Rational Design of an Orthogonal Pair of Bimolecular RNase P Ribozymes through Heterologous Assembly of Their Modular Domains


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plasmid Construction and RNA Preparation
2.2. Ribozyme Activity Assay
2.3. Gel Electrophoresis Mobility Shift Assay (EMSA)
3. Results
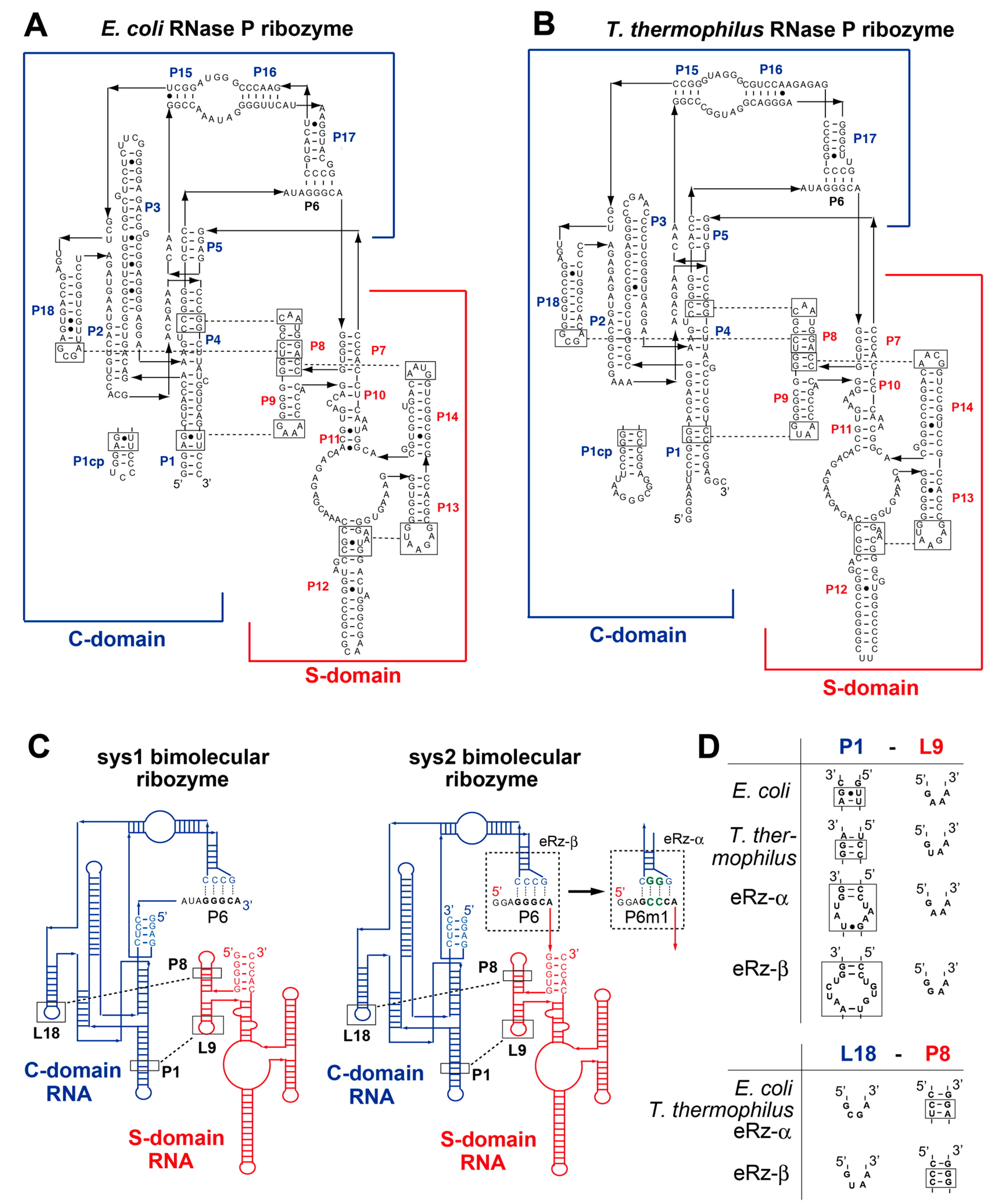
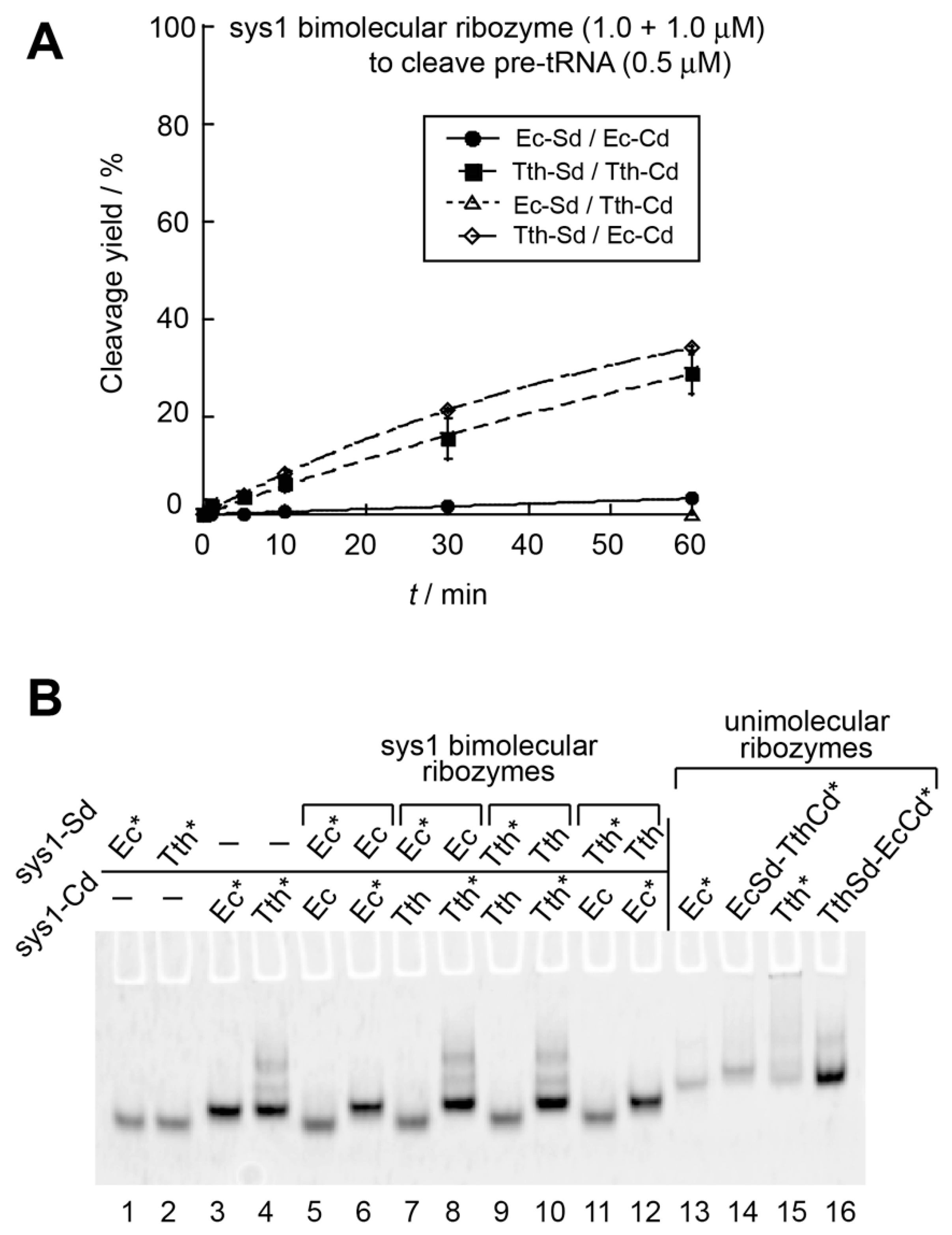
3.1. Optimization of the Bimolecular RNase P Ribozymes by Domain Chimera
3.2. Modular Engineering of L9–P1 Interdomain Interactions
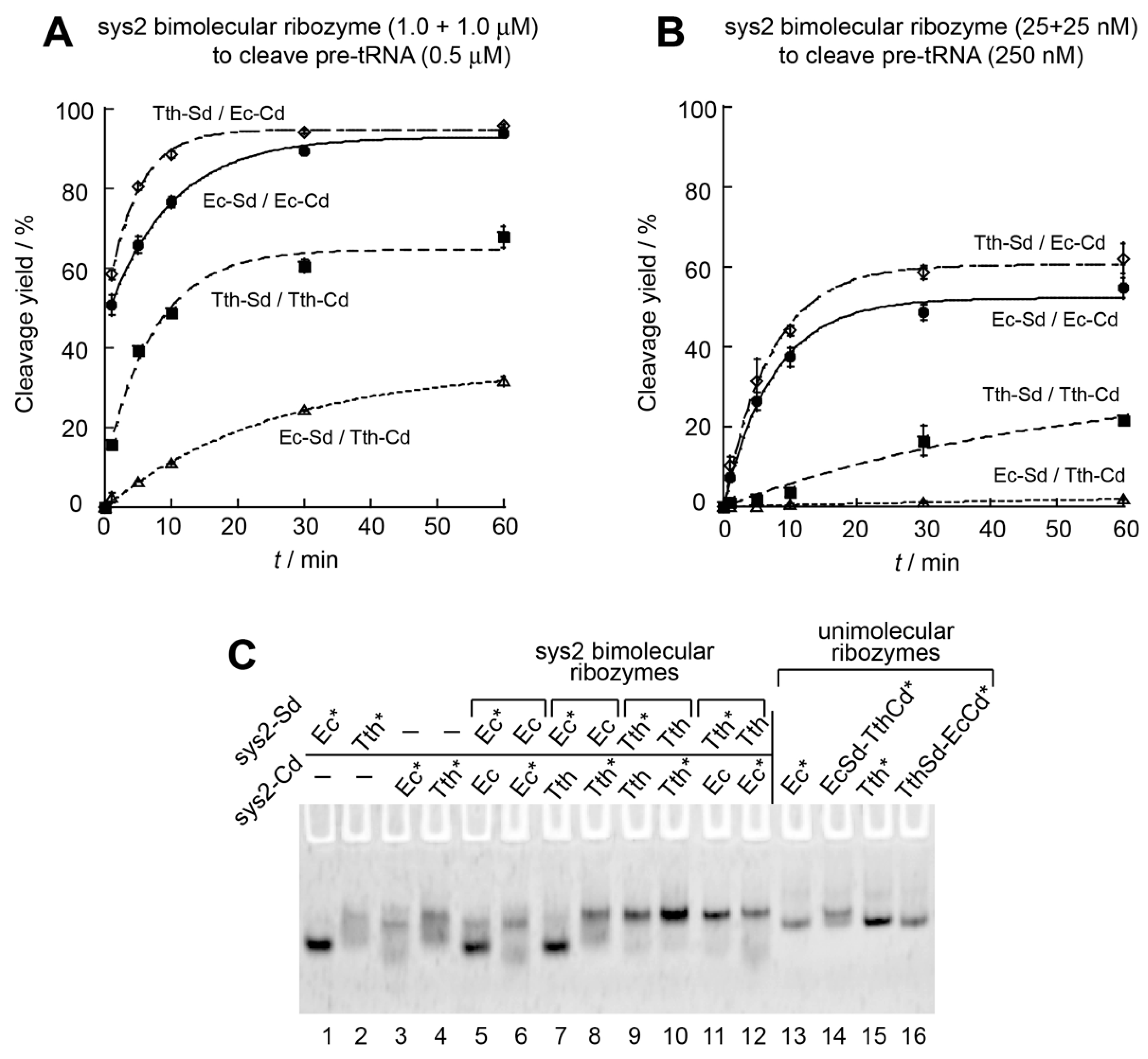
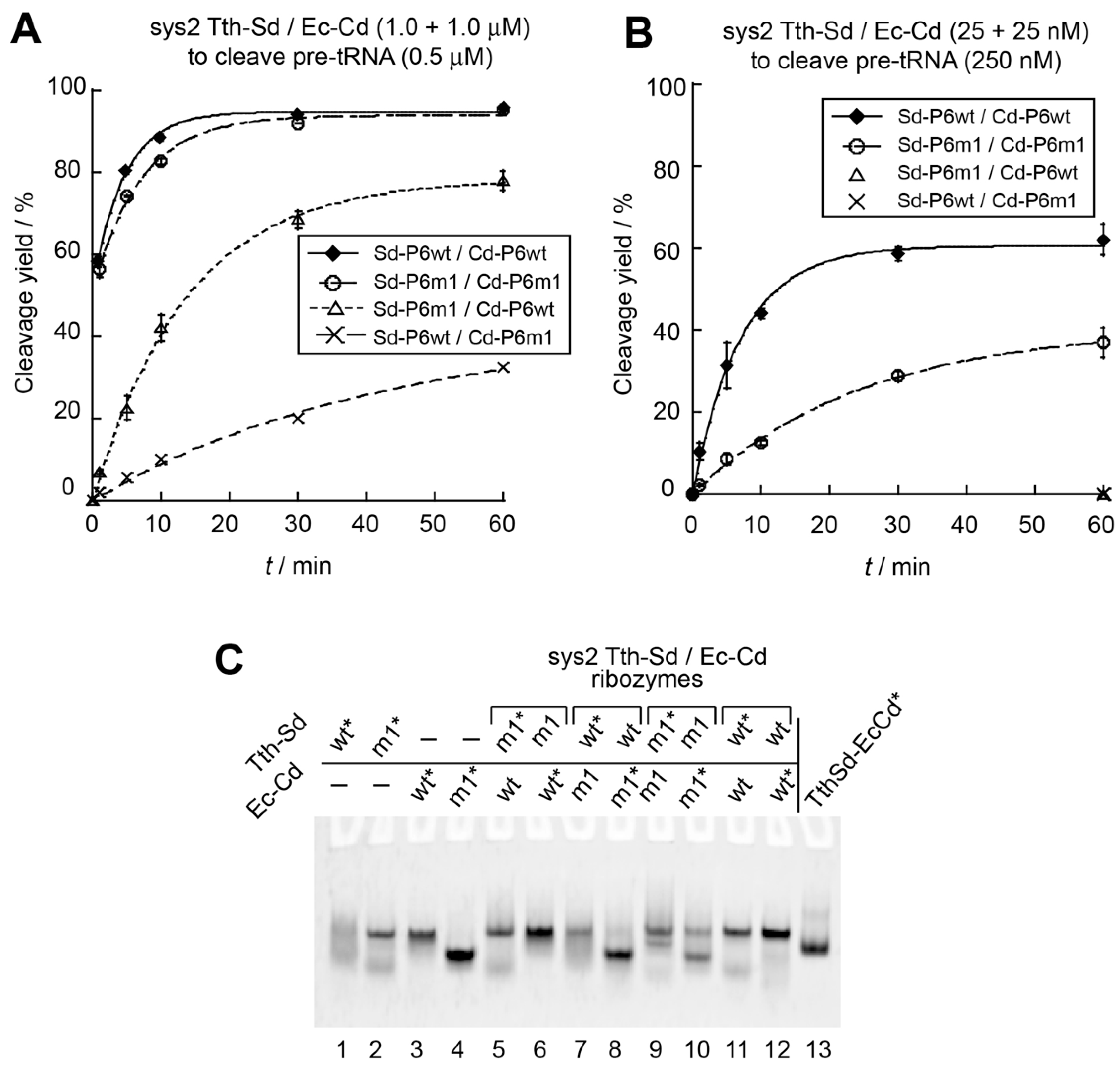
3.3. Engineering of P6 Base Pair Interaction in the Sys2 Ribozyme
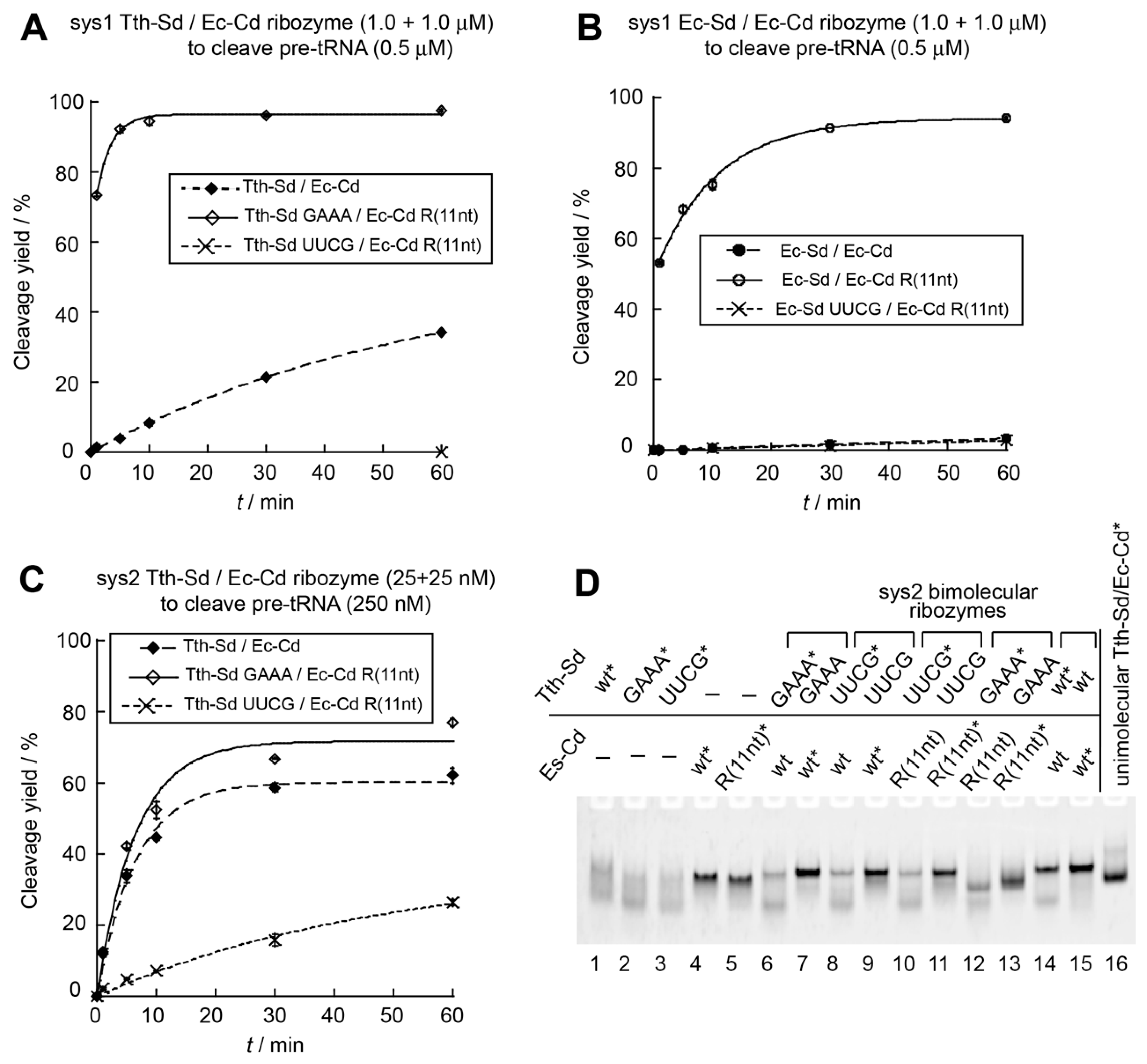
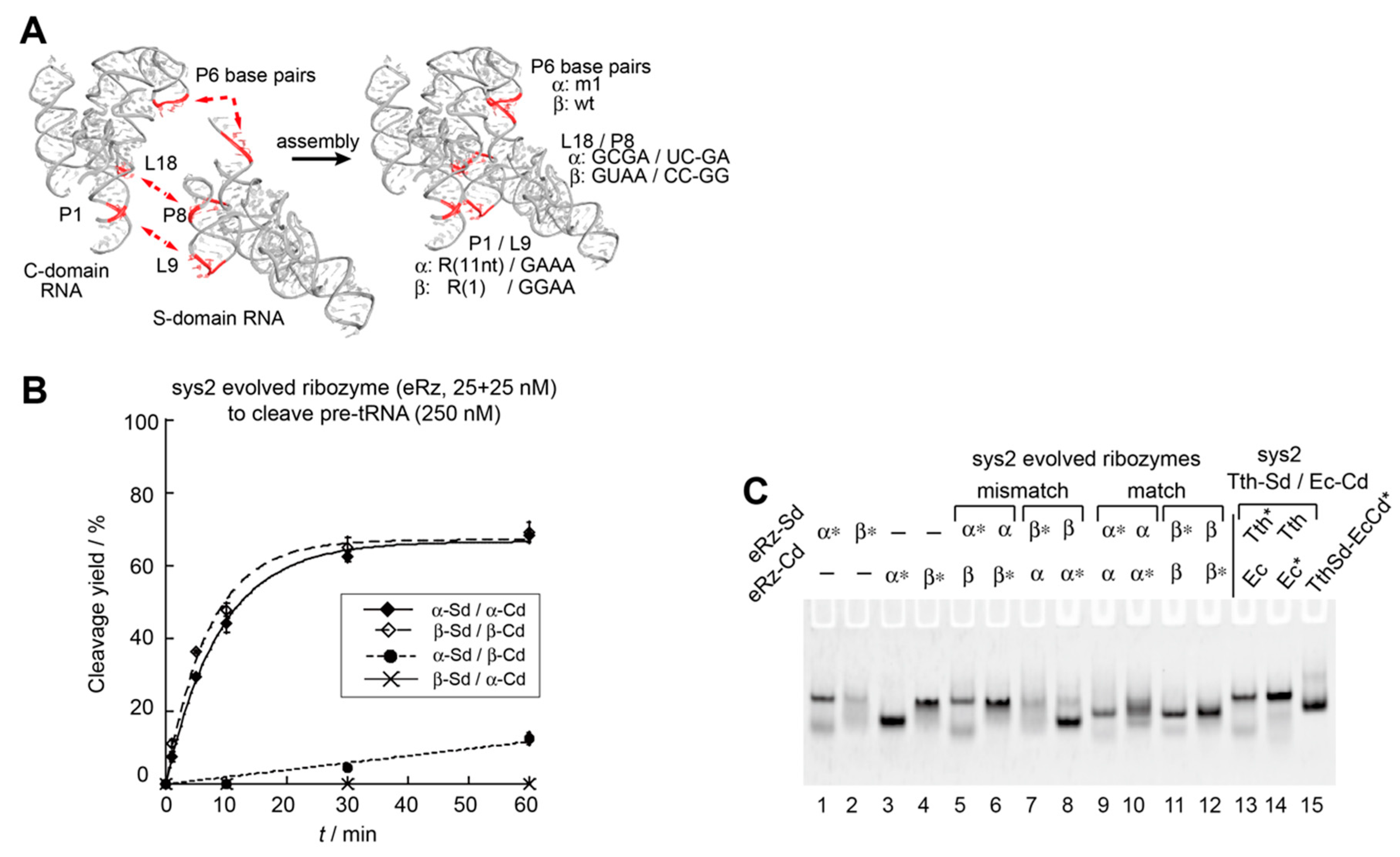
3.4. Construction an Orthogonal Pair of Sys2 Bimolecular Ribozymes with Improved Catalytic Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kummerfeld, S.K.; Teichmann, S.A. Relative rates of gene fusion and fission in multi-domain proteins. Trends Genet. 2005, 21, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Bornberg-Bauer, E.; Beaussart, F.; Kummerfeld, S.K.; Teichmann, S.A.; Weiner, J., 3rd. The evolution of domain arrangements in proteins and interaction networks. Cell. Mol. Life