Postnatal Growth and Development of the Rumen: Integrating Physiological and Molecular Insights


School of Animal Sciences, Virginia Tech, Blacksburg, VA 24061, USA
*
Author to whom correspondence should be addressed.
Biology 2024, 13(4), 269; https://doi.org/10.3390/biology13040269
Submission received: 14 March 2024
/
Revised: 5 April 2024
/
Accepted: 16 April 2024
/
Published: 18 April 2024
(This article belongs to the Section Physiology)
Abstract
:Simple Summary
The rumen is a large organ unique to cattle and other ruminants and is essential for the ingestion, digestion, and absorption of large quantities of plant-based food. Cattle and likely all ruminants, however, are not born with a voluminous, functional rumen. There is remarkable postnatal growth and development in the rumen. This review summarizes the major postnatal changes in the structure and function of the bovine rumen, the major factors causing these changes, and, more importantly, the biological mechanisms underlying these changes.
Abstract
The rumen plays an essential role in the physiology and production of agriculturally important ruminants such as cattle. Functions of the rumen include fermentation, absorption, metabolism, and protection. Cattle are, however, not born with a functional rumen, and the rumen undergoes considerable changes in size, histology, physiology, and transcriptome from birth to adulthood. In this review, we discuss these changes in detail, the factors that affect these changes, and the potential molecular and cellular mechanisms that mediate these changes. The introduction of solid feed to the rumen is essential for rumen growth and functional development in post-weaning calves. Increasing evidence suggests that solid feed stimulates rumen growth and functional development through butyric acid and other volatile fatty acids (VFAs) produced by microbial fermentation of feed in the rumen and that VFAs stimulate rumen growth and functional development through hormones such as insulin and insulin-like growth factor I (IGF-I) or through direct actions on energy production, chromatin modification, and gene expression. Given the role of the rumen in ruminant physiology and performance, it is important to further study the cellular, molecular, genomic, and epigenomic mechanisms that control rumen growth and development in postnatal ruminants. A better understanding of these mechanisms could lead to the development of novel strategies to enhance the growth and development of the rumen and thereby the productivity and health of cattle and other agriculturally important ruminants.
1. Introduction
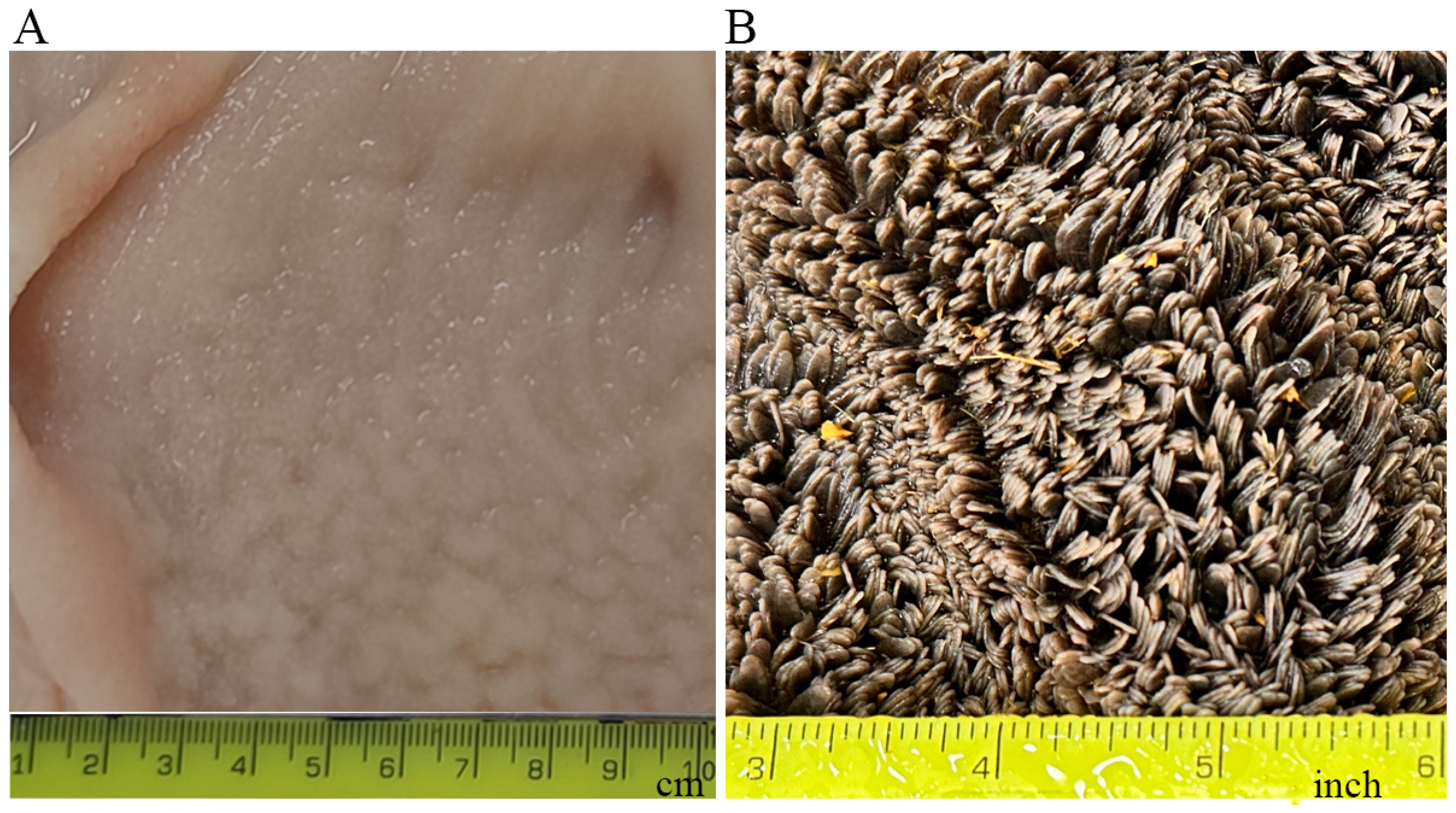
Ruminants possess a complex stomach comprised of four compartments: rumen, reticulum, omasum, and abomasum. Among these, the rumen stands as the largest compartment in adult ruminants [1,2]. The rumen has an oval shape, and the adult rumen is internally divided into different sacs by the inflection of the muscular ruminal walls called ruminal pillars [2,3]. The inner surface of the rumen is covered with numerous finger-like projections called rumen papillae (Figure 1), which play an important role in nutrient absorption and metabolism [1,2]. The rumen plays an essential role in ruminant physiology and production. However, ruminants are not born with a large, functional rumen. The rumen grows considerably in mass and size after weaning [4]. The postnatal development of the bovine rumen can be divided into three phases: the pre-ruminant phase, which spans the initial two to three weeks when the calf relies solely on milk or milk replacer; the transitional phase, commencing once the calf begins to ingest solid feed until weaning; and the ruminant phase, occurring post-weaning when the rumen attains full maturation, and the calf uses volatile fatty acids (VFAs), the major products of fermentation, as its primary energy source [5]. Many of the functions of the rumen such as digestion, absorption, and metabolism are gained after weaning [3,5,6,7]. The development of the rumen from the pre-ruminant phase to the ruminant phase is controlled by a complex interplay of factors and mediators. These factors and mediators include solid feed, microbes, VFAs, hormones, and genes. Much research has been conducted to understand how the rumen develops during the postnatal stage. This review aims to summarize the postnatal changes in the bovine rumen, the factors affecting postnatal rumen growth and development, and the biological mechanisms underlying these effects.
2. Histology of the Rumen
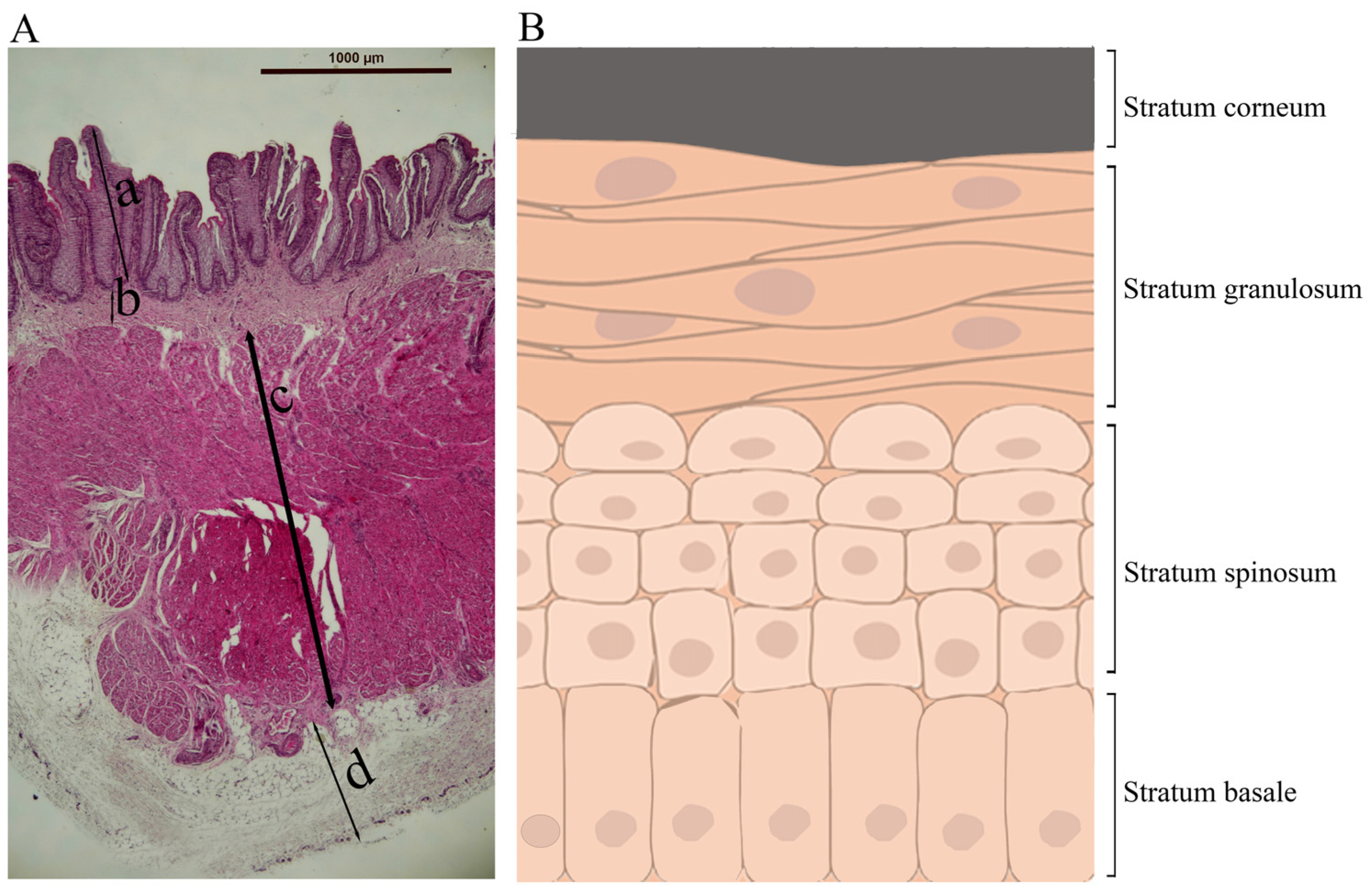
Histologically, the rumen wall is composed of tunica mucosa, lamina propria, tunica submucosa, tunica muscularis, and tunica serosa, from inside to outside [4] (Figure 2A). Tunica mucosa is the innermost part of the rumen, is non-glandular, and has a superficial stratified squamous epithelium. The rumen epithelium consists of four cellular layers or strata: stratum basale, stratum spinosum, stratum granulosum, and stratum corneum, from base to apex (Figure 2B). Cells in these layers differ in morphology and function [4,8]. The cells in the basal layer (stratum basale) are columnar and play a role in nutrient absorption, ketogenesis, energy metabolism, and secretion of immunoglobulins [4,9]. The cells in the spinous layer (stratum spinosum) and granular layer (stratum granulosum) are multilayered and have the presence of tight junctions, adherens junctions, and desmosomes, which provide strong adhesion, resist mechanical stress, and maintain structural integrity [4,10,11]. Neither the stratum corneum nor the stratum basale expresses junctional proteins and hence has no barrier function [12]. The cells of the stratum granulosum lie perpendicular to the cells of the stratum spinosum [13]. The cells in the outermost layer (stratum corneum) of the rumen epithelium are highly keratinized and devoid of nuclei, and they serve as a protective physical barrier against rough, fibrous ingesta, and potentially toxic compounds [10,13]. It is believed that the keratinized cells of the stratum corneum are derived from the cells of the stratum granulosum. During this transition, granular cells lose their organelles, including the nuclei, ribosomes, mitochondria, and Golgi apparatuses, and as such, they cannot proliferate. Subsequently, all cellular filaments along with keratohyalin and endoplasmic reticulum protein complexes are displaced toward the cell periphery. As a result, a flattened horny cell with condensed content is formed. This horny cell is enveloped by the thickened plasma membrane, and during this transformation, only keratinizing epithelia filaments are retained, which subsequently get converted into a cohesive and protective keratinized stratum corneum [14].
