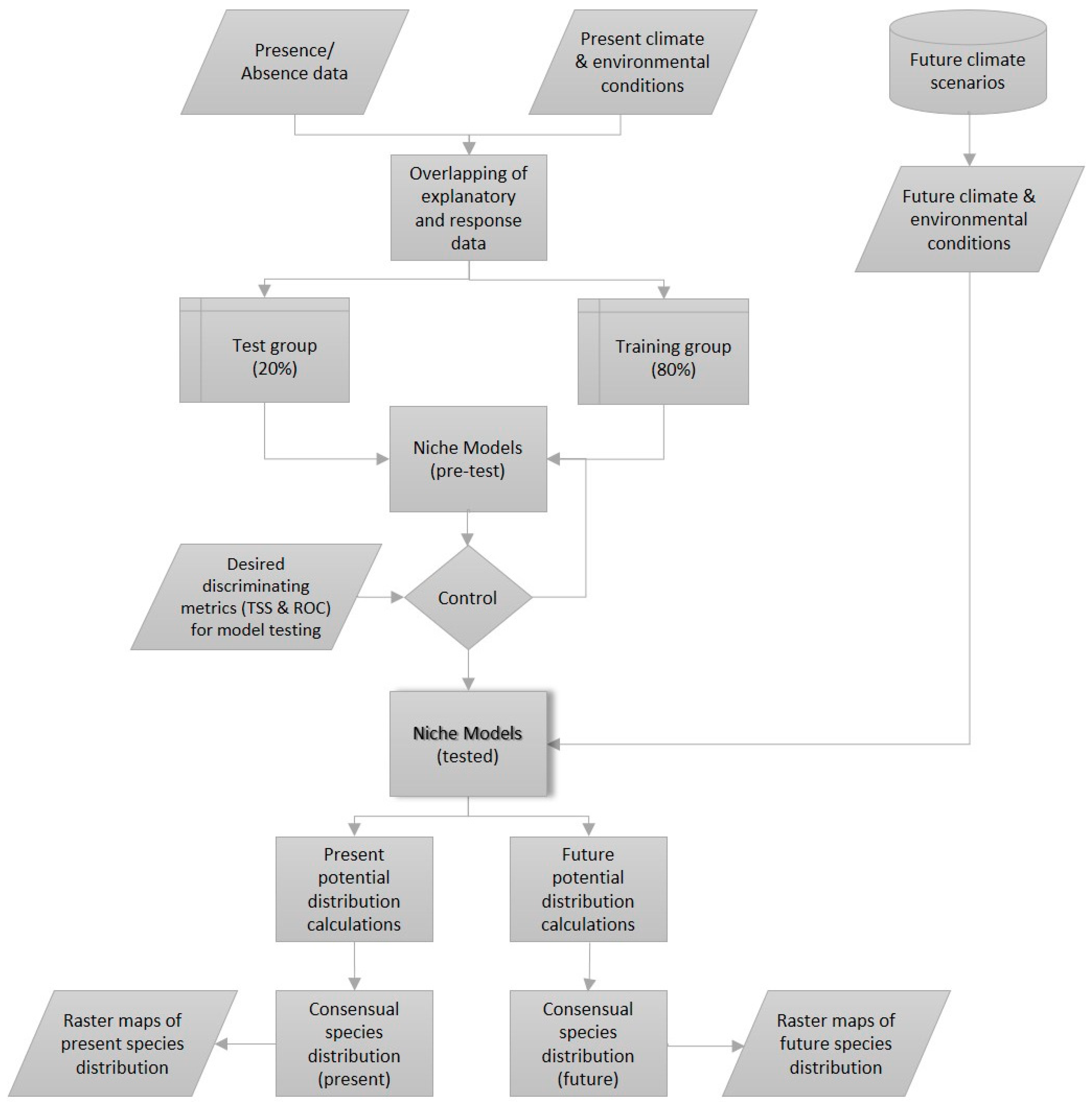
2.2. Methods
2.2.1. Statistical Analyses
2.2.2. Bioclimatic-Based Species Distribution Models
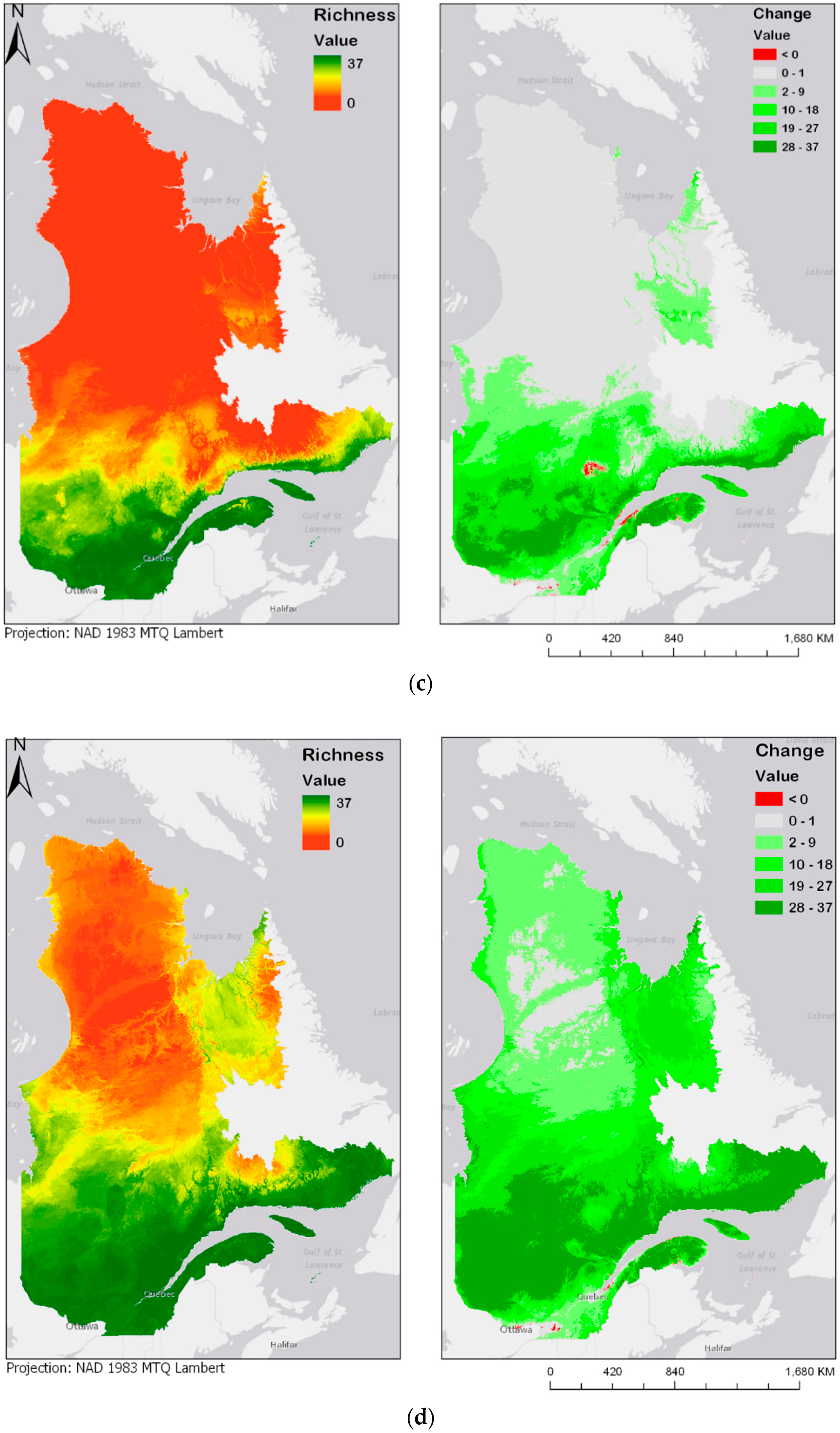
3. Results
3.1. Variable Selection
3.2. Variation Partitioning
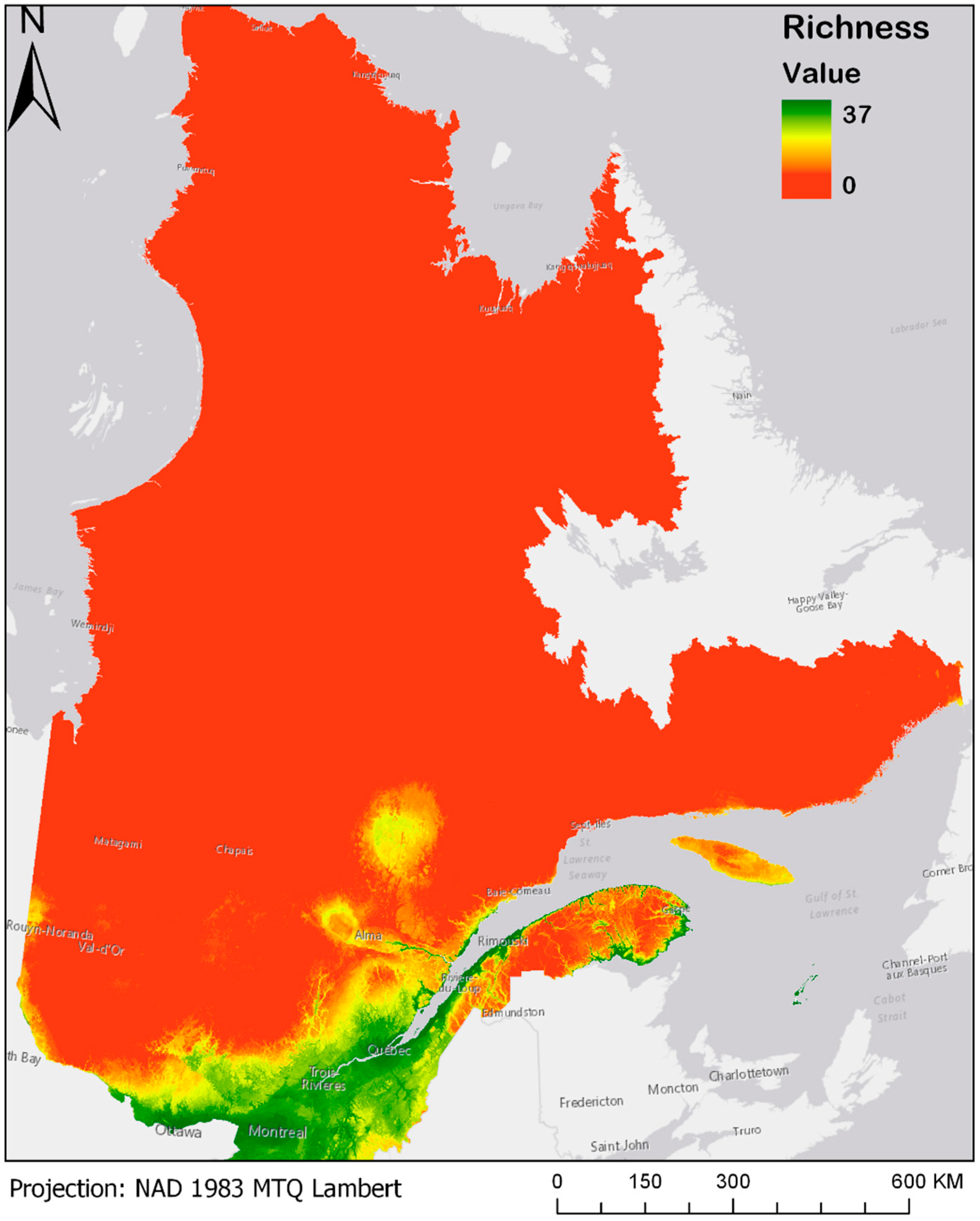
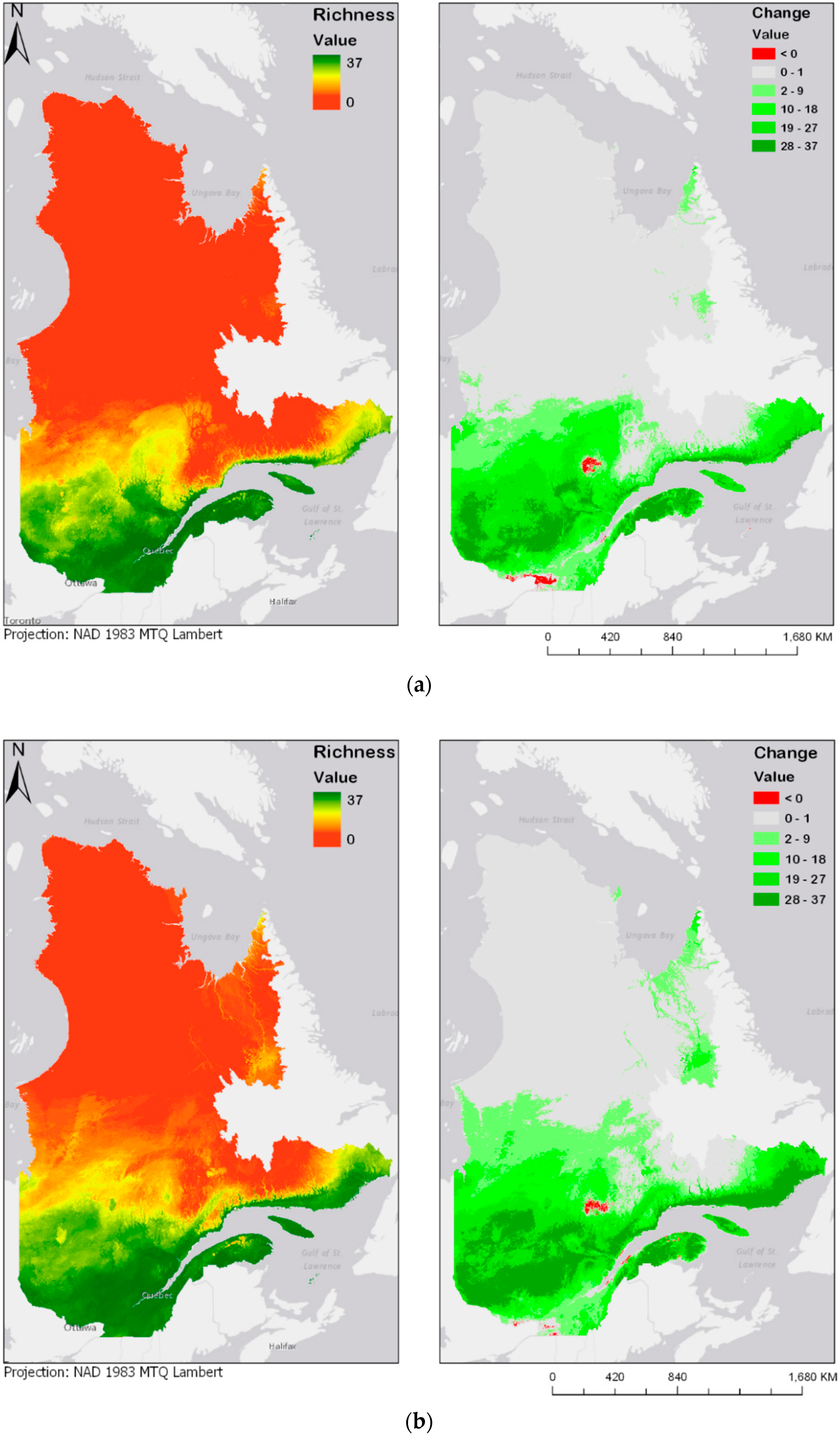
3.3. Bioclimatic Modelling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | English Name |
| Setophaga ruticilla | American Redstard |
| Setophaga castanea | Bay-breasted Warbler |
| Poecile atricapillus | Black-capped Chickadee |
| Poecile hudsonicus | Boreal Chickadee |
| Melanitta americana | Black Scoter |
| Mniotilta varia | Black-and-white Warbler |
| Corvus corax | Common Raven |
| Acanthis flammea | Common Redpoll |
| Junco hyemalis | Dark-eyed Junco |
| Hesperiphona vespertina | Evening Grosbeak |
| Passerella iliaca | Fox Sparrow |
| Regulus satrapa | Golden-crowned Kinglet |
| Perisoreus canadensis | Gray Jay |
| Melospiza lincolnii | Lincoln’s Sparrow |
| Lanius excubitor | Northern Shrike |
| Contopus cooperi | Olive-sided Flycatcher |