Sci. 2005, 62, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Galarneau, A.; Primeau, M.; Trudeau, L.E.; Michnick, S.W. Beta-lactamase protein fragment complementation assays as in vivo and in vitro sensors of protein protein interactions. Nat. Biotechnol. 2002, 20, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Ojennus, D.D.; Lehto, S.E.; Wuttke, D.S. Electrostatic interactions in the reconstitution of an SH2 domain from constituent peptide fragments. Protein Sci. 2003, 12, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, T. Designing split reporter proteins for analytical tools. Anal. Chim. Acta 2006, 556, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Kerppola, T.K. Visualization of molecular interactions using bimolecular fluorescence complementation analysis: Characteristics of protein fragment complementation. Chem. Soc. Rev. 2009, 38, 2876–2886. [Google Scholar] [CrossRef] [PubMed]
- Na, K.; Li, H. Protein Fragment Reconstitution as a Driving Force for Self-Assembling Reversible Protein Hydrogels. Adv. Funct. Mater. 2015, 25, 5593–5601. [Google Scholar]
- Jarrell, K.A.; Dietrich, R.C.; Perlman, P.S. Group II intron domain 5 facilitates a trans-splicing reaction. Mol. Cell. Biol. 1988, 8, 2361–2366. [Google Scholar] [CrossRef]
