β-Lactamase Tools for Establishing Cell Internalization and Cytosolic Delivery of Cell Penetrating Peptides

 ,
,
Abstract
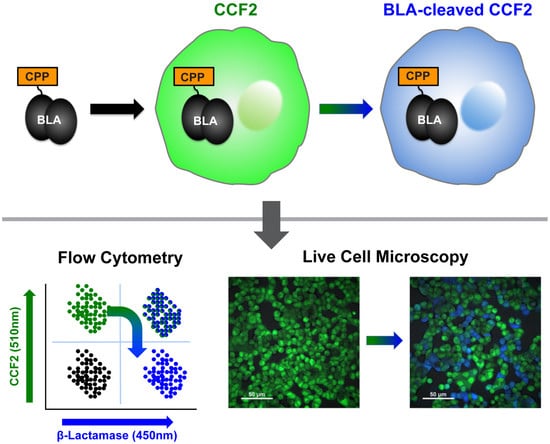
:
1. Introduction
2. Results
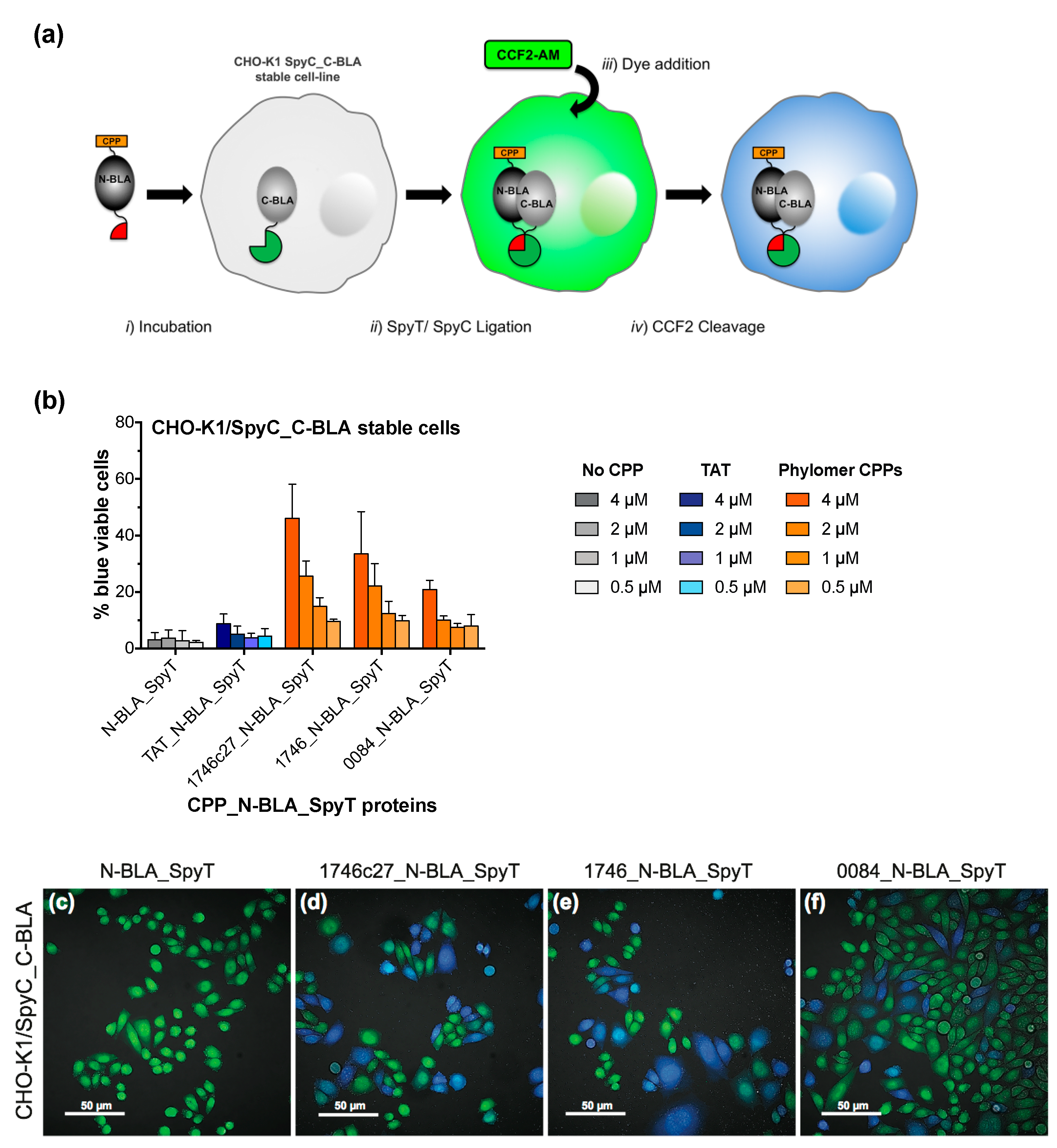
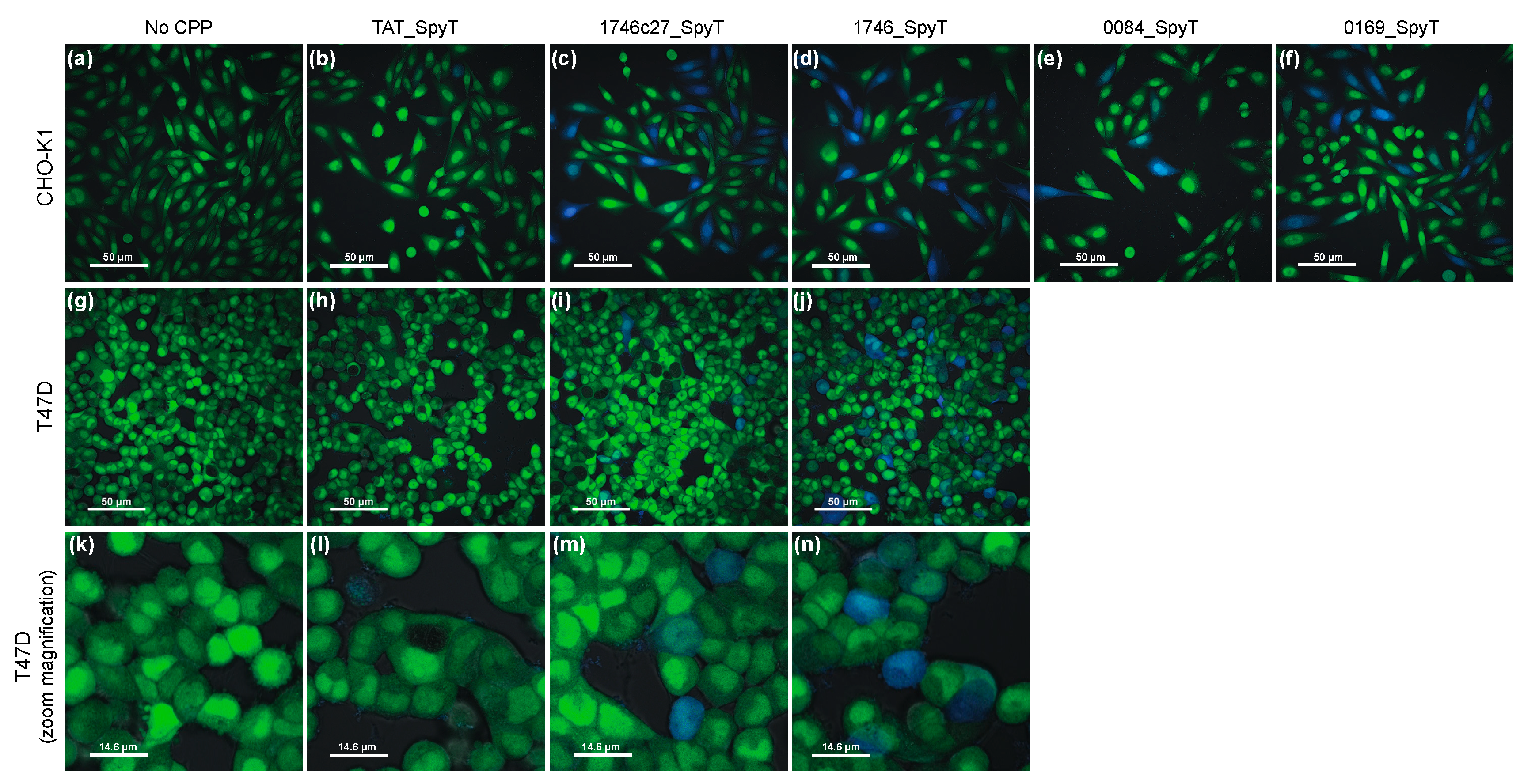
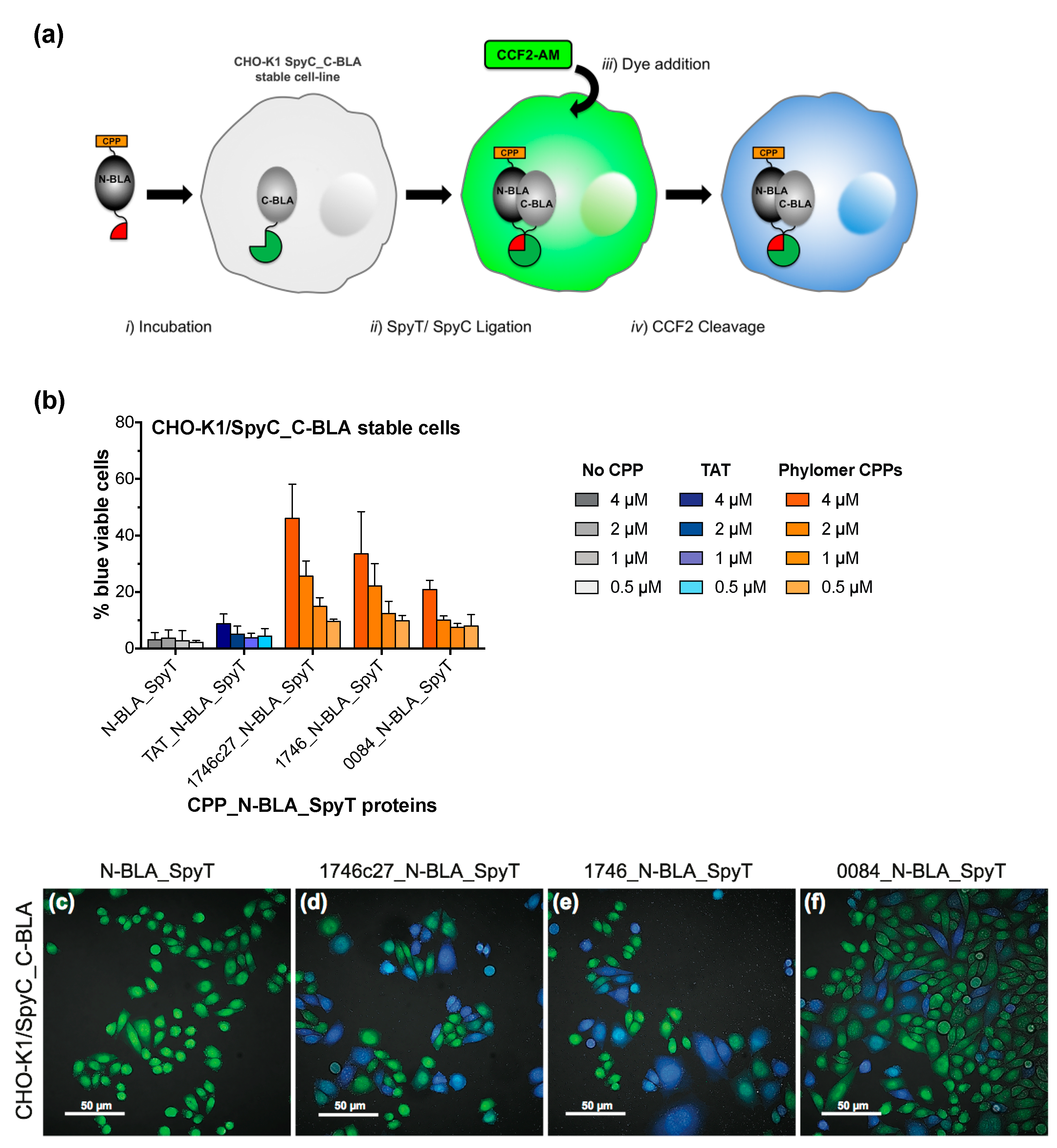
2.1. β-Lactamase Internalisation Assay
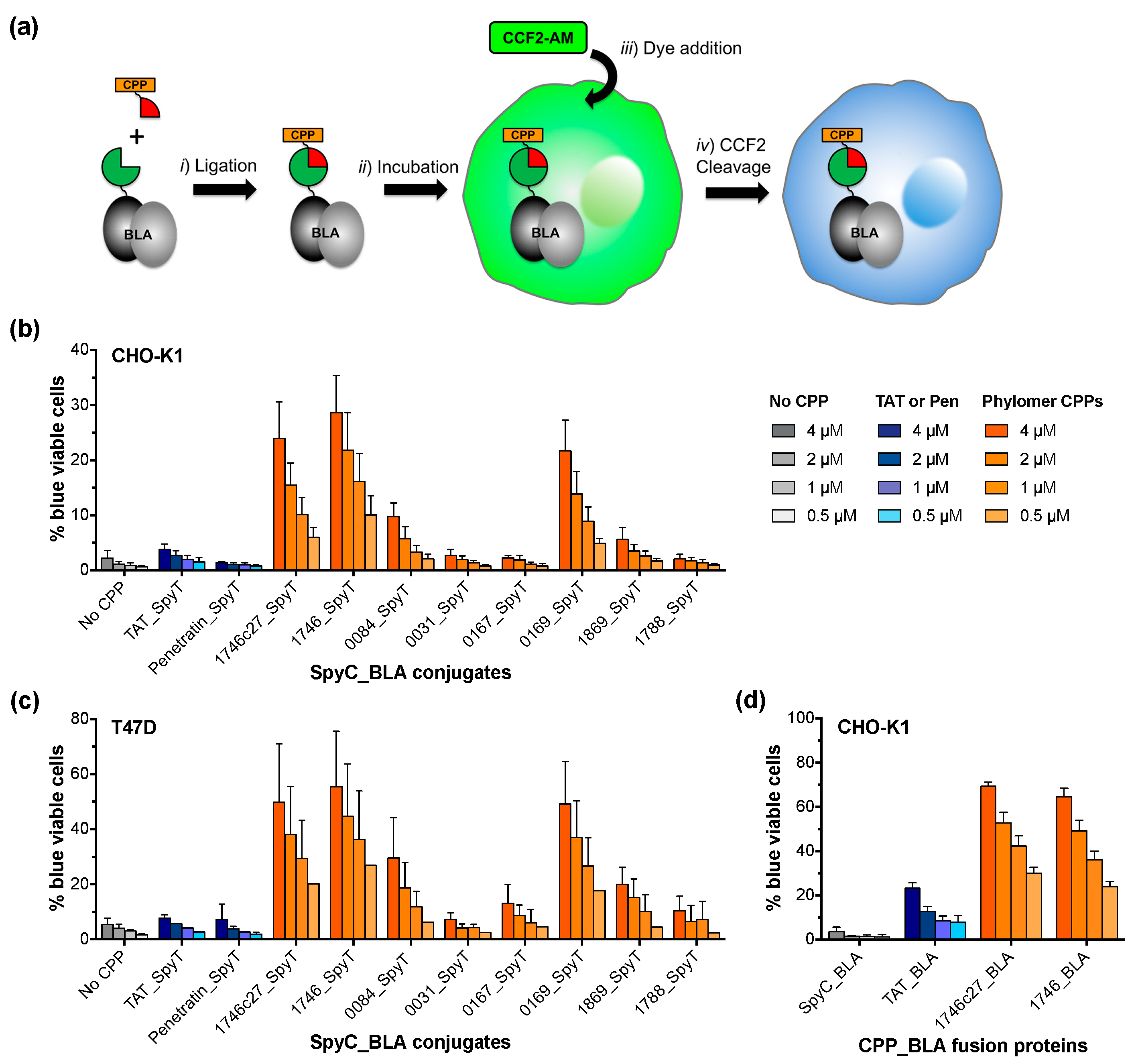
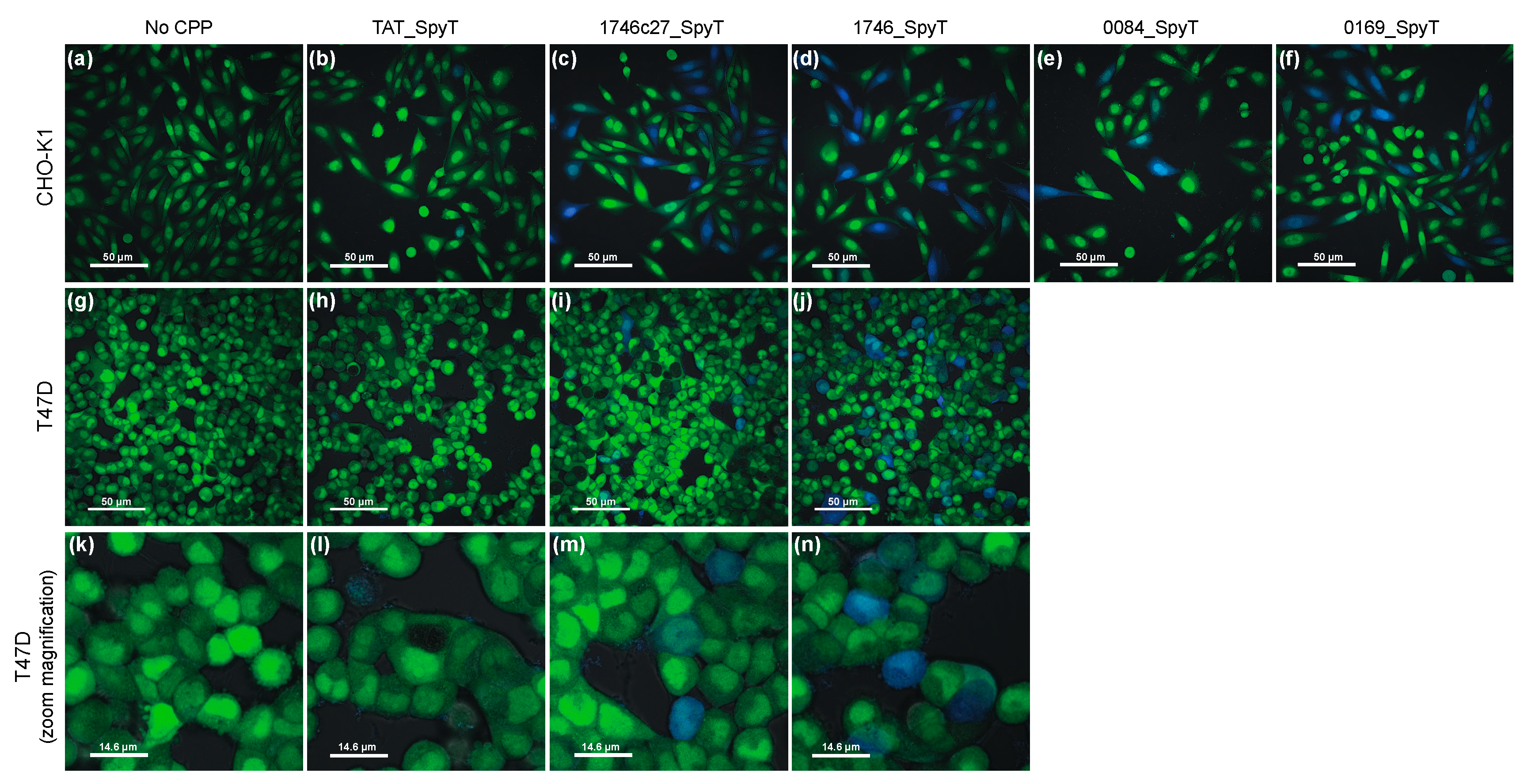
2.2. Split β-Lactamase Cytosolic Delivery Assay
3. Discussion
4. Materials and Methods
4.1. Mammalian Cell Culture
4.2. β-Lactamase Fluorogenic Assay
4.3. Confocal Live-Cell Microscopy
4.4. Peptides
4.5. Recombinant Protein Expression and Purifcation
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence 1 | Length | Net Charge (pH 7) |
|---|---|---|---|
| TAT_SpyT | GRKKRRQRRRGASAHIVMVDAYKPTKG | 27 | 9+ |
| 1746c27_SpyT | KKKKQPPKPKKPKTQEKKKKQPPKPKRGASAHIVMVDAYKPTKG | 44 | 15+ |
| 1746_SpyT | PLKPKKPKTQEKKKKQPPKPKKPKTQEKKKKQPPKPKRGASAHIVMVDAYKPTKG | 55 | 18+ |
| 0031_SpyT | RASPSEQRRKRRRCHRGETQRPDFEAEIEKQQRGASAHIVMVDAYKPTKG | 51 | 6+ |
| 0084_SpyT | RKQKSLQTKLAENPPVPRKKRQSRPRWKQWLQKGASAHIVMVDAYKPTKG | 50 | 12+ |
| 0167_SpyT | PSSTPATNVARPRLNPIRGHKFALAVPNSRTRGASAHIVMVDAYKPTKG | 49 | 7+ |
| 0169_SpyT | PLTQRTLQRGKKPKQRQNWKKARTSSAKTAPKTVVSRTTSQRKGASAHIVMVDAYKPTKG | 60 | 15+ |
| 1788_SpyT | LFVDKATPQIYYTPSESVTVKSKGKNRRKKSGASAHIVMVDAYKPTKG | 48 | 7+ |
| 1869_SpyT | PPHPRPLPAPAQSRKKQKGRAGRGHEKTGASVLRGPQKPHPLPAQLRGASAHIVMVDAYKPTKG | 64 | 11+ |
| Name | Sequence 1 | kDa | pI 2 |
|---|---|---|---|
