
Chromatin’s Influence on Pre-Replication Complex Assembly and Function
1
Department of Cell and Developmental Biology, University of Illinois at Urbana-Champaign, 601S Goodwin Avenue, Urbana, IL 61801, USA
2
Cancer Center at Illinois, University of Illinois Urbana-Champaign, Urbana, IL 61801, USA
*
Author to whom correspondence should be addressed.
Biology 2024, 13(3), 152; https://doi.org/10.3390/biology13030152
Submission received: 10 January 2024
/
Revised: 19 February 2024
/
Accepted: 19 February 2024
/
Published: 27 February 2024
(This article belongs to the Special Issue The Replication Licensing System)
Abstract
:Simple Summary
Genome duplication and the faithful segregation of chromosomes to daughter cells are crucial for the maintenance of genome stability. Replication initiators play a crucial role in ensuring that DNA replication occurs only once per cell division cycle. This review summarizes when, where, and how replication initiates in human cells.
Abstract
In all eukaryotes, the initiation of DNA replication requires a stepwise assembly of factors onto the origins of DNA replication. This is pioneered by the Origin Recognition Complex, which recruits Cdc6. Together, they bring Cdt1, which shepherds MCM2-7 to form the OCCM complex. Sequentially, a second Cdt1-bound hexamer of MCM2-7 is recruited by ORC-Cdc6 to form an MCM double hexamer, which forms a part of the pre-RC. Although the mechanism of ORC binding to DNA varies across eukaryotes, how ORC is recruited to replication origins in human cells remains an area of intense investigation. This review discusses how the chromatin environment influences pre-RC assembly, function, and, eventually, origin activity.
1. Introduction
Eukaryotic cells need to replicate their genome accurately, which requires carefully orchestrated molecular players acting at the right time and place. Replication starts at replication origins, where a sequence of binding events occurs [1]. At the end of mitosis/early G1, the Origin Recognition Complex (ORC) assembles at these replication origins. ORC recruits Cdc6 (cell division cycle 6) and Cdt1 (chromatin licensing and DNA replication factor 1), which helps to load the minichromosome maintenance protein complex (MCM complex consisting of MCM2-7) to form the OCCM complex. The recruitment of a second MCM2-7 hexamer to form the head-to-head hexamers completes the pre-RC assembly (reviewed in [2,3]). Origin licensing, which involves the loading of double hexameric MCM complexes at replication origins, is a highly conserved process. This process ensures that DNA replication occurs only once per cell division cycle.
Our understanding of the intricate steps of replication initiation comes from elaborate work in budding yeast, though many of the principles are also conserved in higher eukaryotes. The initiation of replication is further controlled by Dbf4-dependent kinase (DDK) and cyclin-dependent kinase (CDK), which are sequentially activated during the transition from G1 to the S phase [4,5,6,7,8]. DDK phosphorylates MCM2-7 subunits, which allows the loading of Cdc45 and Sld3, followed by the phosphorylation of Sld2 and Sld3 by CDK [9,10,11,12,13,14]. A series of events ensures that the CMG complex (Cdc45, MCM2-7, and GINS) is activated as the helicase for replication to proceed [15].
The origin recognition complex (ORC) was identified from cell extracts of Saccharomyces cerevisiae as a heterohexameric complex bound to the autonomously replicating sequence (ARS, origins of replication in Saccharomyces cerevisiae), comprising Orc1 through to Orc6 in descending order of their molecular mass [16]. Orthologs of each ORC subunit have been identified in diverse organisms, including humans, frogs, flies, nematodes, and plants [17]. Although the replication origins vary from one species to another, ORC binds to replication origins in diverse organisms. What defines an origin and how ORC recognizes these origins differs significantly amongst different species.
In human cells, Orc2-5 forms the core of the ORC and remains associated as a complex throughout the cell cycle [18,19,20,21]. Several years ago, our laboratory identified ORC-Associated/Leucine-rich and WD-repeat-containing (ORCA/LRWD1) as an ORC-associated factor that stabilized ORC onto chromatin [22,23,24]. The core-ORC and ORCA are stably associated with one another throughout the cell cycle. Human ORC subunits are highly dynamic in their association with chromatin across the cell cycle. Orc2 and ORCA are released from most chromatin sites at the end of G1, but remain associated with heterochromatin sites, including telomeres and centromeres in human cells [25,26,27]. The largest subunit, human Orc1, is spatiotemporally regulated (Figure 1) and is dynamically associated with the ORC on the chromatin and is degraded at the end of G1 following SCF/Skp2-mediated ubiquitination [28]. It is reloaded onto the chromatin during the establishment of the next pre-RC in the late M to early G1 phase [29]. Multiple ORC subunits belong to the AAA+ family of proteins. ORC is an ATPase, and its binding to the origin DNA is ATP-dependent [30].
2. Chromatin Signature at the Origins
In metazoans, replication origins are highly heterogeneous, and their regulation is complex [31]. Replication in metazoans initiates at sites that do not show any clear sequence specificity, unlike in budding yeast, where the initiation sites are, to a large extent, clearly defined [32]. Recent analyses have suggested that GC-rich sequences, which are origin G-rich repeated elements (OGREs) that can form G4 quadraplexes, transcription start sites, and regions of DNAse hypersensitivity, are often found at metazoan origins [33,34,35,36,37,38,39,40,41,42]. Replication initiates in moderately transcribed regions and is associated with DNase hypersensitive sites. Moreover, genome-wide experiments have bolstered the co-regulation model of replication initiation and transcription [43]. These are active areas of study, and we hope to learn the mechanistic details in the coming years.
In higher eukaryotes, the DNA sequence is not the sole determinant of replication initiation events, and origin specification may depend on epigenetic features. The firing of origins is spatially and temporally controlled in a cell-, tissue-, and development-specific manner [37,44,45,46,47,48]. The stochastic model of replication origin firing posits that the firing time of an individual origin in a population of cells is heterogeneous, firing early in some and late in others. The heterogeneity of the stochastic model makes identifying origins challenging. The Dutta and Zang labs have recently suggested that the origins are specified by diverse stochastic events that are dependent on the epigenetic accessibility around promoters. However, intriguingly, the ORC-binding sites or the MCM complex did not overlap with the origins [49]. A series of genome-wide studies with various ORC antibodies, specifically at distinct time points within G1, would be important to pinpoint ORC dynamics in human cells.
It has been suggested that the chromatin environment plays a crucial role in dictating the activation of replication origins [37]. A common feature of all yeast origins seems to be a nucleosome-free and G-rich region that is present upstream of a nucleosome positioned at the initiation sites. These results support the concept of chromatin-dependent origins in budding yeast [50]. The polycomb group (PcG) of proteins comprises two multi-subunit complexes, PRC1 and PRC2, catalyzing H2AK119ub and H3K27me3, respectively. PcG proteins are involved in transcriptional silencing. For example, the association of H3K4me1 with H3K27methylation, catalyzed by the polycomb group protein EZH2, is suggested to provide an environment conducive for DNA replication initiation [51]. Supporting work has shown that PcG proteins are required for origin activation [52,53] and that these proteins structurally restrict origin activation within the polycomb domains. The chromatin mark H3K4me3, which is associated with active transcription and open chromatin, is often found at the origins of mouse stem cells. These origins also have H3K9ac, facilitating accessibility [54]. It is generally accepted that chromatin-modifying factors can create a chromatin environment that enables transcription and replication [55,56,57].
Comprehensive work from the Aladjem lab has demonstrated that common origins are associated most strongly with unmethylated CpG islands, H3K4me3, and H3K9Ac. In contrast, cell-type-specific origins are primarily associated with methylated CpG islands and H3K9me3 and replicate late [42]. They further demonstrated that the early replicating regions often correlate with histone marks associated with transcriptionally active domains, such as H3K4me1/2/3, H3K9ac, H3K18ac, H3K36me3, and H2K27ac. In contrast, late replicating origins are associated with heterochromatin, are often associated with H3 and H4 hypoacetylation, and are enriched with H3K9 and H3K27 methylation [36,37,42]. A list of the various chromatin marks associated with various aspects of replication initiation are listed in Table 1.