The lamina propria fuses with the submucosa and supports, nourishes, and protects the mucosal layer [4]. There is an extension of the lamina propria and submucosa into the papillae [15]. These fused propria-submucosa layers lie beneath the mucosa and contain blood vessels, nerves, and lymphatic vessels, providing structural support, facilitating nutrient and oxygen delivery, and regulating physiological and immune functions [4,10]. The tunica muscularis consists of the longitudinal and circular smooth muscular layers responsible for the movement and contraction of the rumen, promoting effective mixing and fermentation of feed in the rumen [15]. Recently, telocytes have been identified as a novel structural component within and between the longitudinal and circular smooth muscles of the rumen, and they function to support the muscular layer and facilitate intercellular signaling, either directly or indirectly, through the telocyte network [16]. The outermost layer of the rumen is the tunica serosa, a thin layer of smooth tissue that functions to minimize the friction of the rumen with the surrounding organs [4,15].
3. Functions of the Rumen
3.1. Digestion
The rumen itself does not produce enzymes for the breakdown of plants and other feed eaten by a ruminant; however, it provides a habitat for various microbes, which include bacteria, protozoa, archaea, and different kinds of fungi that can break down complex plant and feed materials into readily available products for utilization [17,18,19,20]. These microbes can be categorized as cellulolytic, amylolytic, and proteolytic, responsible for degrading the cellulose, starch, and protein components of the feed, respectively [19]. Bacteria and protozoa are responsible for most of the digestion in the rumen, with bacteria accounting for 80% and protozoa 20% of the digestion [19,21].
Microbial digestion of complex plant polysaccharides produces short-chain fatty acids (SCFAs), which are also known as volatile fatty acids (VFAs), gases, and heat [19,22,23]. Major VFAs produced by rumen fermentation include acetic acid, propionic acid, and butyric acid [3,22]. The typical ratio of these three VFAs in the rumen ranges from 75:15:10 to 40:40:20, depending on dietary composition [22]. Major gases produced in the rumen include methane, ammonia, and carbon dioxide. Microbial digestion of complex polysaccharides in the rumen also yields lactic acid, formic acid, succinic acid, and other short-chain monocarboxylic acids [3].
Rumen microbes can convert esterified plant lipids to unsaturated fatty acids, which are further converted to saturated fatty acids by biohydrogenation [17,24]. Proteases and peptidases from rumen microbes diligently degrade feed proteins into peptides and amino acids, which are then incorporated either into microbial proteins or deaminated to form VFAs [25]. Microbial proteins are eventually digested and absorbed by the abomasum and the small intestines, accounting for 80% of total absorbable proteins in the ruminants [17,25]. Besides digestion, rumen bacteria can synthesize vitamin B complex and vitamin K [15,26]. In addition, the rumen epithelium can absorb ammonia and metabolize it to glutamic acid [27]. The rumen is also involved in waste elimination by moving methane, carbon dioxide, hydrogen, ammonia, and some other end products of rumen fermentation and undigested materials to the hindgut [28]. In addition, the rumen is involved in converting the potentially toxic products of rumen fermentation into metabolites that have less toxicity [29].
Ruminants have a special ability to regurgitate their ingesta from the reticulum to the esophagus and finally to the mouth for re-mastication, also known as cud chewing. This allows them to decrease the particle size and more efficiently digest the feed. It is believed that cud chewing also helps the ruminants provide saliva to maintain optimum rumen pH, increase the concentration of rumen microbes, and maintain the motility of the rumen [30].
3.2. Absorption of VFAs
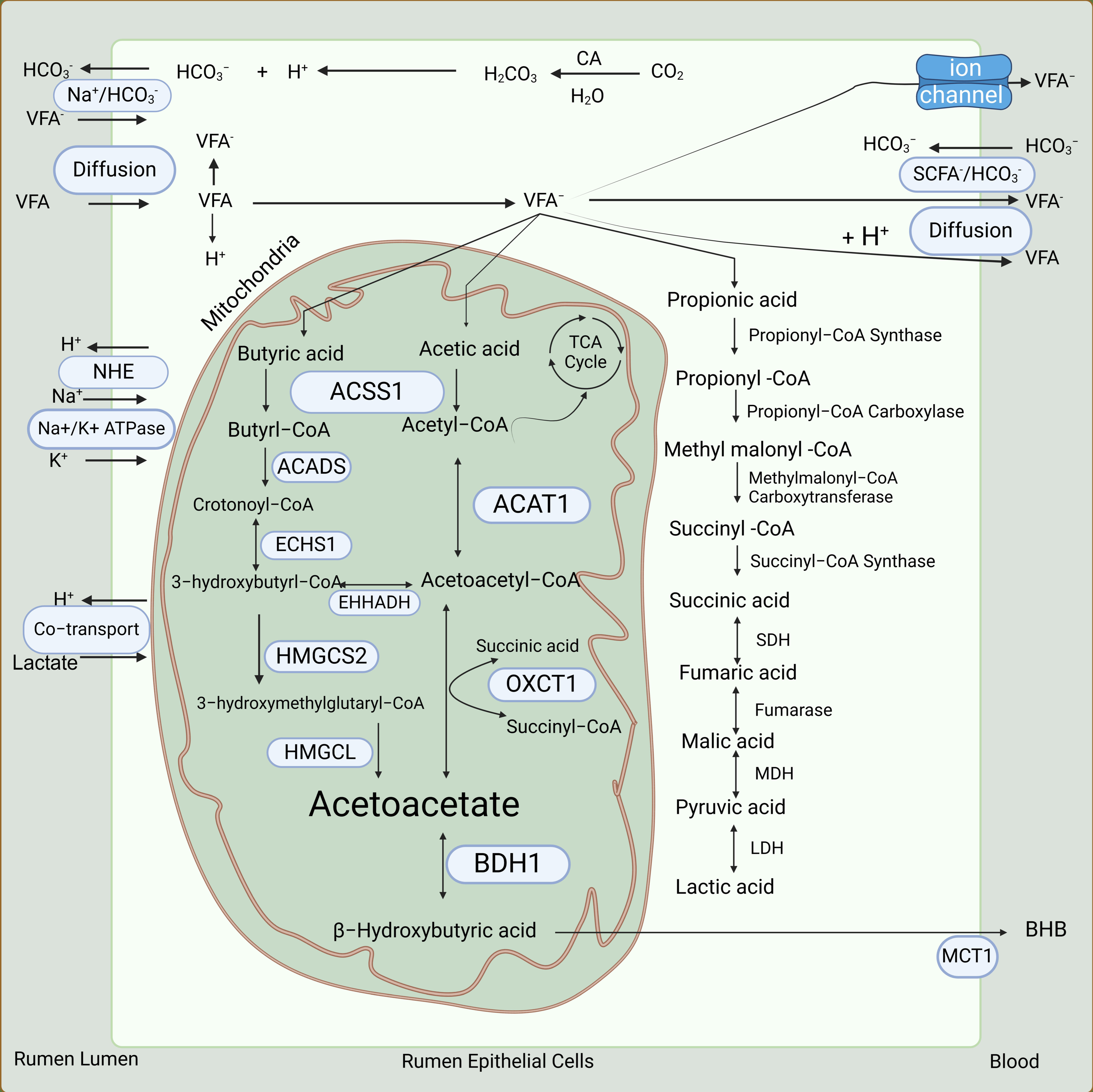
The VFAs are the most important products of rumen fermentation. A dairy cow produces more than 100 moles of VFAs a day in the rumen [31,32]. Nearly 90% of total VFAs produced in the rumen are absorbed by the rumen epithelium [22]. The remaining VFAs are absorbed by the reticulum, omasum, abomasum, and to some extent, the small intestines [33]. The rate of absorption is directly proportional to the increasing chain length of fatty acids; thus, the rate of absorption of butyric acid is higher than that of propionic or acetic acid [22,33]. The number and size of the papillae are directly proportional to the capacity of the rumen to absorb VFAs, as they increase the surface area of the rumen epithelium [34]. Three sequential processes are involved in the absorption and transport of VFAs from the rumen (Figure 3). Firstly, equilibration of VFAs occurs between the rumen and the apical surface of the rumen epithelium. Subsequently, VFAs are absorbed into the rumen epithelium. Finally, the capillaries in the rumen epithelium remove VFAs from the basal side of the rumen epithelium into the blood [32,35].
Due to the high concentration of VFAs in the rumen, a concentration gradient of VFAs exists between the rumen and the bloodstream. This concentration gradient and the lipophilic nature of the rumen epithelium allow for the passive diffusion of non-ionized VFAs from the rumen into the rumen epithelium [18,36,37]. Once the non-ionized VFAs cross the cell membrane, they dissociate into anions and H+, which is the major source of hydrogen ions that leave the rumen epithelium (Figure 3). These hydrogen ions are recycled via a process called sodium/hydrogen exchange. Simultaneously, a bicarbonate ion exchange process occurs, which facilitates the absorption of ionized VFAs from the rumen into the rumen epithelium [18,31]. Bicarbonate ions are produced by the rumen epithelium in a considerable amount [38,39]. This process is important for the absorption of less lipophilic VFAs (e.g., acetic acid) into the rumen epithelium and finally into the bloodstream [38]. After the VFAs enter the rumen epithelial cells, an extensive metabolism of them (Figure 3, discussed in more detail later) occurs [40]. The metabolites of VFAs are absorbed into the bloodstream through a carrier-mediated process facilitated by downregulated in adenoma (DRA, also known as SLC26A3 or CLCA), pendrin anion exchanger 1 (PAT1, also known as SLC26A6), and monocarboxylate transporter 1 (MCT1, also known as SLC16A1) in exchange for chloride and bicarbonate ions [38,39,41,42,43].
3.3. Metabolism of VFAs
According to the older theory, approximately 90% of butyric acid, 50% of propionic acid, and 30% of acetic acid absorbed from the rumen are metabolized in the rumen epithelium to provide nearly 70% of the energy needs of a ruminant [22,40]. However, more recent work suggests that the extent of metabolism of acetic acid and propionic acid, particularly the latter, in the rumen epithelium might have been overestimated [40]. Acetic acid is the least metabolized VFA in the rumen epithelium, and partly because of this, it accounts for more than 90% of total VFAs in the arterial blood [22].
In the rumen epithelium, butyric acid and acetic acid are metabolized to the ketone bodies acetoacetic acid (AcAc) and β-hydroxybutyric acid (BHB) through ketogenesis (Figure 3) [4,44,45]. The 3-hydroxy-3-methylglutaryl-coA synthase, which is a rate-limiting enzyme in ketogenesis, is highly expressed in the adult but absent in the young rumen epithelium [46]. The rumen epithelium exhibits greater ketogenic activity with butyric acid than does the liver, which is the major site of ketogenesis in non-ruminant animals [45,47]. Ketone bodies derived from butyric acid and acetic acid are then used to generate energy in extrahepatic tissues, including the heart, skeletal muscle, adipose tissue, kidney, and mammary gland [40,48].
A trace of butyric acid, which is neither metabolized by the rumen epithelium nor by the liver, enters the adipose tissue or the mammary gland from the bloodstream, and there, is rapidly oxidized for lipogenesis [22]. In the rumen epithelium, a small proportion (about 5%) of propionic acid is converted to lactic acid and pyruvic acid [4,49] (Figure 3). The remaining propionic acid is used by hepatic tissue as a substrate for gluconeogenesis [3]. Most of the acetic acid absorbed from the rumen is not metabolized by the rumen epithelium or the liver and is instead utilized by peripheral tissues [22,50,51]. In the adipose tissue and mammary gland, acetic acid is a major substrate for de novo synthesis of fatty acids [52].
3.4. Barrier Function
The rumen epithelium is subject to challenges from rumen microbes, toxins produced by rumen microbes, changes in the ruminal pH, and many other potential adversities. The multi-layered rumen epithelium functions to efficiently maintain the separation between the ruminal lumen and blood circulation. This separation is achieved through tightly controlled selective absorption mechanisms within cells and intercellular junctions between cells that restrict the paracellular passage of ruminal pathogens and other insults into the rumen epithelium [38,53]. Besides the intercellular junctions, the immune cells within and beneath the rumen epithelium provide an “immune barrier” that protects the rumen epithelium from pernicious stimuli [54,55].
Rumen epithelial integrity and barrier function are achieved mainly through the intercellular junctions in stratum granulosum and stratum spinosum, which include tight junctions (TJ), adherens junctions (AJ), and desmosomes [56,57]. Tight junctions provide barrier functions, intercellular communication, cell adhesion, and selective permeability; adherens junctions and desmosomes provide strong cell adhesion and mechanical anchoring between the cells [58,59]. The keratinized layer present at the upper tip of the rumen papillae also protects the rumen against abrasion caused by feed particles and changes in rumen pH [55].
Various tight junction proteins have been identified in epithelial cells of different species, including claudins, occludin, cingulin, tricellulin, junctional adhesion molecule, and MarvelD3, which form semipermeable transmembrane along with the adapter protein and zona occludens 1, 2, and 3 [57,60,61,62]. Zona occludens link tight junction transmembrane proteins to actin cytoskeleton and help maintain the structure and functioning of tight junctions [58]. In rumen tissue and rumen epithelial cells, researchers have identified the protein presence of claudins 1, 4, and 7, and occludin [53]. Transcriptomic analyses have revealed the mRNA expression of claudins 1, 3, 4, 7, 12, and 23, cingulin, junctional adhesion molecules 2 and 3, occludin, and zona occludens 1 and 3 in rumen epithelial cells [7,63].