- Van der Horst, G.; Christian, A.; Inoue, T. Reconstitution of a group I intron self-splicing reaction with an activator RNA. Proc. Natl. Acad. Sci. USA 1991, 88, 184–188. [Google Scholar] [CrossRef]
- Doudna, J.A.; Cech, T.R. Self-assembly of a group I intron active site from its component tertiary structural domains. RNA 1995, 1, 36–45. [Google Scholar]
- Ikawa, Y.; Shiraishi, H.; Inoue, T. Trans-activation of the Tetrahymena ribozyme by its P2–2.1 domains. J. Biochem. 1998, 123, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Matsumura, S.; Ikawa, Y. Effects of molecular crowding on a bimolecular group I ribozyme and its derivative that self-assembles to form ribozyme oligomers. Biochem. Biophys. Res. Commun. 2018, 507, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Pan, T. Higher order folding and domain analysis of the ribozyme from Bacillus subtilis ribonuclease P. Biochemistry 1995, 34, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Loria, A.; Pan, T. Domain structure of the ribozyme from eubacterial ribonuclease P. RNA 1996, 2, 551–563. [Google Scholar] [PubMed]
- Kim, H.; Poelling, R.R.; Leeper, T.C.; Meyer, M.A.; Schmidt, F.J. In vitro transactivation of Bacillus subtilis RNase P RNA. FEBS Lett. 2001, 506, 235–238. [Google Scholar] [CrossRef]
