Effects of Pulsed Electric Field on the Cell Wall and Infection Activity of Rhizoctonia solani



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2 Experimental Method
2.2.1. Preparation of Bacterial Solution
2.2.2. PEF Treatment
2.2.3. TEM Examination
2.2.4 Determination of Chitin Content
2.2.5. Determination of PG Activity
3. Results
3.1. Effects of PEF Treatment on the Cell Wall of R. solani
3.2. Effect of PEF Treatment on the Infection Activity of R. solani Cells
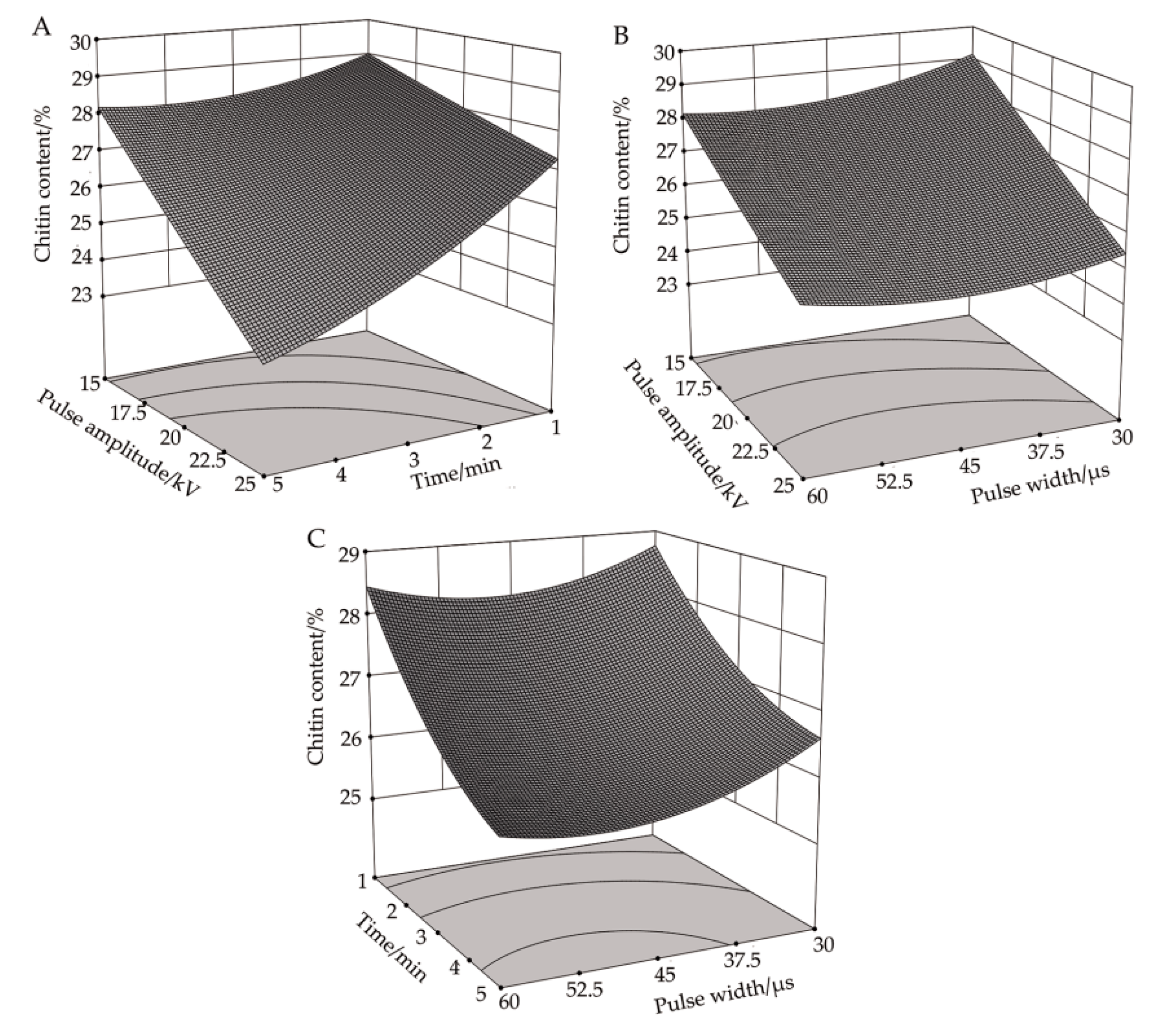
3.3. Effect of High-Voltage PEF Treatment on Chitin Content in R. solani
3.3.1. Response Surface Analysis of the Correlation between Pulse Voltage Amplitude and Processing Time on chitin Content
3.3.2. Response Surface Analysis of the Correlation between Pulse Voltage Amplitude and Pulse Width on Chitin Content
3.3.3. Response Surface Analysis of the Correlation between Pulse Width and Processing Time on Chitin Content
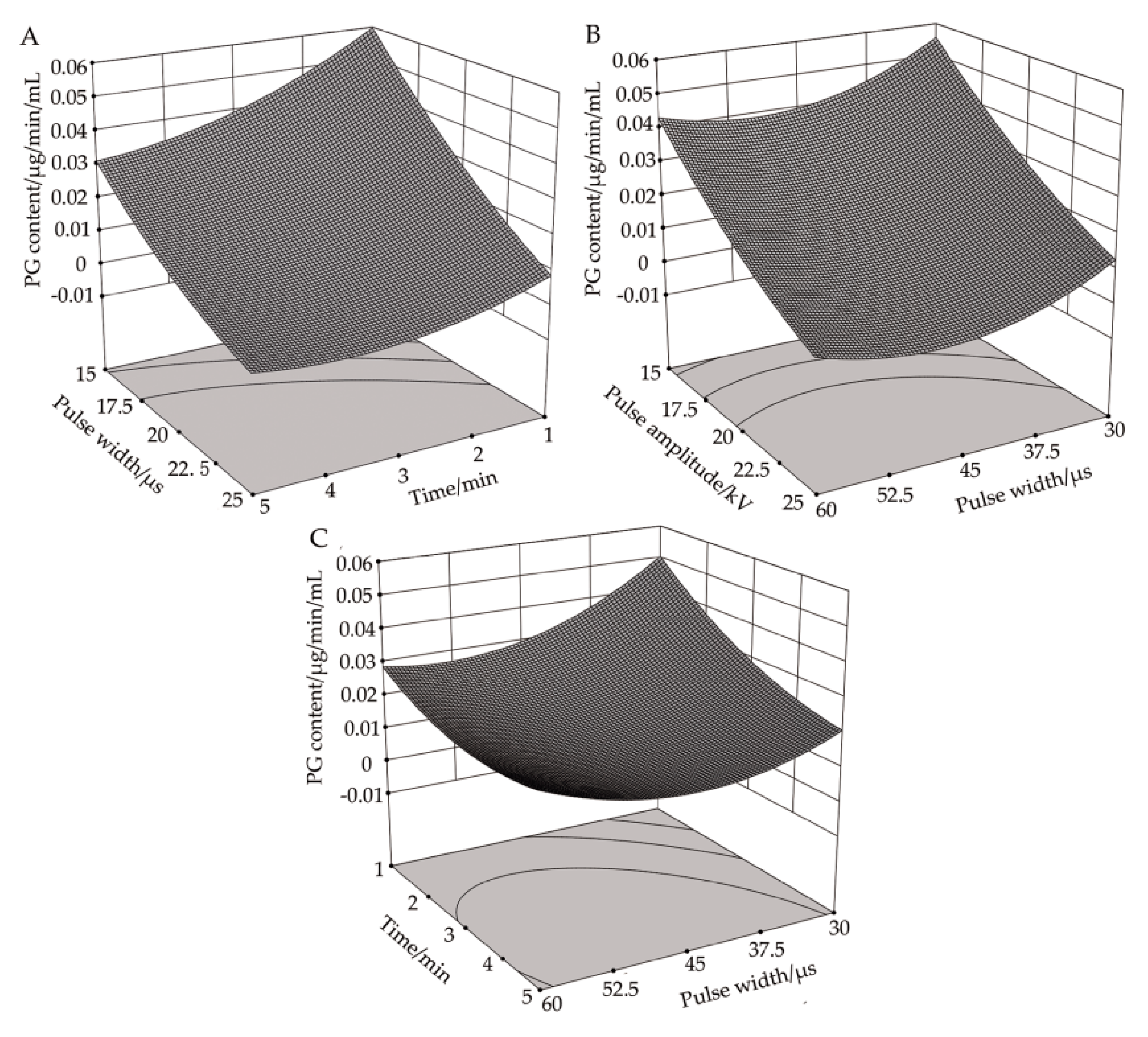
3.4. Effect of High-Voltage PEF Treatment on PG Activity
3.4.1. Response Surface Analysis of the Correlation between Pulse Voltage Amplitude and Processing Time on PG Activity
3.4.2. Response Surface Analysis of the Correlation between Pulse Voltage Amplitude and Pulse Width on PG Activity
3.4.3. Response Surface Analysis of the Correlation between Pulse Width and Processing Time on PG Activity
3.5. Parameter Optimization
3.6. Experimental Verification
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rabindran, R.; Vidhyasekaran, P. Development of a formulation of Pseudomonas fluorescens PfALR2 for management of rice sheath blight. Crop Prot. 1996, 15, 715–721. [Google Scholar] [CrossRef]
- Zuo, S.; Zhang, Y.; Yin, Y.; Li, G.; Zhang, G.; Wang, H.; Chen, Z.; Pan, X. Fine-mapping of qSB-9 TQ, a gene conferring major quantitative resistance to rice sheath blight. Mol. Breed. 2014, 34, 2191–2203. [Google Scholar] [CrossRef]
- Liu, W.; Yang, C. Research progress on biological control of rice sheath blight. Guangxi Agric. Sci. 2009, 40, 512–515. [Google Scholar]