| SpyC_BLA | MGHHHHHHGATLEVLFQGPGGSGSDSATHIKFSKRDEDGKELAGATMELRDSSGKTISTWISDGQVKDFYLYPGKYTFVETAAPDGYEVATAITFTVNEQGQVTVNGKATKGSHPETLVKVKDAEDQLGARVGYIELDLNSGKILESFRPEERFPMMSTFKVLLCGAVLSRIDAGQEQLGRRIHYSQNDLVEYSPVTEKHLTDGMTVRELCSAAITMSDNTAANLLLTTIGGPKELTAFLHNMGDHVTRLDRWEPELNEAIPNDERDTTMPAAMATTLRKLLTGELLTLASRQQLIDWMEADKVAGPLLRSALPAGWFIADKSGAGERGSRGIIAALGPDGKPSRIVVIYTTGSQATMDERNRQIAEIGASLIKHWQLGSASGTTGATSGEF | 42.4 kDa | 5.6 |
| TAT_BLA | MASGRKKRRQRRRASSGSHHHHHHGATLEVLFQGPGGSHPETLVKVKDAEDQLGARVGYIELDLNSGKILESFRPEERFPMMSTFKVLLCGAVLSRIDAGQEQLGRRIHYSQNDLVEYSPVTEKHLTDGMTVRELCSAAITMSDNTAANLLLTTIGGPKELTAFLHNMGDHVTRLDRWEPELNEAIPNDERDTTTPAAMATTLRKLLTGELLTLASRQQLIDWMEADKVAGPLLRSALPAGWFIADKSGAGERGSRGIIAALGPDGKPSRIVVIYTTGSQATMDERNRQIAEIGASLIKHWQLGSASGTTGATSG | 34.3 kDa | 7.2 |
| 1746c27_BLA | MASKKKKQPPKPKKPKTQEKKKKQPPKPKRASSGSHHHHHHGATLEVLFQGPGGSHPETLVKVKDAEDQLGARVGYIELDLNSGKILESFRPEERFPMMSTFKVLLCGAVLSRIDAGQEQLGRRIHYSQNDLVEYSPVTEKHLTDGMTVRELCSAAITMSDNTAANLLLTTIGGPKELTAFLHNMGDHVTRLDRWEPELNEAIPNDERDTTTPAAMATTLRKLLTGELLTLASRQQLIDWMEADKVAGPLLRSALPAGWFIADKSGAGERGSRGIIAALGPDGKPSRIVVIYTTGSQATMDERNRQIAEIGASLIKHWQLGSASGTTGATSG | 36.1 kDa | 9.2 |
| 1746_BLA | MASPLKPKKPKTQEKKKKQPPKPKKPKTQEKKKKQPPKPKRASSGSHHHHHHGATLEVLFQGPGGSHPETLVKVKDAEDQLGARVGYIELDLNSGKILESFRPEERFPMMSTFKVLLCGAVLSRIDAGQEQLGRRIHYSQNDLVEYSPVTEKHLTDGMTVRELCSAAITMSDNTAANLLLTTIGGPKELTAFLHNMGDHVTRLDRWEPELNEAIPNDERDTTTPAAMATTLRKLLTGELLTLASRQQLIDWMEADKVAGPLLRSALPAGWFIADKSGAGERGSRGIIAALGPDGKPSRIVVIYTTGSQATMDERNRQIAEIGASLIKHWQLGSASGTTGATSG | 37.4 kDa | 9.4 |