- Torres-Larios, A.; Swinger, K.K.; Pan, T.; Mondragón, A. Structure of ribonuclease P-a universal ribozyme. Curr. Opin. Struct. Biol. 2006, 16, 327–335. [Google Scholar] [CrossRef]
- Kazantsev, A.V.; Pace, N.R. Bacterial RNase P: A new view of an ancient enzyme. Nat. Rev. Microbiol. 2006, 4, 729–740. [Google Scholar] [CrossRef]
- Evans, D.S.; Marquez, M.; Pace, N.R. RNase P: Interface of the RNA and protein worlds. Trends Biochem. Sci. 2006, 31, 333–341. [Google Scholar] [CrossRef]
- Mondragón, A. Structural studies of RNase P. Annu. Rev. Biophys. 2013, 42, 537–557. [Google Scholar] [CrossRef]
- Nozawa, Y.; Hagihara, M.; Matsumura, S.; Ikawa, Y. Modular architecture of bacterial RNase P ribozymes as a structural platform for RNA nanostructure design. Chimia 2018, 72, 882–887. [Google Scholar] [CrossRef]
- Tsuruga, R.; Uehara, N.; Suzuki, Y.; Furuta, H.; Sugiyama, H.; Endo, M.; Matsumura, S.; Ikawa, Y. Oligomerization of a modular ribozyme assembly of which is controlled by a programmable RNA-RNA interface between two structural modules. J. Biosci. Bioeng. 2019. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Matsumura, S.; Ikawa, Y. Oligomerization of a bimolecular ribozyme modestly rescues its structural defects that disturb interdomain assembly to form the catalytic site. J. Mol. Evol. 2018, 86, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Vieille, C.; Zeikus, G.J. Hyperthermophilic enzymes: Sources, uses, and molecular mechanisms for thermostability. Microbiol. Mol. Biol. Rev. 2001, 65, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Torres-Larios, A.; Swinger, K.K.; Krasilnikov, A.S.; Pan, T.; Mondragón, A. Crystal structure of the RNA component of bacterial ribonuclease P. Nature 2005, 437, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Kazantsev, A.V.; Krivenko, A.A.; Harrington, D.J.; Holbrook, S.R.; Adams, P.D.; Pace, N.R. Crystal structure of a bacterial ribonuclease P RNA. Proc. Natl. Acad. Sci. USA 2005, 102, 13392–13397. [Google Scholar] [CrossRef] [PubMed]
- Krasilnikov, A.S.; Xiao, Y.; Pan, T.; Mondragón, A. Basis for structural diversity in homologous RNAs. Science 2004, 306, 104–107. [Google Scholar] [CrossRef]
- Hartmann, R.K.; Erdmann, V.A. Analysis of the gene encoding the RNA subunit of ribonuclease P from T. thermophilus HB8. Nucleic Acids Res. 1991, 19, 5957–5964. [Google Scholar] [CrossRef]