3. Chromatin Governs the Spatiotemporal Dynamics of Pre-RC Factors
Accumulating evidence points towards the fact that global chromatin structures dictate the replication program, starting from the establishment of pre-RC to the initiation of replication [58]. We have demonstrated that human Orc5 associates with the histone acetyltransferase and GCN5. Tethering Orc5 to a chromatin locus increases acetylation-mediated large-scale decondensation [59]. The ability of Orc5 to induce chromatin unfolding during G1 likely enables the establishment of pre-RC at the origins. Similarly, H4 acetylation at the origins by the ORC-interacting protein HBO1 (human acetylase binding to ORC1) is critical for replication licensing by Cdt1 [60,61]. ORC-mediated chromatin acetylation has been suggested to control DNA replication through pre-RC formation [62].
ORC and ORCA/LRWD1 associate with repressive histone marks, namely H3K9me3, H4K20me3, and H3K27me3 (Figure 2) [63,64,65,66,67]. Recent cryoEM work from the Bleichert lab has suggested that ORCA establishes a ternary complex by simultaneously recognizing Orc2, nucleosomal DNA, and repressive histone trimethylation through an aromatic cage [68]. The authors suggest that this enables reorganization of the local chromatin architecture by diminishing nucleosome self-association. Interestingly, ORC and ORCA also bind to methylated DNA and the methyltransferases that catalyze the above histone modifications and DNA methylation. Our results have previously demonstrated that ORCA promotes late replication, similar to the ORC-mediated stabilization of heterochromatin protein 1 (HP1 α and β in human cells) for establishing late replicating domains [69,70,71,72]. This is supported by new evidence that ORCA is necessary and sufficient to recruit ORC into chromatin condensates marked by H4K20 trimethylation [68]. Although increasing evidence suggests that ORCA plays a key role in recruiting ORC to specific chromatin sites, it is equally intriguing that Orc1 binds to H4K20me2 at the origins via its bromo-adjacent homology (BAH) domain [73]. Further, the loss of this association results in failure of Orc1 to localize to the origins [73,74]. These results support that ORC/ORCA binding to specific chromatin marks is important for the spatiotemporal regulation of DNA replication (Figure 1 and Figure 2).
The histone mark H4K20me1 has also been shown to be present at some human DNA replication origins [75,76,77,78]. Specifically, tethering PR-Set7, the methylase responsible for H4K20 methylation, to a specific genomic locus promotes pre-RC assembly. PR-Set7 is degraded by the CRL4-Cdt2 ubiquitin ligase complex during the S phase of the cell cycle and, accordingly, the stabilization of PR-Set7 or depletion of Cdt2 promotes re-replication. This re-replication phenotype depends on the subsequent trimethylation of H4K20 by Suv4-20h1/h2. Other methylation marks, such as H3K4me2 and me3, are also found at replication origins, but their function still needs to be determined.
Nucleosomes containing the H2A.Z variant can recruit Suv420H1, which is a methyl transferase, that, in turn, facilitates the deposition of H4K20me2, which is eventually recognized by the BAH domain of Orc1 [79]. The genome-wide depletion of H2A.Z leads to decreased H4K20me2 and Orc1 and nascent strand signals throughout the genome. H2A.Z-regulated replication origins fire early and have higher firing efficiency. Nucleosome-free regions next to the location of H2A.Z facilitate pre-RC loading. Although H2A.Z at the promoters seems to be required for epigenetically regulating licensing, the exact mechanism remains to be deciphered. Recently, a new functional genetic element, pG4s, associated with specific nucleosome-free regions (NFRs), was found to be sufficient for replication initiation. The authors suggested that this might be a strategy for the ORC recognition of origins in eukaryotes [80].
Work from the Liu lab has shown that yeast ORC has intrinsic nucleosome remodeling activity, which can evict H2A-H2B dimers, leaving the 2(H3-H4) tetramer on DNA, and this requires the Orc1-BAH domain [81,82]. Exciting work from the Kurat lab has recently established that in yeast, Orc1 collaborates with chromatin remodeling complexes, including INO80, ISW1a, ISW2, and other chromatin modifiers, to establish a Nucleosome Free Region (NFR) and a flanking nucleosomal array at yeast origins [83]. The theme that ORC binding sites are found at NFRs upstream of the transcription start site (TSS) seems to be conserved amongst eukaryotes [60,84,85,86,87,88]. This has provided novel insights into the critical role of ORC in nucleosome organization at the replication origins.
Human Orc1 is intriguing because it is one of the only known proteins to show dynamic spatiotemporal patterning during the G1 phase of the cell cycle (Figure 1b) [29]. Whether Orc1 binds to specific chromatin signatures during different stages within G1 to dictate MCM complex loading or sets the marks where MCM2-7 and finally PCNA would load remains an open question (Figure 1b). In budding yeast and metazoans, Orc1 has an intrinsically disordered region (IDR) that facilitates binding to the minor groove of origin DNA. A domain within the human Orc1 IDR is required for interaction between the Orc1 and CDC6 AAA+ domains in G1 [89]. The IDRs of DNA replication initiators have been shown to drive DNA-dependent phase separation in vitro and chromosome binding in vivo, and these initiator condensates selectively recruit replication-specific partner proteins for the initiation of DNA replication [90].
4. Chromatin Dictates Origin Activation
The acetylation of histones has been linked to open chromatin, and dynamic changes of this modification are known to regulate replication origin activation in yeast [91]. Also, H4K16 deacetylation decreases MCM complex loading onto early origins, dramatically changing the licensing landscape [92,93]. Similarly, at the rDNA locus, the histone deacetylase (SIR proteins) represses a large majority of the replication origins. This epigenetic silencing of origin firing promotes genome stability [94]. SIR proteins can act at the rDNA locus, euchromatic origins, and telomeres [95,96,97]. An increase in the local concentration of histone acetylation has been shown to improve Cdc45 recruitment [98]. Constitutive H3 K56ac sensitizes cells to replicative stress, in part, by negatively influencing the activation of the origins of DNA replication [99].
The methylation of histones has a varied impact on replication origin function. The lack of the ubiquitous mark H3K37me1 from origins facilitates MCM2-7 binding to origins because the presence of this mark impedes MCM2-7 association with chromatin; however, the mechanism is not known [100]. H3K4 di-methylation was found to be important for origin function [101]. Further, H3K36 methylation has been found to regulate the timing of Cdc45 binding to origins, whereas mono-methylation is associated with early replication origins and trimethylation is present at late origins [102].
The chromatin architecture, including the nucleosome positioning and occupancy at origins, impacts origin specification and activation. How this affects ORC and MCM complex loading and what kind of histone marks dictate whether an origin is permissive or restrictive to origin activation remains to be elucidated [103]. This would require a high genome-wide distribution of histone marks during temporally defined windows in G1. An AI-based tool could pinpoint whether a specific signature dictates specific origins and their efficiency, including what has been done to identify the splicing-associated chromatin signatures [104].
5. Euchromatin vs. Heterochromatin: The Replication Timing Issue
Are specific chromatin marks in the human genome dictating replication origin specification and activity? This is a conundrum that has been an active area of investigation. The consensus is that the presence of active marks associated with open chromatin and transcriptional activity, H3K4 methylation, and histone acetylation (H3K9ac, H3K18ac, H3K36ac, and H3K27ac) is correlated with early origin specification. Acetylation at H4K5, H4K8, and H4K12 by HBO1 and ORC-interacting factors is needed for replication licensing and facilitates MCM complex loading [60]. In contrast, the repressive histone marks H3K9 methylation and H3K27 methylation, accompanied by hypoacetylation, are at late-firing origins. These conclusions are based on correlative data, and it is likely a combination of chromatin marks, genomic context, cell-type-, tissue-type-, and developmental-context-dependent states that determines origin specification, ORC–MCM binding, and Cdc45 recruitment. For example, although H3K4me3 may be present at promoters and is likely associated with early origins, data has also suggested this mark represses origin firing. Further, the demethylase of H3K4me3, KDM5C, is needed for fine-tuning the methylation status at origins, and highly elevated levels of this mark result in replication defects due to a Cdc45 loading defects at origins [105].
Heterochromatin marks, including H3K9me3, are usually associated with late origins, and the demethylase KDM4D facilitates the formation of a pre-initiation complex and regulates DNA replication [106]. H3K27me3 is another repressive mark, and the methyltransferase EZH2 is strongly associated with a subset of origins and participates in the activity of these origins [107]. Although H4K20 trimethylation is often linked to heterochromatin sites and is bound by ORCA/LRWD1, H4K20 di-methylation is enriched at origins and is bound by ORC. Finally, the nucleosome containing the histone variant H2A.Z is enriched together with H4K20me2 and Orc1. H3K79 di-methylation is another modification enriched at origins, and the methyl transferase Dot1 is important for preventing genome re-replication [108].