| N-BLA_SpyT | MGHHHHHHGATLEVLFQGPGGSQLHPETLVKVKDAEDQLGARVGYIELDLNSGKILESFRPEERFPMMSTFKVLLCGAVLSRIDAGQEQLGRRIHYSQNDLVEYSPVTEKHLTDGMTVRELCSAAITMSDNTAANLLLTTIGGPKELTAFLHNMGDHVTRLDRWEPELNEAIPNDERDTTMPAAMATTLRKLLTGGGSASGGSGTGATSGAHIVMVDAYKPTKGVDG | 24.5 kDa | 5.7 |
| TAT_N-BLA_SpyT | MASGRKKRRQRRRASSGSHHHHHHGTGAGSGHPETLVKVKDAEDQLGARVGYIELDLNSGKILESFRPEERFPMMSTFKVLLCGAVLSRIDAGQEQLGRRIHYSQNDLVEYSPVTEKHLTDGMTVRELCSAAITMSDNTAANLLLTTIGGPKELTAFLHNMGDHVTRLDRWEPELNEAIPNDERDTTTPAAMATTLRKLLTGGGSASGGSGTGATSGAHIVMVDAYKPTKGGTGGS | 25.3 kDa | 7.9 |
| 1746c27_N-BLA_SpyT | MASKKKKQPPKPKKPKTQEKKKKQPPKPKRASSGSHHHHHHGTGAGSGHPETLVKVKDAEDQLGARVGYIELDLNSGKILESFRPEERFPMMSTFKVLLCGAVLSRIDAGQEQLGRRIHYSQNDLVEYSPVTEKHLTDGMTVRELCSAAITMSDNTAANLLLTTIGGPKELTAFLHNMGDHVTRLDRWEPELNEAIPNDERDTTTPAAMATTLRKLLTGGGSASGGSGTGATSGAHIVMVDAYKPTKGGTGGS | 27.2 kDa | 9.3 |
| 1746_N-BLA_SpyT | MASPLKPKKPKTQEKKKKQPPKPKKPKTQEKKKKQPPKPKRASSGSHHHHHHGTGAGSGHPETLVKVKDAEDQLGARVGYIELDLNSGKILESFRPEERFPMMSTFKVLLCGAVLSRIDAGQEQLGRRIHYSQNDLVEYSPVTEKHLTDGMTVRELCSAAITMSDNTAANLLLTTIGGPKELTAFLHNMGDHVTRLDRWEPELNEAIPNDERDTTTPAAMATTLRKLLTGGGSASGGSGTGATSGAHIVMVDAYKPTKGGTGGS | 28.5 kDa | 9.5 |
| 0084_N-BLA_SpyT | MASRKQKSLQTKLAENPPVPRKKRQSRPRWKQWLQKASSGSHHHHHHGTGAGSGHPETLVKVKDAEDQLGARVGYIELDLNSGKILESFRPEERFPMMSTFKVLLCGAVLSRIDAGQEQLGRRIHYSQNDLVEYSPVTEKHLTDGMTVRELCSAAITMSDNTAANLLLTTIGGPKELTAFLHNMGDHVTRLDRWEPELNEAIPNDERDTTTPAAMATTLRKLLTGGGSASGGSGTGATSGAHIVMVDAYKPTKGGTGGS | 28.1 kDa | 9.0 |
| SpyC_C-BLA | MGGSQLDSATHIKFSKRDEDGKELAGATMELRDSSGKTISTWISDGQVKDFYLYPGKYTFVETAAPDGYEVATAITFTVNEQGQVTVNGKATKGGGSASGTGGSGTGATSGLLTLASRQQLIDWMEADKVAGPLLRSALPAGWFIADKSGAGERGSRGIIAALGPDGKPSRIVVIYTTGSQATMDERNRQIAEIGASLIKHWVDG | 21.4 kDa | 5.7 |