- Ikawa, Y.; Moriyama, S.; Furuta, H. Facile syntheses of BODIPY derivatives for fluorescent labeling of the 3′ and 5′ ends of RNAs. Facile syntheses of BODIPY derivatives for fluorescent labeling of the 3′ and 5′ ends of RNAs. Anal. Biochem. 2008, 378, 166–170. [Google Scholar] [CrossRef]
- Ando, T.; Tanaka, T.; Hori, Y.; Kikuchi, Y. Kinetics of hyperprocessing reaction of human tyrosine tRNA by ribonuclease P ribozyme from Escherichia coli. Biosci. Biotechnol. Biochem. 2002, 66, 1967–1971. [Google Scholar] [CrossRef]
- Brown, J.W.; Haas, E.S.; Pace, N.R. Characterization of ribonuclease P RNAs from thermophilic bacteria. Nucleic Acids Res. 1993, 21, 671–679. [Google Scholar] [CrossRef]
- Paul, R.; Lazarev, D.; Altman, S. Characterization of RNase P from Thermotoga maritima. Nucleic Acids Res. 2001, 29, 880–885. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Marszalkowski, M.; Willkomm, D.K.; Hartmann, R.K. Structural basis of a ribozyme’s thermostability: P1-L9 interdomain interaction in RNase P RNA. RNA 2008, 14, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Schlegl, J.; Hardt, W.D.; Erdmann, V.A.; Hartmann, R.K. Contribution of structural elements to Thermus thermophilus ribonuclease P RNA function. EMBO J. 1994, 13, 4863–4869. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Gössringer, M.; Hartmann, R.K. Archaeal-bacterial chimeric RNase P RNAs: Towards understanding RNA’s architecture, function and evolution. ChemBioChem 2011, 12, 1536–1543. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, L.; Michel, F.; Westhof, E. Involvement of a GNRA tetraloop in long-range RNA tertiary interactions. J. Mol. Biol. 1994, 236, 1271–1276. [Google Scholar] [CrossRef]
- Costa, M.; Michel, F. Frequent use of the same tertiary motif by self-folding RNAs. EMBO J. 1995, 14, 1276–1285. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, J.; Fujita, Y.; Maeda, Y.; Furuta, H.; Ikawa, Y. GNRA/receptor interacting modules: Versatile modular units for natural and artificial RNA architectures. Methods 2011, 54, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Geary, C.; Baudrey, S.; Jaeger, L. Comprehensive features of natural and in vitro selected GNRA tetraloop-binding receptors. Nucleic Acids Res. 2008, 36, 1138–1152. [Google Scholar] [CrossRef] [PubMed]
- Dreyfus, D.H.; Tompkins, S.M.; Fuleihan, R.; Ghoda, L.Y. Gene silencing in the therapy of influenza and other respiratory diseases: Targeting to RNase P by use of External Guide Sequences (EGS). Biologics 2007, 1, 425–432. [Google Scholar]