- Meng, Q.Z.; Liu, Z.H. Research progress of rice sheath blight. Shen Yang Agric. Univ. 2001, 35, 376–381. [Google Scholar]
- Lore, J.S.; Hunjan, M.S.; Singh, P.; Willocquet, L.; Sri, S.; Savary, S. Phenotyping of Partial Physiological Resistance to Rice Sheath Blight. J. Phytopathol. 2013, 161, 224–229. [Google Scholar] [CrossRef]
- Eizenga, G.C.; Agrama, H.A.; Lee, F.N.; Yan, W.; Jia, Y. Identifying Novel Resistance Genes in Newly Introduced Blast Resistant Rice Germplasm. Crop Sci. 2006, 46, 1870–1878. [Google Scholar] [CrossRef]
- Hu, X.R. The Resistance Monitoring of Rhizoctonia Solani Tojinggangmycin and Its Resistance Risk Assessment; Fujian Agriculture and Forestry University: Fuzhou, China, 2006; pp. 1–54. [Google Scholar]
- Shi, F.Y.; Zhu, Y.D. Antagonistic effect of trichoderma elongata T8 on rice sheath blight. Chin. Agric. Sci. Bull. 2005, 21, 264–271. [Google Scholar]
- Novickij, V.; Lastauskiene, E.; Staigvila, G.; Girkontaite, I.; Zinkeviciene, A.; Svediene, J.; Paskevicius, A.; Markovskaja, S.; Novickij, J. Low concentrations of acetic and formic acids enhance the inactivation of Staphylococcus aureus and Pseudomonas aeruginosa with pulsed electric fields. BMC Microbiol. 2019, 19, 73. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Yan, Z.M. Studies on lethal dynamics of S. accha rom ycesce revisiae E.coliand penic illium by treatment time of pulse electric field. Food Mechinery 2007, 23, 11–14. [Google Scholar]
- Raso, J.; Calderón, M.L.; Gongora, M.; Barbosa-Canovas, G.; Swanson, B.G. Inactivation of Mold Ascospores and Conidiospores Suspended in Fruit Juices by Pulsed Electric Fields. LWT Food Sci. Technol. 1998, 31, 668–672. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Li, M.H.; Li, Y. Establishment of Agrobacterium tumefaciens-Mediated Transformation System for Rice Sheath Blight Pathogen Rhizoctonia solani AG-1 IA. Rice Sci. 2011, 18, 297–303. [Google Scholar] [CrossRef]
- Gong, P.; Wang, J.; Liu, B.; Ru, G.; Feng, J. Dissolution of chitin in aqueous KOH. Cellulose 2016, 23, 1705–1711. [Google Scholar] [CrossRef]