References
- Osak, B.; Fitzgerald, L.R. A β-lactamase reporter assay for monitoring the activation of the smoothened pathway. Assay Drug Dev. Technol. 2007, 5, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Robers, M.B.; Machleidt, T.; Carlson, C.B.; Bi, K. Cellular LanthaScreen and β-lactamase reporter assays for high-throughput screening of JAK2 inhibitors. Assay Drug Dev. Technol. 2008, 6, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Guo, V.; Huang, R.; Inglese, J.; Nirenberg, M.; Austin, C.P. A cell-based β-lactamase reporter gene assay for the CREB signaling pathway. Curr. Chem. Genom. 2009, 3, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.M.; Padilla-Parra, S. The β-lactamase assay: Harnessing a FRET biosensor to analyse viral fusion mechanisms. Sensors 2016, 16, 950. [Google Scholar] [CrossRef] [PubMed]
- Zlokarnik, G.; Negulescu, P.A.; Knapp, T.E.; Mere, L.; Burres, N.; Feng, L.; Whitney, M.; Roemer, K.; Tsien, R.Y. Quantitation of transcription and clonal selection of single living cells with β-lactamase as reporter. Science 1998, 279, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.E. Realization of β-lactamase as a versatile fluorogenic reporter. Trends Biotechnol. 2004, 22, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.A. β-Lactamase: An ideal reporter system for monitoring gene expression in live eukaryotic cells. BioTechniques 2007, 42, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, Y.; Zhang, X.; Zhang, W.; Guo, S.; Jin, F. Recent