- Davies-Sala, C.; Soler-Bistué, A.; Bonomo, R.A.; Zorreguieta, A.; Tolmasky, M.E. External guide sequence technology: A path to development of novel antimicrobial therapeutics. Ann. N. Y. Acad. Sci. 2015, 1354, 98–110. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nozawa, Y.; Hagihara, M.; Rahman, M.S.; Matsumura, S.; Ikawa, Y. Rational Design of an Orthogonal Pair of Bimolecular RNase P Ribozymes through Heterologous Assembly of Their Modular Domains. Biology 2019, 8, 65. https://doi.org/10.3390/biology8030065
Nozawa Y, Hagihara M, Rahman MS, Matsumura S, Ikawa Y. Rational Design of an Orthogonal Pair of Bimolecular RNase P Ribozymes through Heterologous Assembly of Their Modular Domains. Biology. 2019; 8(3):65. https://doi.org/10.3390/biology8030065
Chicago/Turabian StyleNozawa, Yuri, Megumi Hagihara, Md Sohanur Rahman, Shigeyoshi Matsumura, and Yoshiya Ikawa. 2019. "Rational Design of an Orthogonal Pair of Bimolecular RNase P Ribozymes through Heterologous Assembly of Their Modular Domains" Biology 8, no. 3: 65. https://doi.org/10.3390/biology8030065
APA StyleNozawa, Y., Hagihara, M., Rahman, M. S., Matsumura, S., & Ikawa, Y. (2019). Rational Design of an Orthogonal Pair of Bimolecular RNase P Ribozymes through Heterologous Assembly of Their Modular Domains. Biology, 8(3), 65. https://doi.org/10.3390/biology8030065