{kind=link}
{kind=link}
Table 1.
Histone modifications with their associated factors and sites.
| Histone Modification/Marks | Downstream Effect on Transcription | Associated Factors | Sites/Functions | References |
|---|---|---|---|---|
| H3K37me1 | Repression | ARS/Set1p/Set2p | Replication origin licensing | Santos-Rosa et al., 2021 [100] |
| H3K4me2 | Activation | Orc1/HAT/SAGA complex | Likely at origins | Rizzardi et al., 2012, Rondinelli et al., 2015 [101,105] |
| H3K79me2 | Activation | CAF-1, 53BP1 | Replication initiation events/G1-S phase | Fu et al., 2013 [108] |
| H3K4me3 | Activation | Data | Early replication origin/pomoter activation | Smith et al., 2016, Cayrou et al., 2015 [37,42] |
| H3K36me3 | Activation | Set2/Cdc45 recruitment | Origin firing during S Phase/Transcription associated mark | Giri and Prasanth, 2015, Unnikrishnan et al., 2010, Smith et al., 2016 [42,65,91] |
| H3K9ac | Activation | Early replication origin/pomoter activation | Smith et al., 2016, Cayrou et al., 2015 [37,42] | |
| H3K27ac | Activation | RNA pol II/Gcn5/KAT2 | Enriched at transcriptionally active regions | Cayrou et al., 2015, Unnikrishnan et al., 2010 [37,91] |
| H3k18ac | Activation | Early replication origins | Mechali et al., 2013 [36] | |
| H3K56ac | Activation | Rtt109 | Enriched in S phase | Tremblay et al., 2023 [99] |
| H4K20me1 | Repression | PR-Set7 to enhance pre-RC formation via H4K20me/high levels in G2/M phase | Abbas et al., 2010, Mechali et al., 2013 [36,75] | |
| H4K20me2 | Repression | ORC1/H2A.Z | Enriched at origins | Long et al., 2020, Kuo et al., 2012, Mechali et al., 2013 [36,73,79] |
| H4ac | Activation | Cdt1 | Condensed chromosomes | Miotto and Struhl, 2010; 2011 [60,61] |
| H4K5, H4K8, H4K12 ac | Activation | HBO1 | MCM loading | Miotto and Struhl, 2010 [60] |
| H4K16ac | Activation | MCM loading/Sas2 | Hoggard et al., 2020, Unnikrishnan et al., 2010 [91,92] | |
| H3K4me1 + H3K27me3 | Activation + Repression | Early origins/replication initiation sites | Cayrou et al., 2015 [37] | |
| H3K9me3, H3K27me3, H4K20me3 | Repression | ORC/ORCA/HP1/PC1 | Early origins/origin licensing | Pasini et al., 2004, Piunti et al., 2014, Kuzmichev et al., 2002, Gorisch et al., 2005, Bartke et al., 2010, Giri et al., 2015, Giri and Prasanth, 2015, Vermeulen et al., 2010, Wang et al., 2017 [51,52,53,54,63,64,65,66,67] |
It is increasingly becoming evident that rather than individual chromatin marks, a combination of marks might dictate origin specification, usage, and function. For example, H3K4me3, H3K9me3, and H3K36me3 are likely associated with late-replicating origins. However, a recent study demonstrated that the presence of all these combinatorically resulted in an early replication phenotype. Furthermore, the addition of H3K56ac and the above three resulted in a higher likelihood of early replication [109,110].
6. Replication-Independent Role of ORC/ORCA in Heterochromatin Organization
ORC is implicated in transcriptional silencing in diverse eukaryotes, including yeast, drosophila, and humans [111]. In humans, the core ORC, Orc2-5, and ORCA associate with heterochromatin sites outside of G1 and bind to heterochromatin proteins HP1α and β. Furthermore, these factors play a role in heterochromatin organization. The loss of ORC and ORCA results in redistribution of the bonafide heterochromatin protein HP1α, with a concomitant alteration in the genome-wide frequency of the H3K9me3 repressive mark [64,72]. This was consistent with chromatin decondensation at satellite repeats. Using a degron-based method, we previously demonstrated that the role of human ORCA in heterochromatin organization is independent of its role in replication licensing, providing important insights into the multiple roles that these replication initiators play during the cell cycle [64]. Corroborating our data, the loss of mouse ORCA resulted in the activation of major satellite repeats, supporting the view that ORC-ORCA may play a role in silencing repeat elements [112].
The post-translation modification of ORC2 by SUMOylation was found to be required for recruiting the demethylase KDM5A. The conversion of H3K4me3 to H3K4me2 increased α-satellite transcription at the centromeres. The aberrant expression of SUMO-less Orc2 caused a reduction in α-satellite transcription and defective pericentric heterochromatin silencing, ultimately causing the re-replication of heterochromatin DNA [113]. These suggest that ORC is required to maintain genomic stability by maintaining accurate centromeric histone methylation, including H3K4, H3K9, and H4K20 trimethylation. Further experimentation is needed to establish whether the function of ORC-ORCA at the centromeric heterochromatin is related to its role in chromatin organization or the establishment of origins during mitosis (Figure 2).
All human ORC proteins are localized to constitutive heterochromatic sites, including centromeres. As suggested in a recent review, the centromeric regions and origins may have been linked in ancestral eukaryotes [1]. Centromeres may represent the origins at which ORC binds during mitosis to establish pre-RC. To support this model, it was reported that ORC is required to maintain alpha-satellite sequences at the centromere [72]. It is also important to note that repressive histone marks are enriched at centromeres, and it remains to be determined whether ORC is enriched at these sites via its interaction with H3K9me3 or H4K20me3, particularly at the pericentromeric regions (Figure 2). DNA combing experiments have suggested that the origin density is higher within the pericentromeric satellite repeats [114]. Also, the telomere repeat factor binding protein TRF2 interacts with Orc2 in pericentromeric DNA, which is required for origin activity [114]. Another possibility is the presence of H3K4 methylation, as is reported at the origins and centromeres of nematodes. With the completion of the T2T assembly of the human genome [115], we will now be able to delve deep into chromatin organization at the centromere.
ORC and ORCA also localize to centromeres and telomeres and associate with the TRF2 protein [25,116,117,118]. TRF2 binds to the origin recognition complex (ORC) and has previously been implicated in loading ORC and MCM complexes onto DNA at telomeres. Work from the Fujita lab has shown that TRF2-mediated ORC recruitment contributes to the suppression of telomere instability [118]. Its interaction with SUMOylated shelterin components facilitates ORCA’s localization to the ALT-telomeres. The loss of ORCA causes global chromatin decondensation, including at the telomeres, primarily because of the loss of H3K9me3, resulting in the deregulation of homologous recombination [25]. We have previously shown that Origin Recognition Complex-Associated (ORCA/LRWD1) protein modulates homologous recombination activity by localizing at the ALT-telomeres (Figure 2).
In addition to binding to H3K9me3, ORC and ORCA also associate with H3K27me3, a mark highly enriched on the inactive X-chromosome [112,119] (Figure 2b). The Zhang lab, using an elegant screen to identify factors involved in X-chromosome inactivation, identified several replication factors, including Orc2 and ORCA [119]. They found that Orc2 co-localized or was adjacent to the H3K27me3 focus or the Xist RNA. Further, the loss of Orc2 was found to disrupt silencing on the inactive X, and its impact on inactive X-silencing occurred through the stabilization of HP1a on the inactive X. ORC-ORCA not only binds to the H3K27me3 repressive mark, but also associates with the PRC2 complex and the methyltransferase EZH2 that catalyzes H3K27me3. ORCA functions as a scaffold, and its loss results in the destabilization of methyltransferase protein levels. Equally confusing is the fact that the loss of these methyltransferases disrupts ORCA binding to chromatin, suggesting that these factors are in a complex with one another. It is important to determine whether ORCA is a methyl-binding protein. These observations provide a unified theme that ORC proteins are multi-talented, and their chromatin association is related to their role in replication initiation and gene regulation.
7. Conclusions
One overarching question that has persisted and evolved over time is understanding the intricate mechanisms governing the regulation of DNA replication initiation within the context of chromatin. Emerging technologies, such as advanced imaging techniques, single-cell genomics, high-throughput sequencing, and the AI-based analyses of genome-wide datasets [120], need to be leveraged to gain deeper insights into the spatiotemporal regulation of chromatin and the pre-RC.