- Redgwell, R.J.; Melton, L.D.; Brasch, D.J. Cell Wall Dissolution in Ripening Kiwifruit. Plant Physiol. 1992, 98, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.F.; Yi, W. The Effect of High Electrostatic Field on Plant Cellular Transmembrane Voltage and Micro Principles. J. Wuhan Inst. Sci. Technol. 2002, 15, 16–18. [Google Scholar]
- Dai, Y.Y.; Cao, J. Research progress of chitin and chitosan in fungi. J. Zhengzhou Inst. Technol. 2004, 25, 88–90. [Google Scholar]
- O’Brien, J.A.; Daudi, A.; Butt, V.S.; Bolwell, G.P. Reactive oxygen species and their role in plant defence and cell wall metabolism. Planta 2012, 236, 765–779. [Google Scholar] [CrossRef]
- Roberts, C.A.; Barton, F.E.; Moore, K. Moore Estimation of Acremonium Coenophialum Mycelium in Infected Tall Fescue. Agron. J. 1988, 80, 737–740. [Google Scholar] [CrossRef]
- Li, H.T.; Deng, W.J. Analysisofinactivation saccharom ycescerevisiae by 13 μs,200 ns and 2 ns pulsed electric fields. Highpower Laser Partilice Beams 2015, 27, 1–6. [Google Scholar]
- Russell, A.E. Unexpected Effects of Chitin, Cellulose, and Lignin Addition on Soil Dynamics in a Wet Tropical Forest. Ecosystems 2014, 17, 918–930. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.C.; Zhang, S.L. Role of Pectolytic Enzymes and Cellulase during Ripening and Woolly Breakdown in Peaches. Acta Hortic. Sin. 2001, 28, 107–111. [Google Scholar]
- Bolin, H.R.; Stafford, A.E. Factors Affecting The Storages Stabilitys of Shredded Lettuce. J. Food Sci. 1977, 42, 1319–1321. [Google Scholar] [CrossRef]
- Qi, L.D.; Wei, J.M. Pectin POIySaCCharjde Degradation in Relation to the texture Softening in Pear Fruit. Sci. Agric. Sin. 2015, 48, 3027–3037. [Google Scholar]
- Chen, X.J.; Zhang, H. Cell Wall Degrading Enzymes Produced by Rhizoctonia solani and Their Pathogenicity to Rice Plants. J. Jiangsu Agric. 2006, 22, 24–28. [Google Scholar]
- Khan, A.; Williams, K.L.; Nevalainen, H.K.M. Effects of Paecilomyces lilacinus protease and chitinase on the eggshell structures and hatching of Meloidogyne javanica juveniles. Biol. Control 2004, 31, 346–352. [Google Scholar] [CrossRef]