progress of cell-penetrating peptides as new carriers for intracellular cargo delivery. J. Control. Release 2014, 174, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.P.; Sharei, A.; Ding, X.; Sahay, G.; Langer, R.; Jensen, K.F. In vitro and ex vivo strategies for intracellular delivery. Nature 2016, 538, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szeto, H.H.; Schiller, P.W.; Zhao, K.; Luo, G. Fluorescent dyes alter intracellular targeting and function of cell-penetrating tetrapeptides. FASEB J. 2005, 19, 118–120. [Google Scholar] [CrossRef] [PubMed]
- El-Andaloussi, S.; Järver, P.; Johansson, H.J.; Langel, U. Cargo-dependent cytotoxicity and delivery efficacy of cell-penetrating peptides: A comparative study. Biochem. J. 2007, 407, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Puckett, C.A.; Barton, J.K. Fluorescein redirects a ruthenium-octaarginine conjugate to the nucleus. J. Am. Chem. Soc. 2009, 131, 8738–8739. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Dougherty, P.G.; Pei, D. Monitoring the cytosolic entry of cell-penetrating peptides using a pH-sensitive fluorophore. Chem. Commun. 2015, 51, 2162–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milech, N.; Longville, B.A.C.; Cunningham, P.T.; Scobie, M.N.; Bogdawa, H.M.; Winslow, S.; Anastasas, M.; Connor, T.; Ong, F.; Stone, S.R.; et al. GFP-complementation assay to detect functional CPP and protein delivery into living cells. Sci. Rep. 2015, 5, 18329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-S.; Choi, D.