At low rumen pH, tight junctions in the rumen epithelium may lose their integrity or function, allowing harmful microbes to enter the bloodstream [56,64]. To combat this, activation of toll-like receptors by rumen microbes and their products, such as lipopolysaccharides (LPS), induces the production of cytokines, which trigger immune cells such as CD4+ T cells to resist the noxious stimuli [65,66] and the epithelial cells to increase the expression of zona occludens 1 and 2 [67]. Probiotics such as Lactobacillus plantarum MB452 also enhance the barrier function of the rumen epithelium by increasing the expression of tight junction genes [68]. Heat stress affects the rumen epithelial barrier negatively to some extent but does not completely disrupt it, due to the upregulation of heat shock proteins [69]. In essence, the strength of the physical barrier of the rumen is intricately linked with the microbes and immune cells in the rumen.
4. Prenatal Development of the Rumen
During bovine gastrointestinal development, which commences around the 29th day of gestation, the caudal region of the esophagus undergoes enlargement, leading to the formation of the definitive stomach compartments, although debates persist regarding the precise origins of these compartments, with evidence leaning toward development from a fusiform primordium [70,71]. By the 6th week of gestation, the rumen undergoes significant expansion and migrates to the left cephalic region, accompanied by the formation of blind sacs and spirally twisting vestibule, the narrow caudal portion of the rumen [70,71]. By around the 58th day of gestation, the dorsal and ventral blind sacs of the rumen expand, forming small evaginations of the main rumen, which later assume their definitive caudal position with a transverse notch, known as the caudal groove of the rumen, between them. During this period before birth, the caudal part of the vestibule develops into atrium ventriculi, while the cranial region merges with the dorsal sac of the rumen in adult animals [71]. At the end of the first trimester, the rumen and reticulum occupy the room between the 7th thoracic vertebrae and the 4th lumbar vertebrae. They extend consistently between the 3rd trimester and the birth to eventually occupy the abdominal cavity between the 9th thoracic and the 3rd lumbar vertebrae [72]. This period also witnesses a surge in cellular proliferation and differentiation within the rumen epithelium, resulting in a marked increase in cell layers [71,73]. Fully developed conical papillae, while still fused with the epithelium, are observable by the end of the pregnancy. By this point, the rumen epithelium already comprises the four layers (Figure 2) [73,74,75].
5. Postnatal Development of the Rumen
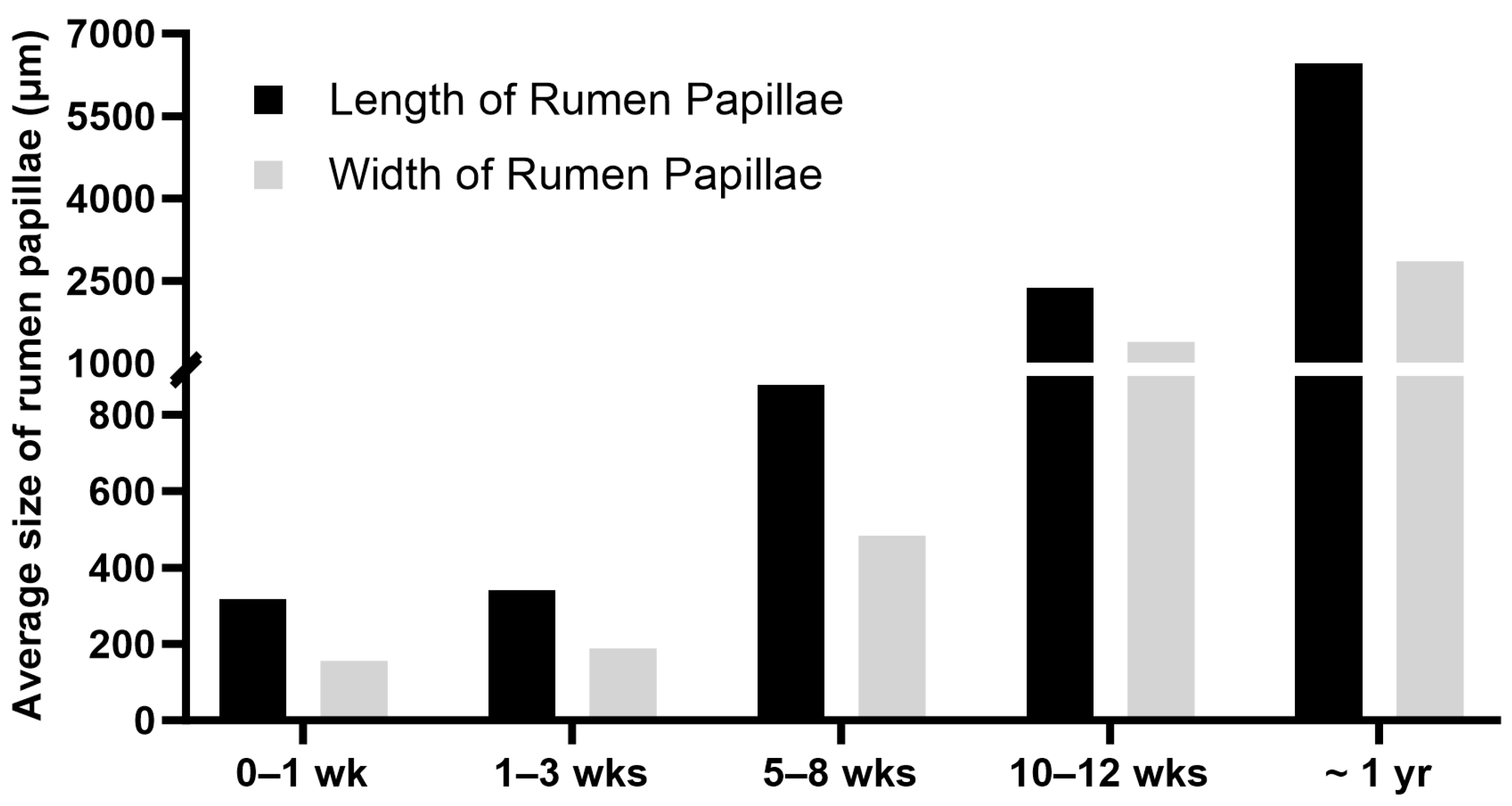
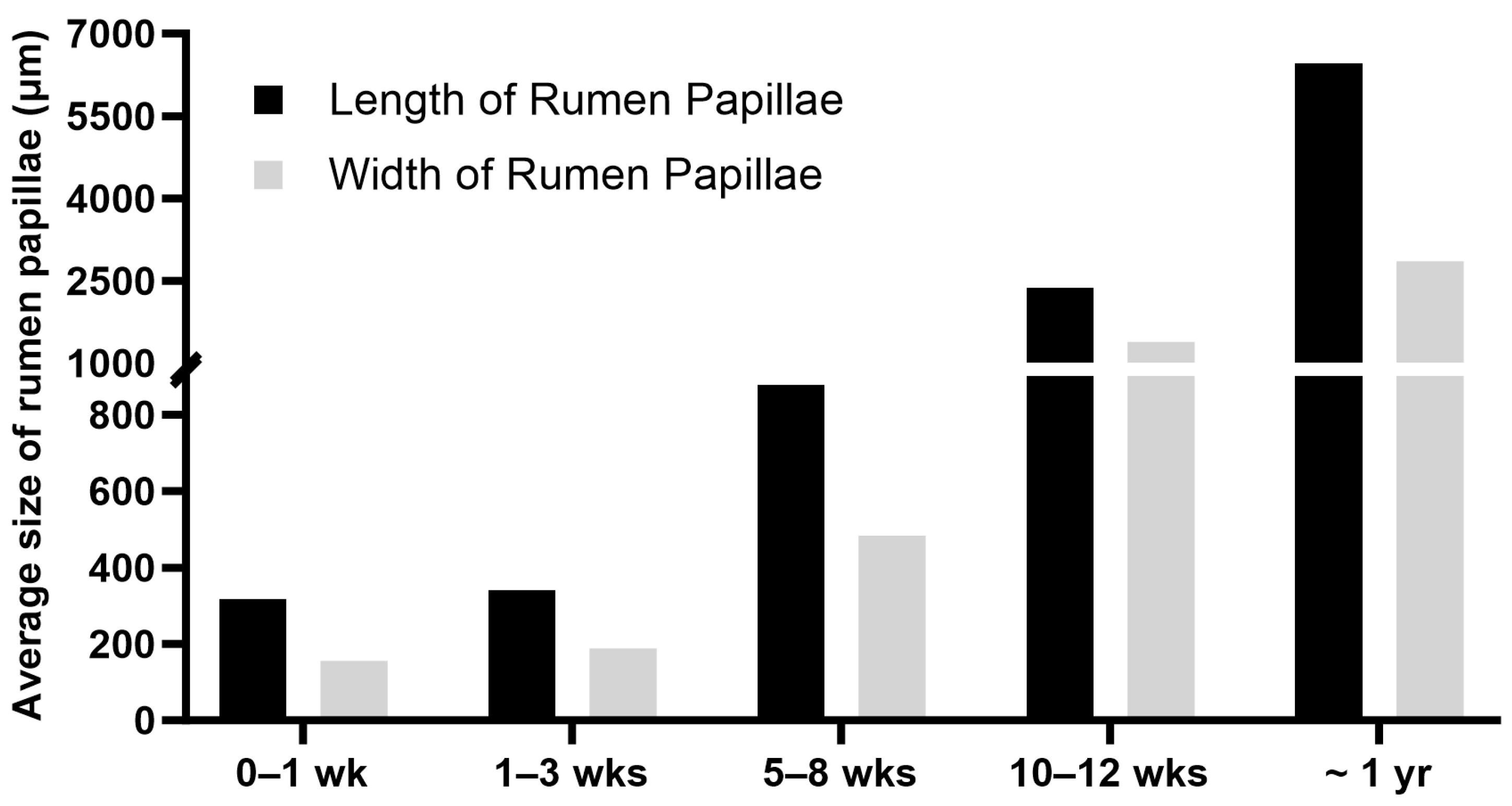
The rumen of a newborn calf is underdeveloped and not functional, and the digestive functions of the stomach are primarily performed by the abomasum, the largest part of the stomach at birth [4,6]. In pre-weaning ruminants, the ingested colostrum, milk, or milk replacer bypasses the rumen, reticulum, and omasum and goes directly from the esophagus to the abomasum through the esophageal groove [4,6,76]. The esophageal groove, also known as the reticular groove, is formed by the muscular fold of the reticulum and is a reflex response triggered by suckling [77]. The rumen epithelium of a newborn calf lacks keratinization and ketogenic activity, and the rumen of a newborn calf lacks an established microbial community [4]. The primary energy source for the neonatal rumen epithelium is derived from nutrients absorbed in the small intestine. Glucose, along with fatty acids, is considered a key energy substrate for the immature rumen epithelial tissue, especially before active fermentation begins in the rumen [78]. During postnatal life, the rumen undergoes significant changes, including increased muscle mass, epithelial growth, visible papillae formation, and colonization of microbial communities [20]. The length and width of rumen papillae increase with ages from birth to one year of age in cattle (Figure 4). The size of rumen papillae and the thickness of the rumen wall are often used as indicators of the degree of rumen development [79]. As the rumen matures, it gains the ability to metabolize VFAs and use VFAs instead of glucose as the major source of energy [80].
As a result of postnatal development, the proportions of the reticulorumen, omasum, and abomasum in the entire stomach undergo significant changes, shifting from 38%, 13%, and 49% at birth to 61%, 13%, and 25% at 8 weeks of age, and reaching 67%, 18%, and 15% at 12–16 weeks of age [76,81,82].
{kind=link}
{kind=link}
{kind=link}
{kind=link}
6. Factors Affecting the Postnatal Development of the Rumen
6.1. Diet
When fed exclusively milk, calves had limited rumen development; however, increased intake of solid feed accelerated rumen development in calves [91]. Rumen papillae grew in weaned calves but not in sucking and milk-continued calves [92]. Calves weaned earlier with higher solid feed intake showed more advanced rumen epithelial, papillary, and muscular growth in the rumen [6,93,94]. Lambs raised on a milk replacer alone showed limited metabolic maturation, while those weaned onto solid feed demonstrated increased VFA concentrations in the rumen and larger rumen papillae [91]. The proliferation rate of cells in the stratum basale was higher in calves fed milk replacer combined with a starter diet than in calves fed milk replacer alone [94]. All these data support the stimulatory effect of solid feed on rumen development in calves.
As mentioned earlier, when milk or milk replacer is drunk by a calf, it bypasses the rumen through the esophageal groove and directly enters the abomasum. Direct infusion of milk into the rumen promoted the growth of rumen papillae in calves, probably due to the availability of substrate for fermentation in the rumen [93]. However, it has also been reported that the introduction of liquid feed into the rumen within the first two weeks of birth can cause defective fermentation and, as a result, digestive disturbances such as ruminal acidosis and bloat occur, which can sometimes be fatal [77,95].