Author Contributions
H.A., N.C. and S.G.P. wrote the manuscript, H.A. prepared the table and modified the figures, N.C. formatted and made changes as per the reviewers’ suggestions. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by NSF (1818286, 2225264), NIH (GM125196), and CCIL awards to SGP.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This review summarizes published data from numerous labs. No new data were generated or analyzed in this article, so data sharing is not applicable.
Acknowledgments
We thank the members of the Prasanth laboratory and K. Prasanth for their insightful suggestions and R. Hsu for the artwork.
Conflicts of Interest
The authors declare no conflicts of interest. The authors declare no competing financial interests.
Abbreviations
IDR: intrinsically disordered region; MCM: minichromosome maintenance; ORC: origin recognition complex; PTM: post-translational modification; pre-RC: pre-Replication Complex; ORCA/LRWD1: ORC-Associated/Leucine-rich and WD-repeat-containing- 1.
References
- Hu, Y.; Stillman, B. Origins of DNA replication in eukaryotes. Mol. Cell 2023, 83, 352–372. [Google Scholar] [CrossRef]
- Attali, I.; Botchan, M.R.; Berger, J.M. Structural Mechanisms for Replicating DNA in Eukaryotes. Annu. Rev. Biochem. 2021, 90, 77–106. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Diffley, J.F.X. The Initiation of Eukaryotic DNA Replication. Annu. Rev. Biochem. 2022, 91, 107–131. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.S.; Gross, M.H.; Sousa, J.; Henrikus, S.S.; Greiwe, J.F.; Nans, A.; Diffley, J.F.X.; Costa, A. Mechanism of replication origin melting nucleated by CMG helicase assembly. Nature 2022, 606, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Stillman, B. The remarkable gymnastics of ORC. eLife 2022, 11, e76475. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Schneider, S.; Dodd, T.; Riera, A.; Bai, L.; Yan, C.; Magdalou, I.; Ivanov, I.; Stillman, B.; Li, H.; et al. Structural mechanism of helicase loading onto replication origin DNA by ORC-Cdc6. Proc. Natl. Acad. Sci. USA 2020, 117, 17747–17756. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Georgescu, R.; Bai, L.; Zhang, D.; Li, H.; O’Donnell, M.E. DNA unwinding mechanism of a eukaryotic replicative CMG helicase. Nat. Commun. 2020, 11, 688. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, P.J.; Blow, J.J. DDK: The Outsourced Kinase of Chromosome Maintenance. Biology 2022, 11, 877. [Google Scholar] [CrossRef] [PubMed]
- Francis, L.I.; Randell, J.C.; Takara, T.J.; Uchima, L.; Bell, S.P. Incorporation into the prereplicative complex activates the Mcm2-7 helicase for Cdc7-Dbf4 phosphorylation. Genes Dev. 2009, 23, 643–654. [Google Scholar] [CrossRef]
- Heller, R.C.; Kang, S.; Lam, W.M.; Chen, S.; Chan, C.S.; Bell, S.P. Eukaryotic origin-dependent DNA replication in vitro reveals sequential action of DDK and S-CDK kinases. Cell 2011, 146, 80–91. [Google Scholar] [CrossRef]
- Sheu, Y.J.; Stillman, B. Cdc7-Dbf4 phosphorylates MCM proteins via a docking site-mediated mechanism to promote S phase progression. Mol. Cell 2006, 24, 101–113. [Google Scholar] [CrossRef]
- Sheu, Y.J.; Stillman, B. The Dbf4-Cdc7 kinase promotes S phase by alleviating an inhibitory activity in Mcm4. Nature 2010, 463, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Umemori, T.; Hirai, K.; Muramatsu, S.; Kamimura, Y.; Araki, H. CDK-dependent phosphorylation of Sld2 and Sld3 initiates DNA replication in budding yeast. Nature 2007, 445, 328–332. [Google Scholar] [CrossRef]
- Zegerman, P.; Diffley, J.F. Phosphorylation of Sld2 and Sld3 by cyclin-dependent kinases promotes DNA replication in budding yeast. Nature 2007, 445, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, S.; Hirai, K.; Tak, Y.S.; Kamimura, Y.; Araki, H. CDK-dependent complex formation between replication proteins Dpb11, Sld2, Pol(epsilon), and GINS in budding yeast. Genes Dev. 2010, 24, 602–612. [Google Scholar] [CrossRef]
- Bell, S.P.; Stillman, B. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature 1992, 357, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.P.; Dutta, A. DNA replication in eukaryotic cells. Annu. Rev. Biochem. 2002, 71, 333–374. [Google Scholar] [CrossRef]
- Bell, S.P.; Kobayashi, R.; Stillman, B. Yeast origin recognition complex functions in transcription silencing and DNA replication. Science 1993, 262, 1844–1849. [Google Scholar] [CrossRef]
- Dhar, S.K.; Delmolino, L.; Dutta, A. Architecture of the human origin recognition complex. J. Biol. Chem. 2001, 276, 29067–29071. [Google Scholar] [CrossRef]
- Siddiqui, K.; Stillman, B. ATP-dependent assembly of the human origin recognition complex. J. Biol. Chem. 2007, 282, 32370–32383. [Google Scholar] [CrossRef]
- Vashee, S.; Simancek, P.; Challberg, M.D.; Kelly, T.J. Assembly of the human origin recognition complex. J. Biol. Chem. 2001, 276, 26666–26673. [Google Scholar] [CrossRef]
- Shen, Z.; Chakraborty, A.; Jain, A.; Giri, S.; Ha, T.; Prasanth, K.V.; Prasanth, S.G. Dynamic association of ORCA with prereplicative complex components regulates DNA replication initiation. Mol. Cell. Biol. 2012, 32, 3107–3120. [Google Scholar] [CrossRef]
- Shen, Z.; Prasanth, S.G. Orc2 protects ORCA from ubiquitin-mediated degradation. Cell Cycle 2012, 11, 3578–3589. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Sathyan, K.M.; Geng, Y.; Zheng, R.; Chakraborty, A.; Freeman, B.; Wang, F.; Prasanth, K.V.; Prasanth, S.G. A WD-repeat protein stabilizes ORC binding to chromatin. Mol. Cell 2010, 40, 99–111. [Google Scholar] [CrossRef]
- Hsu, R.Y.C.; Lin, Y.C.; Redon, C.; Sun, Q.; Singh, D.K.; Wang, Y.; Aggarwal, V.; Mitra, J.; Matur, A.; Moriarity, B.; et al. ORCA/LRWD1 Regulates Homologous Recombination at ALT-Telomeres by Modulating Heterochromatin Organization. iScience 2020, 23, 101038. [Google Scholar] [CrossRef]
- Prasanth, S.G.; Mendez, J.; Prasanth, K.V.; Stillman, B. Dynamics of pre-replication complex proteins during the cell division cycle. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 7–16. [Google Scholar] [CrossRef]
- Prasanth, S.G.; Prasanth, K.V.; Siddiqui, K.; Spector, D.L.; Stillman, B. Human Orc2 localizes to centrosomes, centromeres and heterochromatin during chromosome inheritance. EMBO J. 2004, 23, 2651–2663. [Google Scholar] [CrossRef] [PubMed]
- Mendez, J.; Zou-Yang, X.H.; Kim, S.Y.; Hidaka, M.; Tansey, W.P.; Stillman, B. Human origin recognition complex large subunit is degraded by ubiquitin-mediated proteolysis after initiation of DNA replication. Mol. Cell 2002, 9, 481–491. [Google Scholar] [CrossRef]
- Kara, N.; Hossain, M.; Prasanth, S.G.; Stillman, B. Orc1 Binding to Mitotic Chromosomes Precedes Spatial Patterning during G1 Phase and Assembly of the Origin Recognition Complex in Human Cells. J. Biol. Chem. 2015, 290, 12355–12369. [Google Scholar] [CrossRef] [PubMed]
- Speck, C.; Chen, Z.; Li, H.; Stillman, B. ATPase-dependent cooperative binding of ORC and Cdc6 to origin DNA. Nat. Struct. Mol. Biol. 2005, 12, 965–971. [Google Scholar] [CrossRef]