| Haemorhous purpureus | Purple Finch |
| Pinicola enucleator | Pine Grosbeak |
| Setophaga pinus | Pine Warbler |
| Euphagus carolinus | Rusty Blackbird |
| Sitta canadensis | Red-breasted Nuthatch |
| Regulus calendula | Ruby-crowned Kinglet |
| Loxia curvirostra | Red Crossbill |
| Vireo olivaceus | Red-eyed Vireo |
| Bonasa umbellus | Ruffed Grouse |
| Falcipennis canadensis | Spruce Grouse |
| Tringa solitaria | Solitary Sandpiper |
| Actitis macularius | Spotted Sandpiper |
| Melanitta perspicillata | Surf Scoter |
| Melospiza georgiana | Swamp Sparrow |
| Catharus ustulatus | Swainson’s Thrush |
| Catharus fuscescens | Veery |
| Zonotrichia leucophrys | White-crowned Sparrow |
| Numenius phaeopus | Whimbrel |
| Loxia leucoptera | White-winged Crossbill |
| Melanitta deglandi | White-winged Scoter |
| Empidonax flaviventris | Yellow-bellied Flycatcher |
References
- Virkkala, R.; Heikkinen, R.K.; Leikola, N.; Luoto, M. Projected large-scale range reductions of northern-boreal land bird species due to climate change. Biol. Conserv. 2008, 141, 1343–1353. [Google Scholar] [CrossRef]