- Chen, Y. Inactivation Mechanics of POD and PG Enzymes in High Voltage Pulsed Electric Field. Master’s Thesis, Fujian Agriculture and Forestry University, Fujian, China, 2007. [Google Scholar]
- Andreou, V.; Dimopoulos, G.; Katsaros, G.; Taoukis, P. Comparison of the application of high pressure and pulsed electric fields technologies on the selective inactivation of endogenous enzymes in tomato products. Innov. Food Sci. Emerg. Technol. 2016, 38, 349–355. [Google Scholar] [CrossRef]
- Aguiló-Aguayo, I.; Soliva-Fortuny, R.; Martín-Belloso, O. Comparative study on color, viscosity and related enzymes of tomato juice treated by high-intensity pulsed electric fields or heat. Eur. Food Res. Technol. 2007, 227, 599–606. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, W. Effect of pulsed electric fields on the sterilization and enzyme inactivation of pear juice. Sci. Technol. Food Ind. 2008, 29, 65–68. [Google Scholar]
- Tian, H.Y. Study on the Effect of Pulsed Electric Fields on the Sterilization and Enzyme Inactivation. Master’s Thesis, Dalian Institute of Light Industry, Dalian, China, 2005. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent Name | Reagent Parameter |
|---|---|
| KOH | 100% |
| Polygalacturonic acid | 10 g/L |
| Acetic acid-sodium acetate buffer | 0.05 mol/L, pH=5.5 |
| DNS | — |
| Glutaraldehyde solution | 2.5% |
| Osmic acid solution | 1% |
| Phosphate buffer | 0.1 m, pH = 7.0 |
| Ethanol | 50%, 70%, 80%, 90%, 95%, 100% |
| Acetone | 100% |
| Level | Factors | ||
|---|---|---|---|
| Pulse Width (μs) | Processing Time (min) | Pulse Voltage Amplitude (kV) | |
| -1 | 30 | 1 | 15 |
| 0 | 45 | 3 | 20 |
| 1 | 60 | 5 | 25 |
| Test NO. | Pulse Width (μs) | Processing Time (min) | Pulse Voltage Amplitude (kV) | PG Activity (U) | Chitin Content (%) |
|---|---|---|---|---|---|
| Control check | 0 | 0 | 0 | 0.059 | 29 |
| 1 | 30 | 5 | 20 | 0.024** | 26.53** |
| 2 | 45 | 3 | 20 | 0.016** | 26.59** |
| 3 | 45 | 1 | 25 | 0.012** | 27.12** |
| 4 | 45 | 5 | 25 | 0.002** | 23.26** |