- Marks, A.B.; Fu, H.; Aladjem, M.I. Regulation of Replication Origins. Adv. Exp. Med. Biol. 2017, 1042, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Ganier, O.; Prorok, P.; Akerman, I.; Mechali, M. Metazoan DNA replication origins. Curr. Opin. Cell Biol. 2019, 58, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.C.; Mechali, M. DNA replication origins. Cold Spring Harb. Perspect. Biol. 2013, 5, a010116. [Google Scholar] [CrossRef]
- Prorok, P.; Artufel, M.; Aze, A.; Coulombe, P.; Peiffer, I.; Lacroix, L.; Guedin, A.; Mergny, J.L.; Damaschke, J.; Schepers, A.; et al. Involvement of G-quadruplex regions in mammalian replication origin activity. Nat. Commun. 2019, 10, 3274. [Google Scholar] [CrossRef] [PubMed]
- Mechali, M. Eukaryotic DNA replication origins: Many choices for appropriate answers. Nat. Rev. Mol. Cell Biol. 2010, 11, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Mechali, M.; Yoshida, K.; Coulombe, P.; Pasero, P. Genetic and epigenetic determinants of DNA replication origins, position and activation. Curr. Opin. Genet. Dev. 2013, 23, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Cayrou, C.; Ballester, B.; Peiffer, I.; Fenouil, R.; Coulombe, P.; Andrau, J.C.; van Helden, J.; Mechali, M. The chromatin environment shapes DNA replication origin organization and defines origin classes. Genome Res. 2015, 25, 1873–1885. [Google Scholar] [CrossRef]
- Cayrou, C.; Coulombe, P.; Puy, A.; Rialle, S.; Kaplan, N.; Segal, E.; Mechali, M. New insights into replication origin characteristics in metazoans. Cell Cycle 2012, 11, 658–667. [Google Scholar] [CrossRef] [PubMed]
- Cayrou, C.; Coulombe, P.; Vigneron, A.; Stanojcic, S.; Ganier, O.; Peiffer, I.; Rivals, E.; Puy, A.; Laurent-Chabalier, S.; Desprat, R.; et al. Genome-scale analysis of metazoan replication origins reveals their organization in specific but flexible sites defined by conserved features. Genome Res. 2011, 21, 1438–1449. [Google Scholar] [CrossRef]
- Cayrou, C.; Gregoire, D.; Coulombe, P.; Danis, E.; Mechali, M. Genome-scale identification of active DNA replication origins. Methods 2012, 57, 158–164. [Google Scholar] [CrossRef]
- Smith, O.K.; Aladjem, M.I. Chromatin structure and replication origins: Determinants of chromosome replication and nuclear organization. J. Mol. Biol. 2014, 426, 3330–3341. [Google Scholar] [CrossRef] [PubMed]
- Smith, O.K.; Kim, R.; Fu, H.; Martin, M.M.; Lin, C.M.; Utani, K.; Zhang, Y.; Marks, A.B.; Lalande, M.; Chamberlain, S.; et al. Distinct epigenetic features of differentiation-regulated replication origins. Epigenet. Chromatin 2016, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Brossas, C.; Duriez, B.; Valton, A.L.; Prioleau, M.N. Promoters are key organizers of the duplication of vertebrate genomes. Bioessays 2021, 43, e2100141. [Google Scholar] [CrossRef] [PubMed]
- Besnard, E.; Babled, A.; Lapasset, L.; Milhavet, O.; Parrinello, H.; Dantec, C.; Marin, J.M.; Lemaitre, J.M. Unraveling cell type-specific and reprogrammable human replication origin signatures associated with G-quadruplex consensus motifs. Nat. Struct. Mol. Biol. 2012, 19, 837–844. [Google Scholar] [CrossRef]
- Foulk, M.S.; Urban, J.M.; Casella, C.; Gerbi, S.A. Characterizing and controlling intrinsic biases of lambda exonuclease in nascent strand sequencing reveals phasing between nucleosomes and G-quadruplex motifs around a subset of human replication origins. Genome Res. 2015, 25, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.M.; Ryan, M.; Kim, R.; Zakas, A.L.; Fu, H.; Lin, C.M.; Reinhold, W.C.; Davis, S.R.; Bilke, S.; Liu, H.; et al. Genome-wide depletion of replication initiation events in highly transcribed regions. Genome Res. 2011, 21, 1822–1832. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Lajugie, J.; Fourel, N.; Selzer, A.; Schizas, M.; Bartholdy, B.; Mar, J.; Lin, C.M.; Martin, M.M.; Ryan, M.; et al. Allele-specific genome-wide profiling in human primary erythroblasts reveal replication program organization. PLoS Genet. 2014, 10, e1004319. [Google Scholar] [CrossRef]
- Rao, S.S.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef]
- Tian, M.; Wang, Z.; Su, Z.; Shibata, E.; Shibata, Y.; Dutta, A.; Zang, C. Integrative analysis of DNA replication origins and ORC/MCM binding sites in human cells reveals a lack of overlap. bioRxiv 2023. [Google Scholar] [CrossRef]
- Hoggard, T.; Shor, E.; Muller, C.A.; Nieduszynski, C.A.; Fox, C.A. A Link between ORC-origin binding mechanisms and origin activation time revealed in budding yeast. PLoS Genet. 2013, 9, e1003798. [Google Scholar] [CrossRef]
- Kuzmichev, A.; Nishioka, K.; Erdjument-Bromage, H.; Tempst, P.; Reinberg, D. Histone methyltransferase activity associated with a human multiprotein complex containing the Enhancer of Zeste protein. Genes Dev. 2002, 16, 2893–2905. [Google Scholar] [CrossRef]
- Pasini, D.; Bracken, A.P.; Helin, K. Polycomb group proteins in cell cycle progression and cancer. Cell Cycle 2004, 3, 396–400. [Google Scholar] [CrossRef]
- Piunti, A.; Rossi, A.; Cerutti, A.; Albert, M.; Jammula, S.; Scelfo, A.; Cedrone, L.; Fragola, G.; Olsson, L.; Koseki, H.; et al. Polycomb proteins control proliferation and transformation independently of cell cycle checkpoints by regulating DNA replication. Nat. Commun. 2014, 5, 3649. [Google Scholar] [CrossRef]
- Gorisch, S.M.; Wachsmuth, M.; Toth, K.F.; Lichter, P.; Rippe, K. Histone acetylation increases chromatin accessibility. J. Cell Sci. 2005, 118, 5825–5834. [Google Scholar] [CrossRef] [PubMed]
- Aladjem, M.I. Replication in context: Dynamic regulation of DNA replication patterns in metazoans. Nat. Rev. Genet. 2007, 8, 588–600. [Google Scholar] [CrossRef] [PubMed]
- Hassan-Zadeh, V.; Chilaka, S.; Cadoret, J.C.; Ma, M.K.; Boggetto, N.; West, A.G.; Prioleau, M.N. USF binding sequences from the HS4 insulator element impose early replication timing on a vertebrate replicator. PLoS Biol. 2012, 10, e1001277. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Fu, H.; Lin, C.M.; Conner, A.L.; Zhang, Y.; Aladjem, M.I. Prevention of transcriptional silencing by a replicator-binding complex consisting of SWI/SNF, MeCP1, and hnRNP C1/C2. Mol. Cell. Biol. 2011, 31, 3472–3484. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.; Prasanth, S.G. Replicating and transcribing on twisted roads of chromatin. Brief. Funct. Genom. 2012, 11, 188–204. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.; Chakraborty, A.; Sathyan, K.M.; Prasanth, K.V.; Prasanth, S.G. Orc5 induces large-scale chromatin decondensation in a GCN5-dependent manner. J. Cell Sci. 2016, 129, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Miotto, B.; Struhl, K. HBO1 histone acetylase activity is essential for DNA replication licensing and inhibited by Geminin. Mol. Cell 2010, 37, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Miotto, B.; Struhl, K. JNK1 phosphorylation of Cdt1 inhibits recruitment of HBO1 histone acetylase and blocks replication licensing in response to stress. Mol. Cell 2011, 44, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, M.; Stillman, B. Histone acetyltransferase HBO1 interacts with the ORC1 subunit of the human initiator protein. J. Biol. Chem. 1999, 274, 23027–23034. [Google Scholar] [CrossRef]