- Lawler, J.J.; Shafer, S.L.; White, D.; Kareiva, P.; Maurer, E.P.; Blaustein, A.R.; Bartlein, P.J.; Ecology, S.; Mar, N.; Lawler, J.; et al. Projected Climate-Induced Faunal Change in the Western Hemisphere. Ecology 2009, 90, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C. Ecological and Evolutionary Responses to Recent Climate Change. Annu. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Summary for Policymakers. In Climate Change 2013: The Physical Science Basis; Stocker, T.F., Qin, D.G.-K., Plattner, M., Tignor, S.K., Allen, J., Boschung, A., Nauels, Y., Xia, V.B., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; p. 33. ISBN 110705799X. [Google Scholar]
- Walther, G.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Forister, M.L.; McCall, A.C.; Sanders, N.J.; Fordyce, J.A.; Thorne, J.H.; O’Brien, J.; Waetjen, D.P.; Shapiro, A.M. Compounded effects of climate change and habitat alteration shift patterns of butterfly diversity. Proc. Natl. Acad. Sci. USA 2010, 107, 2088–2092. [Google Scholar] [CrossRef] [PubMed]
- Hurlbert, A.H.; Liang, Z. Spatiotemporal Variation in Avian Migration Phenology: Citizen Science Reveals Effects of Climate Change. PLoS ONE 2012, 7, e31662. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.J. The disproportionate effect of global warming on the arrival dates of short-distance migratory birds in North America. Ibis 2003, 145, 484–495. [Google Scholar] [CrossRef]
- Swanson, D.L.; Palmer, J.S. Spring migration phenology of birds in the Northern Prairie region is correlated with local climate change. J. Field Ornithol. 2009, 80, 351–363. [Google Scholar] [CrossRef]
- Virkkala, R.; Marmion, M.; Heikkinen, R.K.; Thuiller, W.; Luoto, M. Predicting range shifts of northern bird species: Influence of modelling technique and topography. Acta Oecol. 2010, 36, 269–281. [Google Scholar] [CrossRef]
- Thompson, S.D.; Nelson, T.A.; Coops, N.C.; Wulder, M.A.; Lantz, T.C. Global Spatial–Temporal Variability in Terrestrial Productivity and Phenology Regimes between 2000 and 2012. Ann. Am. Assoc. Geogr. 2017, 4452, 1–19. [Google Scholar] [CrossRef]