When inert substances such as nylon bristles, plastic tubes, and wood shavings, which are incapable of undergoing fermentation, were directly inserted into the rumen, it resulted in effective muscular growth in the rumen wall but no significant growth of rumen papillae [6,93]. The surface area of rumen papillae increased in a dose-dependent manner when dairy cows were fed different doses of rumen-fermentable organic matter during lactation and dry periods [96]. These findings suggest that the stimulatory effect of solid feed on rumen papillary growth is primarily attributed to the presence of active fermentation in the rumen, while the effect of solid feed on the growth of rumen muscle is probably due to the physical form of diet activating the stretch receptors in the muscular wall of the rumen [76,97,98].
6.2. Volatile Fatty Acids
As calves consume solid feed, rumen fermentation begins, and, as a result, rumen concentrations of VFAs rise. It has been widely believed that VFAs, particularly, butyric acid, mediate the effect of solid feed intake on rumen development [76,97,99]. Supplementation of sodium butyrate in calf starter promoted rumen development in newborn calves [85]. Intraruminal infusion of sodium butyrate increased the length of rumen papillae, the thickness of the stratum corneum, and the overall size of the rumen epithelium [100]. Sheep with rapid infusion of butyric acid showed greater mitotic activity in the rumen epithelium than sheep with slower infusion of butyric acid [101]. The expression of genes responsible for apoptosis, Caspase-3, and Bax, were significantly reduced in lambs fed sodium butyrate [100]. Volatile fatty acids stimulate rumen epithelial growth by increasing cell proliferation while reducing apoptosis [102,103].
VFAs influence the proliferation and apoptosis of rumen epithelial cells through various mechanisms. Dietary supplementation of ketone bodies, particularly β-hydroxybutyric acid (BHB), enhanced the growth of rumen epithelium [104], suggesting that butyric acid stimulates the growth of rumen epithelium through its metabolite BHB. Butyric acid and propionic acid might stimulate rumen papillary proliferation also by increasing blood flow through the rumen wall [76,105]. Butyrate can function as a histone deacetylase inhibitor to increase histone acetylation and thereby gene expression. There was a correlation between the production of acetic acid and butyric acid and the expressions of genes including MAPK1, PIK3CB, TNFSF1, ITGA6, SNAI2, SAV1, and DLG in rumen epithelium, all of which are related to cell growth [100]. Butyric acid has been shown to increase the expression of CCND1, CDK4, and PPARA, genes that promote cell cycle progression and lipid metabolism [97,106,107]. Thus, butyric acid might stimulate rumen epithelial growth by directly affecting the expression of genes involved in cell proliferation and metabolism through its histone deacetylase inhibitory activity.
6.3. Rumen Microbiota
As discussed above, the growth of rumen epithelium in postnatal ruminants is primarily driven by active fermentation and the production of VFAs within the rumen. Rumen fermentation and production of VFAs are facilitated by a diverse array of microflora consisting of bacteria, fungi, protozoa, and archaea, which colonize the rumen from maternal contact, environmental exposure, and feed intake [1,23,108]. These microorganisms perform fibrolytic, amylolytic, cellulolytic, and proteolytic activities [109]. The establishment of rumen microbiota is influenced by both host genetics and external environmental factors. The colonization of the rumen by microbiota can be observed as early as the first day of birth, transitioning from aerobic to facultative anaerobic and finally to strictly anaerobic bacteria as the animal matures [110,111].
The diverse microbial consortium operates cooperatively, fostering a conducive environment for fermentation and digestion, thereby influencing rumen growth. A correlation exists between ruminal VFA production and bacterial population [112]. Additionally, there is a correlation between the relative abundance of microbial communities and the expression of genes involved in various metabolic functions and molecular processes in rumen tissue [87]. Various taxa present in the rumen are responsible for the production of different enzymes that mediate the degradation of complex dietary materials and plant polysaccharides such as starch to glucose and finally VFAs [113]. Microflora in the rumen also interact with the host’s immune system, modulating various pathways to strengthen it [54,114]. Furthermore, the interaction between rumen microflora and the host contributes significantly to the regulation of the rumen’s barrier function and pH levels [54]. Some rumen bacteria have been found to digest keratinized epithelial cells, which promotes the growth of new cells in the rumen epithelium [115].
6.4. Hormones
Hormones and growth factors have significant effects on the growth and body composition of ruminants, and it would be unwise to disregard their involvement in rumen development and maturation. Insulin has long been suspected to mediate the stimulatory effect of VFAs on the proliferation of rumen epithelial cells in vivo [116]. This notion is supported by multiple lines of evidence. The promoting effect of butyric acid and propionic acid on rumen epithelial growth was associated with improved insulin sensitivity [117]. Infusion of insulin significantly stimulated cell proliferation in the rumen epithelium [116]. Treatment of primary rumen epithelial cells with insulin arrested the inhibitory effect of butyric acid on cell proliferation in vitro and increased the DNA synthesis of the cultured cells in a dose-dependent manner [118]. Nevertheless, not all studies support the role of insulin in mediating the effect of VFAs on rumen growth. For example, Wang et al. found that supplementation of isobutyric acid stimulated rumen development but was associated with decreased blood concentration of insulin in calves [119].
Calves weaned earlier (meaning transitioning to solid feed earlier) showed significantly more growth of the rumen and had higher concentrations of plasma insulin-like growth factor I (IGF-I) than calves weaned later [120]. Similar results were demonstrated in sheep [121]. The stimulatory effect of high-concentrate diets on the growth of rumen papillae was associated with increased systemic IGF-I concentration and increased binding of IGF-I to rumen epithelial IGF-I receptors in goats [121]. Supplementation of isobutyric acid increased rumen development and ketogenesis in calves in a dose-dependent manner, and these increases were accompanied by increased blood concentrations of IGF-I [119]. IGF-I stimulated the proliferation of rumen epithelial cells in vitro [121,122]. These observations suggest that intake of solid feed or butyric acid might stimulate rumen growth by circulating IGF-I [123]. However, short-term intraruminal infusion of butyric acid in castrated bulls increased the size of rumen papillae without increasing the concentration of plasma IGF-I [124], arguing against the role of IGF-I in mediating the effect of solid feed intake or VFAs on the growth of rumen epithelium.
Leptin is a hormone predominantly produced by fat and plays a central role in regulating appetite and energy balance [125]. Interestingly, leptin mRNA was expressed at elevated levels in the rumen and other stomach chambers in calves, indicating a potential involvement of leptin in the initiation of rumen development [126]. Ghrelin, a peptide hormone, is synthesized in the stomach and regulates growth hormone release, appetite, gastrointestinal motility, nutrient absorption, and energy balance in monogastric animals [127,128]. In lambs, ghrelin mRNA is abundantly expressed in the rumen [128], and this suggests a possible paracrine or autocrine action of ghrelin in the rumen. However, the role of ghrelin in postnatal rumen growth is not supported by the data indicating that rumen expression of ghrelin was not different between calves and adult cattle [126].
Epidermal growth factor (EGF) is a growth factor known for its function in regulating epithelial cell proliferation, morphogenesis, and tight junctions in the intestines [97,129]. Proliferation of rumen epithelial cells was stimulated by EGF in vitro [130]. EGF increased mRNA expression of tight junction proteins claudin 1 and occludin in the rumen [129]. These results support the potential role of EGF in regulating epithelial cell proliferation and tight junctions in the rumen.
Transcriptomic analyses suggested that transforming growth factor-β might be another hormone mediating the growth of rumen epithelium from pre-weaned to post-weaned calves [131]. Similarly, integrated analysis of differentially expressed long non-coding RNAs and mRNAs revealed thyroid hormone as another candidate hormone that mediates rumen development in postnatal animals [132,133].
6.5. Weaning
Weaning refers to the transition of young ruminants from dependency on maternal milk or milk replacer to consuming solid feed. Most calves start to consume solid feed after two weeks of age, and the intake of solid feed increases in an inverse relationship with the intake of milk [5,134]. Natural weaning starts when milk alone is insufficient to supply the daily requirements of the calves. This process is accompanied by a gradual reduction in the maternal–filial bond and rejection of dam to nurse the calves, forcing the calves to search for other sources of food [135].
While calves are naturally weaned at the age of six to nine months, it has become a common practice in cattle production to separate calves from dams earlier, as early as at the age of 6 weeks [136,137,138]. One of the benefits of early weaning is early maturation of the rumen [137,138,139,140]. However, as discussed earlier, early weaning could increase the risk of calves suffering from acidosis and other digestive disorders in later life. The development of the rumen following weaning is primarily attributable to the increased availability of substrate for fermentation, leading to the production of VFAs within the rumen. The pace of weaning also influences the development of the rumen. A recent study found that calves weaned gradually showed greater feed intake, higher body weight gain, increased immune functions, higher serum concentrations of β-hydroxybutyric acid, whereas calves weaned abruptly showed higher serum concentrations of non-esterified fatty acids and higher expression of inflammation-related genes such as IL-1β, IL-6, TNF-α, NF-κB, and ICAM in the liver or jejunum, indicating a more functional rumen in the former than the latter [141,142]. Weaning also changes the expression of genes related to rumen epithelial cell proliferation, molecular transport, and energy metabolism [138]. It is advocated that calves are gradually weaned over a period of two to three weeks at around three months of life for smooth transitioning from pre-ruminants to ruminants [138,140,143]. Interested readers can refer to previous reviews by Khan et al., Kertz et al., and Ghaffari and Kertz for recommended forms and amounts of solid feed intake for calves [134,144,145].
6.6. Other Factors
Feeding pasteurized colostrum and milk to neonatal calves has a significant positive impact on their overall health status; calves fed pasteurized colostrum and milk products developed fewer or no respiratory and digestive pathologies compared to those fed unpasteurized products [146,147]. However, feeding pasteurized milk and milk products has been associated with lower rumen mass compared to calves fed whole milk or milk replacers during the weaning phase (60 to 180 days of age) [148]. Feeding pasteurized colostrum and milk products to calves had conflicting effects on the composition of the ruminal microbiome [147].
Probiotics, prebiotics, synbiotics, growth promoters, and digestive enzymes fed to ruminants can affect rumen development indirectly by affecting the digestion of feed and production of VFAs in the rumen [76,84,149]. Supplementing calves’ diet with microbes or probiotics such as Bacillus licheniformis, Saccharomyces cerevisiae, Bacillus subtilis natto, Lactobacillus plantarum, and live or hydrolyzed yeast has the potential to enhance rumen development by influencing the microbial community and digestion in the rumen [150,151,152]. The addition of inulin to whole milk of calves increased the length and width of the rumen papillae [84]. Antibiotics added to feed to increase the productivity of animals could modulate the development and function of the rumen [153,154]. Including pectinase in the diet of lambs increased the production of major VFAs in the rumen, suggesting its effect on rumen development or function in young ruminants [155].
The development of the rumen is influenced by various pathological conditions, including digestive pathologies. Experimental infection of pre-weaned dairy calves with salmonellosis, a major causative agent of digestive problems in the calves, has been shown to negatively affect the growth of ruminal papillae [156]. Similarly, digestive issues induced by the higher intake of lactose, either in milk or in milk replacers in young calves, result in delayed or lesser growth of the rumen compared to those without digestive problems [157]. In adult steers, gastrointestinal toxicosis or acidosis has been observed to cause serious degeneration of the ruminal epithelium; such pathologies can alter the acid-base balance of the rumen, disrupt its microbial ecosystem, and impair its absorptive capacity, ultimately inhibiting rumen development [34,158,159].
7. Genes Involved in Rumen Growth and Development
Structural and functional development of the rumen can be eventually attributed to changes in gene expression in the rumen. The expression of 6 transcription factors (OTX1, SOX21, HOXC8, SOX2, TP63, and PPARG) and 16 ruminant-specific conserved non-coding elements (RSCNEs) contributes significantly to differentiating the rumen from the esophagus [160]. Functioning as enhancers or silencers, RSCNEs contribute to pre- and post-transcriptional regulation, modulation of genes associated with morphological and physiological traits, and chromatin architecture, and play a crucial role in shaping the distinctive features of the rumen [132,161,162]. Zhang et al. identified 6048 differentially expressed genes (DEGs) in the rumen among neonatal, young, and adult cattle [163]. In experiments conducted by other groups, 2048 rumen DEGs were identified between 1-day- and 56-day-old calves, and 871 rumen DEGs were found between suckling and weaned calves [80,164]. Micro RNAs (miRNAs) are small noncoding RNAs that play an important role in post-transcriptional regulation of gene expression. A total of 132 miRNAs were differentially expressed in the rumen between the pre- and post-weaning calves [161].