| 5 | 45 | 3 | 20 | 0.016** | 26.41** |
| 6 | 60 | 5 | 20 | 0.02** | 25.87** |
| 7 | 30 | 3 | 25 | 0.008** | 25.67** |
| 8 | 45 | 3 | 20 | 0.016** | 26.75** |
| 9 | 30 | 1 | 20 | 0.052ns | 28.73ns |
| 10 | 45 | 3 | 20 | 0.015** | 26.35** |
| 11 | 45 | 1 | 15 | 0.058** | 29.14ns |
| 12 | 60 | 3 | 15 | 0.048** | 27.95ns |
| 13 | 45 | 3 | 20 | 0.016** | 26.49** |
| 14 | 45 | 5 | 15 | 0.028** | 28.37** |
| 15 | 60 | 3 | 25 | 0.004** | 25.46** |
| 16 | 60 | 1 | 20 | 0.026** | 28.53ns |
| 17 | 30 | 3 | 15 | 0.056ns | 29.04ns |
| Source of Variation | Chitin Content | PG Activity | ||
|---|---|---|---|---|
| F value | P value | F value | P value | |
| Model | 87.57 | <0.0001 | 41.38 | <0.0001 |
| V | 444.58 | <0.0001 | 245.66 | <0.0001 |
| PW | 12.29 | 0.0099 | 16.11 | 0.0051 |
| t | 237.28 | <0.0001 | 50.02 | 0.0002 |
| V·t | 50.31 | 0.0002 | 7.31 | 0.0305 |
| V·PW | 4.08 | 0.0831 | 0.29 | 0.6055 |
| PW·t | 1.12 | 0.3216 | 8.84 | 0.0207 |
| V2 | 0.11 | 0.7530 | 4.56 | 0.0701 |
| t2 | 15.64 | 0.0055 | 8.81 | 0.0209 |
| PW2 | 20.21 | 0.0028 | 26.90 | 0.0013 |
| R2 | 0.9912 | 0.9815 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; He, P.; Pan, C.; Xu, J.; Xue, B.; Yin, W.; Qian, Y. Effects of Pulsed Electric Field on the Cell Wall and Infection Activity of Rhizoctonia solani. Biology 2019, 8, 73. https://doi.org/10.3390/biology8040073
Feng X, He P, Pan C, Xu J, Xue B, Yin W, Qian Y. Effects of Pulsed Electric Field on the Cell Wall and Infection Activity of Rhizoctonia solani. Biology. 2019; 8(4):73. https://doi.org/10.3390/biology8040073
Chicago/Turabian StyleFeng, Xuebin, Peijun He, Chaoya Pan, Jin Xu, Baoshan Xue, Wenqing Yin, and Yan Qian. 2019. "Effects of Pulsed Electric Field on the Cell Wall and Infection Activity of Rhizoctonia solani" Biology 8, no. 4: 73. https://doi.org/10.3390/biology8040073
APA StyleFeng, X., He, P., Pan, C., Xu, J., Xue, B., Yin, W., & Qian, Y. (2019). Effects of Pulsed Electric Field on the Cell Wall and Infection Activity of Rhizoctonia solani. Biology, 8(4), 73. https://doi.org/10.3390/biology8040073