- Perez, L.; Nelson, T.; Coops, N.C.; Fontana, F.; Drever, C.R. Characterization of spatial relationships between three remotely sensed indirect indicators of biodiversity and climate: A 21 years’ data series review across the Canadian boreal forest. Int. J. Digit. Earth 2016, 9, 676–696. [Google Scholar] [CrossRef]
- Nelson, T.; Coops, N.; Wulder, M.; Perez, L.; Fitterer, J.; Powers, R.; Fontana, F. Predicting Climate Change Impacts to the Canadian Boreal Forest. Diversity 2014, 6, 133–157. [Google Scholar] [CrossRef]
- Stralberg, D.; Bayne, E.M.; Cumming, S.G.; Solymos, P.; Song, S.J.; Schmiegelow, F.K.A. Conservation of future boreal forest bird communities considering lags in vegetation response to climate change: A modified refugia approach. Divers. Distrib. 2015, 21, 1112–1128. [Google Scholar] [CrossRef]
- Pothier, D. Portrait de la forêt boréale québécoise. Le Nat. Can. 2001, 125, 5–9. [Google Scholar]
- MERN. Vegetation Zones and Bioclimatic Domains in Québec. Available online: http://mern.gouv.qc.ca/english/publications/forest/publications/zone-a.pdf (accessed on 1 April 2017).