It is safe to say that the development and functional maturation of the rumen relies on the coordinated expression of various genes involved in diverse molecular functions and biological processes. Genes such as CCNB1, CCNB2, IGF1, IGF2, HMGCL, BDH1, ACAT1, and CREBP could play important roles in driving rumen development because they are known to function in cell proliferation and fatty acid metabolism, and they were expressed at higher levels in the adult cattle than calf rumen [163]. Genes encoding the IGF binding proteins IGFBP2, IGFBP3, and IGFBP6 were expressed at higher levels in the rumen of suckling and milk-continued calves than that of weaned and solid feed-fed calves [92]. Furthermore, the expression levels of IGFBP2, IGFBP3, and IGFBP6 negatively correlated with the growth of rumen papillae [92]. IGFBPs in general function to block the binding of IGF-I to its receptor. These gene expression data support the hypothesis that reduced expression of IGFBP genes in the rumen contributes to solid feed-induced growth of rumen papillae in post-weaning animals, perhaps by increasing the mitogenic action of IGF-I on rumen epithelial cells. Interestingly, in at least one study, IGFBP5 was found to be upregulated during the growth of rumen papillae [164]. Perhaps IGFBP5 has the ability to regulate the growth of rumen papillae independent of IGF-I.
Members of the ruminant-specific PRD-SPRRII gene family were expressed at significantly higher levels in the mature than immature rumen [165]. Genes such as ACTG2, ILK, and KIF4A, which are involved in cell skeleton regulation, cell adhesions, and cell organization, were among the genes expressed at higher levels in adult rather than young ruminants [97,166,167]. Similarly, FABP7, FABP3, ILK, PDGFA, HMGCS2, and AKR1C1 were differentially expressed between the immature and mature rumen [87,168]. Rumen expression of FABP3 was downregulated in weaned calves compared to pre-weaning calves [169]. The upregulation of FABP3 is linked with inhibition of cell growth and proliferation, suggesting a possible role for the FABP3 gene in inducing the proliferation of rumen epithelial cells in post-weaning calves [168,169]. MCT1 (SLC16A1), PAT1 (SLC26A6), ACAT1, HMGCS2, HMGCL, ACAA1, ACAA2, CA4, PPARA, PPARG, SLC26A3, and other members of the SLC family are highly expressed in the adult rumen [80,112,170]. These genes participate in rumen development and functional maturation by facilitating the transportation of VFAs from the ruminal lumen to the rumen epithelial cells and to the bloodstream, or by facilitating beta-oxidation and ketogenesis of VFAs in the rumen epithelium [164,169].
Members of the claudin (CLDN) family, CLDN1, CLDN4, and CLDN7, are highly expressed in the rumen [53], and they may be of prime importance in maintaining the integrity of the rumen epithelium because they encode tight junction proteins [171]. Similarly, genes coding for tight junction-associated proteins MarvelD3, occludin, and tricellulin likely play a role in maintaining the structural integrity and epithelial barrier function of the rumen too [61]. Genes DMRT2, WDR66, COL7A1, EVPL, KRT14, F2RL1, TMPRSS13, and TMPRSS11A participate in the cell junction biological process, are upregulated in the adult rumen [80] and, therefore, may also contribute to the barrier function of the adult rumen.
CEBPB, S100A9, CCL20, CXCL8, TNF, NFKBIA, LBP, PGLYRP2, MUC1, SOCS1, SOCS3, and CCDC3 activate the innate immunity in the rumen [80]. BPIFA1, part of the BPI-like protein family, plays a role in the innate immunity of the epithelial mucosa [97]. These genes are likely involved in both the development and health of the rumen.
8. Conclusions
The proper development of the rumen, particularly the rumen epithelium and papillae, is crucial for the physiology and production of cattle and other ruminants. While the rumen of newborn calves shares anatomical and histological similarities with that of adult cattle, the rumen undergoes significant development during postnatal life, especially after weaning. The rumen encompasses remarkable physical and functional changes in postnatal animals. Physically, the rumen grows in mass and size; functionally, the rumen gains the ability to ferment feed into VFAs (because of colonization of microbes and intake of solid feed), absorb VFAs, metabolize VFAs, and protect against potential insults from the fermentation. Among the various factors influencing postnatal rumen growth and functional maturation, a solid diet stands out as the most significant. Introducing solid feed to the rumen is essential for its growth and functional development in young ruminants. Solid feed stimulates rumen growth and functional development through VFAs, especially butyric acid, produced by rumen fermentation. Butyric acid and other VFAs stimulate rumen growth and functional development through both direct and indirect mechanisms. In direct mechanisms, butyric acid and other VFAs stimulate cell proliferation and metabolism in rumen epithelial cells by altering the expression of genes critical for these processes. In indirect mechanisms, butyric acid and other VFAs stimulate cell proliferation, and metabolism by increasing the concentration and/or action of circulating insulin-like growth factor-I (IGF-I), insulin, and perhaps other hormones and growth factors on the rumen. While these mechanisms have been proposed repeatedly in the literature, they await validation through carefully designed studies. A deeper understanding of how diet and other factors affect rumen development, growth, and function at the cellular, molecular, genomic, and epigenomic levels may help devise novel or more effective management practices to enhance rumen development and functional maturation in cattle and other agriculturally important ruminants.
Author Contributions
Conceptualization, H.J.; writing—original draft preparation, B.P.; writing—review and editing, H.J.; funding acquisition, H.J. All authors have read and agreed to the published version of the manuscript.
Funding
B.P. was supported by a Pratt Fellowship from the College of Agriculture and Life Science, Virginia Tech.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Dehority, B.A. Gastrointestinal Tracts of Herbivores, Particularly the Ruminant: Anatomy, Physiology and Microbial Digestion of Plants. J. Appl. Anim. Res. 2002, 21, 145–160. [Google Scholar] [CrossRef]
- Membrive, C.M.B. Anatomy and physiology of the rumen. In Rumenology; Springer: Cham, Switzerland, 2016; pp. 1–38. [Google Scholar] [CrossRef]
- Harfoot, C.G. Anatomy, physiology and microbiology of the ruminant digestive tract. In Progress in the Chemistry of Fats and Other Lipids; Pergamon Press, Ltd.: London, UK, 1978; Volume 17, pp. 1–19. [Google Scholar] [CrossRef]
- Baldwin, R.L.; Connor, E.E. Rumen Function and Development. In Veterinary Clinics of North America—Food Animal Practice; W.B. Saunders: Philadelphia, PA, USA, 2017; Volume 33, pp. 427–439. [Google Scholar]
- Drackley, J.K. Calf nutrition from birth to breeding. Vet. Clin. N. Am. Food Anim. Pract. 2008, 24, 55–86. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, R.L.; McLeod, K.R.; Klotz, J.L.; Heitmann, R.N. Rumen Development, Intestinal Growth and Hepatic Metabolism In The Pre- and Postweaning Ruminant. J. Dairy Sci. 2004, 87, E55–E65. [Google Scholar] [CrossRef]
- Ji, X.; Tong, H.; Settlage, R.; Yao, W.; Jiang, H. Establishment of a bovine rumen epithelial cell line. J. Anim. Sci. 2021, 99, skab273. [Google Scholar] [CrossRef] [PubMed]
- Dobson, M.J.; Brown, W.C.; Dobson, A.; Phillipson, A.T. A histological study of the organization of the rumen epithelium of sheep. Q. J. Exp. Physiol. Cogn. Med. Sci. 1956, 41, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.; Zhang, Y.H.; Zhang, W. Regulation of Intestinal Epithelial Cells Properties and Functions by Amino Acids. Biomed. Res. Int. 2018, 2018, 2819154. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, R.L.; Jesse, B.W. Technical note: Isolation and characterization of sheep ruminal epithelial cells. J. Anim. Sci. 1991, 69, 3603–3609. [Google Scholar] [CrossRef]
- Steele, M.A.; Penner, G.B.; Chaucheyras-Durand, F.; Guan, L.L. Development and physiology of the rumen and the lower gut: Targets for improving gut health. J. Dairy Sci. 2016, 99, 4955–4966. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.; Simmons, N.L. Functional organization of the bovine rumen epithelium. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R173–R181. [Google Scholar] [CrossRef]
- van Niekerk, J.K.; Middeldorp, M.; Guan, L.L.; Steele, M.A. Preweaning to postweaning rumen papillae structural growth, ruminal fermentation characteristics, and acute-phase proteins in calves. J. Dairy Sci. 2021, 104, 3632–3645. [Google Scholar] [CrossRef]
- Lavker, R.M.; Matoltsy, A.G. Formation of horny cells: The fate of cell organelles and differentiation products in ruminal epithelium. J. Cell Biol. 1970, 44, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Reece, W.O. Functional Anatomy and Physiology of Domestic Animals, 4th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009. [Google Scholar]
- Liang, Y.; Wang, S.; An, T.; Tarique, I.; Vistro, W.A.; Liu, Y.; Wang, Z.; Zhang, H.; Shi, Y.; Haseeb, A.; et al. Telocytes as a Novel Structural Component in the Muscle Layers of the Goat Rumen. Cell Transpl. 2019, 28, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, R.L.; Allison, M.J. Rumen metabolism. J. Anim. Sci. 1983, 57 (Suppl. S2), 461–477. [Google Scholar] [CrossRef]
- Dijkstra, J. Production and absorption of volatile fatty acids in the rumen. Livest. Prod. Sci. 1994, 39, 61–69. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Global Rumen Census, C.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Yin, Y.; Guo, C.; Liu, L.; Mao, S.; Zhu, W.; Liu, J. Transcriptomic analysis reveals the molecular mechanisms of rumen wall morphological and functional development induced by different solid diet introduction in a lamb model. J. Anim. Sci. Biotechnol. 2021, 12, 33. [Google Scholar] [CrossRef] [PubMed]
- Solomon, R.; Jami, E. Rumen protozoa: From background actors to featured role in microbiome research. Environ. Microbiol. Rep. 2021, 13, 45–49. [Google Scholar] [CrossRef]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef]
- Giger-Reverdin, S.; Domange, C.; Broudiscou, L.P.; Sauvant, D.; Berthelot, V. Rumen function in goats, an example of adaptive capacity. J. Dairy Res. 2020, 87, 45–51. [Google Scholar] [CrossRef]
- Jenkins, T.C. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Bach, A.; Calsamiglia, S.; Stern, M.D. Nitrogen metabolism in the rumen. J. Dairy Sci. 2005, 88 (Suppl. S1), E9–E21. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Kang, J.; Zhang, D. Microbial production of vitamin B(12): A review and future perspectives. Microb. Cell Fact. 2017, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Abdoun, K.; Stumpff, F.; Martens, H. Ammonia and urea transport across the rumen epithelium: A review. Anim. Health Res. Rev. 2006, 7, 43–59. [Google Scholar] [CrossRef]
- Li, Y.; Kreuzer, M.; Clayssen, Q.; Ebert, M.O.; Ruscheweyh, H.J.; Sunagawa, S.; Kunz, C.; Attwood, G.; Amelchanka, S.; Terranova, M. The rumen microbiome inhibits methane formation through dietary choline supplementation. Sci. Rep. 2021, 11, 21761. [Google Scholar] [CrossRef]
- Annison, E.F.; Bryden, W.L. Perspectives on ruminant nutrition and metabolism I. Metabolism in the rumen. Nutr. Res. Rev. 1998, 11, 173–198. [Google Scholar] [CrossRef]
- Beauchemin, K.A. Invited review: Current perspectives on eating and rumination activity in dairy cows. J. Dairy Sci. 2018, 101, 4762–4784. [Google Scholar] [CrossRef] [PubMed]
- Aschenbach, J.R.; Penner, G.B.; Stumpff, F.; Gabel, G. Ruminant Nutrition Symposium: Role of fermentation acid absorption in the regulation of ruminal pH. J. Anim. Sci. 2011, 89, 1092–1107. [Google Scholar] [CrossRef]