-K.; Park, S.-W.; Shin, S.-M.; Bae, J.; Kim, D.-M.; Yoo, T.H.; Kim, Y.-S. Quantitative assessment of cellular uptake and cytosolic access of antibody in living cells by an enhanced split GFP complementation assay. Biochem. Biophys. Res. Commun. 2015, 467, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Zakeri, B.; Fierer, J.O.; Celik, E.; Chittock, E.C.; Schwarz-Linek, U.; Moy, V.T.; Howarth, M. Peptide tag forming a rapid covalent bond to a protein, through engineering a bacterial adhesin. Proc. Natl. Acad. Sci. USA 2012, 109, E690–E697. [Google Scholar] [CrossRef] [PubMed]
- Wadia, J.S.; Stan, R.V.; Dowdy, S.F. Transducible TAT-HA fusogenic peptide enhances escape of TAT-fusion proteins after lipid raft macropinocytosis. Nat. Med. 2004, 10, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, I.M.; Wadia, J.S.; Dowdy, S.F. Cationic TAT peptide transduction domain enters cells by macropinocytosis. J. Control. Release 2005, 102, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Lönn, P.; Dowdy, S.F. Cationic PTD/CPP-mediated macromolecular delivery: Charging into the cell. Expert Opin. Drug Deliv. 2015, 12, 1627–1636. [Google Scholar] [CrossRef] [PubMed]
- Galarneau, A.; Primeau, M.; Trudeau, L.-E.; Michnick, S.W. β-Lactamase protein fragment complementation assays as in vivo and in vitro sensors of protein protein interactions. Nat. Biotechnol. 2002, 20, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.R.; Stains, C.I.; Segal, D.J.; Ghosh, I. Split β-lactamase sensor for the sequence-specific detection of DNA methylation. Anal. Chem. 2007, 79, 6702–6708. [Google Scholar] [CrossRef] [PubMed]