- MERN. Bref Portrait de La Forêt Boréale Au Québec. Available online: http://www.mern.gouv.qc.ca/presse/feux-grands.jsp (accessed on 20 July 2005).
- ESRI ArcGIS Desktop, version 10.1; ESRI: Redlands, CA, USA, 2014.
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2014. [Google Scholar]
- eBird: An online Database of Bird Distribution and Abundance [Web Application]. Available online: http://www.ebird.org (accessed on 20 May 2002).
- The Boreal Songbird Initiative Borel Birds. Available online: http://www.borealbirds.org/comprehensive-boreal-bird-guide (accessed on 4 May 2016).
- Royston, J.P. An Extension of Shapiro and Wilk’s W Test for Normality to Large Samples. Appl. Stat. 1982, 31, 115–124. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Natural Resources Canada Canadian Digital Elevation Model. Available online: http://open.canada.ca/data/en/dataset/7f245e4d-76c2-4caa-951a-45d1d2051333 (accessed on 4 May 2016).
- Global Forest Watch Canada Cumulative Anthropogenic Access, Eastern Canada. Available online: http://www.globalforestwatch.ca/data/20100913B (accessed on 4 May 2016).
- Conservation Biology Institute Canada’s Forest Inventory (CanFI) 2001 for the Boreal Eco-Region—Percent Forested, Quebec. Available online: https://databasin.org/datasets/00b4cf27796d4a32be21df7716103e3f (accessed on 20 May 2002).
- Legendre, P.; Legendre, L.F. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlnads, 2012. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. In R Package Version; CRAN-R: Oulu, Finland, 2017. [Google Scholar]
- Liu, Q. Variation Partioning by Partial Redundancy Analysis (RDA). Environmetrics 1997, 8, 75–85. [Google Scholar] [CrossRef]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation Partitioning of Species Data Matrices: Estimation and Comparison of Fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Legendre, P. Studying beta diversity: Ecological variation partitioning by multiple regression and canonical analysis. J. Plant Ecol. 2008, 1, 3–8. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Zweig, M.H.; Campbell, G. Receiver-operating characteristic (ROC) plots: A fundamental evaluation tool in clinical medicine. Clin. Chem. 1993, 39, 561–577. [Google Scholar] [PubMed]
- Fieldgin, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Pearson, R.G.; Thuiller, W.; Araújo, M.B.; Martinez-Meyer, E.; Brotons, L.; McClean, C.; Miles, L.; Segurado, P.; Dawson, T.P.; Lees, D.C. Model-based uncertainty in species range prediction. J. Biogeogr. 2006, 33, 1704–1711. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R. Biomod2: Ensemble Platform for Species Distribution Modeling; CRAN-R: Grenoble, France, 2014. [Google Scholar]
- Hirschi, J.J.M.; Sinha, B.; Josey, S.A. Global warming and changes of continentality since 1948. Weather 2007, 62, 215–221. [Google Scholar] [CrossRef]
- Fitterer, J.L.; Nelson, T.A.; Coops, N.C.; Wulder, M.A.; Mahony, N.A. Exploring the ecological processes driving geographical patterns of breeding bird richness in British Columbia, Canada. Ecol. Appl. 2013, 23, 888–903. [Google Scholar] [CrossRef] [PubMed]
- Renwick, A.R.; Massimino, D.; Newson, S.E.; Chamberlain, D.E.; Pearce-Higgins, J.W.; Johnston, A. Modelling changes in species’ abundance in response to projected climate change. Divers. Distrib. 2012, 18, 121–132. [Google Scholar] [CrossRef]