- Storm, A.C.; Kristensen, N.B. Effects of particle size and dry matter content of a total mixed ration on intraruminal equilibration and net portal flux of volatile fatty acids in lactating dairy cows. J. Dairy Sci. 2010, 93, 4223–4238. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Pathak, N.N. Fundamentals of Animal Nutrition; Springer: Singapore, 2021; pp. 219–246. [Google Scholar] [CrossRef]
- Gholizade, M.; Fayazi, J.; Zali, H.; Asgari, Y. Transcriptomic Changes in the Rumen Epithelium of Cattle after the Induction of Acidosis. Arch. Razi Inst. 2020, 75, 109–121. [Google Scholar] [CrossRef]
- Dobson, A. Blood flow and absorption from the rumen. Q. J. Exp. Physiol. 1984, 69, 599–606. [Google Scholar] [CrossRef]
- Sehested, J.; Diernaes, L.; Moller, P.D.; Skadhauge, E. Ruminal transport and metabolism of short-chain fatty acids (SCFA) in vitro: Effect of SCFA chain length and pH. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1999, 123, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Sehested, J.; Diernaes, L.; Moller, P.D.; Skadhauge, E. Transport of butyrate across the isolated bovine rumen epithelium--interaction with sodium, chloride and bicarbonate. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1999, 123, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Aschenbach, J.R.; Zebeli, Q.; Patra, A.K.; Greco, G.; Amasheh, S.; Penner, G.B. Symposium review: The importance of the ruminal epithelial barrier for a healthy and productive cow. J. Dairy Sci. 2019, 102, 1866–1882. [Google Scholar] [CrossRef] [PubMed]
- Bilk, S.; Huhn, K.; Honscha, K.U.; Pfannkuche, H.; Gabel, G. Bicarbonate exporting transporters in the ovine ruminal epithelium. J. Comp. Physiol. B 2005, 175, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Gabel, G.; Aschenbach, J.R.; Muller, F. Transfer of energy substrates across the ruminal epithelium: Implications and limitations. Anim. Health Res. Rev. 2002, 3, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Aschenbach, J.R.; Bilk, S.; Tadesse, G.; Stumpff, F.; Gabel, G. Bicarbonate-dependent and bicarbonate-independent mechanisms contribute to nondiffusive uptake of acetate in the ruminal epithelium of sheep. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1098–G1107. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.; Gatherar, I.; Haslam, I.; Glanville, M.; Simmons, N.L. Expression and localization of monocarboxylate transporters and sodium/proton exchangers in bovine rumen epithelium. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R997–R1007. [Google Scholar] [CrossRef] [PubMed]
- Muller, F.; Aschenbach, J.R.; Gabel, G. Role of Na+/H+ exchange and HCO3− transport in pHi recovery from intracellular acid load in cultured epithelial cells of sheep rumen. J. Comp. Physiol. B 2000, 170, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Beck, U.; Emmanuel, B.; Giesecke, D. The ketogenic effect of glucose in rumen epithelium of ovine (Ovis aries) and bovine (Bos taurus) origin. Comp. Biochem. Physiol. B 1984, 77, 517–521. [Google Scholar] [CrossRef]
- Giesecke, D.; Beck, U.; Wiesmayr, S.; Stangassinger, M. The effect of rumen epithelial development on metabolic activities and ketogenesis by the tissue in vitro. Comp. Biochem. Physiol. B 1979, 62, 459–463. [Google Scholar] [CrossRef]
- Lane, M.A.; Baldwin, R.L.; Jesse, B.W. Developmental changes in ketogenic enzyme gene expression during sheep rumen development. J. Anim. Sci. 2002, 80, 1538–1544. [Google Scholar] [CrossRef] [PubMed]
- Bush, R.S.; Milligan, L.P.; Krishnamurti, C.R. Effects of Propionate on Ketogenesis from Butyrate by Bovine Tissues. Can. J. Anim. Sci. 1970, 50, 210. [Google Scholar] [CrossRef]
- Goosen, P.C. Metabolism in rumen epithelium oxidation of substrates and formation of ketone bodies by pieces of rumen epithelium. Z. Tierphysiol. Tierernahr. Futtermittelkunde 1976, 37, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Weekes, T.E. The in vitro metabolism of propionate and glucose by the rumen epithelium. Comp. Biochem. Physiol. B 1974, 49, 393–406. [Google Scholar] [CrossRef]
- Urrutia, N.; Bomberger, R.; Matamoros, C.; Harvatine, K.J. Effect of dietary supplementation of sodium acetate and calcium butyrate on milk fat synthesis in lactating dairy cows. J. Dairy Sci. 2019, 102, 5172–5181. [Google Scholar] [CrossRef] [PubMed]
- Black, A.L.; Luick, J.; Moller, F.; Anand, R.S. Pyruvate and propionate metabolism in lactating cows. Effect of butyrate on pyruvate metabolism. J. Biol. Chem. 1966, 241, 5233–5237. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Zhang, Y.; Zhang, H.; Sun, Y.; Mao, Y.; Yang, Z.; Li, M. Transcriptional regulation of milk fat synthesis in dairy cattle. J. Funct. Foods 2022, 96, 105208. [Google Scholar] [CrossRef]
- Stumpff, F.; Georgi, M.I.; Mundhenk, L.; Rabbani, I.; Fromm, M.; Martens, H.; Gunzel, D. Sheep rumen and omasum primary cultures and source epithelia: Barrier function aligns with expression of tight junction proteins. J. Exp. Biol. 2011, 214, 2871–2882. [Google Scholar] [CrossRef] [PubMed]
- Malago, J.J. Contribution of microbiota to the intestinal physicochemical barrier. Benef. Microbes 2015, 6, 295–311. [Google Scholar] [CrossRef]
- Shen, H.; Xu, Z.; Shen, Z.; Lu, Z. The Regulation of Ruminal Short-Chain Fatty Acids on the Functions of Rumen Barriers. Front. Physiol. 2019, 10, 1305. [Google Scholar] [CrossRef]
- Liu, J.H.; Xu, T.T.; Liu, Y.J.; Zhu, W.Y.; Mao, S.Y. A high-grain diet causes massive disruption of ruminal epithelial tight junctions in goats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R232–R241. [Google Scholar] [CrossRef]
- Naydenov, N.G.; Hopkins, A.M.; Ivanov, A.I. c-Jun N-terminal kinase mediates disassembly of apical junctions in model intestinal epithelia. Cell Cycle 2009, 8, 2110–2121. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Takai, Y. Recent advances in understanding tight junctions. Fac. Rev. 2021, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.A.; Uppada, S.; Achkar, I.W.; Hashem, S.; Yadav, S.K.; Shanmugakonar, M.; Al-Naemi, H.A.; Haris, M.; Uddin, S. Tight Junction Proteins and Signaling Pathways in Cancer and Inflammation: A Functional Crosstalk. Front. Physiol. 2018, 9, 1942. [Google Scholar] [CrossRef]
- Mineta, K.; Yamamoto, Y.; Yamazaki, Y.; Tanaka, H.; Tada, Y.; Saito, K.; Tamura, A.; Igarashi, M.; Endo, T.; Takeuchi, K.; et al. Predicted expansion of the claudin multigene family. FEBS Lett. 2011, 585, 606–612. [Google Scholar] [CrossRef]
- Raleigh, D.R.; Marchiando, A.M.; Zhang, Y.; Shen, L.; Sasaki, H.; Wang, Y.; Long, M.; Turner, J.R. Tight junction-associated MARVEL proteins marveld3, tricellulin, and occludin have distinct but overlapping functions. Mol. Biol. Cell 2010, 21, 1200–1213. [Google Scholar] [CrossRef]
- Zhang, K.; Meng, M.; Gao, L.; Tu, Y.; Bai, Y. Rumen-derived lipopolysaccharide induced ruminal epithelium barrier damage in goats fed a high-concentrate diet. Microb. Pathog. 2019, 131, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, R.L.t.; Wu, S.; Li, W.; Li, C.; Bequette, B.J.; Li, R.W. Quantification of Transcriptome Responses of the Rumen Epithelium to Butyrate Infusion using RNA-seq Technology. Gene Regul. Syst. Bio 2012, 6, 67–80. [Google Scholar] [CrossRef]
- Meissner, S.; Hagen, F.; Deiner, C.; Gunzel, D.; Greco, G.; Shen, Z.; Aschenbach, J.R. Key role of short-chain fatty acids in epithelial barrier failure during ruminal acidosis. J. Dairy Sci. 2017, 100, 6662–6675. [Google Scholar] [CrossRef]
- Kandel, A.; Masello, M.; Xiao, Z. CD4+ T Cell Responses to Pathogens in Cattle. In Bovine Science—Challenges and Advances; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Spiljar, M.; Merkler, D.; Trajkovski, M. The Immune System Bridges the Gut Microbiota with Systemic Energy Homeostasis: Focus on TLRs, Mucosal Barrier, and SCFAs. Front. Immunol. 2017, 8, 1353. [Google Scholar] [CrossRef]
- Cario, E.; Gerken, G.; Podolsky, D.K. Toll-like receptor 2 enhances ZO-1-associated intestinal epithelial barrier integrity via protein kinase C. Gastroenterology 2004, 127, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Park, Z.; McCann, M.J.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010, 10, 316. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Gao, S.; Ding, J.; He, J.; Ma, L.; Bu, D. Effects of Heat Stress on the Ruminal Epithelial Barrier of Dairy Cows Revealed by Micromorphological Observation and Transcriptomic Analysis. Front. Genet. 2021, 12, 768209. [Google Scholar] [CrossRef] [PubMed]
- Kalenberg, C.A.; Stoffel, M.H. The embryonic development of the bovine stomach revisited. Anat. Histol. Embryol. 2020, 49, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Warner, E.D. The organogenesis and early histogenesis of the bovine stomach. Am. J. Anat. 1958, 102, 33–63. [Google Scholar] [CrossRef] [PubMed]
- Al Masri, S.; Reincke, R.; Huenigen, H.; Gemeinhardt, O.; Richardson, K.C.; Plendl, J. Computed tomography study of the fetal development of the dairy cow stomach complex. J. Dairy Sci. 2018, 101, 1719–1729. [Google Scholar] [CrossRef] [PubMed]
- Arias, J.L.; Cabrera, R.; Valencia, A. Observations on the histological development of the bovine rumen papillae. Morphological changes due to age. Anat. Histol. Embryol. 1978, 7, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.; Masot, J.; Franco, A.; Gazquez, A.; Redondo, E. Histomorphometric and immunohistochemical study of the goat rumen during prenatal development. Anat. Rec. 2012, 295, 776–785. [Google Scholar] [CrossRef]
- Stallcup, O.T.; Kreider, D.L.; Rakes, J.M. Histological development and histochemical localization of enzymes in rumen and reticulum in bovine fetuses. J. Anim. Sci. 1990, 68, 1773–1789. [Google Scholar] [CrossRef]
- Diao, Q.; Zhang, R.; Fu, T. Review of strategies to promote rumen development in calves. Animals 2019, 9, 490. [Google Scholar] [CrossRef]
- Kaba, T.; Abera, B.; Kassa, T. Esophageal groove dysfunction: A cause of ruminal bloat in newborn calves. BMC Vet. Res. 2018, 14, 276. [Google Scholar] [CrossRef] [PubMed]
- White, R.G.; Leng, R.A. Glucose metabolism in feeding and postabsorptive lambs and mature sheep. Comp. Biochem. Physiol. Part A Physiol. 1980, 67, 223–229. [Google Scholar] [CrossRef]
- Lesmeister, K.E.; Tozer, P.R.; Heinrichs, A.J. Development and analysis of a rumen tissue sampling procedure. J. Dairy Sci. 2004, 87, 1336–1344. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Li, Z.; Li, B.; Zhao, C.; Wang, Y.; Chen, Y.; Jiang, Y. Dynamics of rumen gene expression, microbiome colonization, and their interplay in goats. BMC Genom. 2021, 22, 288. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.L.; Drackley, J.K. The Development, Nutrition, and Management of the Young Calf; Iowa State University Press: Ames, IA, USA, 1998; pp. 1–50. [Google Scholar]
- Diao, Q.-y.; Zhang, R.; Tu, Y. Current research progresses on calf rearing and nutrition in China. J. Integr. Agric. 2017, 16, 2805–2814. [Google Scholar] [CrossRef]
- Alves Costa, N.; Pansani, A.P.; de Castro, C.H.; Basile Colugnati, D.; Xaxier, C.H.; Guimaraes, K.C.; Antas Rabelo, L.; Nunes-Souza, V.; Souza Caixeta, L.F.; Nassar Ferreira, R. Milk restriction or oligosaccharide supplementation in calves improves compensatory gain and digestive tract development without changing hormone levels. PLoS ONE 2019, 14, e0214626. [Google Scholar] [CrossRef] [PubMed]
- Arne, A.; Ilgaza, A. Prebiotic and synbiotic effect on rumen papilla length development and rumen pH in 12-week-old calves. Vet. World 2021, 14, 2883–2888. [Google Scholar] [CrossRef] [PubMed]
- Gorka, P.; Kowalski, Z.M.; Pietrzak, P.; Kotunia, A.; Jagusiak, W.; Holst, J.J.; Guilloteau, P.; Zabielski, R. Effect of method of delivery of sodium butyrate on rumen development in newborn calves. J. Dairy Sci. 2011, 94, 5578–5588. [Google Scholar] [CrossRef] [PubMed]