- Saunders, J.C.; Young, L.M.; Mahood, R.A.; Jackson, M.P.; Revill, C.H.; Foster, R.J.; Smith, D.A.; Ashcroft, A.E.; Brockwell, D.J.; Radford, S.E. An in vivo platform for identifying inhibitors of protein aggregation. Nat. Chem. Biol. 2016, 12, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Jacoby, G.A. Updated functional classification of β-lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, H.; Ozkirimli, E.; Özgür, A. Classification of beta-lactamases and penicillin binding proteins using ligand-centric network models. PLoS ONE 2015, 10, e0117874. [Google Scholar] [CrossRef] [PubMed]
- Verdurmen, W.P.R.; Thanos, M.; Ruttekolk, I.R.; Gulbins, E.; Brock, R. Cationic cell-penetrating peptides induce ceramide formation via acid sphingomyelinase: Implications for uptake. J. Control. Release 2010, 147, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Elmquist, A.; Langel, U. In vitro uptake and stability study of pVEC and its all-D analog. Biol. Chem. 2003, 384, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Pujals, S.; Fernández-Carneado, J.; Ludevid, M.D.; Giralt, E. D-SAP: A new, noncytotoxic, and fully protease resistant cell-penetrating peptide. ChemMedChem 2008, 3, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Najjar, K.; Erazo-Oliveras, A.; Brock, D.J.; Wang, T.-Y.; Pellois, J.-P. An l- to d-Amino acid conversion in an endosomolytic analog of the cell-penetrating peptide TAT influences proteolytic stability, endocytic uptake, and endosomal escape. J. Biol. Chem. 2017, 292, 847–861. [Google Scholar] [CrossRef] [PubMed]