- Moudrý, V.; Šímová, P. Relative importance of climate, topography, and habitats for breeding wetland birds with different latitudinal distributions in the Czech Republic. Appl. Geogr. 2013, 44, 165–171. [Google Scholar] [CrossRef]
- Revermann, R.; Schmid, H.; Zbinden, N.; Spaar, R.; Schröder, B. Habitat at the mountain tops: How long can Rock Ptarmigan (Lagopus muta helvetica) survive rapid climate change in the Swiss Alps? A multi-scale approach. J. Ornithol. 2012, 153, 891–905. [Google Scholar] [CrossRef]
- Berteaux, D.; de Blois, S.; Angers, J.F.; Bonin, J.; Casajus, N.; Darveau, M.; Fournier, F.; Humphries, M.M.; McGill, B.; Larivée, J.; et al. The CC-Bio project: Studying the effects of climate change on Quebec biodiversity. Diversity 2010, 2, 1181–1204. [Google Scholar] [CrossRef]





| Variable Name | Description | |
|---|---|---|
| Bioclimatic Variables | BIO1 | Annual Mean Temperature |
| BIO2 | Mean Diurnal Range (Mean of monthly (max temp–min temp)) | |
| BIO3 | Isothermality (BIO2/BIO7) (×100) | |
| BIO4 | Temperature Seasonality (standard deviation ×100) | |
| BIO5 | Max Temperature of Warmest Month | |
| BIO6 | Min Temperature of Coldest Month | |
| BIO7 | Temperature Annual Range (BIO5-BIO6) | |
| BIO8 | Mean Temperature of Wettest Quarter | |
| BIO9 | Mean Temperature of Driest Quarter | |
| BIO10 | Mean Temperature of Warmest Quarter | |
| BIO11 | Mean Temperature of Coldest Quarter | |
| BIO12 | Annual Precipitation | |
| BIO13 | Precipitation of Wettest Month | |
| BIO14 | Precipitation of Driest Month | |
| BIO15 | Precipitation Seasonality (Coefficient of Variation) | |
| BIO16 | Precipitation of Wettest Quarter | |
| BIO17 | Precipitation of Driest Quarter | |
| BIO18 | Precipitation of Warmest Quarter | |
| BIO19 | Precipitation of Coldest Quarter | |
| Other Variables | elevation | Elevation in meter |
| pct_for | Percentage of forested area | |
| pct_wet | Percentage of wet area | |
| linear | Linear disturbances | |
| Acronym | Name | Description | Source |
|---|---|---|---|
| RF | Random Forest | Measures variable importance using multiple permutations. | Breiman, 2001 |
| MARS | Multivariate Adaptive Regression Splines | Allows to generate non-linear models comprising interaction. | Friedman, 1991 |
| MAXENT | Maximum Entropy | Allows presence-only modelling. Automated learning approach, or machine-learning. | Phillips et al., 2004 |
| Name | Latitude | Longitude | Bioclimatic Domain | Name | Latitude | Longitude | Bioclimatic Domain |
|---|---|---|---|---|---|---|---|
| row0 | 55.8447 | −76.2316 | Forest tundra | row25 | 51.9439 | −68.9568 | Spruce-moss |
| row1 | 48.6628 | −69.6323 | Fir-white birch | row26 | 55.5781 | −75.4543 | Spruce-lichen |
| row2 | 46.8135 | −78.6928 | Maple-yellow birch | row27 | 56.4672 | −67.1192 | Spruce-lichen |
| row3 | 51.6501 | −59.9633 | Spruce-moss | row28 | 51.3266 | −61.8963 | Spruce-moss |
| row4 | 47.6502 | −73.5156 | Fir-Yellow birch | row29 | 52.6880 | −73.1066 | Spruce-lichen |
| row5 | 49.9449 | −71.3324 | Spruce-moss | row30 | 57.1851 | −67.7253 | Spruce-lichen |
| row6 | 53.0530 | −75.8372 | Spruce-lichen | row31 | 56.9220 | −65.2494 | Forest tundra |
| row7 | 52.4532 | −77.3685 | Spruce-lichen | row32 | 51.4333 | −77.3326 | Spruce-moss |
| row8 | 50.1640 | −77.6107 | Spruce-moss | row33 | 57.1362 | −66.8704 | Spruce-lichen |