- Kern, R.J.; Lindholm-Perry, A.K.; Freetly, H.C.; Kuehn, L.A.; Rule, D.C.; Ludden, P.A. Rumen papillae morphology of beef steers relative to gain and feed intake and the association of volatile fatty acids with kallikrein gene expression. Livest. Sci. 2016, 187, 24–30. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Liang, G.; Guan, L.L. Regulation of rumen development in neonatal ruminants through microbial metagenomes and host transcriptomes. Genome Biol. 2019, 20, 172. [Google Scholar] [CrossRef]
- Novak, T.E.; Rodriguez-Zas, S.L.; Southey, B.R.; Starkey, J.D.; Stockler, R.M.; Alfaro, G.F.; Moisa, S.J. Jersey steer ruminal papillae histology and nutrigenomics with diet changes. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1694–1707. [Google Scholar] [CrossRef] [PubMed]
- Yohe, T.T.; Dennis, T.S.; Buss, L.N.; Croft, E.J.D.; Quigley, J.D.; Hill, T.M.; Suarez-Mena, F.X.; Aragona, K.M.; Laarman, A.H.; Costa, J.H.C.; et al. Performance and visceral tissue growth and development of Holstein calves fed differing milk replacer allowances and starch concentrations in pelleted starter. J. Dairy Sci. 2022, 105, 4099–4115. [Google Scholar] [CrossRef]
- Yohe, T.T.; O’Diam, K.M.; Daniels, K.M. Growth, ruminal measurements, and health characteristics of Holstein bull calves fed an Aspergillus oryzae fermentation extract. J. Dairy Sci. 2015, 98, 6163–6175. [Google Scholar] [CrossRef]
- Lane, M.A.; Baldwin, R.L.t.; Jesse, B.W. Sheep rumen metabolic development in response to age and dietary treatments. J. Anim. Sci. 2000, 78, 1990–1996. [Google Scholar] [CrossRef]
- Nishihara, K.; Suzuki, Y.; Kim, D.; Roh, S. Growth of rumen papillae in weaned calves is associated with lower expression of insulin-like growth factor-binding proteins 2, 3, and 6. Anim. Sci. J. 2019, 90, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- Tamate, H.; McGilliard, A.D.; Jacobson, N.L.; Getty, R. Effect of Various Dietaries on the Anatomical Development of the Stomach in the Calf. J. Dairy Sci. 1962, 45, 408–420. [Google Scholar] [CrossRef]
- Yohe, T.T.; Schramm, H.; Parsons, C.L.M.; Tucker, H.L.M.; Enger, B.D.; Hardy, N.R.; Daniels, K.M. Form of calf diet and the rumen. I: Impact on growth and development. J. Dairy Sci. 2019, 102, 8486–8501. [Google Scholar] [CrossRef]
- Gentile, A.; Sconza, S.; Lorenz, I.; Otranto, G.; Rademacher, G.; Famigli-Bergamini, P.; Klee, W. D-Lactic acidosis in calves as a consequence of experimentally induced ruminal acidosis. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2004, 51, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Dieho, K.; Bannink, A.; Geurts, I.A.L.; Schonewille, J.T.; Gort, G.; Dijkstra, J. Morphological adaptation of rumen papillae during the dry period and early lactation as affected by rate of increase of concentrate allowance. J. Dairy Sci. 2016, 99, 2339–2352. [Google Scholar] [CrossRef]
- Connor, E.E.; Baldwin, R.L.t.; Li, C.J.; Li, R.W.; Chung, H. Gene expression in bovine rumen epithelium during weaning identifies molecular regulators of rumen development and growth. Funct. Integr. Genom. 2013, 13, 133–142. [Google Scholar] [CrossRef]
- Rosenboom, R.W. Rumen Transition from Weaning to 400 Pounds. Vet. Clin. N. Am. Food Anim. Pract. 2022, 38, 153–164. [Google Scholar] [CrossRef]
- Niwinska, B.; Hanczakowska, E.; Arciszewski, M.B.; Klebaniuk, R. Review: Exogenous butyrate: Implications for the functional development of ruminal epithelium and calf performance. Animal 2017, 11, 1522–1530. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sun, D.; Mao, S.; Zhu, W.; Liu, J. Infusion of sodium butyrate promotes rumen papillae growth and enhances expression of genes related to rumen epithelial VFA uptake and metabolism in neonatal twin lambs. J. Anim. Sci. 2019, 97, 909–921. [Google Scholar] [CrossRef]
- Sakata, T.; Tamate, H. Rumen epithelial cell proliferation accelerated by rapid increase in intraruminal butyrate. J. Dairy Sci. 1978, 61, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.A.; Jesse, B.W. Effect of volatile fatty acid infusion on development of the rumen epithelium in neonatal sheep. J. Dairy Sci. 1997, 80, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Mentschel, J.; Leiser, R.; Mulling, C.; Pfarrer, C.; Claus, R. Butyric acid stimulates rumen mucosa development in the calf mainly by a reduction of apoptosis. Arch. Tierernahr. 2001, 55, 85–102. [Google Scholar] [CrossRef]
- Chai, J.; Liu, Z.; Wu, J.; Kang, Y.; Abdelsattar, M.M.; Zhao, W.; Wang, S.; Yang, S.; Deng, F.; Li, Y.; et al. Dietary beta-hydroxybutyric acid improves the growth performance of young ruminants based on rumen microbiota and volatile fatty acid biosynthesis. Front. Microbiol. 2023, 14, 1296116. [Google Scholar] [CrossRef]
- Flatt, W.P.; Warner, R.G.; Loosli, J.K. Influence of purified materials on the development of the ruminant stomach. J. Dairy Sci. 1958, 41, 1593–1600. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, Y.; Li, P.F. PPARalpha: An emerging target of metabolic syndrome, neurodegenerative and cardiovascular diseases. Front. Endocrinol. 2022, 13, 1074911. [Google Scholar] [CrossRef]
- Malhi, M.; Gui, H.; Yao, L.; Aschenbach, J.R.; Gabel, G.; Shen, Z. Increased papillae growth and enhanced short-chain fatty acid absorption in the rumen of goats are associated with transient increases in cyclin D1 expression after ruminal butyrate infusion. J. Dairy Sci. 2013, 96, 7603–7616. [Google Scholar] [CrossRef]
- Ahmad, A.A.; Zhang, J.; Liang, Z.; Du, M.; Yang, Y.; Zheng, J.; Yan, P.; Long, R.; Tong, B.; Han, J.; et al. Age-dependent variations in rumen bacterial community of Mongolian cattle from weaning to adulthood. BMC Microbiol. 2022, 22, 213. [Google Scholar] [CrossRef] [PubMed]
- Sujani, S.; Seresinhe, R.T. Exogenous Enzymes in Ruminant Nutrition: A Review. Asian J. Anim. Sci. 2015, 9, 85–99. [Google Scholar] [CrossRef]
- Li, K.; Shi, B.; Na, R. The Colonization of Rumen Microbiota and Intervention in Pre-Weaned Ruminants. Animals 2023, 13, 994. [Google Scholar] [CrossRef] [PubMed]
- Minato, H.; Otsuka, M.; Shirasaka, S.; Itabashi, H.; Mitsumori, M. Colonization of microorganisms in the rumen of young calves. J. Gen. Appl. Microbiol. 1992, 38, 447–456. [Google Scholar] [CrossRef]
- Hu, R.; Zou, H.; Wang, Z.; Cao, B.; Peng, Q.; Jing, X.; Wang, Y.; Shao, Y.; Pei, Z.; Zhang, X.; et al. Nutritional Interventions Improved Rumen Functions and Promoted Compensatory Growth of Growth-Retarded Yaks as Revealed by Integrated Transcripts and Microbiome Analyses. Front. Microbiol. 2019, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Xie, F.; Sun, D.; Liu, J.; Zhu, W.; Mao, S. Ruminal microbiome-host crosstalk stimulates the development of the ruminal epithelium in a lamb model. Microbiome 2019, 7, 83. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, Y.; Yu, Z.; Xu, Q.; Zheng, N.; Zhao, S.; Huang, G.; Wang, J. Ruminal microbiota-host interaction and its effect on nutrient metabolism. Anim. Nutr. 2021, 7, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Na, S.W.; Guan, L.L. Understanding the role of rumen epithelial host-microbe interactions in cattle feed efficiency. Anim. Nutr. 2022, 10, 41–53. [Google Scholar] [CrossRef]
- Sakata, T.; Hikosaka, K.; Shiomura, Y.; Tamate, H. Stimulatory effect of insulin on ruminal epithelium cell mitosis in adult sheep. Br. J. Nutr. 1980, 44, 325–331. [Google Scholar] [CrossRef]
- Kato, S.; Sato, K.; Chida, H.; Roh, S.G.; Ohwada, S.; Sato, S.; Guilloteau, P.; Katoh, K. Effects of Na-butyrate supplementation in milk formula on plasma concentrations of GH and insulin, and on rumen papilla development in calves. J. Endocrinol. 2011, 211, 241–248. [Google Scholar] [CrossRef]
- Gálfi, P.; Neogrády, S.; Sakata, T. Effects of Volatile Fatty Acids on the Epithelial Cell Proliferation of the Digestive Tract and Its Hormonal Mediation. In Physiological Aspects of Digestion and Metabolism in Ruminants; Elsevier: Amsterdam, The Netherlands, 1991; pp. 49–59. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Q.; Zhang, Y.L.; Pei, C.X.; Zhang, S.L.; Guo, G.; Huo, W.J.; Yang, W.Z.; Wang, H. Effects of isobutyrate supplementation in pre- and post-weaned dairy calves diet on growth performance, rumen development, blood metabolites and hormone secretion. Animal 2017, 11, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Zitnan, R.; Kuhla, S.; Sanftleben, P.; Bilska, A.; Schneider, F.; Zupcanova, M.; Voigt, J. Diet induced ruminal papillae development in neonatal calves not correlating with rumen butyrate. Veterinární Medicína 2005, 50, 472–479. [Google Scholar] [CrossRef]
- Shen, Z.; Seyfert, H.M.; Lohrke, B.; Schneider, F.; Zitnan, R.; Chudy, A.; Kuhla, S.; Hammon, H.M.; Blum, J.W.; Martens, H.; et al. An energy-rich diet causes rumen papillae proliferation associated with more IGF type 1 receptors and increased plasma IGF-1 concentrations in young goats. J. Nutr. 2004, 134, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, K.; Suzuki, Y.; Roh, S. Ruminal epithelial insulin-like growth factor-binding proteins 2, 3, and 6 are associated with epithelial cell proliferation. Anim. Sci. J. 2020, 91, e13422. [Google Scholar] [CrossRef]
- Tarnawski, A.S.; Ahluwalia, A. The Critical Role of Growth Factors in Gastric Ulcer Healing: The Cellular and Molecular Mechanisms and Potential Clinical Implications. Cells 2021, 10, 1964. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Kuhla, S.; Zitnan, R.; Seyfert, H.M.; Schneider, F.; Hagemeister, H.; Chudy, A.; Lohrke, B.; Blum, J.W.; Hammon, H.M.; et al. Intraruminal infusion of n-butyric acid induces an increase of ruminal papillae size independent of IGF-1 system in castrated bulls. Arch. Anim. Nutr. 2005, 59, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Pico, C.; Palou, M.; Pomar, C.A.; Rodriguez, A.M.; Palou, A. Leptin as a key regulator of the adipose organ. Rev. Endocr. Metab. Disord. 2022, 23, 13–30. [Google Scholar] [CrossRef]
- Hayashi, H.; Yamakado, M.; Yamaguchi, M.; Kozakai, T. Leptin and ghrelin expressions in the gastrointestinal tracts of calves and cows. In Journal of Veterinary Medical Science; Japanese Society of Veterinary Science: Tokyo, Japan, 2020; Volume 82, pp. 475–478. [Google Scholar] [CrossRef]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cheng, L.; Guo, J.; Ma, Y.; Li, F. Expression of Ghrelin in gastrointestinal tract and the effect of early weaning on Ghrelin expression in lambs. Mol. Biol. Rep. 2014, 41, 909–914. [Google Scholar] [CrossRef]
- Greco, G.; Amasheh, S.; Shen, Z.; Lu, Z.; Aschenbach, J.R. Effects of glucagon-like peptides 1 and 2 and epidermal growth factor on the epithelial barrier of the rumen of adult sheep. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1727–1738. [Google Scholar] [CrossRef]
- Baldwin, R.L. The proliferative actions of insulin, insulin-like growth factor-I, epidermal growth factor, butyrate and propionate on ruminal epithelial cells in vitro. Small Rumin. Res. 1999, 32, 261–268. [Google Scholar] [CrossRef]
- Connor, E.E.; Baldwin, R.L.t.; Walker, M.P.; Ellis, S.E.; Li, C.; Kahl, S.; Chung, H.; Li, R.W. Transcriptional regulators transforming growth factor-beta1 and estrogen-related receptor-alpha identified as putative mediators of calf rumen epithelial tissue development and function during weaning. J. Dairy Sci. 2014, 97, 4193–4207. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xue, M.; Zhang, L.; Li, L.; Lian, H.; Li, M.; Gao, T.; Fu, T.; Tu, Y. Integration of Long Non-Coding RNA and mRNA Profiling Reveals the Mechanisms of Different Dietary NFC/NDF Ratios Induced Rumen Development in Calves. Animals 2022, 12, 650. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fan, H.; Li, M.; Zhao, K.; Xia, S.; Chen, Y.; Shao, J.; Tang, T.; Bai, X.; Liu, Z.; et al. Integration of Non-Coding RNA and mRNA Profiles Reveals the Mechanisms of Rumen Development Induced by Different Types of Diet in Calves. Genes 2023, 14, 1093. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Weary, D.M.; von Keyserlingk, M.A. Invited review: Effects of milk ration on solid feed intake, weaning, and performance in dairy heifers. J. Dairy Sci. 2011, 94, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Enriquez, D.; Hotzel, M.J.; Ungerfeld, R. Minimising the stress of weaning of beef calves: A review. Acta Vet. Scand. 2011, 53, 28. [Google Scholar] [CrossRef] [PubMed]