- Baghirova, S.; Hughes, B.G.; Hendzel, M.J.; Schulz, R. Sequential fractionation and isolation of subcellular proteins from tissue or cultured cells. MethodsX 2015, 2, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Holden, P.; Horton, W.A. Crude subcellular fractionation of cultured mammalian cell lines. BMC Res. Notes 2009, 2, 243. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stone, S.R.; Heinrich, T.; Juraja, S.M.; Satiaputra, J.N.; Hall, C.M.; Anastasas, M.; Mills, A.D.; Chamberlain, C.A.; Winslow, S.; Priebatsch, K.; et al. β-Lactamase Tools for Establishing Cell Internalization and Cytosolic Delivery of Cell Penetrating Peptides. Biomolecules 2018, 8, 51. https://doi.org/10.3390/biom8030051
Stone SR, Heinrich T, Juraja SM, Satiaputra JN, Hall CM, Anastasas M, Mills AD, Chamberlain CA, Winslow S, Priebatsch K, et al. β-Lactamase Tools for Establishing Cell Internalization and Cytosolic Delivery of Cell Penetrating Peptides. Biomolecules. 2018; 8(3):51. https://doi.org/10.3390/biom8030051
Chicago/Turabian StyleStone, Shane R., Tatjana Heinrich, Suzy M. Juraja, Jiulia N. Satiaputra, Clinton M. Hall, Mark Anastasas, Anna D. Mills, Christopher A. Chamberlain, Scott Winslow, Kristin Priebatsch, and et al. 2018. "β-Lactamase Tools for Establishing Cell Internalization and Cytosolic Delivery of Cell Penetrating Peptides" Biomolecules 8, no. 3: 51. https://doi.org/10.3390/biom8030051