- Bartke, T.; Vermeulen, M.; Xhemalce, B.; Robson, S.C.; Mann, M.; Kouzarides, T. Nucleosome-interacting proteins regulated by DNA and histone methylation. Cell 2010, 143, 470–484. [Google Scholar] [CrossRef]
- Giri, S.; Aggarwal, V.; Pontis, J.; Shen, Z.; Chakraborty, A.; Khan, A.; Mizzen, C.; Prasanth, K.V.; Ait-Si-Ali, S.; Ha, T.; et al. The preRC protein ORCA organizes heterochromatin by assembling histone H3 lysine 9 methyltransferases on chromatin. eLife 2015, 4, e06496. [Google Scholar] [CrossRef]
- Giri, S.; Prasanth, S.G. Association of ORCA/LRWD1 with repressive histone methyl transferases mediates heterochromatin organization. Nucleus 2015, 6, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, M.; Eberl, H.C.; Matarese, F.; Marks, H.; Denissov, S.; Butter, F.; Lee, K.K.; Olsen, J.V.; Hyman, A.A.; Stunnenberg, H.G.; et al. Quantitative interaction proteomics and genome-wide profiling of epigenetic histone marks and their readers. Cell 2010, 142, 967–980. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Khan, A.; Marks, A.B.; Smith, O.K.; Giri, S.; Lin, Y.C.; Creager, R.; MacAlpine, D.M.; Prasanth, K.V.; Aladjem, M.I.; et al. Temporal association of ORCA/LRWD1 to late-firing origins during G1 dictates heterochromatin replication and organization. Nucleic Acids Res. 2017, 45, 2490–2502. [Google Scholar] [CrossRef]
- Sahu, S.; Ekundayo, B.E.; Kumar, A.; Bleichert, F. A dual role for the chromatin reader ORCA/LRWD1 in targeting the origin recognition complex to chromatin. EMBO J. 2023, 42, e114654. [Google Scholar] [CrossRef]
- Chakraborty, A.; Shen, Z.; Prasanth, S.G. “ORCanization” on heterochromatin: Linking DNA replication initiation to chromatin organization. Epigenetics 2011, 6, 665–670. [Google Scholar] [CrossRef]
- Schwaiger, M.; Kohler, H.; Oakeley, E.J.; Stadler, M.B.; Schubeler, D. Heterochromatin protein 1 (HP1) modulates replication timing of the Drosophila genome. Genome Res. 2010, 20, 771–780. [Google Scholar] [CrossRef]
- Pak, D.T.; Pflumm, M.; Chesnokov, I.; Huang, D.W.; Kellum, R.; Marr, J.; Romanowski, P.; Botchan, M.R. Association of the origin recognition complex with heterochromatin and HP1 in higher eukaryotes. Cell 1997, 91, 311–323. [Google Scholar] [CrossRef]
- Prasanth, S.G.; Shen, Z.; Prasanth, K.V.; Stillman, B. Human origin recognition complex is essential for HP1 binding to chromatin and heterochromatin organization. Proc. Natl. Acad. Sci. USA 2010, 107, 15093–15098. [Google Scholar] [CrossRef]
- Kuo, A.J.; Song, J.; Cheung, P.; Ishibe-Murakami, S.; Yamazoe, S.; Chen, J.K.; Patel, D.J.; Gozani, O. The BAH domain of ORC1 links H4K20me2 to DNA replication licensing and Meier-Gorlin syndrome. Nature 2012, 484, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Sherstyuk, V.V.; Shevchenko, A.I.; Zakian, S.M. Epigenetic landscape for initiation of DNA replication. Chromosoma 2014, 123, 183–199. [Google Scholar] [CrossRef]
- Abbas, T.; Shibata, E.; Park, J.; Jha, S.; Karnani, N.; Dutta, A. CRL4(Cdt2) regulates cell proliferation and histone gene expression by targeting PR-Set7/Set8 for degradation. Mol. Cell 2010, 40, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Beck, D.B.; Burton, A.; Oda, H.; Ziegler-Birling, C.; Torres-Padilla, M.E.; Reinberg, D. The role of PR-Set7 in replication licensing depends on Suv4-20h. Genes Dev. 2012, 26, 2580–2589. [Google Scholar] [CrossRef]
- Beck, D.B.; Oda, H.; Shen, S.S.; Reinberg, D. PR-Set7 and H4K20me1: At the crossroads of genome integrity, cell cycle, chromosome condensation, and transcription. Genes Dev. 2012, 26, 325–337. [Google Scholar] [CrossRef]
- Tardat, M.; Brustel, J.; Kirsh, O.; Lefevbre, C.; Callanan, M.; Sardet, C.; Julien, E. The histone H4 Lys 20 methyltransferase PR-Set7 regulates replication origins in mammalian cells. Nat. Cell Biol. 2010, 12, 1086–1093. [Google Scholar] [CrossRef]
- Long, H.; Zhang, L.; Lv, M.; Wen, Z.; Zhang, W.; Chen, X.; Zhang, P.; Li, T.; Chang, L.; Jin, C.; et al. H2A.Z facilitates licensing and activation of early replication origins. Nature 2020, 577, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Poulet-Benedetti, J.; Tonnerre-Doncarli, C.; Valton, A.L.; Laurent, M.; Gerard, M.; Barinova, N.; Parisis, N.; Massip, F.; Picard, F.; Prioleau, M.N. Dimeric G-quadruplex motifs-induced NFRs determine strong replication origins in vertebrates. Nat. Commun. 2023, 14, 4843. [Google Scholar] [CrossRef]
- Li, S.; Wasserman, M.R.; Yurieva, O.; Bai, L.; O’Donnell, M.E.; Liu, S. Origin recognition complex harbors an intrinsic nucleosome remodeling activity. Proc. Natl. Acad. Sci. USA 2022, 119, e2211568119. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wasserman, M.R.; Yurieva, O.; Bai, L.; O’Donnell, M.E.; Liu, S. Nucleosome-directed replication origin licensing independent of a consensus DNA sequence. Nat. Commun. 2022, 13, 4947. [Google Scholar] [CrossRef] [PubMed]
- Chacin, E.; Reusswig, K.U.; Furtmeier, J.; Bansal, P.; Karl, L.A.; Pfander, B.; Straub, T.; Korber, P.; Kurat, C.F. Establishment and function of chromatin organization at replication origins. Nature 2023, 616, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Azmi, I.F.; Watanabe, S.; Maloney, M.F.; Kang, S.; Belsky, J.A.; MacAlpine, D.M.; Peterson, C.L.; Bell, S.P. Nucleosomes influence multiple steps during replication initiation. eLife 2017, 6, e22512. [Google Scholar] [CrossRef] [PubMed]
- Berbenetz, N.M.; Nislow, C.; Brown, G.W. Diversity of eukaryotic DNA replication origins revealed by genome-wide analysis of chromatin structure. PLoS Genet. 2010, 6, e1001092. [Google Scholar] [CrossRef]
- Liu, J.; Zimmer, K.; Rusch, D.B.; Paranjape, N.; Podicheti, R.; Tang, H.; Calvi, B.R. DNA sequence templates adjacent nucleosome and ORC sites at gene amplification origins in Drosophila. Nucleic Acids Res. 2015, 43, 8746–8761. [Google Scholar] [CrossRef] [PubMed]
- Lubelsky, Y.; Sasaki, T.; Kuipers, M.A.; Lucas, I.; Le Beau, M.M.; Carignon, S.; Debatisse, M.; Prinz, J.A.; Dennis, J.H.; Gilbert, D.M. Pre-replication complex proteins assemble at regions of low nucleosome occupancy within the Chinese hamster dihydrofolate reductase initiation zone. Nucleic Acids Res. 2011, 39, 3141–3155. [Google Scholar] [CrossRef]
- Yin, S.; Deng, W.; Hu, L.; Kong, X. The impact of nucleosome positioning on the organization of replication origins in eukaryotes. Biochem. Biophys. Res. Commun. 2009, 385, 363–368. [Google Scholar] [CrossRef]
- Hossain, M.; Bhalla, K.; Stillman, B. Multiple, short protein binding motifs in ORC1 and CDC6 control the initiation of DNA replication. Mol. Cell 2021, 81, 1951–1969.e6. [Google Scholar] [CrossRef]
- Parker, M.W.; Bell, M.; Mir, M.; Kao, J.A.; Darzacq, X.; Botchan, M.R.; Berger, J.M. A new class of disordered elements controls DNA replication through initiator self-assembly. eLife 2019, 8, e48562. [Google Scholar] [CrossRef]