- Carballo, O.C.; Khan, M.A.; Knol, F.W.; Lewis, S.J.; Stevens, D.R.; Laven, R.A.; McCoard, S.A. Impact of weaning age on rumen development in artificially reared lambs1. J. Anim. Sci. 2019, 97, 3498–3510. [Google Scholar] [CrossRef] [PubMed]
- Mikus, T.; Marzel, R.; Mikus, O. Early weaning: New insights on an ever-persistent problem in the dairy industry. J. Dairy Res. 2020, 87, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Schwarzkopf, S.; Kinoshita, A.; Huther, L.; Salm, L.; Kehraus, S.; Sudekum, K.H.; Huber, K.; Danicke, S.; Frahm, J. Weaning age influences indicators of rumen function and development in female Holstein calves. BMC Vet. Res. 2022, 18, 102. [Google Scholar] [CrossRef]
- Liu, S.; Ma, J.; Li, J.; Alugongo, G.M.; Wu, Z.; Wang, Y.; Li, S.; Cao, Z. Effects of Pair Versus Individual Housing on Performance, Health, and Behavior of Dairy Calves. Animals 2019, 10, 50. [Google Scholar] [CrossRef]
- Eckert, E.; Brown, H.E.; Leslie, K.E.; DeVries, T.J.; Steele, M.A. Weaning age affects growth, feed intake, gastrointestinal development, and behavior in Holstein calves fed an elevated plane of nutrition during the preweaning stage. J. Dairy Sci. 2015, 98, 6315–6326. [Google Scholar] [CrossRef] [PubMed]
- Agustinho, B.C.; Wolfe, A.; Tsai, C.Y.; Pereira, L.M.; Konetchy, D.E.; Laarman, A.H.; Rezamand, P. Effect of weaning age and pace on blood metabolites, cortisol concentration, and the mRNA abundance of inflammation-related genes in gastrointestinal, adipose, and liver tissue of Holstein dairy calves. J. Dairy Sci. 2024. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Lee, H.J.; Lee, W.S.; Kim, H.S.; Ki, K.S.; Hur, T.Y.; Suh, G.H.; Kang, S.J.; Choi, Y.J. Structural growth, rumen development, and metabolic and immune responses of Holstein male calves fed milk through step-down and conventional methods. J. Dairy Sci. 2007, 90, 3376–3387. [Google Scholar] [CrossRef] [PubMed]
- Schwarzkopf, S.; Kinoshita, A.; Kluess, J.; Kersten, S.; Meyer, U.; Huber, K.; Danicke, S.; Frahm, J. Weaning Holstein Calves at 17 Weeks of Age Enables Smooth Transition from Liquid to Solid Feed. Animals 2019, 9, 1132. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, M.H.; Kertz, A.F. Review: Effects of different forms of calf starters on feed intake and growth rate: A systematic review and Bayesian meta-analysis of studies from 1938 to 2021. Appl. Anim. Sci. 2021, 37, 273–293. [Google Scholar] [CrossRef]
- Kertz, A.F.; Hill, T.M.; Quigley, J.D., 3rd; Heinrichs, A.J.; Linn, J.G.; Drackley, J.K. A 100-Year Review: Calf nutrition and management. J. Dairy Sci. 2017, 100, 10151–10172. [Google Scholar] [CrossRef] [PubMed]
- Armengol, R.; Fraile, L. Colostrum and milk pasteurization improve health status and decrease mortality in neonatal calves receiving appropriate colostrum ingestion. J. Dairy Sci. 2016, 99, 4718–4725. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Khan, M.Z.; Xiao, J.; Alugongo, G.M.; Chen, X.; Li, S.; Wang, Y.; Cao, Z. An Overview of Waste Milk Feeding Effect on Growth Performance, Metabolism, Antioxidant Status and Immunity of Dairy Calves. Front. Vet. Sci. 2022, 9, 898295. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, W.B.; Bi, Y.L.; Tu, Y.; Beckers, Y.; Du, H.C.; Diao, Q.Y. Early Feeding Regime of Waste Milk, Milk, and Milk Replacer for Calves Has Different Effects on Rumen Fermentation and the Bacterial Community. Animals 2019, 9, 443. [Google Scholar] [CrossRef]
- Arowolo, M.A.; He, J. Use of probiotics and botanical extracts to improve ruminant production in the tropics: A review. Anim. Nutr. 2018, 4, 241–249. [Google Scholar] [CrossRef]
- Fomenky, B.E.; Do, D.N.; Talbot, G.; Chiquette, J.; Bissonnette, N.; Chouinard, Y.P.; Lessard, M.; Ibeagha-Awemu, E.M. Direct-fed microbial supplementation influences the bacteria community composition of the gastrointestinal tract of pre- and post-weaned calves. Sci. Rep. 2018, 8, 14147. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, N.A.; Chethan, H.S.; Srivastava, R.; Gabbur, A.B. Role of probiotics in ruminant nutrition as natural modulators of health and productivity of animals in tropical countries: An overview. Trop. Anim. Health Prod. 2022, 54, 110. [Google Scholar] [CrossRef] [PubMed]
- McCann, J.C.; Elolimy, A.A.; Loor, J.J. Rumen Microbiome, Probiotics, and Fermentation Additives. Vet. Clin. N. Am. Food Anim. Pract. 2017, 33, 539–553. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Baran, M.; Kmet, V. Effect of pectinase on rumen fermentation in sheep and lambs. Arch. Tierernahr. 1987, 37, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.T.; Anderson, K.L.; Yoon, I.; Scott, M.F.; Carlson, S.A. Amelioration of salmonellosis in pre-weaned dairy calves fed Saccharomyces cerevisiae fermentation products in feed and milk replacer. Vet. Microbiol. 2014, 172, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Laksesvela, B.; Ommundsen, A.; Landsverk, T. Indigestion in young calves. V. The influence of grass silage and fine hay. Acta Vet. Scand. 1978, 19, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Chester-Jones, H.; Fontenot, J.P.; Veit, H.P. Physiological and pathological effects of feeding high levels of magnesium to steers. J. Anim. Sci. 1990, 68, 4400–4413. [Google Scholar] [CrossRef]
- Mu, Y.; Qi, W.; Zhang, T.; Zhang, J.; Mao, S. Multi-omics Analysis Revealed Coordinated Responses of Rumen Microbiome and Epithelium to High-Grain-Induced Subacute Rumen Acidosis in Lactating Dairy Cows. mSystems 2022, 7, e0149021. [Google Scholar] [CrossRef]
- Pan, X.; Ma, Z.; Sun, X.; Li, H.; Zhang, T.; Zhao, C.; Wang, N.; Heller, R.; Hung Wong, W.; Wang, W.; et al. CNEReg Interprets Ruminant-specific Conserved Non-coding Elements by Developmental Gene Regulatory Network. Genom. Proteom. Bioinform. 2023, 21, 632–648. [Google Scholar] [CrossRef] [PubMed]
- Do, D.N.; Dudemaine, P.L.; Fomenky, B.E.; Ibeagha-Awemu, E.M. Integration of miRNA weighted gene co-expression network and miRNA-mRNA co-expression analyses reveals potential regulatory functions of miRNAs in calf rumen development. Genomics 2019, 111, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Wray, G.A. The evolutionary significance of cis-regulatory mutations. Nat. Rev. Genet. 2007, 8, 206–216. [Google Scholar] [CrossRef]
- Zhang, Y.; Cai, W.; Li, Q.; Wang, Y.; Wang, Z.; Zhang, Q.; Xu, L.; Xu, L.; Hu, X.; Zhu, B.; et al. Transcriptome Analysis of Bovine Rumen Tissue in Three Developmental Stages. Front. Genet. 2022, 13, 821406. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, K.; Kato, D.; Suzuki, Y.; Kim, D.; Nakano, M.; Yajima, Y.; Haga, S.; Nakano, M.; Ishizaki, H.; Kawahara-Miki, R.; et al. Comparative transcriptome analysis of rumen papillae in suckling and weaned Japanese Black calves using RNA sequencing. J. Anim. Sci. 2018, 96, 2226–2237. [Google Scholar] [CrossRef]
- Chen, C.; Yin, Y.; Li, H.; Zhou, B.; Zhou, J.; Zhou, X.; Li, Z.; Liu, G.; Pan, X.; Zhang, R.; et al. Ruminant-specific genes identified using high-quality genome data and their roles in rumen evolution. Sci. Bull. 2022, 67, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Wu, C. ILK: A pseudokinase in the center stage of cell-matrix adhesion and signaling. Curr. Opin. Cell Biol. 2012, 24, 607–613. [Google Scholar] [CrossRef]
- Sha, Y.; He, Y.; Liu, X.; Zhao, S.; Hu, J.; Wang, J.; Li, S.; Li, W.; Shi, B.; Hao, Z. Rumen Epithelial Development- and Metabolism-Related Genes Regulate Their Micromorphology and VFAs Mediating Plateau Adaptability at Different Ages in Tibetan Sheep. Int. J. Mol. Sci. 2022, 23, 16078. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, Y.; Andreyev, O.; Hoyt, R.F., Jr.; Singh, A.; Hunt, T.; Horvath, K.A. Overexpression of FABP3 inhibits human bone marrow derived mesenchymal stem cell proliferation but enhances their survival in hypoxia. Exp. Cell Res. 2014, 323, 56–65. [Google Scholar] [CrossRef]
- Kato, D.; Suzuki, Y.; Haga, S.; So, K.; Yamauchi, E.; Nakano, M.; Ishizaki, H.; Choi, K.; Katoh, K.; Roh, S.G. Utilization of digital differential display to identify differentially expressed genes related to rumen development. Anim. Sci. J. 2016, 87, 584–590. [Google Scholar] [CrossRef]
- Lu, J.; Zhao, H.; Xu, J.; Zhang, L.; Yan, L.; Shen, Z. Elevated cyclin D1 expression is governed by plasma IGF-1 through Ras/Raf/MEK/ERK pathway in rumen epithelium of goats supplying a high metabolizable energy diet. J. Anim. Physiol. Anim. Nutr. 2013, 97, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Hernandez, V.; Quiros, M.; Nusrat, A. Intestinal epithelial claudins: Expression and regulation in homeostasis and inflammation. Ann. N. Y. Acad. Sci. 2017, 1397, 66–79. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Macroscopic view of the internal surface of the rumen. (A) The rumen of a newborn calf; (B) The rumen of an adult steer. Note that compared to the newborn rumen, the inner surface of the adult rumen is covered with large rumen papillae.
Figure 1.
Macroscopic view of the internal surface of the rumen. (A) The rumen of a newborn calf; (B) The rumen of an adult steer. Note that compared to the newborn rumen, the inner surface of the adult rumen is covered with large rumen papillae.

Figure 2.
(A) Microscopic view of the rumen wall. The section was cut from the rumen of a newborn calf and stained with hematoxylin and eosin. a: rumen papilla; b: mucosal and submucosal layer; c: muscular layer; d: serosal layer. (B) Schematic representation of different cellular layers of the rumen epithelium.
Figure 2.
(A) Microscopic view of the rumen wall. The section was cut from the rumen of a newborn calf and stained with hematoxylin and eosin. a: rumen papilla; b: mucosal and submucosal layer; c: muscular layer; d: serosal layer. (B) Schematic representation of different cellular layers of the rumen epithelium.

Figure 3.
Schematic representation of absorption and metabolism of volatile fatty acids. CA: carbonic anhydrase; VFA: volatile fatty acids; ACSS1: acyl-CoA synthetase short-chain family member 1; ACADS: acyl-CoA dehydrogenase short-chain; ECHS1: enoyl-CoA hydratase, short chain 1; ACAT1: acetyl-CoA acetyltransferase 1; EHHADH: enoyl-CoA hydratase and 3-hydroxyacyl CoA dehydrogenase; HMGCS2: 3-hydroxy-3-methylglutaryl-CoA synthase 2; HMGCL: 3-hydroxy-3-methylglutaryl-CoA lyase; OXCT1: 3-oxoacid CoA-transferase 1; BDH1: 3-hydroxybutyrate dehydrogenase 1; SDH: succinate dehydrogenase; MDH: malate dehydrogenase; LDH: lactate dehydrogenase.
Figure 3.
Schematic representation of absorption and metabolism of volatile fatty acids. CA: carbonic anhydrase; VFA: volatile fatty acids; ACSS1: acyl-CoA synthetase short-chain family member 1; ACADS: acyl-CoA dehydrogenase short-chain; ECHS1: enoyl-CoA hydratase, short chain 1; ACAT1: acetyl-CoA acetyltransferase 1; EHHADH: enoyl-CoA hydratase and 3-hydroxyacyl CoA dehydrogenase; HMGCS2: 3-hydroxy-3-methylglutaryl-CoA synthase 2; HMGCL: 3-hydroxy-3-methylglutaryl-CoA lyase; OXCT1: 3-oxoacid CoA-transferase 1; BDH1: 3-hydroxybutyrate dehydrogenase 1; SDH: succinate dehydrogenase; MDH: malate dehydrogenase; LDH: lactate dehydrogenase.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pokhrel, B.; Jiang, H. Postnatal Growth and Development of the Rumen: Integrating Physiological and Molecular Insights. Biology 2024, 13, 269. https://doi.org/10.3390/biology13040269
AMA Style
Pokhrel B, Jiang H. Postnatal Growth and Development of the Rumen: Integrating Physiological and Molecular Insights. Biology. 2024; 13(4):269. https://doi.org/10.3390/biology13040269
Chicago/Turabian StylePokhrel, Binod, and Honglin Jiang. 2024. "Postnatal Growth and Development of the Rumen: Integrating Physiological and Molecular Insights" Biology 13, no. 4: 269. https://doi.org/10.3390/biology13040269
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.