| row9 | 55.8104 | −75.5142 | Spruce-lichen | row34 | 58.8476 | −77.3046 | Shrub tundra |
| row10 | 47.9788 | −70.2513 | Fir-white birch | row35 | 51.7193 | −62.2129 | Spruce-moss |
| row11 | 49.1433 | −78.3106 | Fir-white birch | row36 | 54.1290 | −68.6697 | Spruce-lichen |
| row12 | 54.6786 | −70.7768 | Forest tundra | row37 | 48.3239 | −72.4839 | Fir-white birch |
| row13 | 57.3887 | −71.8150 | Forest tundra | row38 | 52.9978 | −77.6026 | Spruce-lichen |
| row14 | 47.6650 | −78.3112 | Fir-Yellow birch | row39 | 52.2419 | −68.9173 | Spruce-moss |
| row15 | 50.5063 | −60.6872 | Spruce-moss | row40 | 53.3667 | −73.6408 | Spruce-lichen |
| row16 | 45.4110 | −70.6361 | Maple-yellow birch | row41 | 49.8007 | −69.2638 | Spruce-moss |
| row17 | 50.7644 | −75.8232 | Spruce-moss | row42 | 54.0512 | −72.7190 | Spruce-lichen |
| row18 | 52.0158 | −76.7739 | Spruce-moss | row43 | 48.1273 | −66.8305 | Fir-white birch |
| row19 | 49.9230 | −67.9408 | Spruce-moss | row44 | 61.7936 | −75.4240 | Herbaceous arctic tundra |
| row20 | 54.8004 | −79.0797 | Spruce-lichen | row45 | 53.1172 | −73.9403 | Spruce-lichen |
| row21 | 58.4981 | −71.2060 | Arctic tundra | row46 | 46.9410 | −78.6283 | Maple-yellow birch |
| row22 | 48.7750 | −66.8054 | Fir-white birch | row47 | 57.5095 | −69.6575 | Forest tundra |
| row23 | 53.7060 | −76.5934 | Spruce-lichen | row48 | 56.9296 | −73.6560 | Forest tundra |
| row24 | 52.4247 | −74.0392 | Spruce-lichen | row49 | 47.6405 | −72.4772 | Fir-Yellow birch |
| Variables | Df | Var | F | N.perm | Pr (>F) | VIF |
|---|---|---|---|---|---|---|
| elevation | 1 | 49.512 | 23.8522 | 99 | 0.01 | 3.359 |
| pct_wet | 1 | 4.443 | 2.1403 | 99 | 0.07 | 1.232 |
| BIO11 | 1 | 67.141 | 32.3447 | 99 | 0.01 | 9.217 |
| BIO15 | 1 | 10.6 | 5.1063 | 99 | 0.01 | 7.199 |
| BIO16 | 1 | 14.793 | 7.1266 | 99 | 0.01 | 5.181 |
| BIO7 | 1 | 9.725 | 4.685 | 99 | 0.01 | 1.779 |
| residuals | 43 | 89.259 |
| Variables | Df | Var | F | N.perm | Pr (>F) |
|---|---|---|---|---|---|
| RDA1 | 1 | 117.189 | 56.455 | 199 | 0.005 |
| RDA2 | 1 | 18.856 | 9.0835 | 199 | 0.005 |
| RDA3 | 1 | 13.101 | 6.3114 | 199 | 0.005 |
| RDA4 | 1 | 4.093 | 1.972 | 999 | 0.071 |
| RDA5 | 1 | 2.017 | 0.9718 | 99 | 0.44 |
| RDA6 | 1 | 0.958 | 0.4614 | 99 | 0.88 |
| residuals | 43 | 89.259 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaudreau, J.; Perez, L.; Harati, S. Towards Modelling Future Trends of Quebec’s Boreal Birds’ Species Distribution under Climate Change. ISPRS Int. J. Geo-Inf. 2018, 7, 335. https://doi.org/10.3390/ijgi7090335
Gaudreau J, Perez L, Harati S. Towards Modelling Future Trends of Quebec’s Boreal Birds’ Species Distribution under Climate Change. ISPRS International Journal of Geo-Information. 2018; 7(9):335. https://doi.org/10.3390/ijgi7090335
Chicago/Turabian StyleGaudreau, Jonathan, Liliana Perez, and Saeed Harati. 2018. "Towards Modelling Future Trends of Quebec’s Boreal Birds’ Species Distribution under Climate Change" ISPRS International Journal of Geo-Information 7, no. 9: 335. https://doi.org/10.3390/ijgi7090335
APA StyleGaudreau, J., Perez, L., & Harati, S. (2018). Towards Modelling Future Trends of Quebec’s Boreal Birds’ Species Distribution under Climate Change. ISPRS International Journal of Geo-Information, 7(9), 335. https://doi.org/10.3390/ijgi7090335