- Unnikrishnan, A.; Gafken, P.R.; Tsukiyama, T. Dynamic changes in histone acetylation regulate origins of DNA replication. Nat. Struct. Mol. Biol. 2010, 17, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Hoggard, T.; Muller, C.A.; Nieduszynski, C.A.; Weinreich, M.; Fox, C.A. Sir2 mitigates an intrinsic imbalance in origin licensing efficiency between early- and late-replicating euchromatin. Proc. Natl. Acad. Sci. USA 2020, 117, 14314–14321. [Google Scholar] [CrossRef] [PubMed]
- Hoggard, T.A.; Chang, F.; Perry, K.R.; Subramanian, S.; Kenworthy, J.; Chueng, J.; Shor, E.; Hyland, E.M.; Boeke, J.D.; Weinreich, M.; et al. Yeast heterochromatin regulators Sir2 and Sir3 act directly at euchromatic DNA replication origins. PLoS Genet. 2018, 14, e1007418. [Google Scholar] [CrossRef]
- Pasero, P.; Bensimon, A.; Schwob, E. Single-molecule analysis reveals clustering and epigenetic regulation of replication origins at the yeast rDNA locus. Genes Dev. 2002, 16, 2479–2484. [Google Scholar] [CrossRef]
- Aparicio, O.M.; Billington, B.L.; Gottschling, D.E. Modifiers of position effect are shared between telomeric and silent mating-type loci in S. cerevisiae. Cell 1991, 66, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Rine, J.; Herskowitz, I. Four genes responsible for a position effect on expression from HML and HMR in Saccharomyces cerevisiae. Genetics 1987, 116, 9–22. [Google Scholar] [CrossRef]
- Stevenson, J.B.; Gottschling, D.E. Telomeric chromatin modulates replication timing near chromosome ends. Genes Dev. 1999, 13, 146–151. [Google Scholar] [CrossRef]
- Vogelauer, M.; Rubbi, L.; Lucas, I.; Brewer, B.J.; Grunstein, M. Histone acetylation regulates the time of replication origin firing. Mol. Cell 2002, 10, 1223–1233. [Google Scholar] [CrossRef]
- Tremblay, R.; Mehrjoo, Y.; Ahmed, O.; Simoneau, A.; McQuaid, M.E.; Affar, E.B.; Nislow, C.; Giaever, G.; Wurtele, H. Persistent Acetylation of Histone H3 Lysine 56 Compromises the Activity of DNA Replication Origins. Mol. Cell. Biol. 2023, 43, 566–595. [Google Scholar] [CrossRef]
- Santos-Rosa, H.; Millan-Zambrano, G.; Han, N.; Leonardi, T.; Klimontova, M.; Nasiscionyte, S.; Pandolfini, L.; Tzelepis, K.; Bartke, T.; Kouzarides, T. Methylation of histone H3 at lysine 37 by Set1 and Set2 prevents spurious DNA replication. Mol. Cell 2021, 81, 2793–2807.e8. [Google Scholar] [CrossRef]
- Rizzardi, L.F.; Dorn, E.S.; Strahl, B.D.; Cook, J.G. DNA replication origin function is promoted by H3K4 di-methylation in Saccharomyces cerevisiae. Genetics 2012, 192, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Pryde, F.; Jain, D.; Kerr, A.; Curley, R.; Mariotti, F.R.; Vogelauer, M. H3 k36 methylation helps determine the timing of cdc45 association with replication origins. PLoS ONE 2009, 4, e5882. [Google Scholar] [CrossRef]
- Lee, C.S.K.; Weibeta, M.; Hamperl, S. Where and when to start: Regulating DNA replication origin activity in eukaryotic genomes. Nucleus 2023, 14, 2229642. [Google Scholar] [CrossRef] [PubMed]
- Agirre, E.; Oldfield, A.J.; Bellora, N.; Segelle, A.; Luco, R.F. Splicing-associated chromatin signatures: A combinatorial and position-dependent role for histone marks in splicing definition. Nat. Commun. 2021, 12, 682. [Google Scholar] [CrossRef] [PubMed]
- Rondinelli, B.; Schwerer, H.; Antonini, E.; Gaviraghi, M.; Lupi, A.; Frenquelli, M.; Cittaro, D.; Segalla, S.; Lemaitre, J.M.; Tonon, G. H3K4me3 demethylation by the histone demethylase KDM5C/JARID1C promotes DNA replication origin firing. Nucleic Acids Res. 2015, 43, 2560–2574. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Wang, Z.; Zhang, H.; Gan, H.; Zhang, Z. H3K9me3 demethylase Kdm4d facilitates the formation of pre-initiative complex and regulates DNA replication. Nucleic Acids Res. 2017, 45, 169–180. [Google Scholar] [CrossRef]
- Prorok, P.; Forouzanfar, F.; Murugarren, N.; Peiffer, I.; Charton, R.; Akerman, I.; Mechali, M. Loss of Ezh2 function remodels the DNA replication initiation landscape. Cell Rep. 2023, 42, 112280. [Google Scholar] [CrossRef]
- Fu, H.; Maunakea, A.K.; Martin, M.M.; Huang, L.; Zhang, Y.; Ryan, M.; Kim, R.; Lin, C.M.; Zhao, K.; Aladjem, M.I. Methylation of histone H3 on lysine 79 associates with a group of replication origins and helps limit DNA replication once per cell cycle. PLoS Genet. 2013, 9, e1003542. [Google Scholar] [CrossRef]
- Ding, Q.; Edwards, M.M.; Wang, N.; Zhu, X.; Bracci, A.N.; Hulke, M.L.; Hu, Y.; Tong, Y.; Hsiao, J.; Charvet, C.J.; et al. The genetic architecture of DNA replication timing in human pluripotent stem cells. Nat. Commun. 2021, 12, 6746. [Google Scholar] [CrossRef]
- Van Rechem, C.; Ji, F.; Chakraborty, D.; Black, J.C.; Sadreyev, R.I.; Whetstine, J.R. Collective regulation of chromatin modifications predicts replication timing during cell cycle. Cell Rep. 2021, 37, 109799. [Google Scholar] [CrossRef]
- Duncker, B.P.; Chesnokov, I.N.; McConkey, B.J. The origin recognition complex protein family. Genome Biol. 2009, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.M.; Zhang, Z. Leucine-rich repeat and WD repeat-containing protein 1 is recruited to pericentric heterochromatin by trimethylated lysine 9 of histone H3 and maintains heterochromatin silencing. J. Biol. Chem. 2012, 287, 15024–15033. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Cheng, J.; Bawa-Khalfe, T.; Yao, X.; Chin, Y.E.; Yeh, E.T.H. SUMOylated ORC2 Recruits a Histone Demethylase to Regulate Centromeric Histone Modification and Genomic Stability. Cell Rep. 2016, 15, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Bauwens, S.; Lototska, L.; Koundrioukoff, S.; Debatisse, M.; Ye, J.; Gilson, E.; Mendez-Bermudez, A. The Telomeric Protein TRF2 Regulates Replication Origin Activity within Pericentromeric Heterochromatin. Life 2021, 11, 267. [Google Scholar] [CrossRef]
- Nurk, S.; Koren, S.; Rhie, A.; Rautiainen, M.; Bzikadze, A.V.; Mikheenko, A.; Vollger, M.R.; Altemose, N.; Uralsky, L.; Gershman, A.; et al. The complete sequence of a human genome. Science 2022, 376, 44–53. [Google Scholar] [CrossRef]
- Deng, Z.; Dheekollu, J.; Broccoli, D.; Dutta, A.; Lieberman, P.M. The origin recognition complex localizes to telomere repeats and prevents telomere-circle formation. Curr. Biol. 2007, 17, 1989–1995. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA binding to TRF2 facilitates heterochromatin formation and ORC recruitment at telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef]
- Higa, M.; Matsuda, Y.; Fujii, J.; Sugimoto, N.; Yoshida, K.; Fujita, M. TRF2-mediated ORC recruitment underlies telomere stability upon DNA replication stress. Nucleic Acids Res. 2021, 49, 12234–12251. [Google Scholar] [CrossRef]
- Chan, K.M.; Zhang, H.; Malureanu, L.; van Deursen, J.; Zhang, Z. Diverse factors are involved in maintaining X chromosome inactivation. Proc. Natl. Acad. Sci. USA 2011, 108, 16699–16704. [Google Scholar] [CrossRef]
- Caudai, C.; Galizia, A.; Geraci, F.; Le Pera, L.; Morea, V.; Salerno, E.; Via, A.; Colombo, T. AI applications in functional genomics. Comput. Struct. Biotechnol. J. 2021, 19, 5762–5790. [Google Scholar] [CrossRef]
Figure 1.
(a) The temporal association of Orc1 to H4K20me2 and ORCA to H4K20me3 during G1 dictates early and late origins in humans. Live cell imaging, immunofluorescence, and genome-wide ChIP studies have demonstrated the spatiotemporal localization of Orc1 and ORCA during the cell cycle. Time-lapse imaging studies predict that human Orc1 associates with all origins in early G1 and then is sequentially released from most sites that represent heterochromatic sites, except at late origins. Orc1 associates with H4K20me2/3 marks. ORCA is associated with repressive histone marks, primarily H3K9me3 and H4K20me3, and is likely enriched only at late-firing origins in G1. (b) Model depicting organized patterns of Orc1 during the G1 phase and how it anticipates the spatiotemporal dynamics of MCMs and PCNA in sequential order. During early G1, Orc1 is present as punctate foci throughout the nucleus, and as cells progress through G1, it is enriched only at heterochromatic sites. The MCM complex shows similar patterns to Orc1, but it is homogenously distributed in G1, shows temporal patterning during the S phase, and is lost from most sites by the end of the S phase. Finally, PCNA shows punctate foci in the early S phase and is redistributed to heterochromatic sites during the mid and late S phase.
Figure 1.
(a) The temporal association of Orc1 to H4K20me2 and ORCA to H4K20me3 during G1 dictates early and late origins in humans. Live cell imaging, immunofluorescence, and genome-wide ChIP studies have demonstrated the spatiotemporal localization of Orc1 and ORCA during the cell cycle. Time-lapse imaging studies predict that human Orc1 associates with all origins in early G1 and then is sequentially released from most sites that represent heterochromatic sites, except at late origins. Orc1 associates with H4K20me2/3 marks. ORCA is associated with repressive histone marks, primarily H3K9me3 and H4K20me3, and is likely enriched only at late-firing origins in G1. (b) Model depicting organized patterns of Orc1 during the G1 phase and how it anticipates the spatiotemporal dynamics of MCMs and PCNA in sequential order. During early G1, Orc1 is present as punctate foci throughout the nucleus, and as cells progress through G1, it is enriched only at heterochromatic sites. The MCM complex shows similar patterns to Orc1, but it is homogenously distributed in G1, shows temporal patterning during the S phase, and is lost from most sites by the end of the S phase. Finally, PCNA shows punctate foci in the early S phase and is redistributed to heterochromatic sites during the mid and late S phase.
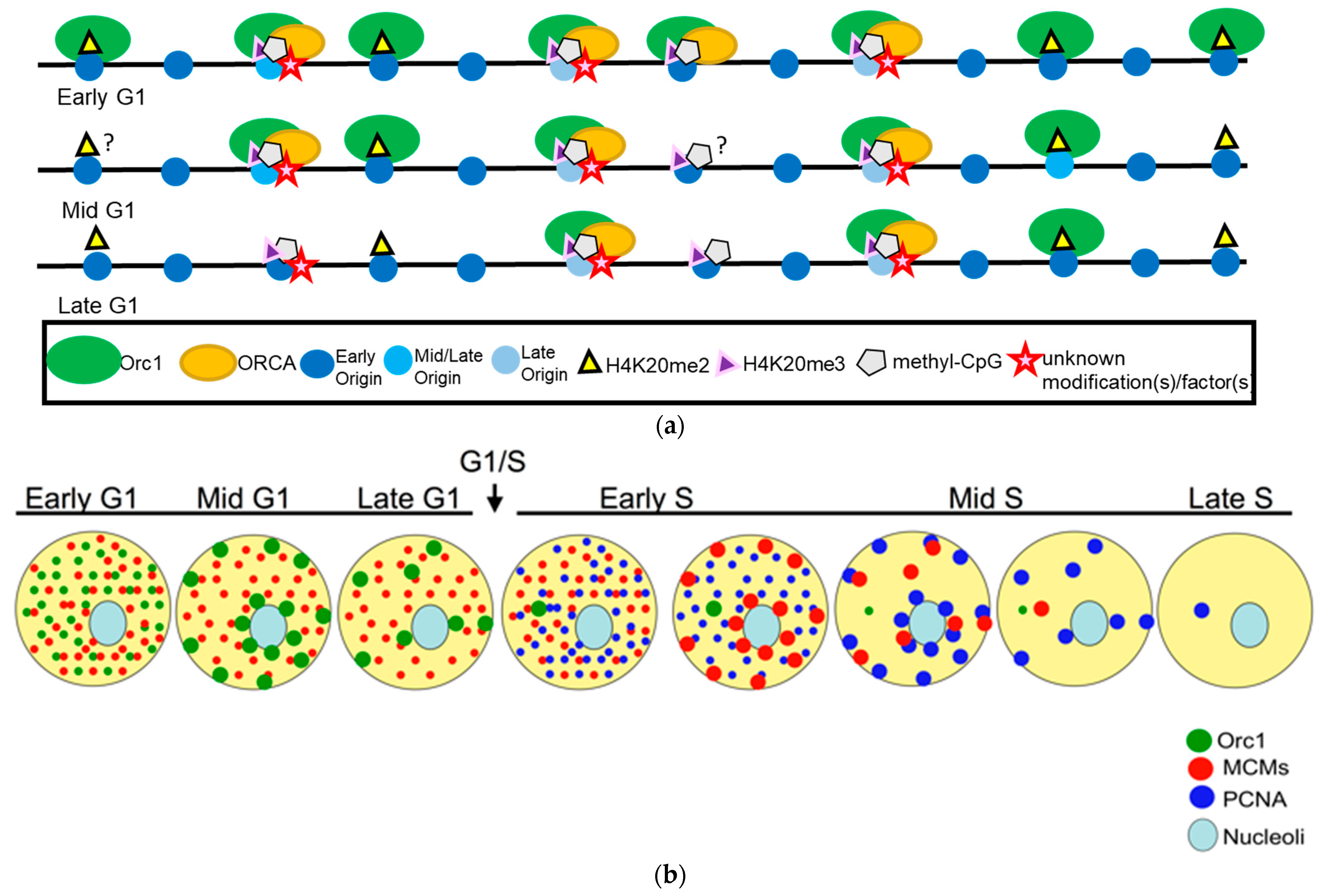
Figure 2.
(a) Replication initiators controlling diverse cellular functions, coordinating the completion of duplication with chromosome segregation and cell division. At origins: a model depicting the binding of ORCA to specific replication origins enriched for H3K9me3. At telomeres: the binding of ORCA to ALT-telomeres could be through binding to H3K9me3 and G-quadruplex DNA through associations with RPA. APB: ALT-associated PML body. At centromeres: the binding of ORCA to various chromatin marks that represent origins or for mitotic regulation. (b) At facultative heterochromatin: the binding of ORCA to an inactive X-chromosome. The figure models have been adapted and modified from Wang et al., 2017 and Giri et al., 2015 [64,67].
Figure 2.
(a) Replication initiators controlling diverse cellular functions, coordinating the completion of duplication with chromosome segregation and cell division. At origins: a model depicting the binding of ORCA to specific replication origins enriched for H3K9me3. At telomeres: the binding of ORCA to ALT-telomeres could be through binding to H3K9me3 and G-quadruplex DNA through associations with RPA. APB: ALT-associated PML body. At centromeres: the binding of ORCA to various chromatin marks that represent origins or for mitotic regulation. (b) At facultative heterochromatin: the binding of ORCA to an inactive X-chromosome. The figure models have been adapted and modified from Wang et al., 2017 and Giri et al., 2015 [64,67].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ahmad, H.; Chetlangia, N.; Prasanth, S.G. Chromatin’s Influence on Pre-Replication Complex Assembly and Function. Biology 2024, 13, 152. https://doi.org/10.3390/biology13030152
AMA Style
Ahmad H, Chetlangia N, Prasanth SG. Chromatin’s Influence on Pre-Replication Complex Assembly and Function. Biology. 2024; 13(3):152. https://doi.org/10.3390/biology13030152
Chicago/Turabian StyleAhmad, Hina, Neha Chetlangia, and Supriya G. Prasanth. 2024. "Chromatin’s Influence on Pre-Replication Complex Assembly and Function" Biology 13, no. 3: 152. https://doi.org/10.3390/biology13030152
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.